
High-Throughput Transcriptomics of Celf1 Conditional Knockout Lens Identifies Downstream Networks Linked to Cataract Pathology
 , , , and
, , , and 
Abstract
:1. Introduction
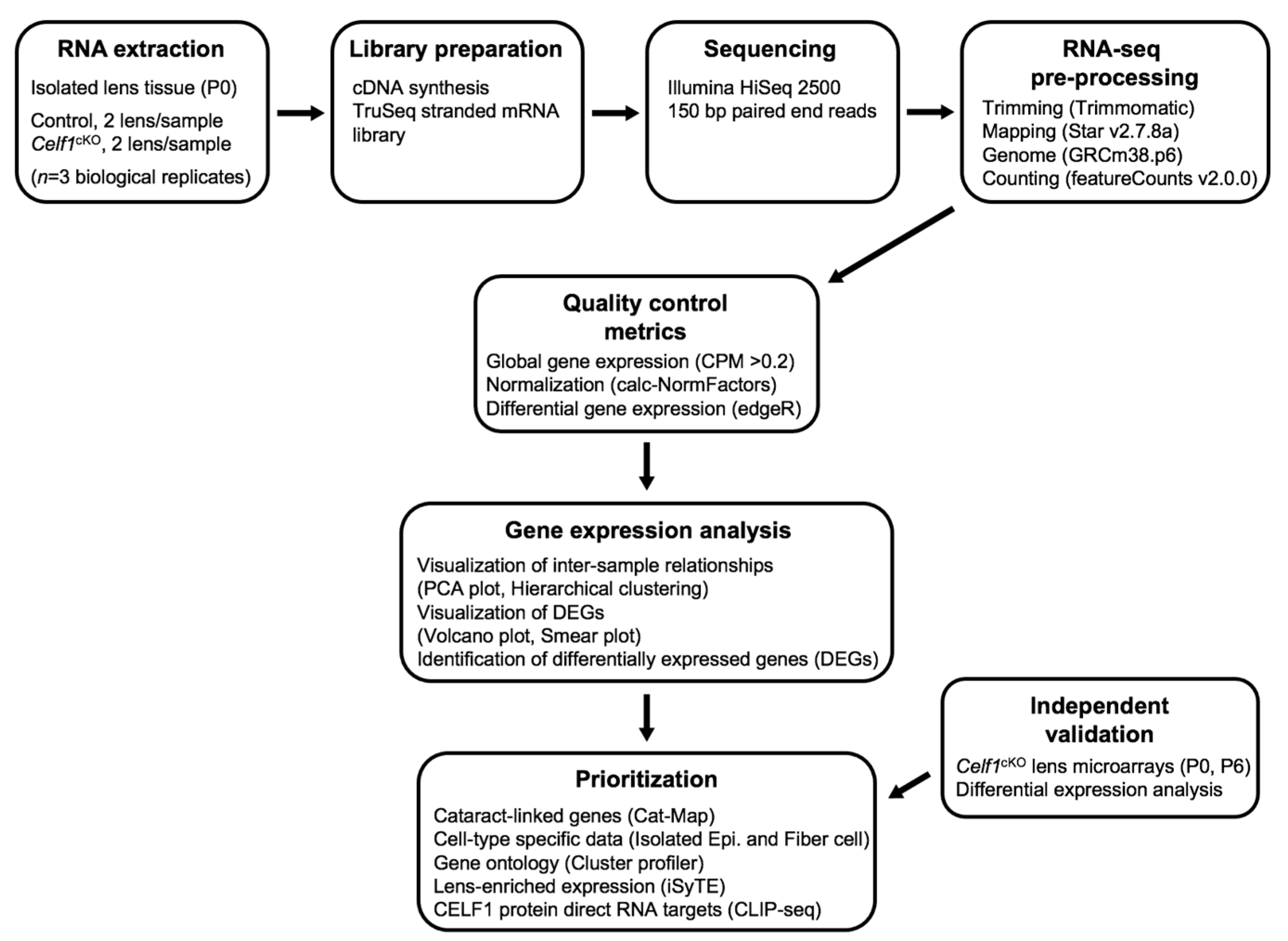
2. Materials and Methods
2.1. Animals
2.2. Lens RNA Isolation
2.3. RNA-Sequencing and Analysis
2.4. Microarray Analysis
2.5. Prioritization of DEGs by Cat-Map, iSyTE, Expression in Fiber vs. Epi, and Pathway Analysis
2.5.1. Cat-Map: Cataract Associated Genes
2.5.2. iSyTE: Gene Expression Enrichment in the Lens
2.5.3. Celf1cKO DEGs Preferentially Expressed in Normal Lens Fiber Cells or Epithelial Cells
2.5.4. Celf1cKO DEGs Independently Identified as RNA Targets of CELF1 Protein by CLIP-Seq in a Human Cell Line
2.5.5. Gene Ontology (GO) Term and Pathways Analysis
2.5.6. Immunostaining Analysis
3. Results
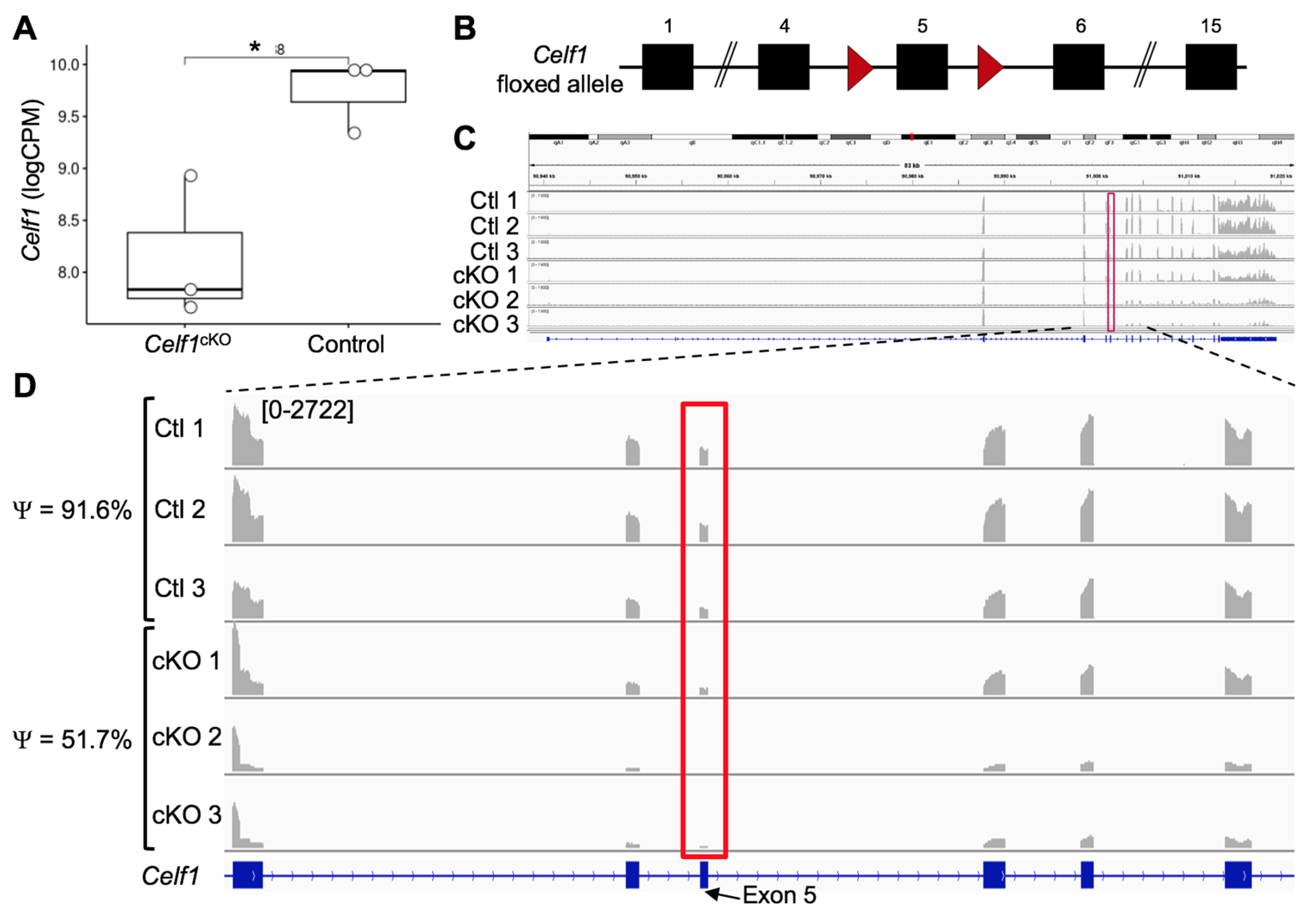
3.1. Generation of RNA-Seq Datasets from Celf1cKO and Control Lenses
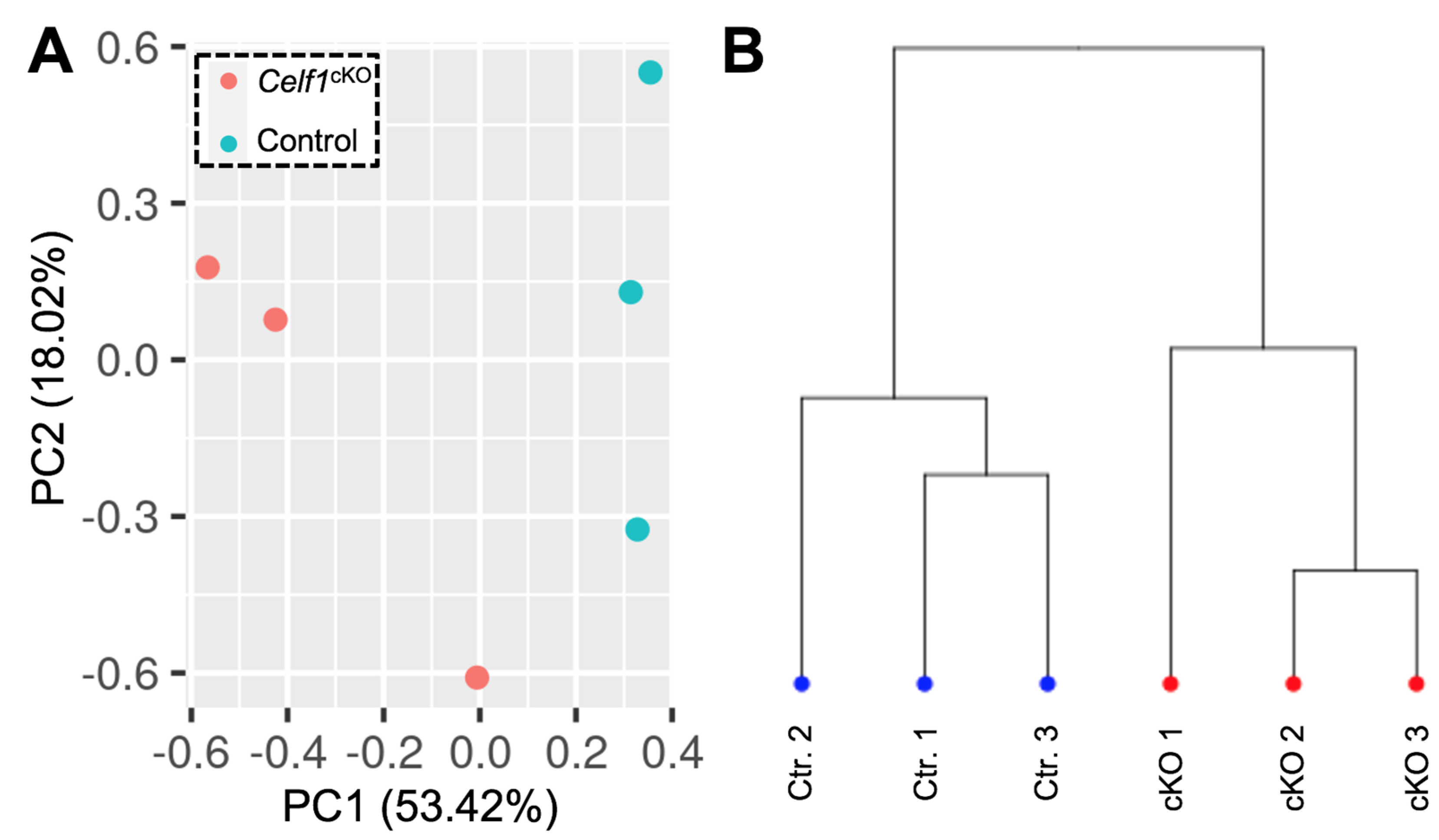
3.2. Quality Control of RNA-Seq Datasets
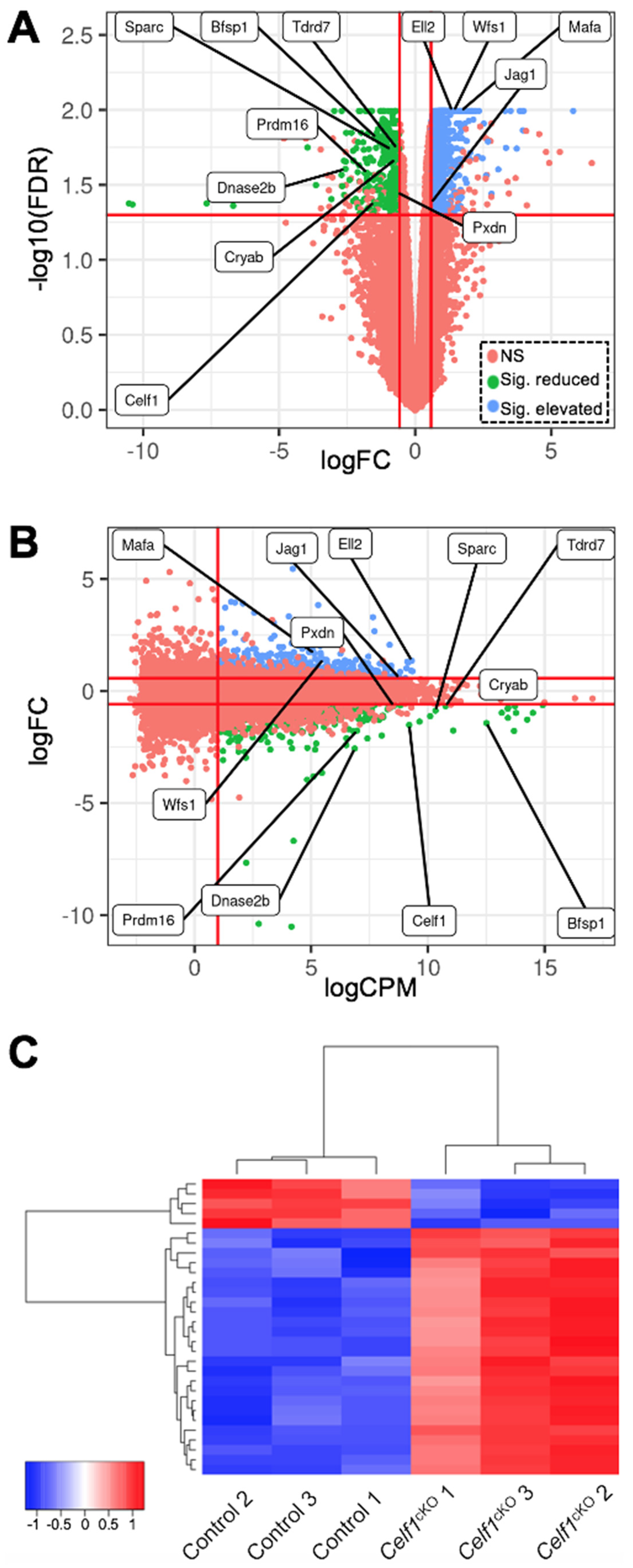
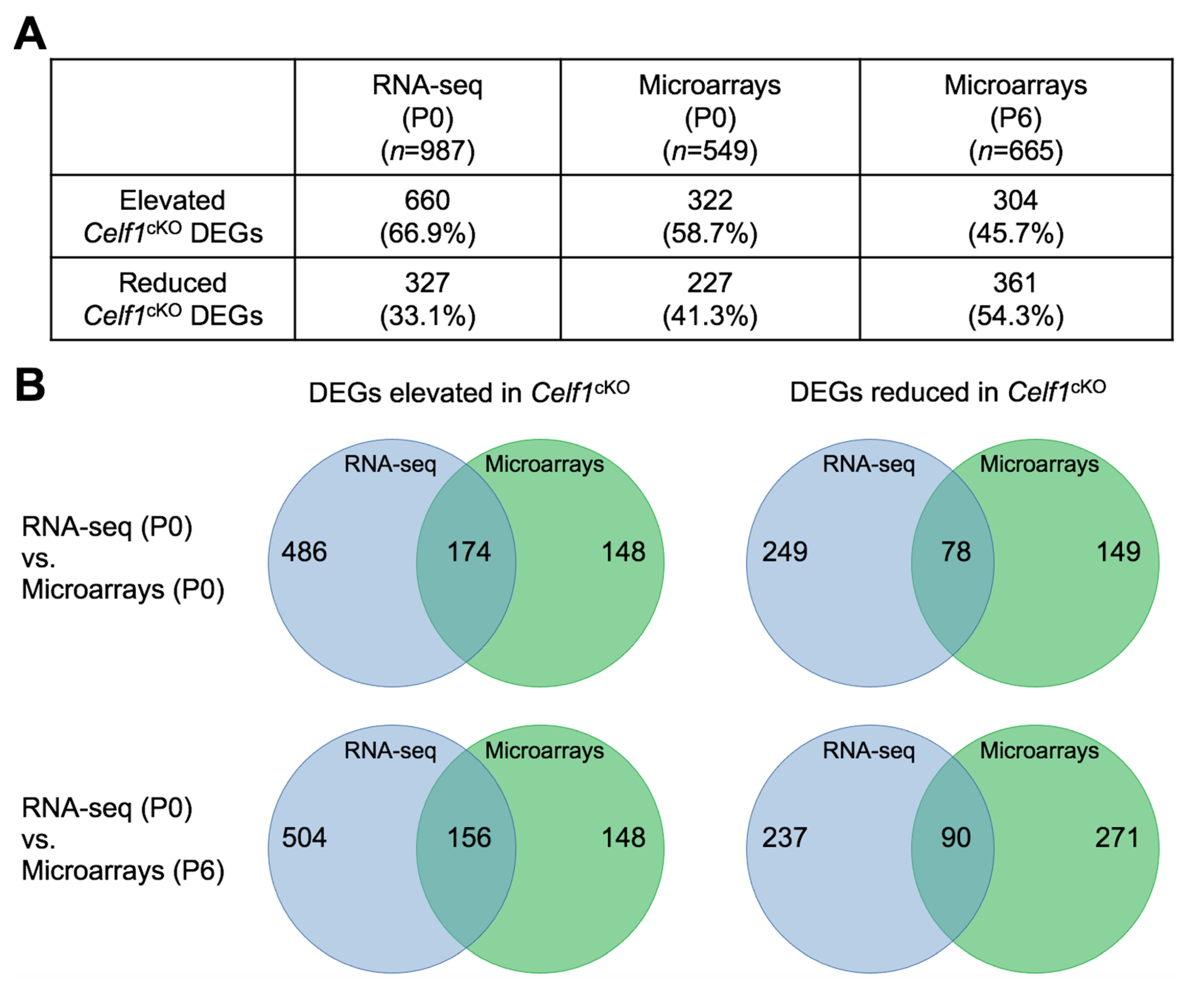
3.3. Identification of Differentially Expressed Genes (DEGs) in Celf1cKO Lens
3.4. Relevance of Celf1cKO Lens DEGs to Lens Development and Cataracts
3.4.1. Prioritization of Celf1cKO Lens DEGs Using the Cat-Map Database
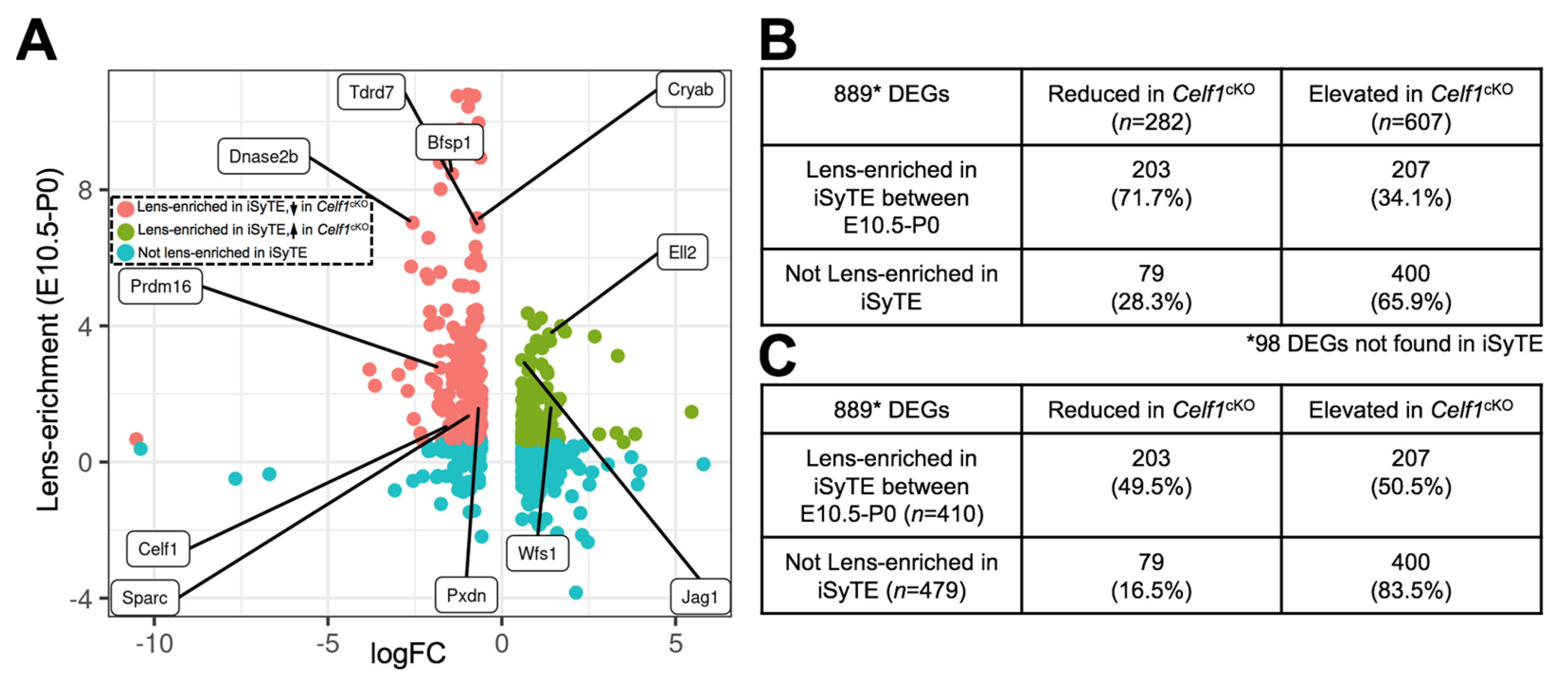
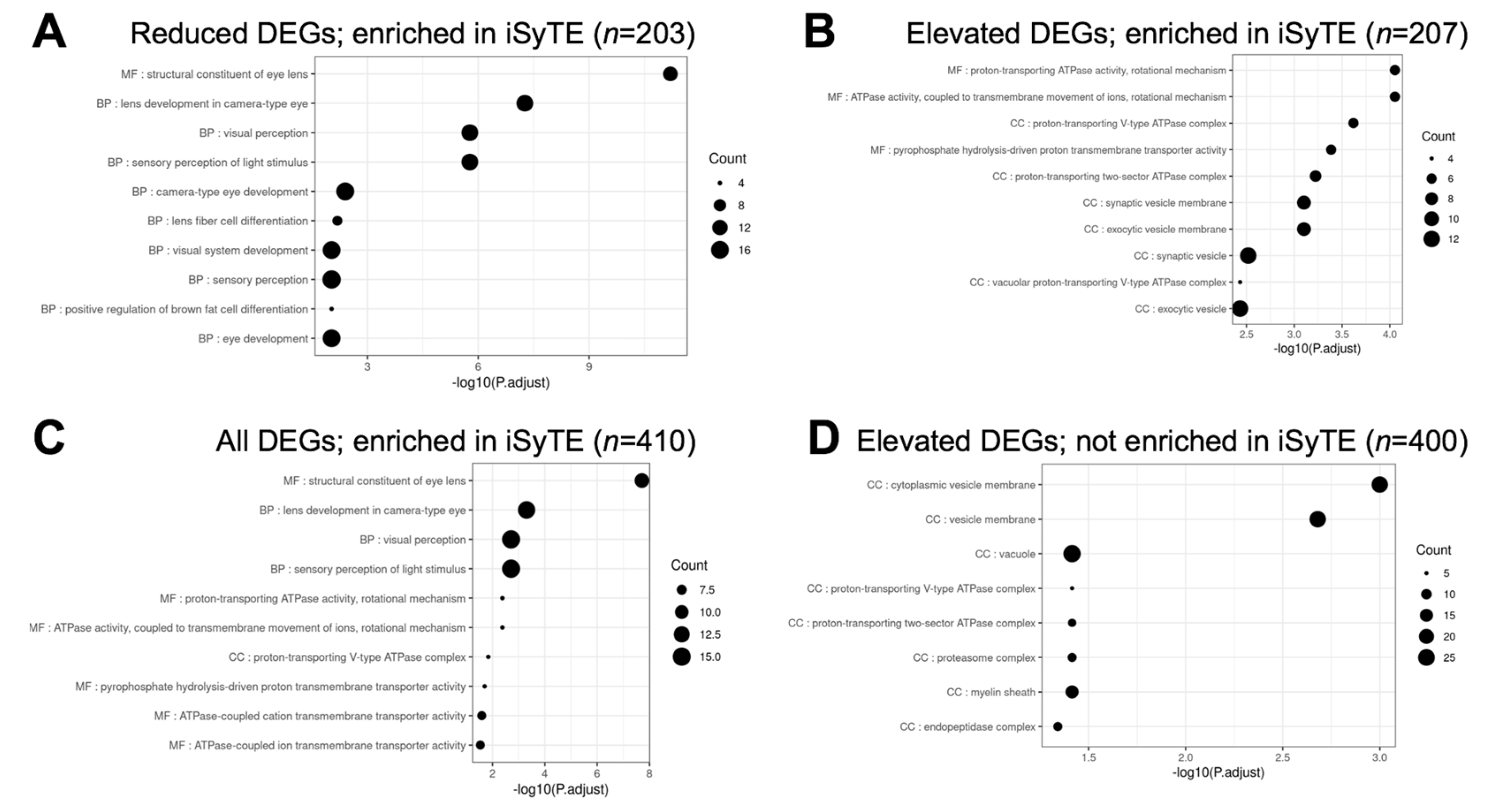
3.4.2. Prioritization of Celf1cKO Lens DEGs Using the iSyTE Database
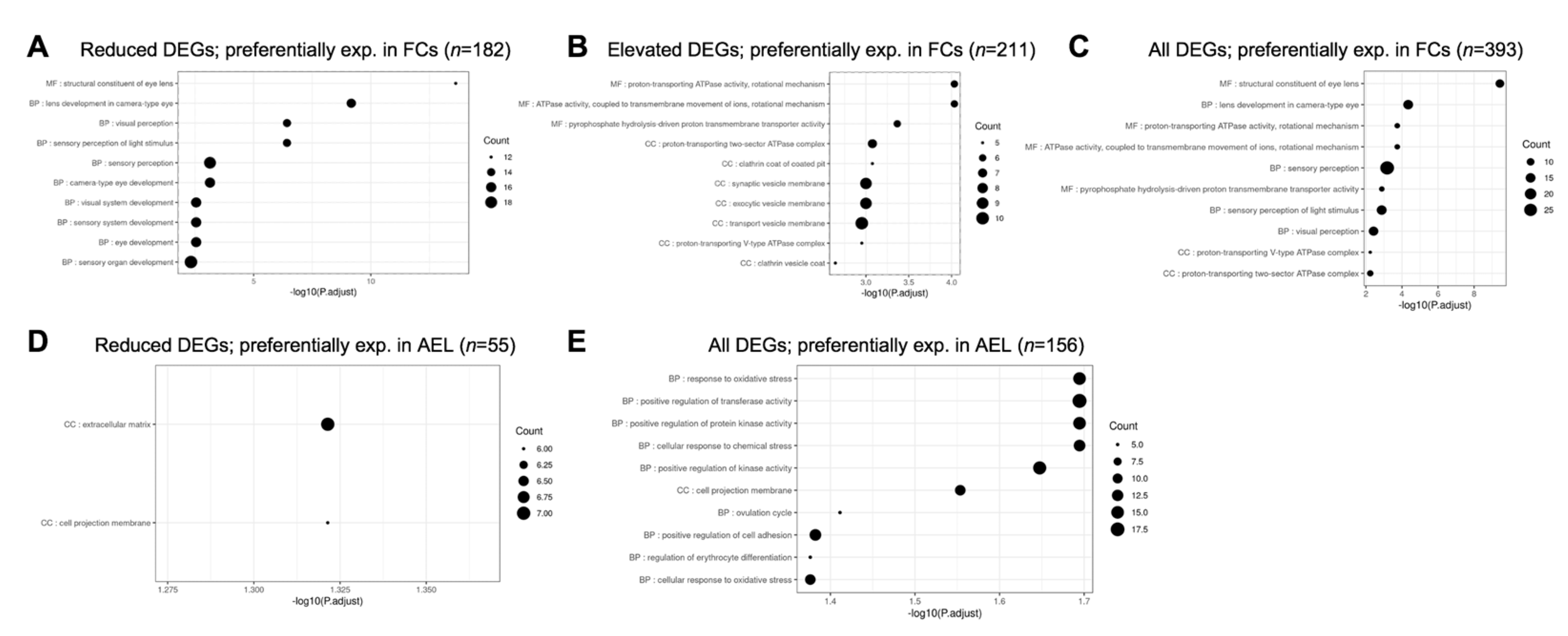
3.4.3. Prioritization of Celf1cKO Lens DEGs Using Isolated Epithelial and Fiber Cell Transcriptome Data
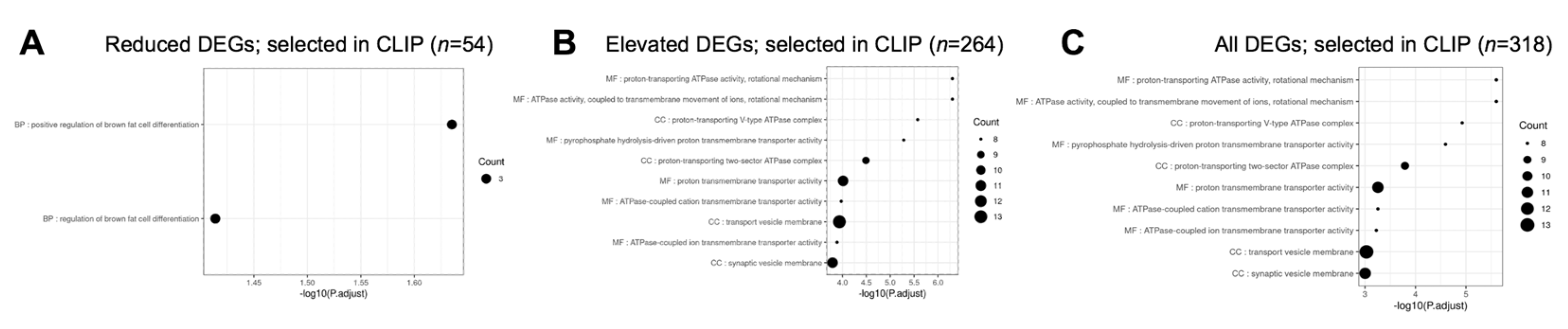
3.4.4. Prioritization of Celf1cKO Lens DEGs Using CLIP-Seq Data Identifying Direct RNA Targets of CELF1 Protein in a Human Cell Line
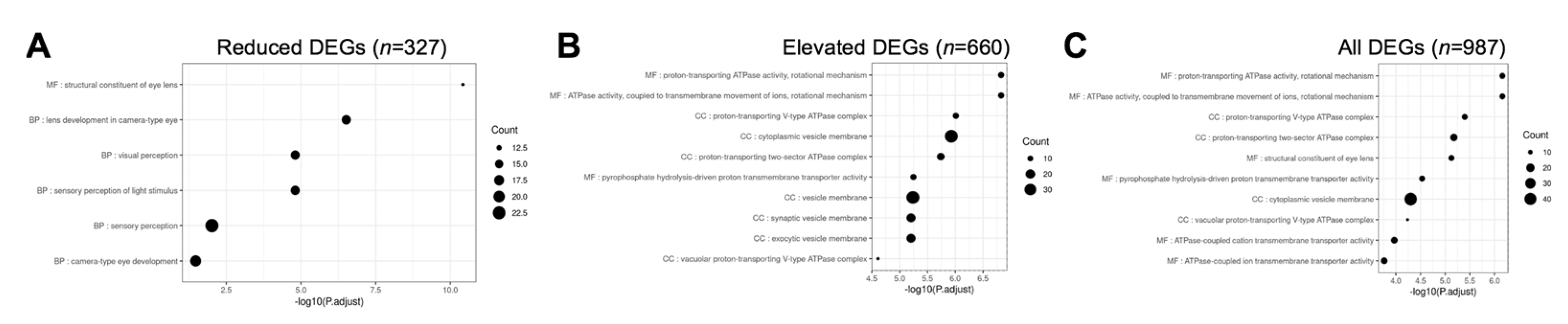
3.5. Gene Ontology and Pathway Analysis of Celf1cKO Lens DEGs
3.6. Comparative Analysis of Celf1cKO Lens DEGs Identified by RNA-Seq and Microarrays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harland, R.M.; Grainger, R.M. Xenopus Research: Metamorphosed by Genetics and Genomics. Trends Genet. 2011, 27, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graw, J. Mouse Models of Cataract. J. Genet. 2009, 88, 469–486. [Google Scholar] [CrossRef]
- Shiels, A.; Hejtmancik, J.F. Inherited Cataracts: Genetic Mechanisms and Pathways New and Old. Exp. Eye Res. 2021, 209, 108662. [Google Scholar] [CrossRef]
- Cvekl, A.; Zhang, X. Signaling and Gene Regulatory Networks in Mammalian Lens Development. Trends Genet. 2017, 33, 677–702. [Google Scholar] [CrossRef]
- Lachke, S.A.; Maas, R.L. Building the Developmental Oculome: Systems Biology in Vertebrate Eye Development and Disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 305–323. [Google Scholar] [CrossRef] [Green Version]
- Lachke, S.A. RNA-Binding Proteins and Post-Transcriptional Regulation in Lens Biology and Cataract: Mediating Spatiotemporal Expression of Key Factors That Control the Cell Cycle, Transcription, Cytoskeleton and Transparency. Exp. Eye Res. 2022, 214, 108889. [Google Scholar] [CrossRef]
- Dash, S.; Siddam, A.D.; Barnum, C.E.; Janga, S.C.; Lachke, S.A. RNA-Binding Proteins in Eye Development and Disease: Implication of Conserved RNA Granule Components. Wiley Interdiscip. Rev. RNA 2016, 7, 527–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, S.; Dang, C.A.; Beebe, D.C.; Lachke, S.A. Deficiency of the RNA Binding Protein Caprin2 Causes Lens Defects and Features of Peters Anomaly. Dev. Dyn. 2015, 244, 1313–1327. [Google Scholar] [CrossRef] [Green Version]
- Siddam, A.D.; Gautier-Courteille, C.; Perez-Campos, L.; Anand, D.; Kakrana, A.; Dang, C.A.; Legagneux, V.; Méreau, A.; Viet, J.; Gross, J.M.; et al. The RNA-Binding Protein Celf1 Post-Transcriptionally Regulates P27Kip1 and Dnase2b to Control Fiber Cell Nuclear Degradation in Lens Development. PLoS Genet. 2018, 14, e1007278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, S.; Viet, J.; Weatherbee, B.A.T.; Siddam, A.D.; Hernandez, F.G.; Gautier-Courteille, C.; Paillard, L.; Lachke, S.A. The Cataract-Linked RNA-Binding Protein Celf1 Post-Transcriptionally Controls the Spatiotemporal Expression of the Key Homeodomain Transcription Factors Pax6 and Prox1 in Lens Development. Hum. Genet. 2020, 139, 1541–1554. [Google Scholar] [CrossRef]
- Dash, S.; Brastrom, L.K.; Patel, S.D.; Scott, C.A.; Slusarski, D.C.; Lachke, S.A. The Master Transcription Factor SOX2, Mutated in Anophthalmia/Microphthalmia, Is Post-Transcriptionally Regulated by the Conserved RNA-Binding Protein RBM24 in Vertebrate Eye Development. Hum. Mol. Genet. 2020, 29, 591–604. [Google Scholar] [CrossRef]
- Lachke, S.A.; Alkuraya, F.S.; Kneeland, S.C.; Ohn, T.; Aboukhalil, A.; Howell, G.R.; Saadi, I.; Cavallesco, R.; Yue, Y.; Tsai, A.C.-H.; et al. Mutations in the RNA Granule Component TDRD7 Cause Cataract and Glaucoma. Science 2011, 331, 1571–1576. [Google Scholar] [CrossRef] [Green Version]
- Barnum, C.E.; Al Saai, S.; Patel, S.D.; Cheng, C.; Anand, D.; Xu, X.; Dash, S.; Siddam, A.D.; Glazewski, L.; Paglione, E.; et al. The Tudor-Domain Protein TDRD7, Mutated in Congenital Cataract, Controls the Heat Shock Protein HSPB1 (HSP27) and Lens Fiber Cell Morphology. Hum. Mol. Genet. 2020, 29, 2076–2097. [Google Scholar] [CrossRef] [PubMed]
- Lorén, C.E.; Schrader, J.W.; Ahlgren, U.; Gunhaga, L. FGF Signals Induce Caprin2 Expression in the Vertebrate Lens. Differentiation 2009, 77, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, K.; Shichino, Y.; Iwasaki, S.; Shiina, N. Implications of RNG140 (Caprin2)-Mediated Translational Regulation in Eye Lens Differentiation. J. Biol. Chem. 2020, 295, 15029–15044. [Google Scholar] [CrossRef]
- Shao, M.; Lu, T.; Zhang, C.; Zhang, Y.-Z.; Kong, S.-H.; Shi, D.-L. Rbm24 Controls Poly(A) Tail Length and Translation Efficiency of Crystallin MRNAs in the Lens via Cytoplasmic Polyadenylation. Proc. Natl. Acad. Sci. USA 2020, 117, 7245–7254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, Q.; Cabrera, P.E.; Zhong, Z.; Sun, W.; Jiao, X.; Chen, Y.; Govindarajan, G.; Naeem, M.A.; Khan, S.N.; et al. Molecular Genetic Analysis of Pakistani Families with Autosomal Recessive Congenital Cataracts by Homozygosity Screening. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2207–2217. [Google Scholar] [CrossRef]
- Fernández-Alcalde, C.; Nieves-Moreno, M.; Noval, S.; Peralta, J.M.; Montaño, V.E.F.; Del Pozo, Á.; Santos-Simarro, F.; Vallespín, E. Molecular and Genetic Mechanism of Non-Syndromic Congenital Cataracts. Mutation Screening in Spanish Families. Genes 2021, 12, 580. [Google Scholar] [CrossRef]
- Kandaswamy, D.K.; Prakash, M.V.S.; Graw, J.; Koller, S.; Magyar, I.; Tiwari, A.; Berger, W.; Santhiya, S.T. Application of WES Towards Molecular Investigation of Congenital Cataracts: Identification of Novel Alleles and Genes in a Hospital-Based Cohort of South India. Int. J. Mol. Sci. 2020, 21, 9569. [Google Scholar] [CrossRef]
- Tan, Y.-Q.; Tu, C.; Meng, L.; Yuan, S.; Sjaarda, C.; Luo, A.; Du, J.; Li, W.; Gong, F.; Zhong, C.; et al. Loss-of-Function Mutations in TDRD7 Lead to a Rare Novel Syndrome Combining Congenital Cataract and Nonobstructive Azoospermia in Humans. Genet. Med. 2019, 21, 1209–1217. [Google Scholar] [CrossRef]
- Tanaka, T.; Hosokawa, M.; Vagin, V.V.; Reuter, M.; Hayashi, E.; Mochizuki, A.L.; Kitamura, K.; Yamanaka, H.; Kondoh, G.; Okawa, K.; et al. Tudor Domain Containing 7 (Tdrd7) Is Essential for Dynamic Ribonucleoprotein (RNP) Remodeling of Chromatoid Bodies during Spermatogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 10579–10584. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Wu, M.; He, C.-Y.; An, X.-J.; Sun, M.; Chen, C.-L.; Ye, J. RNA Granule Component TDRD7 Gene Polymorphisms in a Han Chinese Population with Age-Related Cataract. J. Int. Med. Res. 2014, 42, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choquet, H.; Melles, R.B.; Anand, D.; Yin, J.; Cuellar-Partida, G.; Wang, W.; 23andMe Research Team; Hoffmann, T.J.; Nair, K.S.; Hysi, P.G.; et al. A Large Multiethnic GWAS Meta-Analysis of Cataract Identifies New Risk Loci and Sex-Specific Effects. Nat. Commun. 2021, 12, 3595. [Google Scholar] [CrossRef] [PubMed]
- Bauermeister, D.; Claußen, M.; Pieler, T. A Novel Role for Celf1 in Vegetal RNA Localization during Xenopus Oogenesis. Dev. Biol. 2015, 405, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlasova-St Louis, I.; Dickson, A.M.; Bohjanen, P.R.; Wilusz, C.J. CELFish Ways to Modulate MRNA Decay. Biochim. Biophys. Acta 2013, 1829, 695–707. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Miskimins, W.K. CUG-Binding Protein Represses Translation of P27Kip1 MRNA through Its Internal Ribosomal Entry Site. RNA Biol. 2011, 8, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Ladd, A.N.; Charlet, N.; Cooper, T.A. The CELF Family of RNA Binding Proteins Is Implicated in Cell-Specific and Developmentally Regulated Alternative Splicing. Mol. Cell. Biol. 2001, 21, 1285–1296. [Google Scholar] [CrossRef] [Green Version]
- Vlasova, I.A.; Tahoe, N.M.; Fan, D.; Larsson, O.; Rattenbacher, B.; Sternjohn, J.R.; Vasdewani, J.; Karypis, G.; Reilly, C.S.; Bitterman, P.B.; et al. Conserved GU-Rich Elements Mediate MRNA Decay by Binding to CUG-Binding Protein 1. Mol. Cell. 2008, 29, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Rowan, S.; Conley, K.W.; Le, T.T.; Donner, A.L.; Maas, R.L.; Brown, N.L. Notch Signaling Regulates Growth and Differentiation in the Mammalian Lens. Dev. Biol. 2008, 321, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Lachke, S.A.; Ho, J.W.K.; Kryukov, G.V.; O’Connell, D.J.; Aboukhalil, A.; Bulyk, M.L.; Park, P.J.; Maas, R.L. ISyTE: Integrated Systems Tool for Eye Gene Discovery. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Rowan, S.; Siggers, T.; Lachke, S.A.; Yue, Y.; Bulyk, M.L.; Maas, R.L. Precise Temporal Control of the Eye Regulatory Gene Pax6 via Enhancer-Binding Site Affinity. Genes. Dev. 2010, 24, 980–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From Reads to Genes to Pathways: Differential Expression Analysis of RNA-Seq Experiments Using Rsubread and the EdgeR Quasi-Likelihood Pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef] [Green Version]
- Shiels, A.; Bennett, T.M.; Hejtmancik, J.F. Cat-Map: Putting Cataract on the Map. Mol. Vis. 2010, 16, 2007–2015. [Google Scholar]
- Kakrana, A.; Yang, A.; Anand, D.; Djordjevic, D.; Ramachandruni, D.; Singh, A.; Huang, H.; Ho, J.W.K.; Lachke, S.A. ISyTE 2.0: A Database for Expression-Based Gene Discovery in the Eye. Nucleic Acids Res. 2018, 46, D875–D885. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, D.; Cvekl, A. A Comprehensive Spatial-Temporal Transcriptomic Analysis of Differentiating Nascent Mouse Lens Epithelial and Fiber Cells. Exp. Eye Res. 2018, 175, 56–72. [Google Scholar] [CrossRef]
- Le Tonquèze, O.; Gschloessl, B.; Legagneux, V.; Paillard, L.; Audic, Y. Identification of CELF1 RNA Targets by CLIP-Seq in Human HeLa Cells. Genom. Data 2016, 8, 97–103. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Agrawal, S.A.; Anand, D.; Siddam, A.D.; Kakrana, A.; Dash, S.; Scheiblin, D.A.; Dang, C.A.; Terrell, A.M.; Waters, S.M.; Singh, A.; et al. Compound Mouse Mutants of BZIP Transcription Factors Mafg and Mafk Reveal a Regulatory Network of Non-Crystallin Genes Associated with Cataract. Hum. Genet. 2015, 134, 717–735. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-Performance Genomics Data Visualization and Exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Beisang, D.; Bohjanen, P.R.; Vlasova-St. Louis, I.A. CELF1, a Multifunctional Regulator of Posttranscriptional Networks. In Binding Protein; Abdelmohsen, K., Ed.; InTech: London, UK, 2012; ISBN 978-953-51-0758-3. [Google Scholar]
- Kress, C.; Gautier-Courteille, C.; Osborne, H.B.; Babinet, C.; Paillard, L. Inactivation of CUG-BP1/CELF1 Causes Growth, Viability, and Spermatogenesis Defects in Mice. Mol. Cell. Biol. 2007, 27, 1146–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifdaloz, M.; Osterloh, L.; Graña, O.; Riveiro-Falkenbach, E.; Ximénez-Embún, P.; Muñoz, J.; Tejedo, C.; Calvo, T.G.; Karras, P.; Olmeda, D.; et al. Systems Analysis Identifies Melanoma-Enriched pro-Oncogenic Networks Controlled by the RNA Binding Protein CELF1. Nat. Commun. 2017, 8, 2249. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, A.; Cheema, S.; Fachini, J.M.; Kongchan, N.; Lu, G.; Simon, L.M.; Wang, T.; Mao, S.; Rosen, D.G.; Ittmann, M.M.; et al. CELF1 Is a Central Node in Post-Transcriptional Regulatory Programmes Underlying EMT. Nat. Commun. 2016, 7, 13362. [Google Scholar] [CrossRef] [PubMed]
- House, R.P.; Talwar, S.; Hazard, E.S.; Hill, E.G.; Palanisamy, V. RNA-Binding Protein CELF1 Promotes Tumor Growth and Alters Gene Expression in Oral Squamous Cell Carcinoma. Oncotarget 2015, 6, 43620–43634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, T.; Sasaki, A.; Akazawa, N.; Otani, H.; Bessho, Y. Celf1 Regulation of Dmrt2a Is Required for Somite Symmetry and Left-Right Patterning during Zebrafish Development. Development 2012, 139, 3553–3560. [Google Scholar] [CrossRef] [Green Version]
- Timchenko, N.A.; Cai, Z.J.; Welm, A.L.; Reddy, S.; Ashizawa, T.; Timchenko, L.T. RNA CUG Repeats Sequester CUGBP1 and Alter Protein Levels and Activity of CUGBP1. J. Biol. Chem. 2001, 276, 7820–7826. [Google Scholar] [CrossRef] [Green Version]
- Philips, A.V.; Timchenko, L.T.; Cooper, T.A. Disruption of Splicing Regulated by a CUG-Binding Protein in Myotonic Dystrophy. Science 1998, 280, 737–741. [Google Scholar] [CrossRef]
- Xiao, J.; Jin, S.; Wang, X.; Huang, J.; Zou, H. CELF1 Selectively Regulates Alternative Splicing of DNA Repair Genes Associated With Cataract in Human Lens Cell Line. Biochem. Genet. 2022. [Google Scholar] [CrossRef]
- Xiao, J.; Tian, X.; Jin, S.; He, Y.; Song, M.; Zou, H. CELF1 Promotes Matrix Metalloproteinases Gene Expression at Transcriptional Level in Lens Epithelial Cells. BMC Ophthalmol. 2022, 22, 122. [Google Scholar] [CrossRef]
- Nishimoto, S.; Kawane, K.; Watanabe-Fukunaga, R.; Fukuyama, H.; Ohsawa, Y.; Uchiyama, Y.; Hashida, N.; Ohguro, N.; Tano, Y.; Morimoto, T.; et al. Nuclear Cataract Caused by a Lack of DNA Degradation in the Mouse Eye Lens. Nature 2003, 424, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, D.T.; Lyon, G.J.; Carlton, M.B.; Sanes, J.R.; Cunningham, J.M.; Anderson, J.R.; Hogan, B.L.; Evans, M.J.; Colledge, W.H. Mice Deficient for the Secreted Glycoprotein SPARC/Osteonectin/BM40 Develop Normally but Show Severe Age-Onset Cataract Formation and Disruption of the Lens. EMBO J. 1998, 17, 1860–1870. [Google Scholar] [CrossRef] [PubMed]
- Anand, D.; Lachke, S.A. Systems Biology of Lens Development: A Paradigm for Disease Gene Discovery in the Eye. Exp. Eye Res. 2017, 156, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Anand, D.; Kakrana, A.; Siddam, A.D.; Huang, H.; Saadi, I.; Lachke, S.A. RNA Sequencing-Based Transcriptomic Profiles of Embryonic Lens Development for Cataract Gene Discovery. Hum. Genet. 2018, 137, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Aryal, S.; Anand, D.; Hernandez, F.G.; Weatherbee, B.A.T.; Huang, H.; Reddy, A.P.; Wilmarth, P.A.; David, L.L.; Lachke, S.A. MS/MS in Silico Subtraction-Based Proteomic Profiling as an Approach to Facilitate Disease Gene Discovery: Application to Lens Development and Cataract. Hum. Genet. 2020, 139, 151–184. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | logFC | logCPM | FDR | Symbol | logFC | logCPM | FDR |
|---|---|---|---|---|---|---|---|
| Lgsn | −3.957 | 4.826 | 0.02 | Tdrd7 | −0.673 | 10.755 | 0.02 |
| Gjb6 | −3.08 | 1.224 | 0.04 | Cryba4 | −0.671 | 13.735 | 0.02 |
| Dnase2b | −2.565 | 6.866 | 0.03 | Pxdn | −0.667 | 8.535 | 0.04 |
| Lctl | −1.985 | 8.025 | 0.01 | Flnb | −0.664 | 5.836 | 0.02 |
| Crybb2 | −1.763 | 11.087 | 0.03 | Crybb1 | −0.623 | 14.92 | 0.02 |
| Celf1 | −1.511 | 9.204 | 0.04 | Jag1 | 0.582 | 8.78 | 0.04 |
| Bfsp1 | −1.428 | 12.513 | 0.02 | Psmc3 | 0.587 | 7.955 | 0.03 |
| Adgrl2 | −1.326 | 5.119 | 0.03 | Ube2a | 0.597 | 4.673 | 0.03 |
| Crygb | −1.275 | 14.257 | 0.02 | Nploc4 | 0.6 | 6.468 | 0.02 |
| Cryba2 | −1.195 | 13.429 | 0.02 | Sec23a | 0.681 | 7.201 | 0.01 |
| Gja3 | −1.12 | 10.1 | 0.02 | Klc1 | 0.724 | 8.255 | 0.02 |
| Dnmbp | −1.109 | 8.04 | 0.01 | Ercc6 | 0.803 | 4.453 | 0.02 |
| Ulk4 | −1.07 | 2.037 | 0.02 | Atm | 0.811 | 4.526 | 0.05 |
| Crygd | −0.973 | 14.5 | 0.01 | Rnf149 | 0.857 | 3.842 | 0.02 |
| Cryga | −0.968 | 13.166 | 0.01 | Agps | 0.907 | 5.422 | 0.02 |
| Sparc | −0.874 | 10.326 | 0.02 | Ptn | 1.047 | 5.652 | 0.01 |
| Sord | −0.815 | 4.669 | 0.02 | Polr3b | 1.08 | 4.807 | 0.02 |
| Loxl1 | −0.793 | 6.509 | 0.02 | Pqbp1 | 1.247 | 5.673 | 0.01 |
| Crygc | −0.788 | 13.425 | 0.01 | Pgrmc1 | 1.258 | 6.345 | 0.02 |
| Ace | −0.785 | 5.561 | 0.046 | Wfs1 | 1.421 | 5.521 | 0.01 |
| Fkrp | −0.745 | 4.324 | 0.022 | Mafa | 1.684 | 5.134 | 0.01 |
| Cryab | −0.73 | 13.439 | 0.021 |
| Total DEGs (n = 987) | Reduced in Celf1cKO Lens (n = 327) | Elevated in Celf1cKO Lens (n = 660) |
|---|---|---|
| Preferentially exp. in FCs | 182 | 211 |
| Preferentially exp. in AEL | 55 | 101 |
| Not preferentially exp. in FCs or AEL | 90 | 348 |
| DEGs Selected by CLIP (n = 297 1) | Reduced in Celf1cKO Lens (n = 50) | Elevated in Celf1cKO Lens (n = 247) |
|---|---|---|
| Lens enriched-exp. in iSyTE | 26 | 77 |
| Not lens enriched-exp. in iSyTE | 24 | 170 |
| DEGs Selected by CLIP (n = 318) | Reduced in Celf1cKO Lens (n = 54) | Elevated in Celf1cKO Lens (n = 264) |
|---|---|---|
| Preferentially exp. in FCs | 30 | 74 |
| Preferentially exp. in AEL | 9 | 41 |
| Not preferentially exp. in FCs or AEL | 15 | 149 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddam, A.D.; Duot, M.; Coomson, S.Y.; Anand, D.; Aryal, S.; Weatherbee, B.A.T.; Audic, Y.; Paillard, L.; Lachke, S.A. High-Throughput Transcriptomics of Celf1 Conditional Knockout Lens Identifies Downstream Networks Linked to Cataract Pathology. Cells 2023, 12, 1070. https://doi.org/10.3390/cells12071070
Siddam AD, Duot M, Coomson SY, Anand D, Aryal S, Weatherbee BAT, Audic Y, Paillard L, Lachke SA. High-Throughput Transcriptomics of Celf1 Conditional Knockout Lens Identifies Downstream Networks Linked to Cataract Pathology. Cells. 2023; 12(7):1070. https://doi.org/10.3390/cells12071070
Chicago/Turabian StyleSiddam, Archana D., Matthieu Duot, Sarah Y. Coomson, Deepti Anand, Sandeep Aryal, Bailey A. T. Weatherbee, Yann Audic, Luc Paillard, and Salil A. Lachke. 2023. "High-Throughput Transcriptomics of Celf1 Conditional Knockout Lens Identifies Downstream Networks Linked to Cataract Pathology" Cells 12, no. 7: 1070. https://doi.org/10.3390/cells12071070