
StressMatic: A Novel Automated System to Induce Depressive- and Anxiety-like Phenotype in Rats
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Rack Development & Stress Categories
2.3. Weight Gain Monitoring
2.4. Sucrose Consumption Test (SCT)
2.5. Sweet Drive Test (SDT)
2.6. Elevated Plus Maze Test (EPM)
2.7. Novelty Suppressed Feeding Test (NSF)
2.8. Novel Object Recognition Test (NOR)
2.9. Forced Swim Test (FST)
2.10. Corticosterone Levels Measurements
2.11. Immunostaining Procedures
2.12. Statistical Analysis
3. Results
3.1. Behavioral Validation of the auCMS Protocol
3.2. Endocrine Stress-Induced Changes
3.3. Cellular Proliferation in the Hippocampus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans-Lacko, S.; Aguilar-Gaxiola, S.; Al-Hamzawi, A.; Alonso, J.; Benjet, C.; Bruffaerts, R.; Chiu, W.T.; Florescu, S.; de Girolamo, G.; Gureje, O.; et al. Socio-Economic Variations in the Mental Health Treatment Gap for People with Anxiety, Mood, and Substance Use Disorders: Results from the WHO World Mental Health (WMH) Surveys. Psychol. Med. 2018, 48, 1560–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, S.H. Core Symptoms of Major Depressive Disorder: Relevance to Diagnosis and Treatment. Dialogues Clin. Neurosci. 2008, 10, 271–277. [Google Scholar] [CrossRef]
- Epel, E.S.; Crosswell, A.D.; Mayer, S.E.; Prather, A.A.; Slavich, G.M.; Puterman, E.; Mendes, W.B. More than a Feeling: A Unified View of Stress Measurement for Population Science. Front. Neuroendocrinol. 2018, 49, 146–169. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, A.; Reif, A.; Cryan, J.F.; Slattery, D.A. The Future of Rodent Models in Depression Research. Nat. Rev. Neurosci. 2019, 20, 686–701. [Google Scholar] [CrossRef]
- Hao, Y.; Ge, H.; Sun, M.; Gao, Y. Selecting an Appropriate Animal Model of Depression. Int. J. Mol. Sci. 2019, 20, 4827. [Google Scholar] [CrossRef] [Green Version]
- Planchez, B.; Surget, A.; Belzung, C. Animal Models of Major Depression: Drawbacks and Challenges. J. Neural Transm. 2019, 126, 1383–1408. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Timberlake, M.A.; Prall, K.; Dwivedi, Y. The Recent Progress in Animal Models of Depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 77, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Tornese, P.; Sala, N.; Bonini, D.; Bonifacino, T.; la Via, L.; Milanese, M.; Treccani, G.; Seguini, M.; Ieraci, A.; Mingardi, J.; et al. Chronic Mild Stress Induces Anhedonic Behavior and Changes in Glutamate Release, BDNF Trafficking and Dendrite Morphology Only in Stress Vulnerable Rats. The Rapid Restorative Action of Ketamine. Neurobiol. Stress 2019, 10, 100160. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.; Donovan, M.H.; Ross, M.N.; Richardson, D.R.; Reister, R.; Farnbauch, L.A.; Fischer, S.J.; Riethmacher, D.; Gershenfeld, H.K.; Lagace, D.C.; et al. Stress-Induced Anxiety- and Depressive-like Phenotype Associated with Transient Reduction in Neurogenesis in Adult Nestin-CreERT2/Diphtheria Toxin Fragment A Transgenic Mice. PLoS ONE 2016, 11, e0147256. [Google Scholar] [CrossRef] [PubMed]
- Algamal, M.; Ojo, J.O.; Lungmus, C.P.; Muza, P.; Cammarata, C.; Owens, M.J.; Mouzon, B.C.; Diamond, D.M.; Mullan, M.; Crawford, F. Chronic Hippocampal Abnormalities and Blunted HPA Axis in an Animal Model of Repeated Unpredictable Stress. Front. Behav. Neurosci. 2018, 12, 150. [Google Scholar] [CrossRef]
- Herman, J.P. Regulation of the HPA Axis by Acute and Chronic Stress. In Encyclopedia of Behavioral Neuroscience; Elsevier: Amsterdam, The Netherlands, 2010; pp. 149–153. [Google Scholar]
- Karin, O.; Raz, M.; Tendler, A.; Bar, A.; Korem Kohanim, Y.; Milo, T.; Alon, U. A New Model for the HPA Axis Explains Dysregulation of Stress Hormones on the Timescale of Weeks. Mol. Syst. Biol. 2020, 16, e9510. [Google Scholar] [CrossRef] [PubMed]
- Starr, L.R.; Dienes, K.; Li, Y.I.; Shaw, Z.A. Chronic Stress Exposure, Diurnal Cortisol Slope, and Implications for Mood and Fatigue: Moderation by Multilocus HPA-Axis Genetic Variation. Psychoneuroendocrinology 2019, 100, 156–163. [Google Scholar] [CrossRef]
- Willner, P. Reliability of the Chronic Mild Stress Model of Depression: A User Survey. Neurobiol. Stress 2017, 6, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willner, P. The Chronic Mild Stress (CMS) Model of Depression: History, Evaluation and Usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [Green Version]
- Patrício, P.; Mateus-Pinheiro, A.; Irmler, M.; Alves, N.D.; Machado-Santos, A.R.; Morais, M.; Correia, J.S.; Korostynski, M.; Piechota, M.; Stoffel, R.; et al. Differential and Converging Molecular Mechanisms of Antidepressants’ Action in the Hippocampal Dentate Gyrus. Neuropsychopharmacology 2015, 40, 338–349. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.R.; Geiger, L.; Wiborg, O.; Czéh, B. Stress-Induced Morphological, Cellular and Molecular Changes in the Brain-Lessons Learned from the Chronic Mild Stress Model of Depression. Cells 2020, 9, 1026. [Google Scholar] [CrossRef] [Green Version]
- Morais, M.; Santos, P.A.R.; Mateus-Pinheiro, A.; Patrício, P.; Pinto, L.; Sousa, N.; Pedroso, P.; Almeida, S.; Filipe, A.; Bessa, J.M. The Effects of Chronic Stress on Hippocampal Adult Neurogenesis and Dendritic Plasticity Are Reversed by Selective MAO-A Inhibition. J. Psychopharmacol. 2014, 28, 1178–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessa, J.M.; Mesquita, A.R.; Oliveira, M.; Pêgo, J.M.; Cerqueira, J.J.; Palha, J.A.; Almeida, O.F.X.; Sousa, N. A Trans-Dimensional Approach to the Behavioral Aspects of Depression. Front. Behav. Neurosci. 2009, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Bessa, J.M.; Ferreira, D.; Melo, I.; Marques, F.; Cerqueira, J.J.; Palha, J.A.; Almeida, O.F.X.; Sousa, N. The Mood-Improving Actions of Antidepressants Do Not Depend on Neurogenesis but Are Associated with Neuronal Remodeling. Mol. Psychiatry 2009, 14, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Mateus-Pinheiro, A.; Patrício, P.; Alves, N.D.; Machado-Santos, A.R.; Morais, M.; Bessa, J.M.; Sousa, N.; Pinto, L. The Sweet Drive Test: Refining Phenotypic Characterization of Anhedonic Behavior in Rodents. Front. Behav. Neurosci. 2014, 8, 74. [Google Scholar] [CrossRef]
- Cox, B.M.; Alsawah, F.; McNeill, P.C.; Galloway, M.P.; Perrine, S.A. Neurochemical, Hormonal, and Behavioral Effects of Chronic Unpredictable Stress in the Rat. Behav. Brain Res. 2011, 220, 106–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.N.; Hellemans, K.G.C.; Verma, P.; Gorzalka, B.B.; Weinberg, J. Neurobiology of Chronic Mild Stress: Parallels to Major Depression. Neurosci. Biobehav. Rev. 2012, 36, 2085–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, V.; Nestler, E.J. Animal Models of Depression: Molecular Perspectives. Curr. Top. Behav. Neurosci. 2011, 7, 121–147. [Google Scholar] [CrossRef] [Green Version]
- Nestler, E.J.; Gould, E.; Manji, H. Preclinical Models: Status of Basic Research in Depression. Biol. Psychiatry 2002, 52, 503–528. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Leonard, B.E. The Olfactory Bulbectomised Rat as a Model of Depression. Neurosci. Biobehav. Rev. 2005, 29, 627–647. [Google Scholar] [CrossRef]
- Song, J.; Kim, Y.K. Animal Models for the Study of Depressive Disorder. CNS Neurosci. Ther. 2021, 27, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Will, C.C.; Aird, F.; Redei, E.E. Selectively Bred Wistar-Kyoto Rats: An Animal Model of Depression and Hyper-Responsiveness to Antidepressants. Mol. Psychiatry 2003, 8, 925–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willner, P. Stress and Depression: Insights from Animal Models. Stress Med. 1997, 13, 229–233. [Google Scholar] [CrossRef]
- Willner, P. Validity, Reliability and Utility of the Chronic Mild Stress Model of Depression: A 10-Year Review and Evaluation. Psychopharmacology 1997, 134, 319–329. [Google Scholar] [CrossRef]
- Willner, P. Chronic Mild Stress (CMS) Revisited: Consistency and Behavioural- Neurobiological Concordance in the Effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef]
- Becker, M.; Pinhasov, A.; Ornoy, A. Animal Models of Depression: What Can They Teach Us about the Human Disease? Diagnostics 2021, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Holland, I.; Davies, J.A. Automation in the Life Science Research Laboratory. Front. Bioeng. Biotechnol. 2020, 8, 571777. [Google Scholar] [CrossRef] [PubMed]
- Ottenweller, J.E.; Servatius, R.J.; Natelson, B.H.; Servatius, R.J.; Natelson, B.H. Repeated Stress Persistently Elevates Morning, But Not Evening, Plasma Corticosterone Levels in Male Rats. Physiol. Behav. 1994, 55, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Balu, D.T.; Lucki, I. Adult Hippocampal Neurogenesis: Regulation, Functional Implications, and Contribution to Disease Pathology. Neurosci. Biobehav. Rev. 2009, 33, 232–252. [Google Scholar] [CrossRef] [Green Version]
- Patrício, P.; Mateus-Pinheiro, A.; Sousa, N.; Pinto, L. Re-Cycling Paradigms: Cell Cycle Regulation in Adult Hippocampal Neurogenesis and Implications for Depression. Mol. Neurobiol. 2013, 48, 84–96. [Google Scholar] [CrossRef] [PubMed]





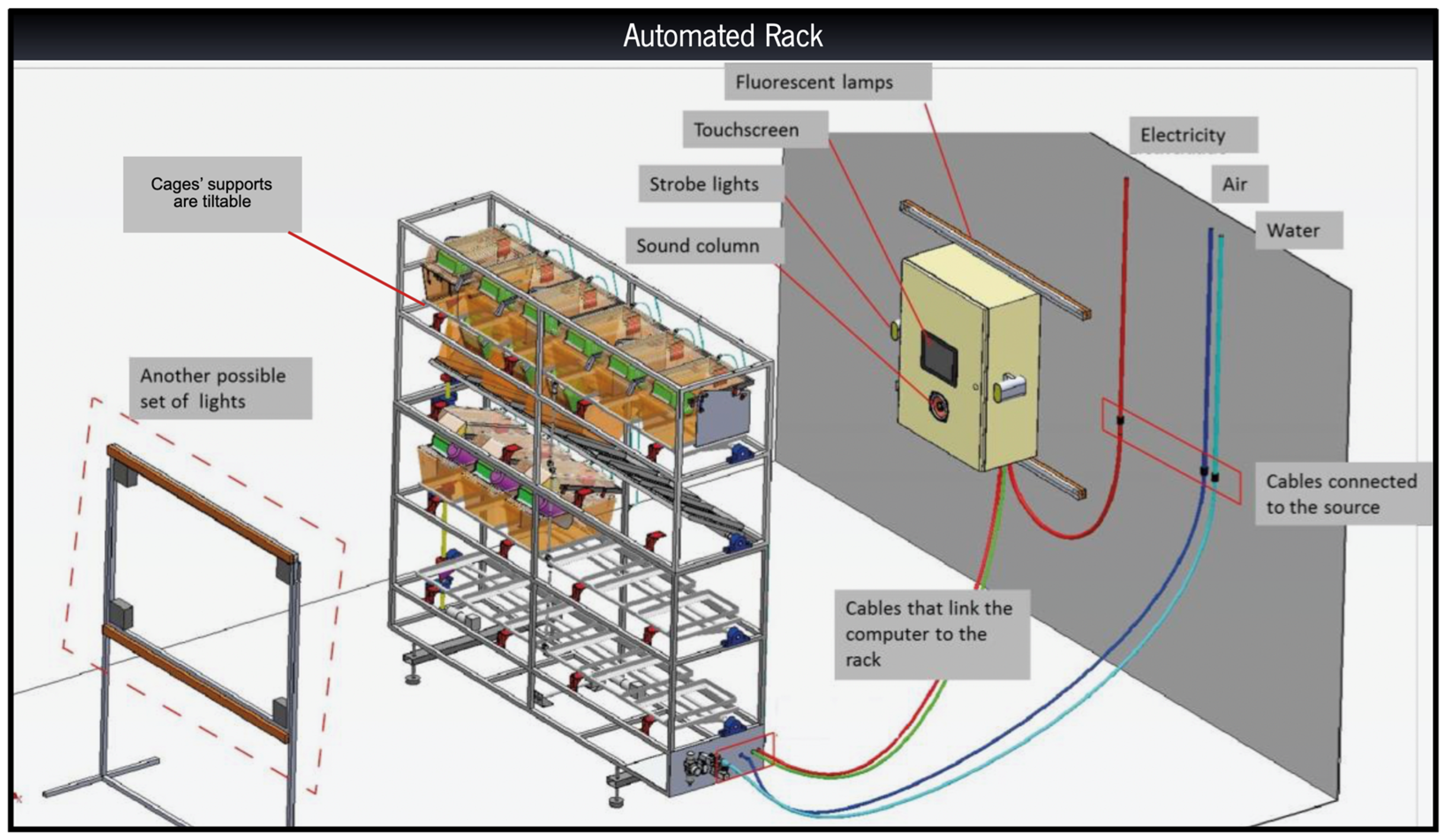{kind=link}
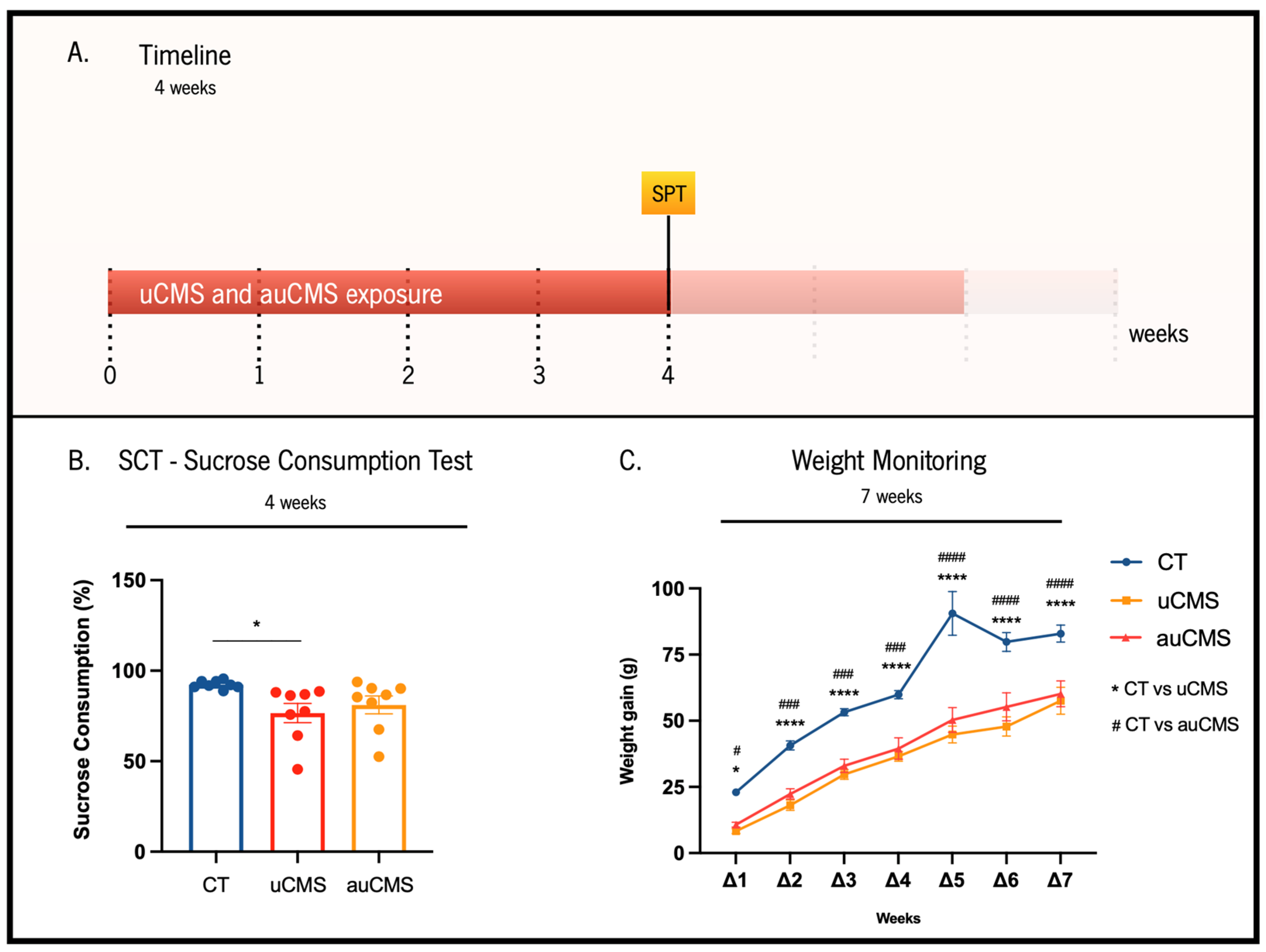{kind=link}
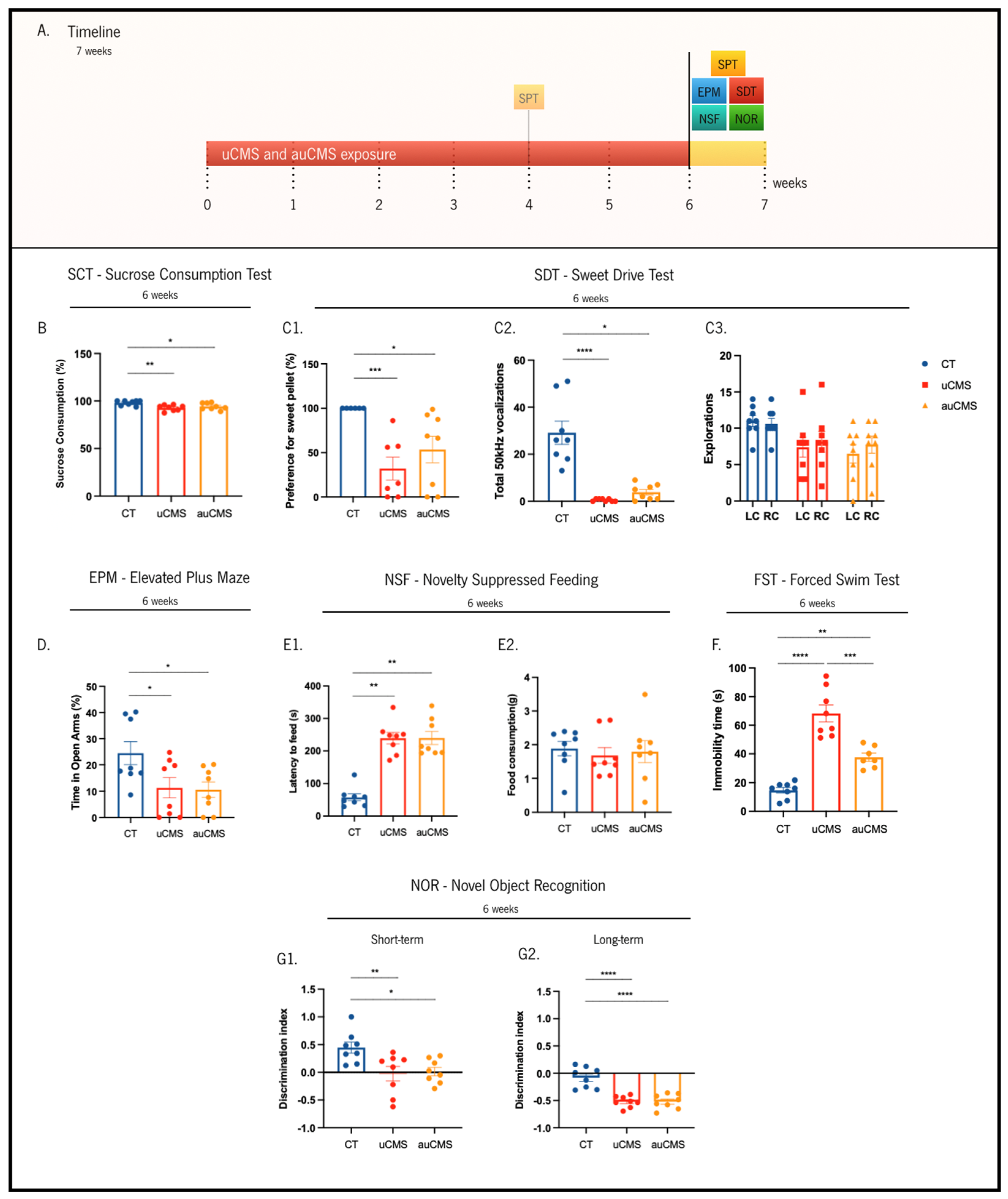{kind=link}
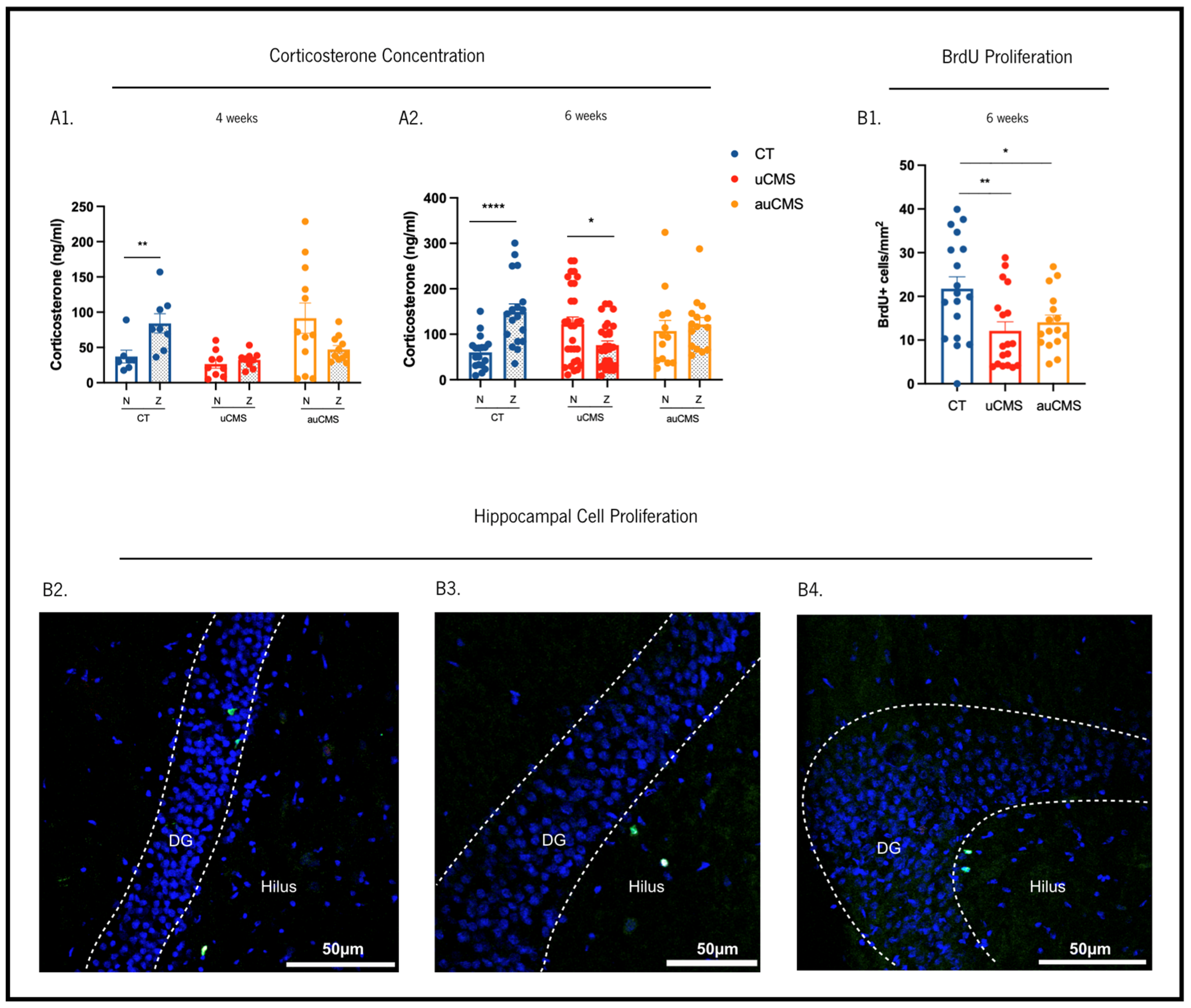{kind=link}
| Fully automated | A. | Tilted cage (approximately 45°) |
| B. | Housing on damp bedding during the night | |
| C. | Overnight illumination | |
| D. | Inverted light/dark cycle | |
| E. | Exposure to strobe lights | |
| F. | Startle noise | |
| Manual | G. | Food deprivation followed by exposure to inaccessible food |
| H. | Water deprivation followed by exposure to an empty bottle | |
| I. | Overcrowding | |
| J. | Cage switch | |
| Partially automated | K. | Confinement to a restricted space |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins-Macedo, J.; Mateus-Pinheiro, A.; Alves, C.; Veloso, F.; Gomes, E.D.; Ribeiro, I.; Correia, J.S.; Silveira-Rosa, T.; Alves, N.D.; Rodrigues, A.J.; et al. StressMatic: A Novel Automated System to Induce Depressive- and Anxiety-like Phenotype in Rats. Cells 2023, 12, 381. https://doi.org/10.3390/cells12030381
Martins-Macedo J, Mateus-Pinheiro A, Alves C, Veloso F, Gomes ED, Ribeiro I, Correia JS, Silveira-Rosa T, Alves ND, Rodrigues AJ, et al. StressMatic: A Novel Automated System to Induce Depressive- and Anxiety-like Phenotype in Rats. Cells. 2023; 12(3):381. https://doi.org/10.3390/cells12030381
Chicago/Turabian StyleMartins-Macedo, Joana, António Mateus-Pinheiro, Cátia Alves, Fernando Veloso, Eduardo D. Gomes, Inês Ribeiro, Joana S. Correia, Tiago Silveira-Rosa, Nuno D. Alves, Ana J. Rodrigues, and et al. 2023. "StressMatic: A Novel Automated System to Induce Depressive- and Anxiety-like Phenotype in Rats" Cells 12, no. 3: 381. https://doi.org/10.3390/cells12030381