
The Male Mouse Meiotic Cilium Emanates from the Mother Centriole at Zygotene Prior to Centrosome Duplication
 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
2.2. Squashing Procedure and Immunofluorescence Microscopy
2.3. Histology
2.4. Quantification Analysis
3. Results
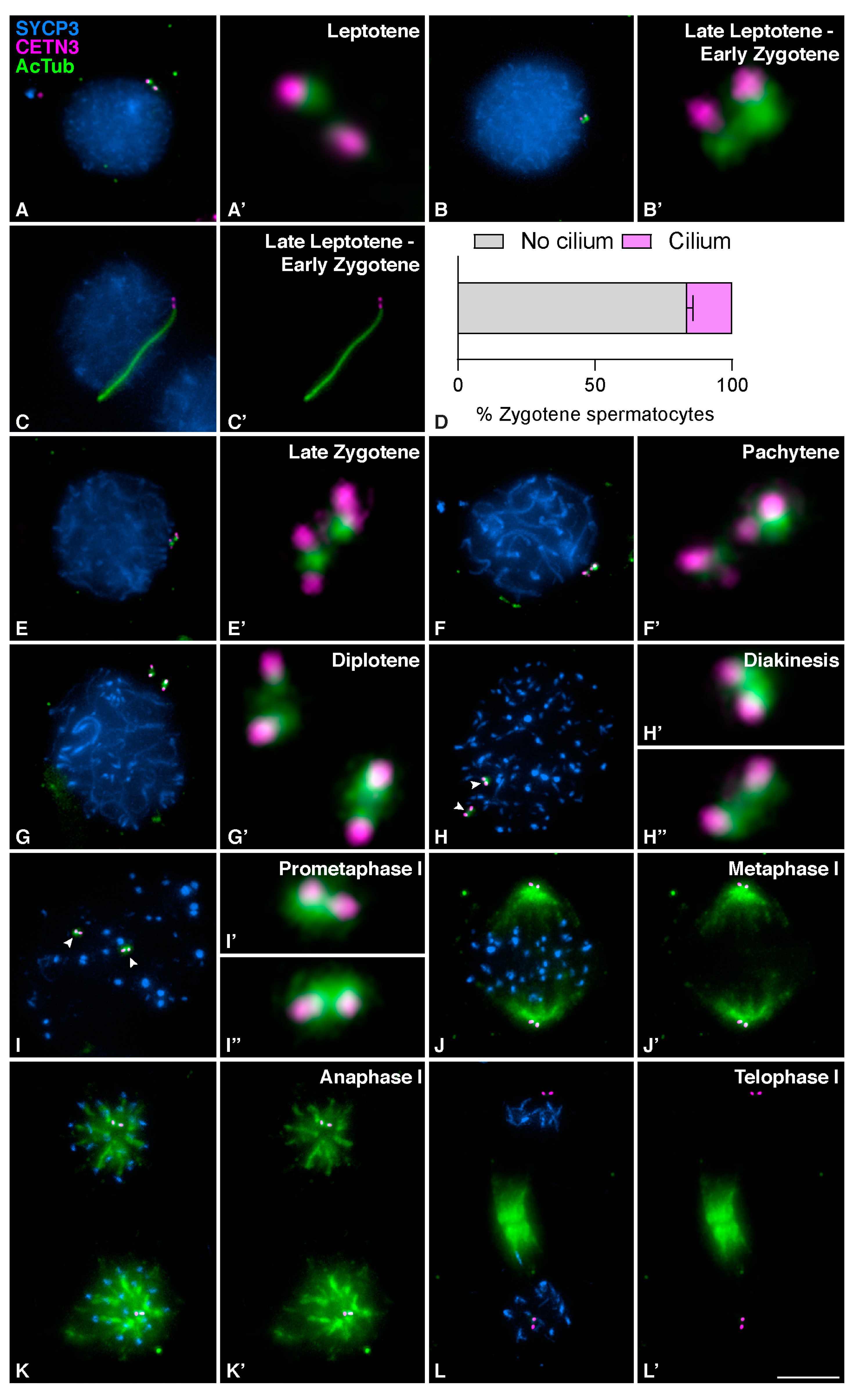
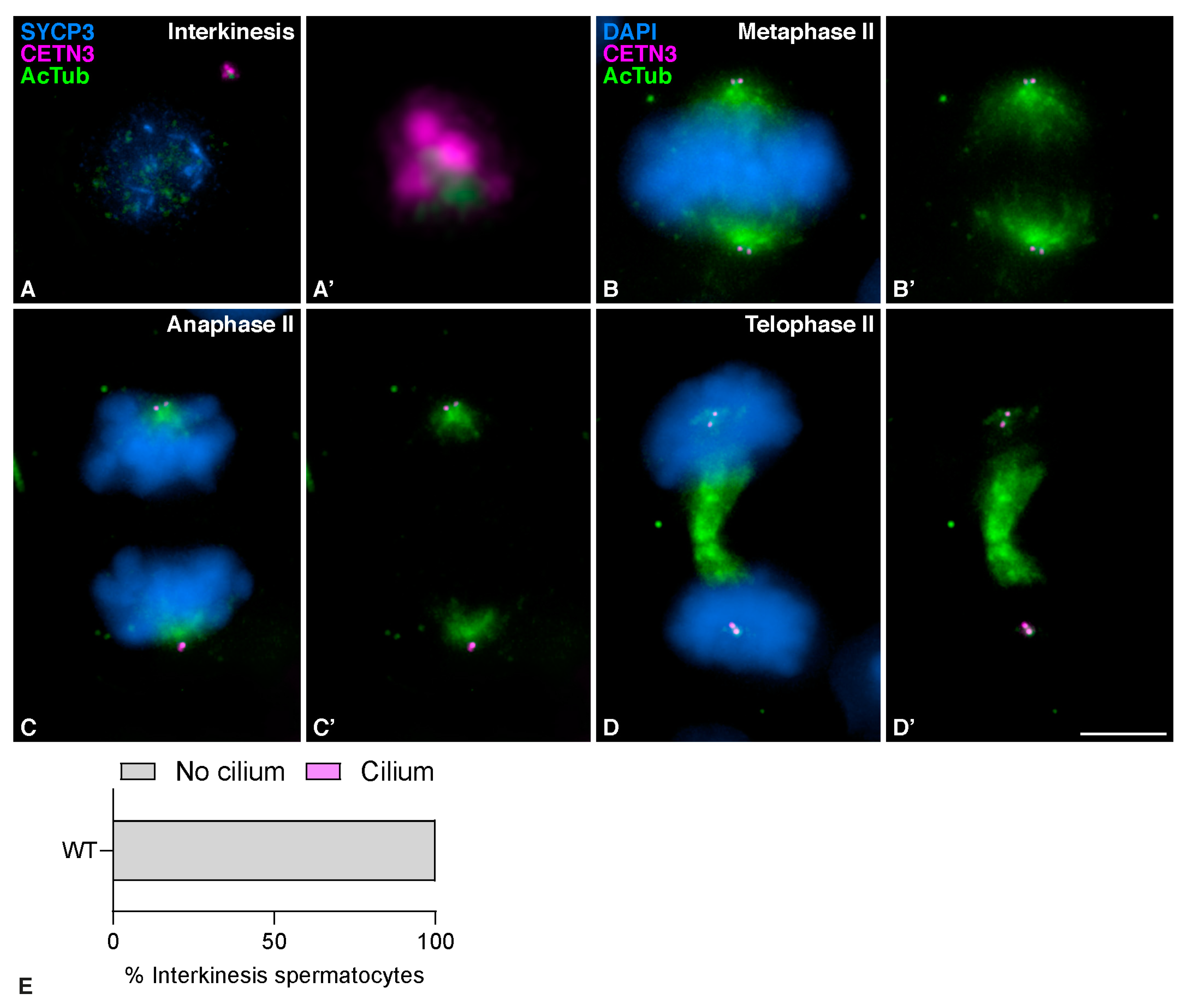
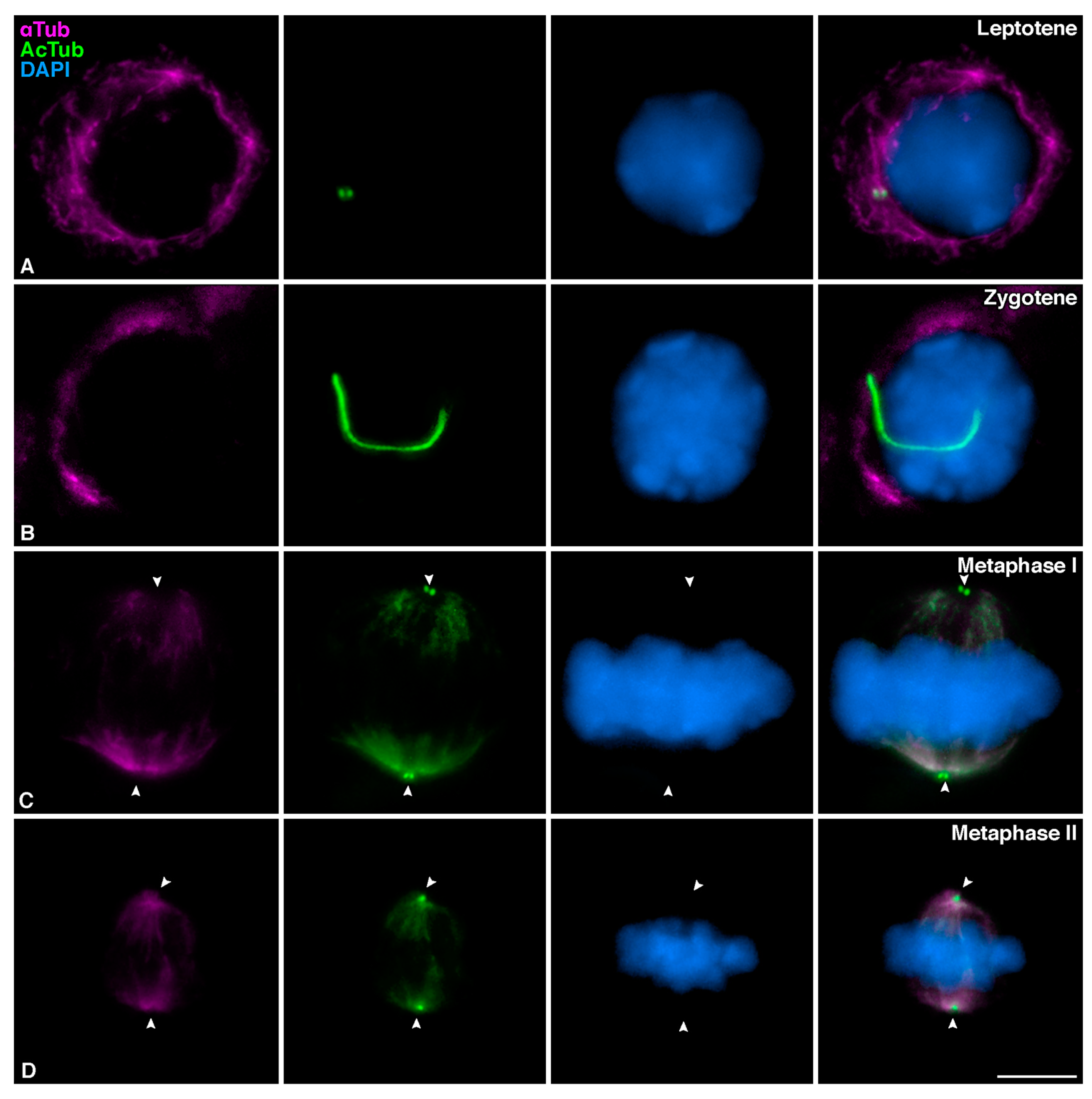
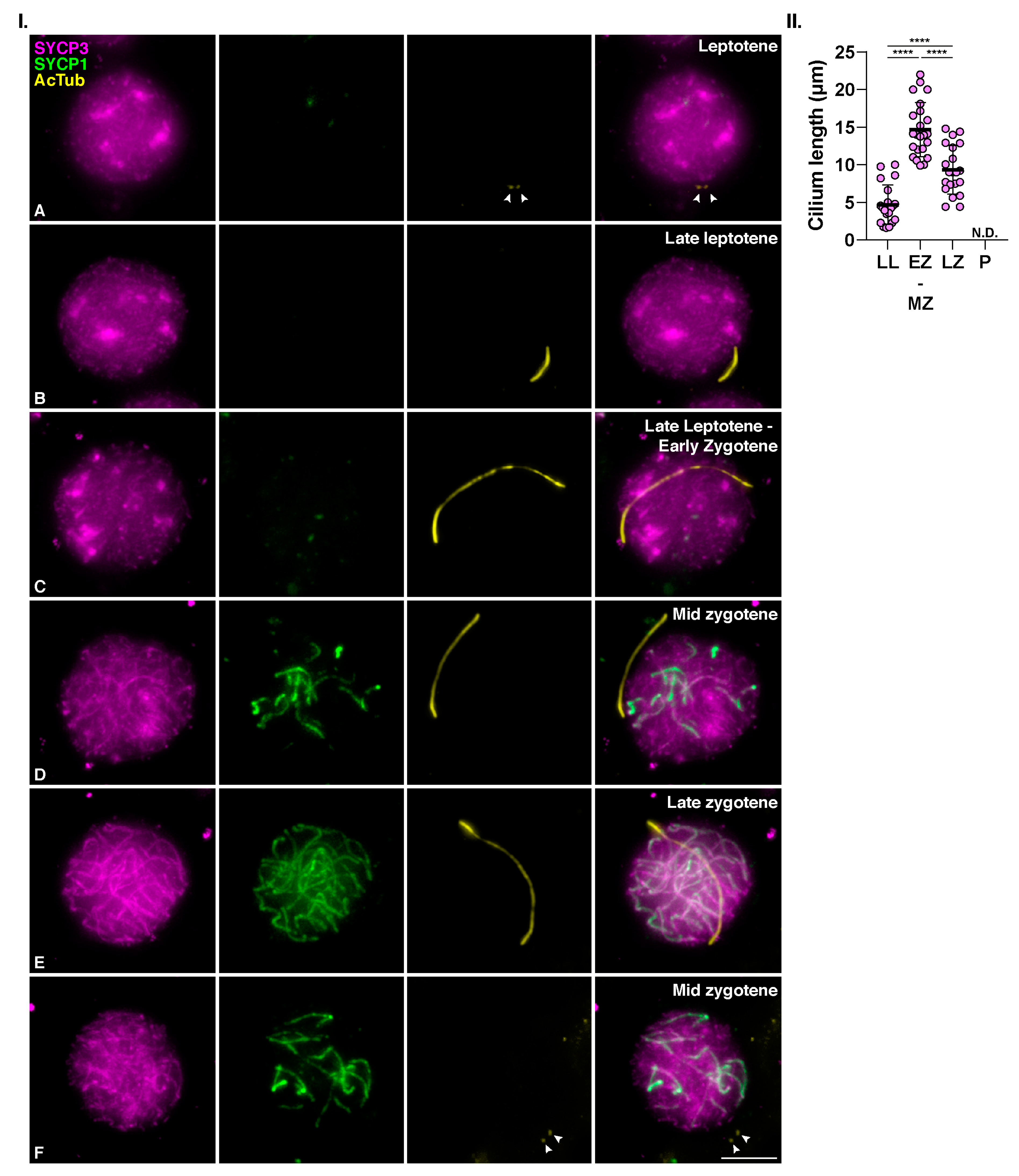
3.1. Mouse Meiotic Cilia Are Present in Zygotene spermatocytes prior to Centrosome Duplication
3.2. Meiotic Cilia Appear at the Onset of Synapsis
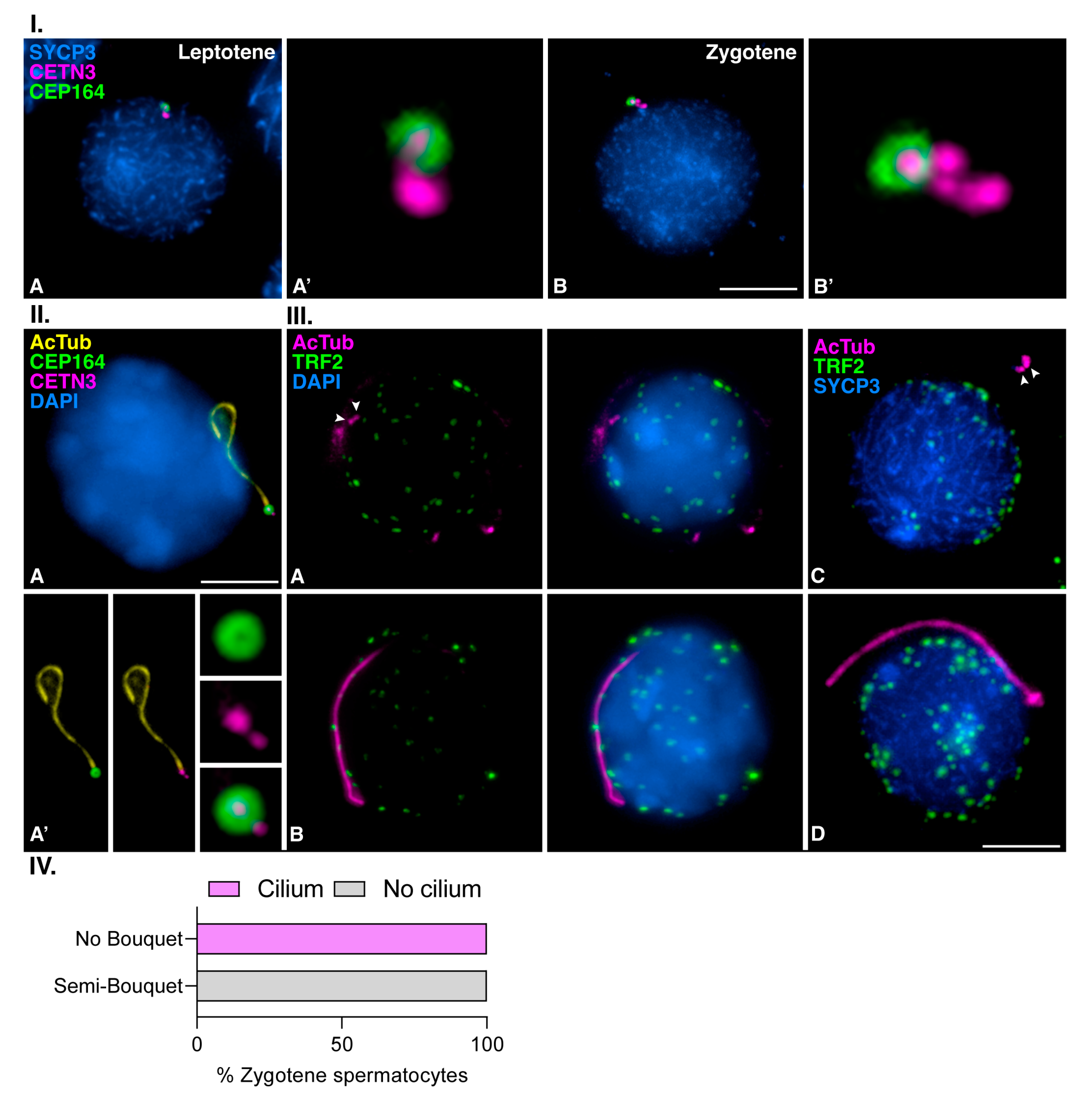
3.3. Mouse Meiotic Cilia Emanate from the Mother Centriole Prior to Centrosome Duplication
3.4. Meiotic Bouquet Conformation and Cilia Formation Are Not Concurrent Events
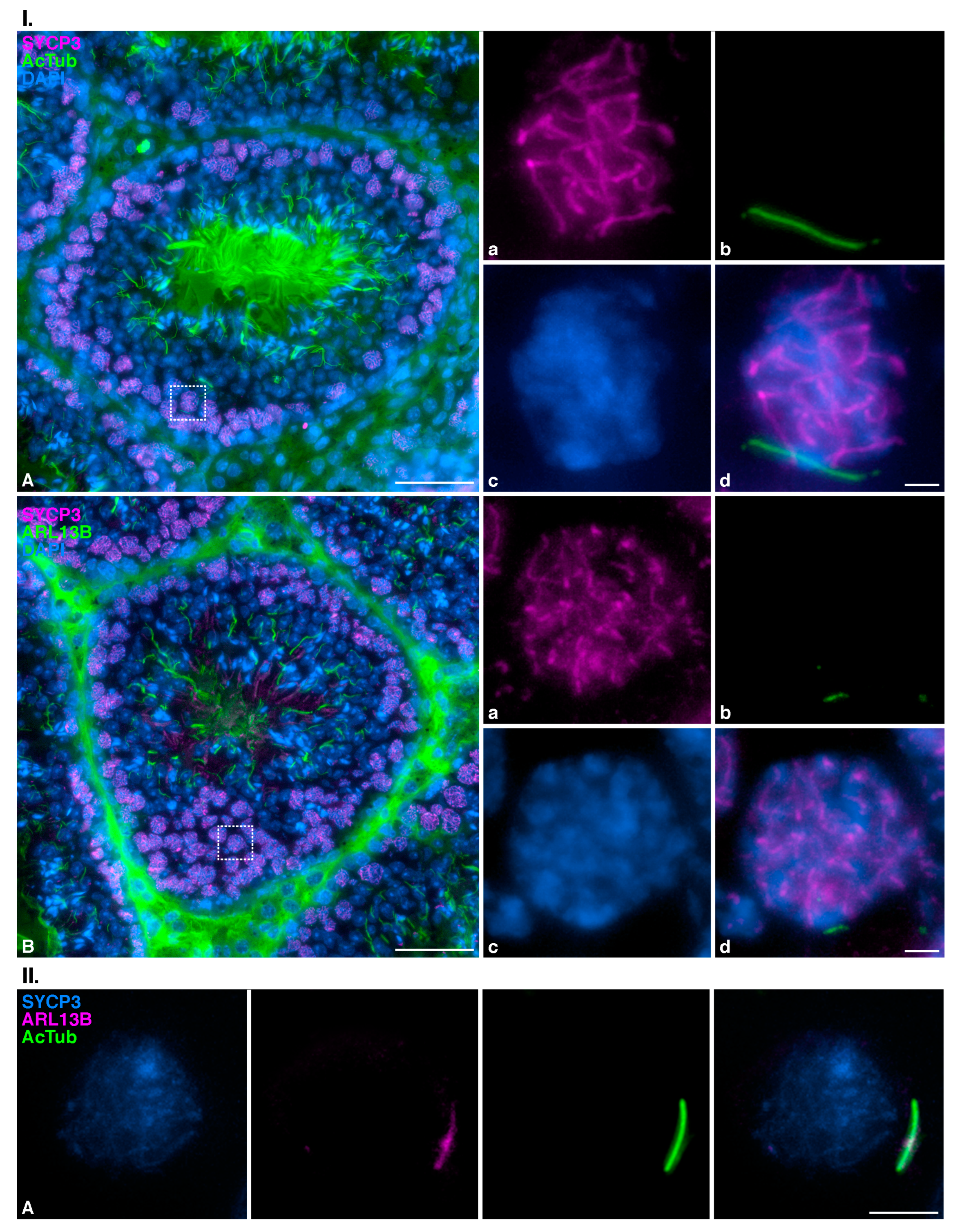
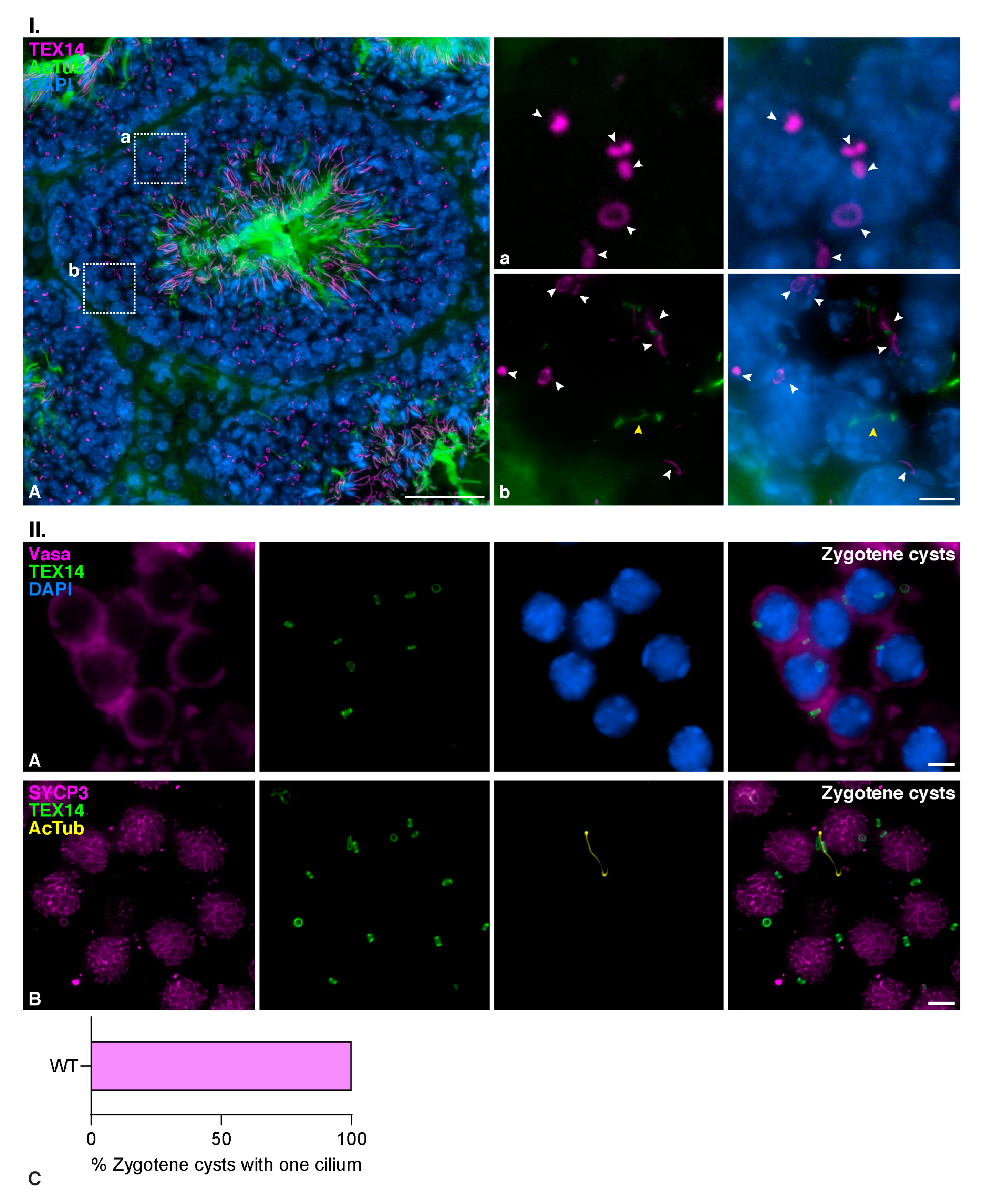
3.5. Zygotene TEX14 Connected Cysts
4. Discussion
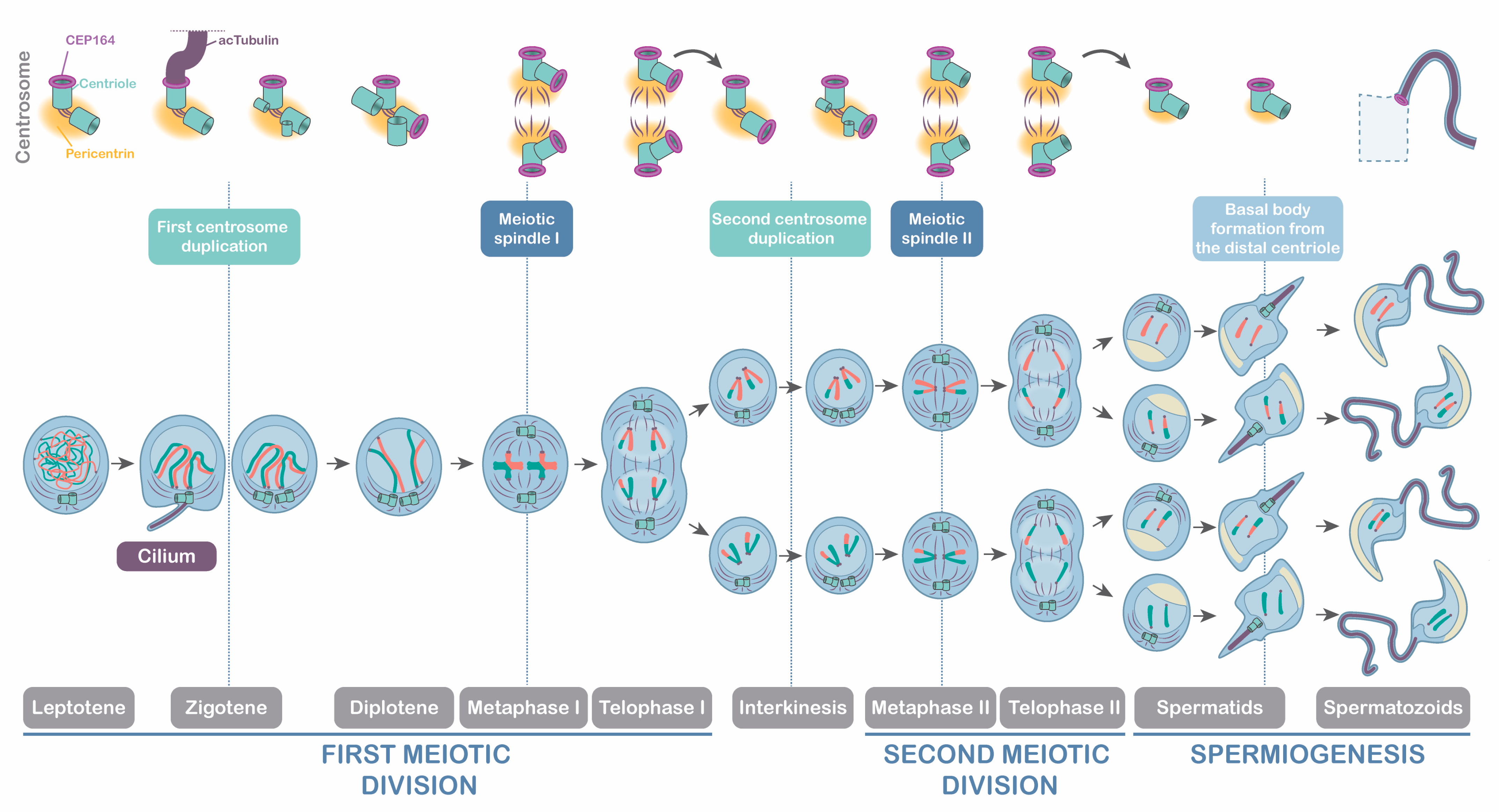
4.1. The Meiotic Cilium in Mouse Spermatogenesis
4.2. Meiotic Centrosomes and Their Relation to Ciliogenesis and Flagelogenesis
4.3. Tubulin Acetylation during Male Mouse Meiosis
4.4. Zygotene Cilia and Bouquet Formation Are Not Directly Related in Male Mouse Meiosis
4.5. Signaling Functions of the Cilia Could Be Spread to Cyst-Connected Spermatocytes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bornens, M. The centrosome in cells and organisms. Science 2012, 335, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Bornens, M. Centrosome organization and functions. Curr. Opin. Struct. Biol. 2021, 66, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A.; Stearns, T. The centrosome cycle: Centriole biogenesis, duplication and inherent asymmetries. Nat. Cell Biol. 2011, 13, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Lattao, R.; Kovács, L.; Glover, D.M. The Centrioles, Centrosomes, Basal Bodies, and Cilia of Drosophila melanogaster. Genetics 2017, 206, 33–53. [Google Scholar] [CrossRef]
- Schwarz, A.; Sankaralingam, P.; O’Connell, K.F.; Müller-Reichert, T. Revisiting Centrioles in Nematodes-Historic Findings and Current Topics. Cells 2018, 7, 101. [Google Scholar] [CrossRef]
- Azimzadeh, J.; Marshall, W.F. Building the centriole. Curr. Biol. 2010, 20, R816–R825. [Google Scholar] [CrossRef]
- Dumont, J.; Desai, A. Acentrosomal spindle assembly and chromosome segregation during oocyte meiosis. Trends Cell Biol. 2012, 22, 241–249. [Google Scholar] [CrossRef]
- Woodruff, J.B.; Wueseke, O.; Hyman, A.A. Pericentriolar material structure and dynamics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130459. [Google Scholar] [CrossRef]
- Conduit, P.T.; Wainman, A.; Raff, J.W. Centrosome function and assembly in animal cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 611–624. [Google Scholar] [CrossRef]
- Arbi, M.; Pefani, D.E.; Taraviras, S.; Lygerou, Z. Controlling centriole numbers: Geminin family members as master regulators of centriole amplification and multiciliogenesis. Chromosoma 2018, 127, 151–174. [Google Scholar] [CrossRef] [PubMed]
- Bloodgood, R.A. Sensory reception is an attribute of both primary cilia and motile cilia. J. Cell Sci. 2010, 123, 505–509. [Google Scholar] [CrossRef]
- Gadadhar, S.; Hirschmugl, T.; Janke, C. The tubulin code in mammalian sperm development and function. Semin. Cell Dev. Biol. 2022, 137, 26–37. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, F.R.; Reiter, J.F. Scoring a backstage pass: Mechanisms of ciliogenesis and ciliary access. J. Cell Biol. 2012, 197, 697–709. [Google Scholar] [CrossRef]
- Satir, P.; Christensen, S.T. Structure and function of mammalian cilia. Histochem. Cell Biol. 2008, 129, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Joukov, V.; De Nicolo, A. The Centrosome and the Primary Cilium: The Yin and Yang of a Hybrid Organelle. Cells 2019, 8, 701. [Google Scholar] [CrossRef] [PubMed]
- Eggenschwiler, J.T.; Anderson, K.V. Cilia and developmental signaling. Annu. Rev. Cell Dev. Biol. 2007, 23, 345–373. [Google Scholar] [CrossRef]
- McEwen, D.P.; Jenkins, P.M.; Martens, J.R. Olfactory cilia: Our direct neuronal connection to the external world. Curr. Top. Dev. Biol. 2008, 85, 333–370. [Google Scholar] [CrossRef]
- Wong, S.Y.; Seol, A.D.; So, P.L.; Ermilov, A.N.; Bichakjian, C.K.; Epstein, E.H., Jr.; Dlugosz, A.A.; Reiter, J.F. Primary cilia can both mediate and suppress Hedgehog pathway-dependent tumorigenesis. Nat. Med. 2009, 15, 1055–1061. [Google Scholar] [CrossRef]
- Sarkisian, M.R.; Semple-Rowland, S.L. Emerging Roles of Primary Cilia in Glioma. Front. Cell. Neurosci. 2019, 13, 55. [Google Scholar] [CrossRef]
- Braun, D.A.; Hildebrandt, F. Ciliopathies. Cold Spring Harb. Perspect. Biol. 2017, 9, a028191. [Google Scholar] [CrossRef] [PubMed]
- Eguether, T.; Hahne, M. Mixed signals from the cell’s antennae: Primary cilia in cancer. EMBO Rep. 2018, 19, e46589. [Google Scholar] [CrossRef]
- Wheatley, D.N.; Wang, A.M.; Strugnell, G.E. Expression of primary cilia in mammalian cells. Cell Biol. Int. 1996, 20, 73–81. [Google Scholar] [CrossRef]
- Graser, S.; Stierhof, Y.D.; Lavoie, S.B.; Gassner, O.S.; Lamla, S.; Le Clech, M.; Nigg, E.A. Cep164, a novel centriole appendage protein required for primary cilium formation. J. Cell Biol. 2007, 179, 321–330. [Google Scholar] [CrossRef]
- Vertii, A.; Hehnly, H.; Doxsey, S. The Centrosome, a Multitalented Renaissance Organelle. Cold Spring Harb. Perspect. Biol. 2016, 8, a025049. [Google Scholar] [CrossRef]
- Siller, S.S.; Sharma, H.; Li, S.; Yang, J.; Zhang, Y.; Holtzman, M.J.; Winuthayanon, W.; Colognato, H.; Holdener, B.C.; Li, F.Q.; et al. Conditional knockout mice for the distal appendage protein CEP164 reveal its essential roles in airway multiciliated cell differentiation. PLoS Genet. 2017, 13, e1007128. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.N.; Kuhns, S.; Neuner, A.; Hub, B.; Zentgraf, H.; Pereira, G. Cep164 mediates vesicular docking to the mother centriole during early steps of ciliogenesis. J. Cell Biol. 2012, 199, 1083–1101. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Dynlacht, B.D. Regulating the transition from centriole to basal body. J. Cell Biol. 2011, 193, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dynlacht, B.D. The regulation of cilium assembly and disassembly in development and disease. Development 2018, 145, dev151407. [Google Scholar] [CrossRef]
- Griswold, M.D. Spermatogenesis: The Commitment to Meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef]
- Handel, M.A.; Schimenti, J.C. Genetics of mammalian meiosis: Regulation, dynamics and impact on fertility. Nat. Rev. Genet. 2010, 11, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Bolcun-Filas, E.; Handel, M.A. Meiosis: The chromosomal foundation of reproduction. Biol. Reprod. 2018, 99, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, E.; Lopez-Jimenez, P.; Gonzalez-Martinez, J.; Malumbres, M.; Suja, J.A.; Gomez, R. PLK1 regulates centrosome migration and spindle dynamics in male mouse meiosis. EMBO Rep. 2021, 22, e51030. [Google Scholar] [CrossRef]
- Kimmins, S.; Kotaja, N.; Davidson, I.; Sassone-Corsi, P. Testis-specific transcription mechanisms promoting male germ-cell differentiation. Reproduction 2004, 128, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Tapia Contreras, C.; Hoyer-Fender, S. The Transformation of the Centrosome into the Basal Body: Similarities and Dissimilarities between Somatic and Male Germ Cells and Their Relevance for Male Fertility. Cells 2021, 10, 2266. [Google Scholar] [CrossRef]
- Avidor-Reiss, T.; Carr, A.; Fishman, E.L. The sperm centrioles. Mol. Cell Endocrinol. 2020, 518, 110987. [Google Scholar] [CrossRef] [PubMed]
- Sironen, A.; Shoemark, A.; Patel, M.; Loebinger, M.R.; Mitchison, H.M. Sperm defects in primary ciliary dyskinesia and related causes of male infertility. Cell Mol. Life Sci. 2020, 77, 2029–2048. [Google Scholar] [CrossRef]
- Girardet, L.; Augiere, C.; Asselin, M.P.; Belleannee, C. Primary cilia: Biosensors of the male reproductive tract. Andrology 2019, 7, 588–602. [Google Scholar] [CrossRef]
- Mytlis, A.; Kumar, V.; Qiu, T.; Deis, R.; Hart, N.; Levy, K.; Masek, M.; Shawahny, A.; Ahmad, A.; Eitan, H.; et al. Control of meiotic chromosomal bouquet and germ cell morphogenesis by the zygotene cilium. Science 2022, 376, eabh3104. [Google Scholar] [CrossRef]
- Xie, H.; Wang, X.; Jin, M.; Li, L.; Zhu, J.; Kang, Y.; Chen, Z.; Sun, Y.; Zha, C. Cilia regulate meiotic recombination in zebrafish. J. Mol. Cell Biol. 2022. [Google Scholar] [CrossRef]
- Fisher, S.; Kuna, D.; Caspary, T.; Kahn, R.A.; Sztul, E. ARF family GTPases with links to cilia. Am. J. Physiol. Cell Physiol. 2020, 319, C404–C418. [Google Scholar] [CrossRef] [PubMed]
- Caspary, T.; Larkins, C.E.; Anderson, K.V. The graded response to Sonic Hedgehog depends on cilia architecture. Dev. Cell 2007, 12, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Hor, C.H.; Goh, E.L. Small GTPases in hedgehog signalling: Emerging insights into the disease mechanisms of Rab23-mediated and Arl13b-mediated ciliopathies. Curr. Opin. Genet. Dev. 2019, 56, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Papanikos, F.; Clement, J.A.J.; Testa, E.; Ravindranathan, R.; Grey, C.; Dereli, I.; Bondarieva, A.; Valerio-Cabrera, S.; Stanzione, M.; Schleiffer, A.; et al. Mouse ANKRD31 Regulates Spatiotemporal Patterning of Meiotic Recombination Initiation and Ensures Recombination between X and Y Sex Chromosomes. Mol. Cell 2019, 74, 1069–1085. [Google Scholar] [CrossRef] [PubMed]
- Fraune, J.; Schramm, S.; Alsheimer, M.; Benavente, R. The mammalian synaptonemal complex: Protein components, assembly and role in meiotic recombination. Exp. Cell Res. 2012, 318, 1340–1346. [Google Scholar] [CrossRef]
- Sillibourne, J.E.; Specht, C.G.; Izeddin, I.; Hurbain, I.; Tran, P.; Triller, A.; Darzacq, X.; Dahan, M.; Bornens, M. Assessing the localization of centrosomal proteins by PALM/STORM nanoscopy. Cytoskeleton 2011, 68, 619–627. [Google Scholar] [CrossRef]
- Wellard, S.R.; Zhang, Y.; Shults, C.; Zhao, X.; McKay, M.; Murray, S.A.; Jordan, P.W. Overlapping roles for PLK1 and Aurora A during meiotic centrosome biogenesis in mouse spermatocytes. EMBO Rep. 2021, 22, e51023. [Google Scholar] [CrossRef]
- Hoque, M.; Chen, D.; Hess, R.A.; Li, F.Q.; Takemaru, K.I. CEP164 is essential for efferent duct multiciliogenesis and male fertility. Reproduction 2021, 162, 129–139. [Google Scholar] [CrossRef]
- Liebe, B.; Petukhova, G.; Barchi, M.; Bellani, M.; Braselmann, H.; Nakano, T.; Pandita, T.K.; Jasin, M.; Fornace, A.; Meistrich, M.L.; et al. Mutations that affect meiosis in male mice influence the dynamics of the mid-preleptotene and bouquet stages. Exp. Cell Res. 2006, 312, 3768–3781. [Google Scholar] [CrossRef]
- Scherthan, H.; Sfeir, A.; de Lange, T. Rap1-independent telomere attachment and bouquet formation in mammalian meiosis. Chromosoma 2011, 120, 151–157. [Google Scholar] [CrossRef]
- Scherthan, H.; Weich, S.; Schwegler, H.; Heyting, C.; Harle, M.; Cremer, T. Centromere and telomere movements during early meiotic prophase of mouse and man are associated with the onset of chromosome pairing. J. Cell Biol. 1996, 134, 1109–1125. [Google Scholar] [CrossRef]
- Greenbaum, M.P.; Iwamori, T.; Buchold, G.M.; Matzuk, M.M. Germ cell intercellular bridges. Cold Spring Harb. Perspect. Biol. 2011, 3, a005850. [Google Scholar] [CrossRef]
- Dym, M.; Fawcett, D.W. Further observations on the numbers of spermatogonia, spermatocytes, and spermatids connected by intercellular bridges in the mammalian testis. Biol. Reprod. 1971, 4, 195–215. [Google Scholar] [CrossRef]
- Greenbaum, M.P.; Yan, W.; Wu, M.H.; Lin, Y.N.; Agno, J.E.; Sharma, M.; Braun, R.E.; Rajkovic, A.; Matzuk, M.M. TEX14 is essential for intercellular bridges and fertility in male mice. Proc. Natl. Acad. Sci. USA 2006, 103, 4982–4987. [Google Scholar] [CrossRef] [PubMed]
- Azizi, H.; Ranjbar, M.; Rahaiee, S.; Govahi, M.; Skutella, T. Investigation of VASA Gene and Protein Expression in Neonate and Adult Testicular Germ Cells in Mice In Vivo and In Vitro. Cell J. 2020, 22, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.; Talib, S.Z.A.; Kaldis, P. Diverse roles for CDK-associated activity during spermatogenesis. FEBS Lett. 2019, 593, 2925–2949. [Google Scholar] [CrossRef]
- Riparbelli, M.G.; Callaini, G.; Megraw, T.L. Assembly and persistence of primary cilia in dividing Drosophila spermatocytes. Dev. Cell 2012, 23, 425–432. [Google Scholar] [CrossRef]
- Alieva, I.B.; Vorobjev, I.A. Vertebrate primary cilia: A sensory part of centrosomal complex in tissue cells, but a “sleeping beauty” in cultured cells? Cell Biol. Int. 2004, 28, 139–150. [Google Scholar] [CrossRef]
- Zhao, H.; Khan, Z.; Westlake, C.J. Ciliogenesis membrane dynamics and organization. Semin. Cell Dev. Biol. 2022, 133, 20–31. [Google Scholar] [CrossRef]
- Zitouni, S.; Nabais, C.; Jana, S.C.; Guerrero, A.; Bettencourt-Dias, M. Polo-like kinases: Structural variations lead to multiple functions. Nat. Rev. Mol. Cell Biol. 2014, 15, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Piperno, G.; Fuller, M.T. Monoclonal antibodies specific for an acetylated form of alpha-tubulin recognize the antigen in cilia and flagella from a variety of organisms. J. Cell Biol. 1985, 101, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Wloga, D.; Joachimiak, E.; Louka, P.; Gaertig, J. Posttranslational Modifications of Tubulin and Cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028159. [Google Scholar] [CrossRef] [PubMed]
- Janke, C.; Magiera, M.M. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, S. Centrioles and the formation of rudimentary cilia by fibroblasts and smooth muscle cells. J. Cell Biol. 1962, 15, 363–377. [Google Scholar] [CrossRef]
- Breslow, D.K.; Holland, A.J. Mechanism and Regulation of Centriole and Cilium Biogenesis. Annu. Rev. Biochem. 2019, 88, 691–724. [Google Scholar] [CrossRef]
- Garcia, G., 3rd; Reiter, J.F. A primer on the mouse basal body. Cilia 2016, 5, 17. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Schrøder, J.M.; Satir, P.; Christensen, S.T. The ciliary cytoskeleton. Compr. Physiol. 2012, 2, 779–803. [Google Scholar] [CrossRef]
- Kochanski, R.S.; Borisy, G.G. Mode of centriole duplication and distribution. J. Cell Biol. 1990, 110, 1599–1605. [Google Scholar] [CrossRef]
- Edde, B.; Rossier, J.; Le Caer, J.P.; Desbruyeres, E.; Gros, F.; Denoulet, P. Posttranslational glutamylation of alpha-tubulin. Science 1990, 247, 83–85. [Google Scholar] [CrossRef]
- Rasamizafy, S.F.; Delsert, C.; Rabeharivelo, G.; Cau, J.; Morin, N.; van Dijk, J. Mitotic Acetylation of Microtubules Promotes Centrosomal PLK1 Recruitment and Is Required to Maintain Bipolar Spindle Homeostasis. Cells 2021, 10, 1859. [Google Scholar] [CrossRef] [PubMed]
- Piperno, G.; LeDizet, M.; Chang, X.J. Microtubules containing acetylated alpha-tubulin in mammalian cells in culture. J. Cell Biol. 1987, 104, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Reed, N.A.; Cai, D.; Blasius, T.L.; Jih, G.T.; Meyhofer, E.; Gaertig, J.; Verhey, K.J. Microtubule acetylation promotes kinesin-1 binding and transport. Curr. Biol. 2006, 16, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Ikeda, M.; Chiba, S.; Kanno, S.; Mizuno, K. Furry promotes acetylation of microtubules in the mitotic spindle by inhibition of SIRT2 tubulin deacetylase. J. Cell Sci. 2013, 126, 4369–4380. [Google Scholar] [CrossRef]
- Akera, T. Tubulin post-translational modifications in meiosis. Semin. Cell Dev. Biol. 2021, 137, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, S.; Cui, Z.; Dai, X.; Zhang, M.; Miao, Y.; Zhou, C.; Ou, X.; Xiong, B. The cohesion establishment factor Esco1 acetylates alpha-tubulin to ensure proper spindle assembly in oocyte meiosis. Nucleic Acids Res. 2018, 46, 2335–2346. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Gao, M.; Han, L.; Qiu, D.; Wang, H.; Xiong, B.; Sun, S.C.; Liu, H.; Gu, L. HDAC3 promotes meiotic apparatus assembly in mouse oocytes by modulating tubulin acetylation. Development 2017, 144, 3789–3797. [Google Scholar] [CrossRef]
- Parra, M.T.; Viera, A.; Gomez, R.; Page, J.; Benavente, R.; Santos, J.L.; Rufas, J.S.; Suja, J.A. Involvement of the cohesin Rad21 and SCP3 in monopolar attachment of sister kinetochores during mouse meiosis I. J. Cell Sci. 2004, 117, 1221–1234. [Google Scholar] [CrossRef]
- Siegfried, K.R. It starts at the ends: The zebrafish meiotic bouquet is where it all begins. PLoS Genet. 2019, 15, e1007854. [Google Scholar] [CrossRef]
- Link, J.; Jantsch, V. Meiotic chromosomes in motion: A perspective from Mus musculus and Caenorhabditis elegans. Chromosoma 2019, 128, 317–330. [Google Scholar] [CrossRef]
- Page, J.; Berrios, S.; Rufas, J.S.; Parra, M.T.; Suja, J.A.; Heyting, C.; Fernandez-Donoso, R. The pairing of X and Y chromosomes during meiotic prophase in the marsupial species Thylamys elegans is maintained by a dense plate developed from their axial elements. J. Cell Sci. 2003, 116, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, E.; Criollo, A.; Schiattarella, G.G.; Altamirano, F.; French, K.M.; May, H.I.; Jiang, N.; Nguyen, N.U.N.; Romero, D.; Roa, J.C.; et al. Fibroblast Primary Cilia Are Required for Cardiac Fibrosis. Circulation 2019, 139, 2342–2357. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, V.; Weisz, O.A. Discerning the role of mechanosensors in regulating proximal tubule function. Am. J. Physiol. Ren. Physiol. 2016, 310, F1–F5. [Google Scholar] [CrossRef] [PubMed]
- Broekhuis, J.R.; Leong, W.Y.; Jansen, G. Regulation of cilium length and intraflagellar transport. Int. Rev. Cell Mol. Biol. 2013, 303, 101–138. [Google Scholar] [CrossRef]
- Challis, R.C.; Tian, H.; Yin, W.; Ma, M. Genetic Ablation of Type III Adenylyl Cyclase Exerts Region-Specific Effects on Cilia Architecture in the Mouse Nose. PLoS One 2016, 11, e0150638. [Google Scholar] [CrossRef]
- Zemirli, N.; Boukhalfa, A.; Dupont, N.; Botti, J.; Codogno, P.; Morel, E. The primary cilium protein folliculin is part of the autophagy signaling pathway to regulate epithelial cell size in response to fluid flow. Cell Stress 2019, 3, 100–109. [Google Scholar] [CrossRef]
- Spasic, M.; Jacobs, C.R. Lengthening primary cilia enhances cellular mechanosensitivity. Eur. Cell Mater. 2017, 33, 158–168. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Ito, S.; Slautterback, D. The occurrence of intercellular bridges in groups of cells exhibiting synchronous differentiation. J. Biophys. Biochem. Cytol. 1959, 5, 453–460. [Google Scholar] [CrossRef]
- Greenbaum, M.P.; Iwamori, N.; Agno, J.E.; Matzuk, M.M. Mouse TEX14 is required for embryonic germ cell intercellular bridges but not female fertility. Biol. Reprod. 2009, 80, 449–457. [Google Scholar] [CrossRef]
- Lei, L.; Spradling, A.C. Mouse primordial germ cells produce cysts that partially fragment prior to meiosis. Development 2013, 140, 2075–2081. [Google Scholar] [CrossRef]
- Moss, S.B.; VanScoy, H.; Gerton, G.L. Mapping of a haploid transcribed and translated sperm-specific gene to the mouse X chromosome. Mamm. Genome 1997, 8, 37–38. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Mruk, D.D.; Cheng, Y.H.; Tang, E.I.; Han, D.; Lee, W.M.; Wong, E.W.; Cheng, C.Y. Actin binding proteins, spermatid transport and spermiation. Semin. Cell Dev. Biol. 2014, 30, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Mruk, D.D.; Cheng, C.Y. Regulation of actin dynamics and protein trafficking during spermatogenesis--insights into a complex process. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 153–172. [Google Scholar] [CrossRef]
- Linck, R.W.; Chemes, H.; Albertini, D.F. The axoneme: The propulsive engine of spermatozoa and cilia and associated ciliopathies leading to infertility. J. Assist. Reprod. Genet. 2016, 33, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Page, J.; Suja, J.A.; Santos, J.L.; Rufas, J.S. Squash procedure for protein immunolocalization in meiotic cells. Chromosome Res. 1998, 6, 639–642. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Jiménez, P.; Pérez-Martín, S.; Hidalgo, I.; García-Gonzalo, F.R.; Page, J.; Gómez, R. The Male Mouse Meiotic Cilium Emanates from the Mother Centriole at Zygotene Prior to Centrosome Duplication. Cells 2023, 12, 142. https://doi.org/10.3390/cells12010142
López-Jiménez P, Pérez-Martín S, Hidalgo I, García-Gonzalo FR, Page J, Gómez R. The Male Mouse Meiotic Cilium Emanates from the Mother Centriole at Zygotene Prior to Centrosome Duplication. Cells. 2023; 12(1):142. https://doi.org/10.3390/cells12010142
Chicago/Turabian StyleLópez-Jiménez, Pablo, Sara Pérez-Martín, Inés Hidalgo, Francesc R. García-Gonzalo, Jesús Page, and Rocio Gómez. 2023. "The Male Mouse Meiotic Cilium Emanates from the Mother Centriole at Zygotene Prior to Centrosome Duplication" Cells 12, no. 1: 142. https://doi.org/10.3390/cells12010142