
Pre-Synaptic GABAA in NaV1.8+ Primary Afferents Is Required for the Development of Punctate but Not Dynamic Mechanical Allodynia following CFA Inflammation

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animals
2.3. Genotyping
2.4. Anaesthesia and Housing
2.5. Characterization of Sensory and Motor Function in Transgenic Mice
2.6. Assessment of Sensitivity to Light Touch
2.7. Hot/Cold Plate Assessment of Sensitivity to Noxious Hot and Cold
2.8. Assessment of Sensorimotor Function
2.9. Induction of CFA Inflammation
2.10. Assessment of Mechanical and Thermal Sensitivity after CFA
2.11. Mechanical Stimulus Protocol following CFA Inflammation
2.12. Preparation of Mouse Spinal Cord
2.13. Dual-Labeling of c-Fos Protein and c-fos mRNA
2.14. Imaging
2.15. Cell Quantification and Image Analysis
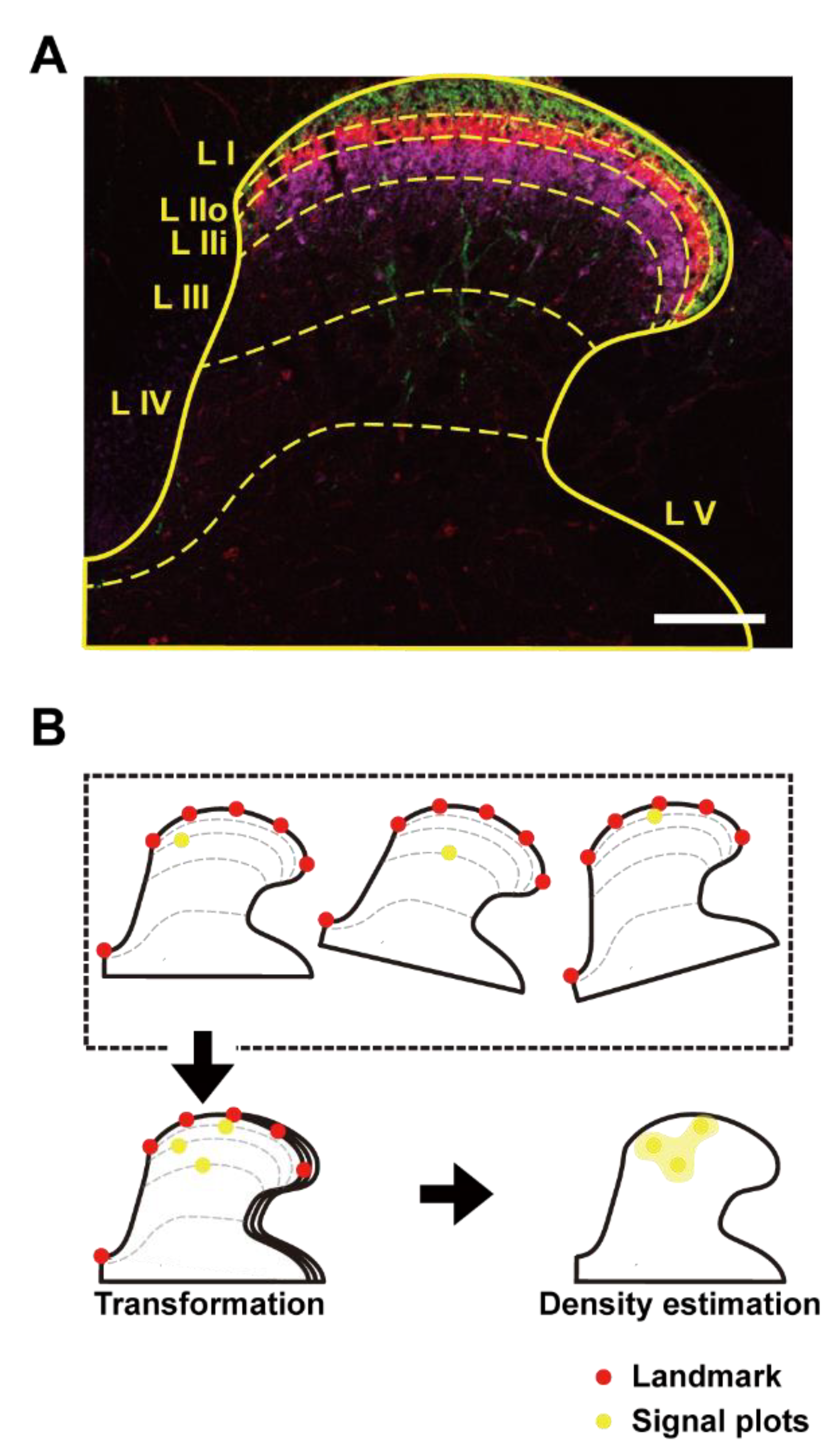
2.16. Processing of Dorsal Horn Images and Spatial Density Estimation
2.17. Mouse Sural Nerve Preparation
2.18. Compound Action Potential Recordings
2.19. Assessment of Electrical Excitability in A-Fibres
2.20. Assessment of Electrical Excitability in C-Fibres
2.21. Induction and Assessment of Activity Depending Slowing
2.22. Chemicals
2.23. Data Analysis and Statistics
3. Results
3.1. GABAA-β3 Deletion in NaV1.8+ Afferents Did Not Affect Voluntary Motor Behavior
3.2. GABAA-β3 Deletion in NaV1.8+ Afferents Increased Basal Sensitivity to Heat
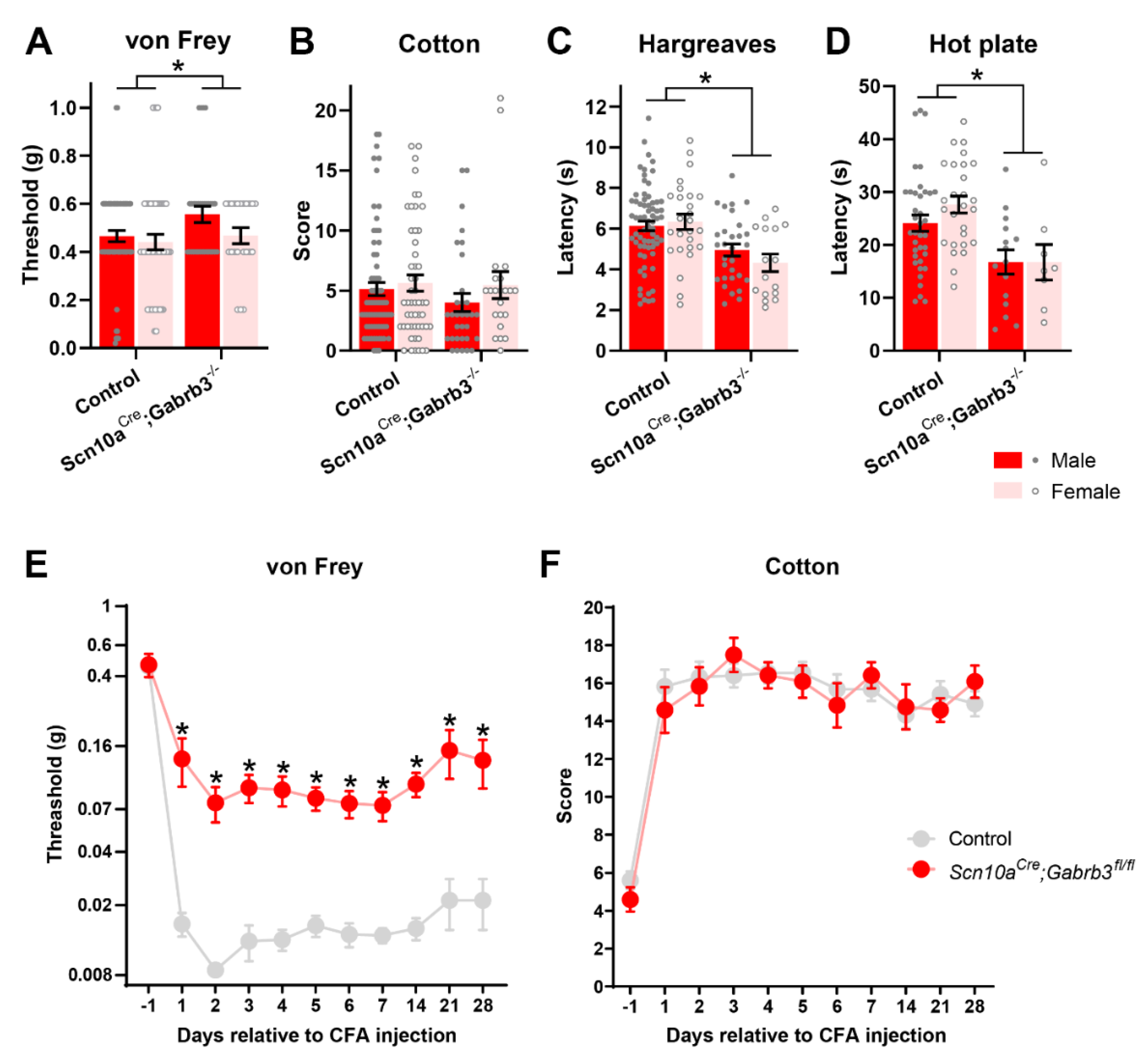
3.3. GABAA-β3 Deletion in NaV1.8+ Afferents Blunted Punctate Mechanical Allodynia after CFA
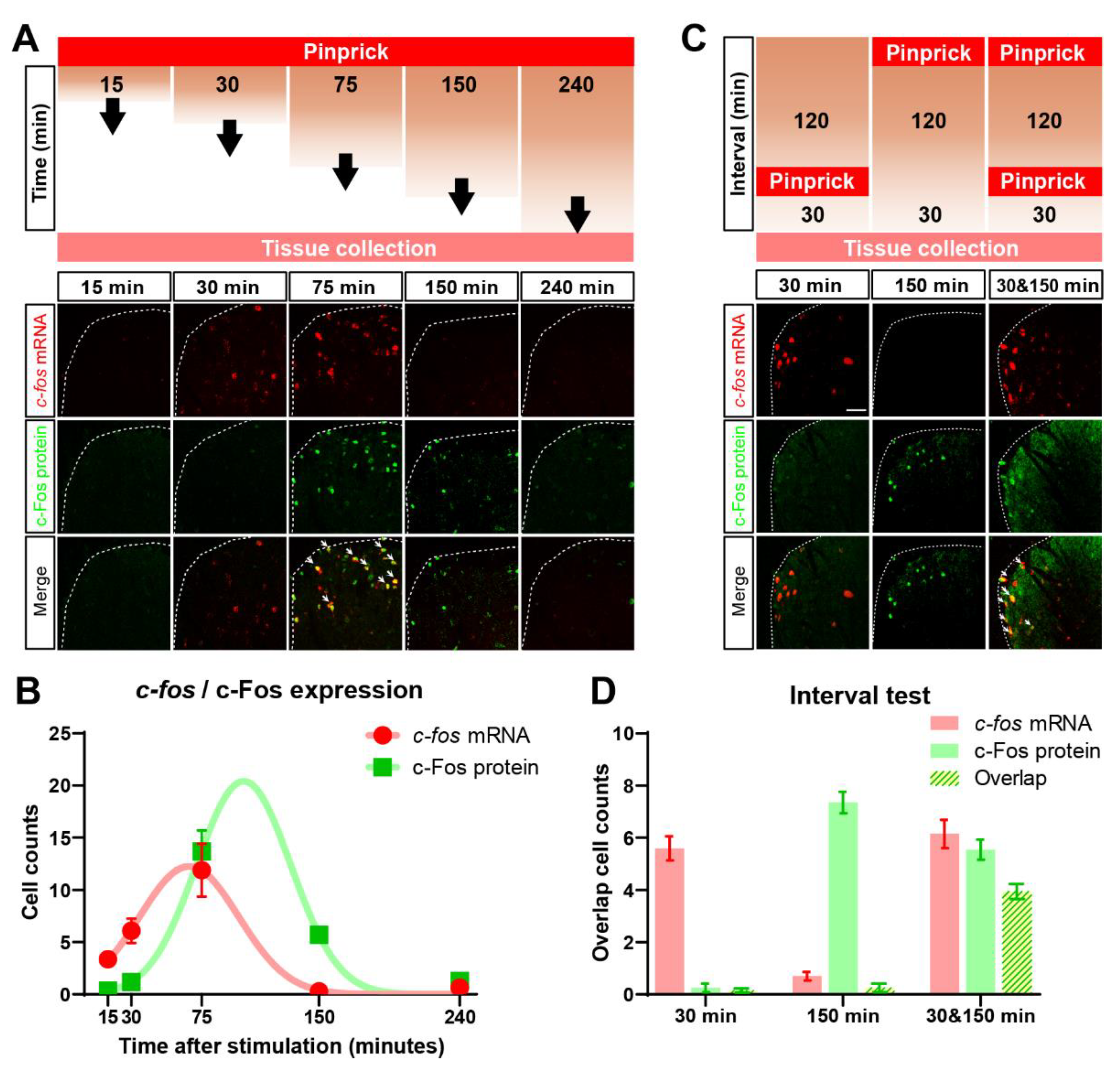
3.4. Identification of Cells in the Dorsal Horn Activated by Time Separated Punctate and Dynamic Mechanical Stimuli
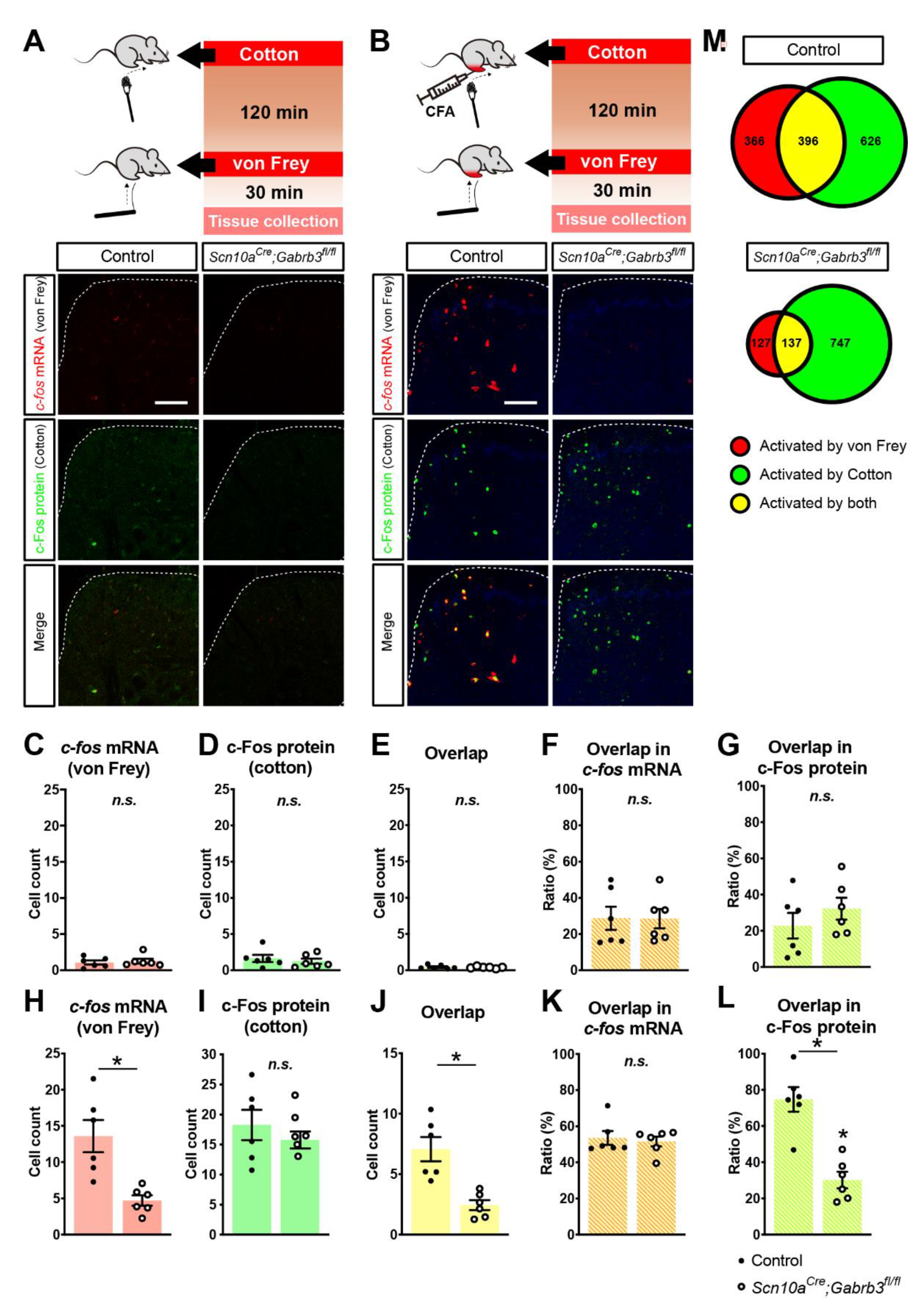
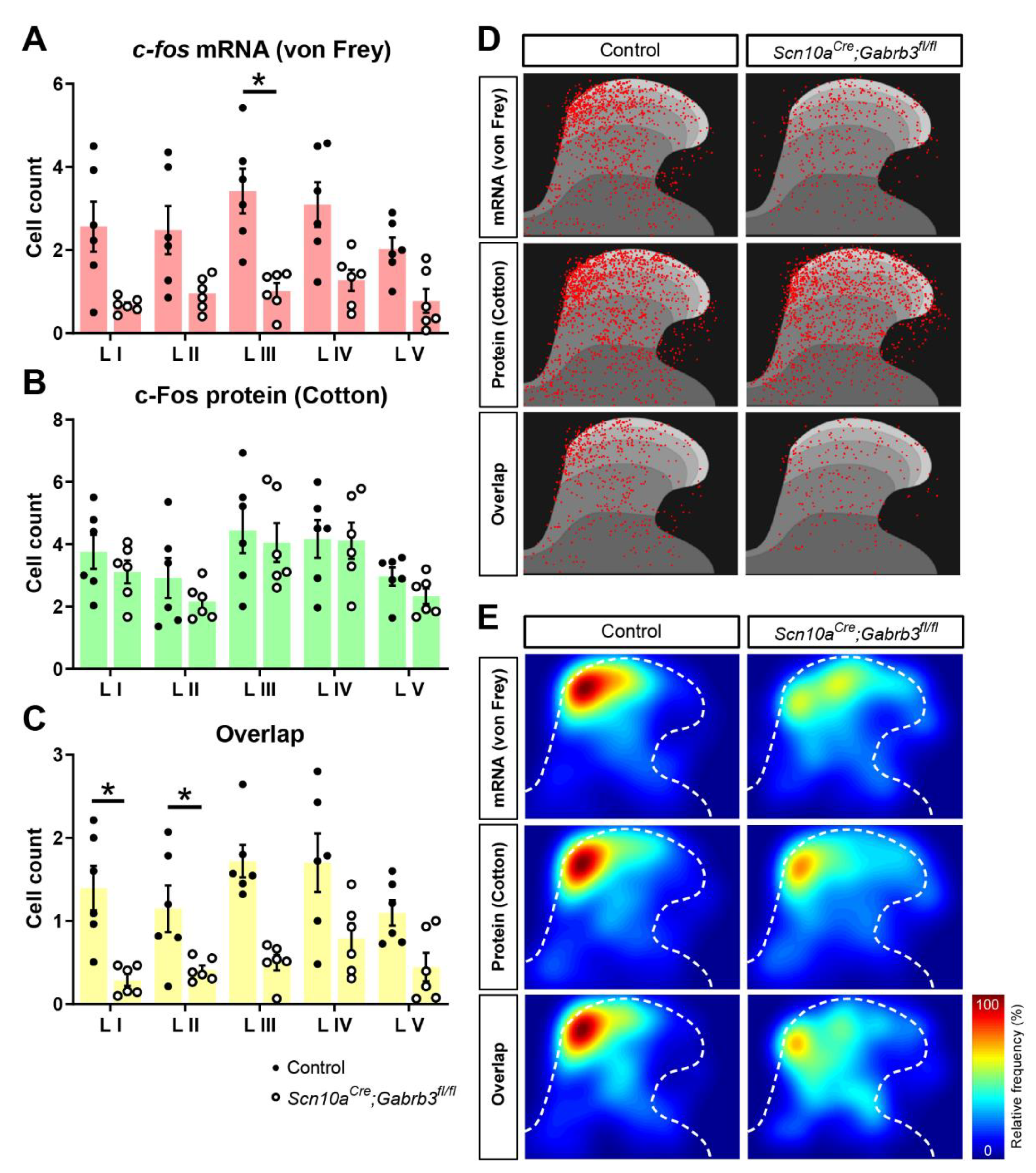
3.5. Fewer c-fos-Labelled Cells in Spinal Dorsal Horn Responding to Punctate Mechanical Stimuli after CFA Injury in Mice Lacking GABAA-β3 in NaV1.8+ Afferents
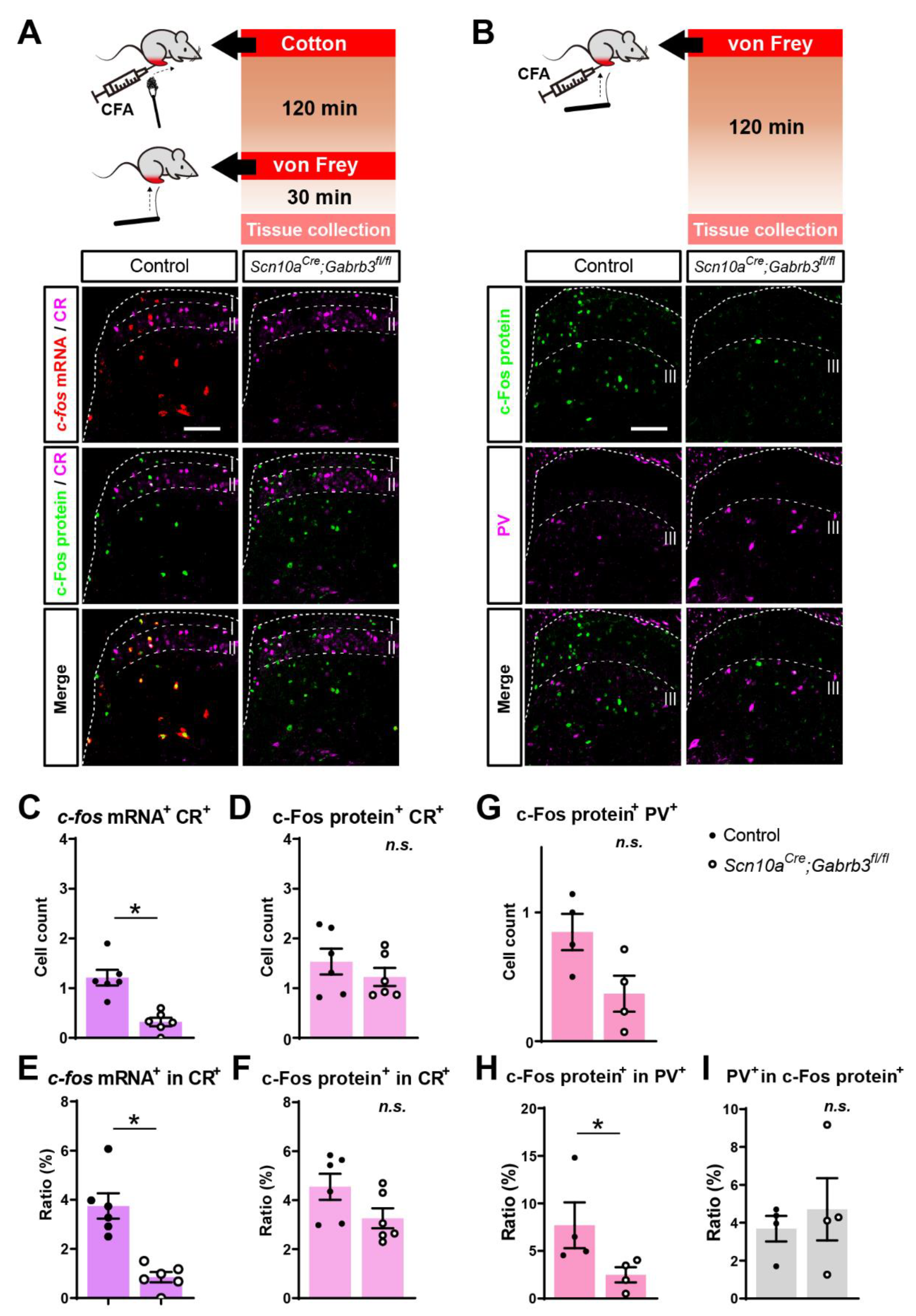
3.6. Fewer Calretinin-Positive and Fewer Parvalbumin-Positive Cells Responded to Punctate but Not Dynamic Mechanical Stimuli after CFA Injury in Mice Lacking NaV1.8+ Pre-Synaptic GABAA
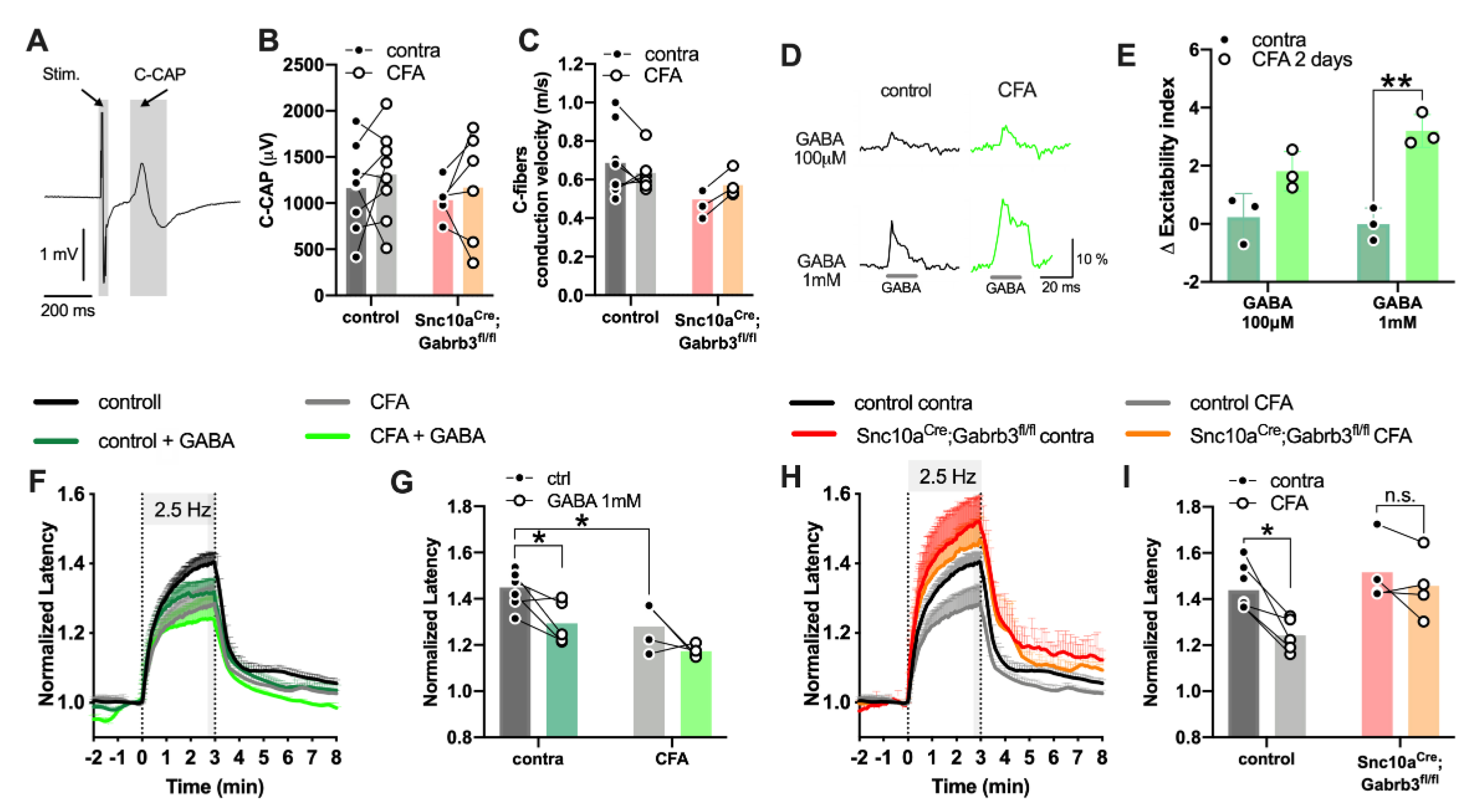
3.7. Loss of Axonal GABAA in NaV1.8+-Afferents Did Not Affect A-Fibre Excitability but Reduced the Capacity of C-Fibres to Sustain Action Potential Firing
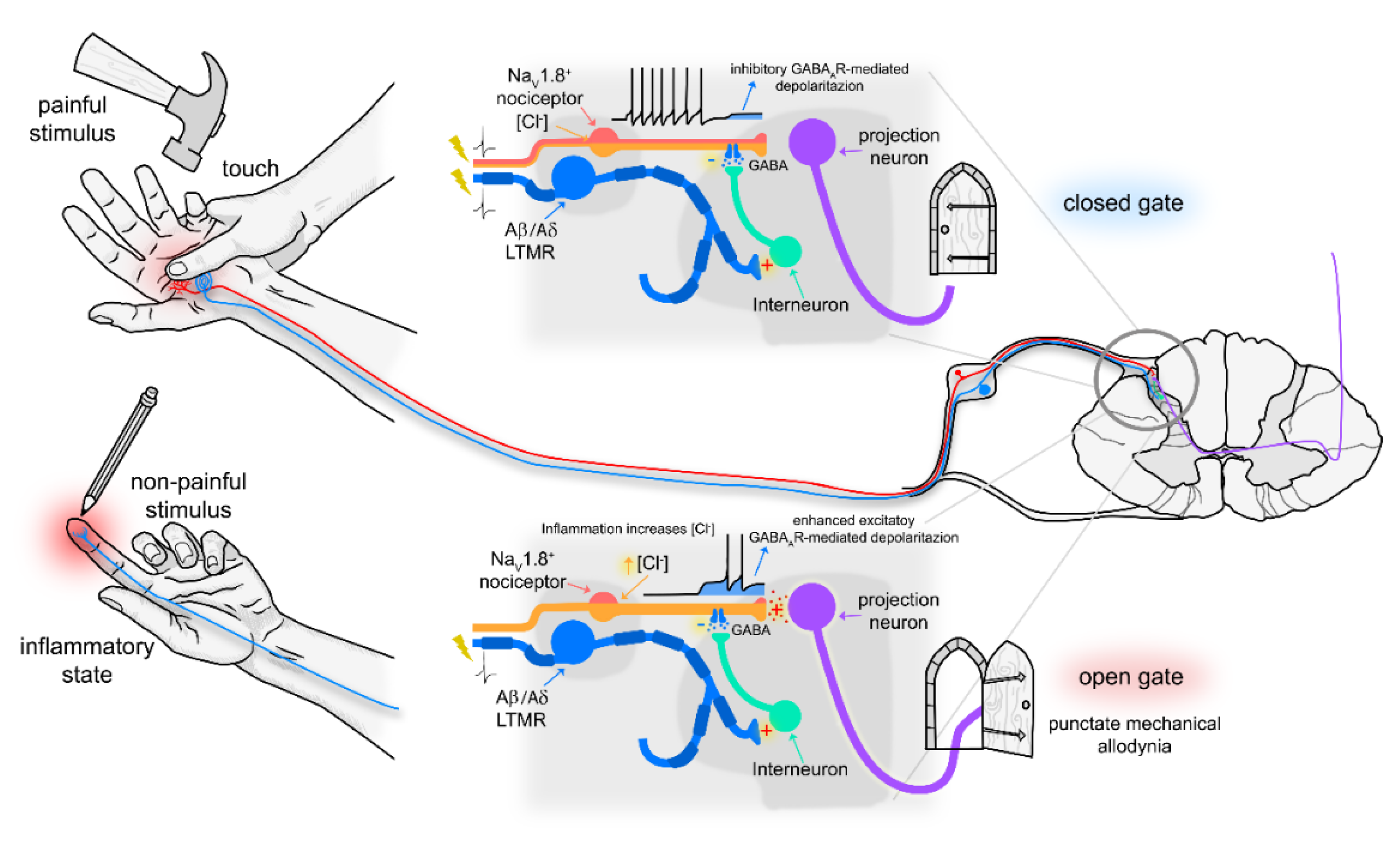
4. Discussion
Role of PV- and CR-Positive Neurons in Punctate Mechanical Allodynia
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, T.S.; Finnerup, N.B. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Campbell, J.N.; Raja, S.N.; Meyer, R.A.; Mackinnon, S.E. Myelinated afferents signal the hyperalgesia associated with nerve injury. Pain 1988, 32, 89–94. [Google Scholar] [CrossRef]
- Torsney, C.; MacDermott, A.B. Disinhibition opens the gate to pathological pain signaling in superficial neurokinin 1 recep-tor-expressing neurons in rat spinal cord. J. Neurosci. 2006, 26, 1833–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, H.; Doubell, T.P.; Woolf, C.J. Peripheral inflammation facilitates Abeta fiber-mediated synaptic input to the substantia gelatinosa of the adult rat spinal cord. J. Neurosci. 1999, 19, 859–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.T.-C.; Guo, D.; Campanelli, D.; Frattini, F.; Mayer, F.L.; Zhou, L.; Kuner, R.; Heppenstall, P.A.; Knipper, M.; Hu, J. Presynaptic GABAergic inhibition regulated by BDNF contributes to neuropathic pain induction. Nat. Commun. 2014, 5, 5331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.; Hu, J. Spinal presynaptic inhibition in pain control. Neuroscience 2014, 283, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Peirs, C.; Dallel, R.; Todd, A.J. Recent advances in our understanding of the organization of dorsal horn neuron populations and their contribution to cutaneous mechanical allodynia. J. Neural Transm. 2020, 127, 505–525. [Google Scholar] [CrossRef] [Green Version]
- Todd, A.J. GABA and Glycine in Synaptic Glomeruli of the Rat Spinal Dorsal Horn. Eur. J. Neurosci. 1996, 8, 2492–2498. [Google Scholar] [CrossRef]
- Hughes, D.I.; Mackie, M.; Nagy, G.G.; Riddell, J.S.; Maxwell, D.J.; Szabó, G.; Erdélyi, F.; Veress, G.; Szűcs, P.; Antal, M.; et al. P boutons in lamina IX of the rodent spinal cord express high levels of glutamic acid decarboxylase-65 and originate from cells in deep medial dorsal horn. Proc. Natl. Acad. Sci. USA 2005, 102, 9038–9043. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Bardoni, R.; Tong, C.-K.; Engelman, H.S.; Joseph, D.J.; Magherini, P.C.; MacDermott, A.B. Functional Expression of AMPA Receptors on Central Terminals of Rat Dorsal Root Ganglion Neurons and Presynaptic Inhibition of Glutamate Release. Neuron 2002, 35, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.L.; Kovatsis, E.M.; Pozsgai, R.Y.; Tasnim, A.; Zhang, Q.; Ginty, D.D. Distinct Modes of Presynaptic Inhibition of Cutaneous Afferents and Their Functions in Behavior. Neuron 2019, 102, 420–434.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Doroshenko, M.; Lauzadis, J.; Kanjiya, M.P.; Rebecchi, M.J.; Kaczocha, M.; Puopolo, M. Presynaptic Inhibition of Primary Nociceptive Signals to Dorsal Horn Lamina I Neurons by Dopamine. J. Neurosci. 2018, 38, 8809–8821. [Google Scholar] [CrossRef] [Green Version]
- Bardoni, R.; Tawfik, V.L.; Wang, D.; François, A.; Solorzano, C.; Shuster, S.A.; Choudhury, P.; Betelli, C.; Cassidy, C.; Smith, K.; et al. Delta Opioid Receptors Presynaptically Regulate Cutaneous Mechanosensory Neuron Input to the Spinal Cord Dorsal Horn. Neuron 2014, 81, 1312–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro-Da-Silva, A.; Pignatelli, D.; Coimbra, A. Synaptic architecture of glomeruli in superficial dorsal horn of rat spinal cord, as shown in serial reconstructions. J. Neurocytol. 1985, 14, 203–220. [Google Scholar] [CrossRef]
- Todd, A.; Watt, C.; Spike, R.; Sieghart, W. Colocalization of GABA, glycine, and their receptors at synapses in the rat spinal cord. J. Neurosci. 1996, 16, 974–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.I.; Sikander, S.; Kinnon, C.M.; Boyle, K.A.; Watanabe, M.; Callister, R.J.; Graham, B.A. Morphological, neurochemical and electrophysiological features of parvalbumin-expressing cells: A likely source of axo-axonic inputs in the mouse spinal dorsal horn. J. Physiol. 2012, 590, 3927–3951. [Google Scholar] [CrossRef]
- Ribeiro-Da-Silva, A.; Tagari, P.; Cuello, A.C. Morphological characterization of substance P-like immunoreactive glomeruli in the superficial dorsal horn of the rat spinal cord and trigeminal subnucleus caudalis: A quantitative study. J. Comp. Neurol. 1989, 281, 497–515. [Google Scholar] [CrossRef]
- Agnati, L.F.; Fuxe, K.; Zoli, M.; Ozini, I.; Toffano, G.; Ferraguti, F. A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: The volume transmission and the wiring transmission. Acta Physiol. Scand. 1986, 128, 201–207. [Google Scholar]
- Ma, W.; Saunders, P.A.; Somogyi, R.; Poulter, M.O.; Barker, J.L. Ontogeny of GABAA receptor subunit mRNAs in rat spinal cord and dorsal root ganglia. J. Comp. Neurol. 1993, 338, 337–359. [Google Scholar] [CrossRef]
- Du, X.; Hao, H.; Yang, Y.; Huang, S.; Wang, C.; Gigout, S.; Ramli, R.; Li, X.; Jaworska, E.; Edwards, I.; et al. Local GABAergic signaling within sensory ganglia controls peripheral nociceptive transmission. J. Clin. Investig. 2017, 127, 1741–1756. [Google Scholar] [CrossRef] [Green Version]
- Bonalume, V.; Caffino, L.; Castelnovo, L.F.; Faroni, A.; Liu, S.; Hu, J.; Milanese, M.; Bonanno, G.; Sohns, K.; Carr, R.; et al. Axonal GABAA stabilizes excitability in unmyelinated sensory axons secondary to NKCC1 activity. J. Physiol. 2021, 599, 4065–4084. [Google Scholar] [CrossRef] [PubMed]
- Deschenes, M.; Feltz, P.; Lamour, Y. A model for an estimate in vivo of the ionic basis of presynaptic inhibition: An intracellular analysis of the GABA-induced depolarization in rat dorsal root ganglia. Brain Res. 1976, 118, 486–493. [Google Scholar] [CrossRef]
- Gilbert, D.; Franjic-Würtz, C.; Funk, K.; Gensch, T.; Frings, S.; Möhrlen, F. Differential maturation of chloride homeostasis in primary afferent neurons of the somatosensory system. Int. J. Dev. Neurosci. 2007, 25, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, H.P.I.; Frings, S.; Gensch, T. Determination of Intracellular Chloride Concentration in Dorsal Root Ganglion Neurons by Fluorescence Lifetime Imaging. In Calcium-Activated Chloride Channels; Fuller, C.M., Ed.; Academic: Boston, USA, 2002; pp. 167–189. [Google Scholar]
- Alvarez-Leefmans, F.J.; Gamiño, S.M.; Giraldez, F.; Noguerón, I. Intracellular chloride regulation in amphibian dorsal root ganglion neurones studied with ion-selective microelectrodes. J. Physiol. 1988, 406, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, H.; Yamada, J.; Ueno, S.; Okabe, A.; Kato, H.; Sato, K.; Hashimoto, K.; Fukuda, A. Differential functional expression of cation-Cl- cotransporter mRNAs (KCC1, KCC2, and NKCC1) in rat trigeminal nervous system. Brain Res. Mol. Brain Res. 2005, 133, 12–18. [Google Scholar] [CrossRef]
- Funk, K.; Woitecki, A.; Franjic-Würtz, C.; Gensch, T.; Möhrlen, F.; Frings, S. Modulation of Chloride Homeostasis by Inflammatory Mediators in Dorsal Root Ganglion Neurons. Mol. Pain 2008, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Lu, S.; Gold, M. Persistent inflammation increases GABA-induced depolarization of rat cutaneous dorsal root ganglion neurons in vitro. Neuroscience 2012, 220, 330–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melzack, R.; Wall, P.D. Pain mechanisms: A new theory. Science 1965, 150, 971–979. [Google Scholar] [CrossRef]
- Fernandes, E.; Pechincha, C.; Luz, L.; Kokai, E.; Szucs, P.; Safronov, B. Primary afferent-driven presynaptic inhibition of C-fiber inputs to spinal lamina I neurons. Prog. Neurobiol. 2020, 188, 101786. [Google Scholar] [CrossRef]
- Agarwal, N.; Offermanns, S.; Kuner, R. Conditional gene deletion in primary nociceptive neurons of trigeminal ganglia and dorsal root ganglia. Genesis 2004, 38, 122–129. [Google Scholar] [CrossRef]
- Homanics, G.E.; DeLorey, T.M.; Firestone, L.L.; Quinlan, J.J.; Handforth, A.; Harrison, N.L.; Krasowski, M.D.; Rick, C.E.M.; Korpi, E.R.; Olsen, R.W.; et al. Mice devoid of gamma-aminobutyrate type A receptor beta3 subunit have epilepsy, cleft palate, and hypersen-sitive behavior. Proc. Natl. Acad. Sci. USA 1997, 94, 4143–4148. [Google Scholar] [CrossRef] [Green Version]
- DeLorey, T.M.; Sahbaie, P.; Hashemi, E.; Li, W.W.; Salehi, A.; Clark, D.J. Somatosensory and sensorimotor consequences associated with the heterozygous disruption of the autism can-didate gene. Gabrb3. Behav Brain Res. 2011, 216, 36–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schallert, T.; Upchurch, M.; Lobaugh, N.; Farrar, S.; Spirduso, W.; Gilliam, P.; Vaughn, D.; Wilcox, R. Tactile extinction: Distinguishing between sensorimotor and motor asymmetries in rats with unilateral nigrostriatal damage. Pharmacol. Biochem. Behav. 1982, 16, 455–462. [Google Scholar] [CrossRef]
- Fehrenbacher, J.C.; Vasko, M.R.; Duarte, D.B. Models of inflammation: Carrageenan- or complete Freund’s Adjuvant (CFA)-induced edema and hypersensitivity in the rat. In Current Protocols in Pharmacology; John Wiley & Sons, Inc.: Hoboken NJ, USA, 2012; Chapter 5. [Google Scholar]
- Zhang, Q.; He, Q.; Wang, J.; Fu, C.; Hu, H. Use of TAI-FISH to visualize neural ensembles activated by multiple stimuli. Nat. Protoc. 2018, 13, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.W.; Ribeiro-Da-Silva, A. Non-Peptidergic Primary Afferents are Presynaptic to Neurokinin-1 Receptor Immunoreactive Lamina I Projection Neurons in Rat Spinal Cord. Mol. Pain 2012, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Henry, M.A.; Nousek-Goebl, N.A.; Westrum, L.E. Light and electron microscopic localization of calcitonin gene-related peptide immunoreactivity in lamina II of the feline trigeminal pars caudalis/medullary dorsal horn: A qualitative study. Synapse 1993, 13, 99–107. [Google Scholar] [CrossRef]
- Malmberg, A.B.; Chen, C.; Tonegawa, S.; Basbaum, A.I. Preserved acute pain and reduced neuropathic pain in mice lacking PKCgamma. Science 1997, 278, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Kiernan, M.C.; Burke, D.; Andersen, K.V.; Bostock, H. Multiple measures of axonal excitability: A new approach in clinical testing. Muscle Nerve 2000, 23, 399–409. [Google Scholar] [CrossRef]
- Bostock, H.; Cikurel, K.; Burke, D. Threshold tracking techniques in the study of human peripheral nerve. Muscle Nerve 1998, 21, 137–158. [Google Scholar] [CrossRef]
- De Col, R.; Messlinger, K.; Carr, R.W. Conduction velocity is regulated by sodium channel inactivation in unmyelinated axons innervating the rat cranial meninges. J. Physiol. 2008, 586, 1089–1103. [Google Scholar] [CrossRef]
- Orefice, L.L.; Zimmerman, A.L.; Chirila, A.M.; Sleboda, S.J.; Head, J.P.; Ginty, D.D. Peripheral Mechanosensory Neuron Dysfunction Underlies Tactile and Behavioral Deficits in Mouse Models of ASDs. Cell 2016, 166, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Zhou, S.; Coggeshall, R.; Carlton, S. N-methyl-d-aspartate-induced excitation and sensitization of normal and inflamed nociceptors. Neuroscience 2003, 118, 547–562. [Google Scholar] [CrossRef]
- Abraira, V.E.; Kuehn, E.D.; Chirila, A.M.; Springel, M.W.; Toliver, A.A.; Zimmerman, A.L.; Orefice, L.L.; Boyle, K.A.; Bai, L.; Song, B.J.; et al. The Cellular and Synaptic Architecture of the Mechanosensory Dorsal Horn. Cell 2016, 168, 295–310.e19. [Google Scholar] [CrossRef] [Green Version]
- Todd, A.J.; McGill, M.M.; Shehab, S.A.S. Neurokinin 1 receptor expression by neurons in laminae I, III and IV of the rat spinal dorsal horn that project to the brainstem. Eur. J. Neurosci. 2000, 12, 689–700. [Google Scholar] [CrossRef]
- Brown, P.B.; Fuchs, J.L. Somatotopic representation of hindlimb skin in cat dorsal horn. J. Neurophysiol. 1975, 38, 1–9. [Google Scholar] [CrossRef]
- Petitjean, H.; Bourojeni, F.; Tsao, D.; Davidova, A.; Sotocinal, S.G.; Mogil, J.S.; Kania, A.; Sharif-Naeini, R. Recruitment of Spinoparabrachial Neurons by Dorsal Horn Calretinin Neurons. Cell Rep. 2019, 28, 1429–1438.e4. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, M.C.; Bostock, H.; Park, S.B.; Kaji, R.; Krarup, C.; Krishnan, A.; Kuwabara, S.; Lin, C.S.-Y.; Misawa, S.; Moldovan, M.; et al. Measurement of axonal excitability: Consensus guidelines. Clin. Neurophysiol. 2020, 131, 308–323. [Google Scholar] [CrossRef]
- Renganathan, M.; Cummins, T.R.; Hormuzdiar, W.N.; Waxman, S.G. α-SNS Produces the Slow TTX-Resistant Sodium Current in Large Cutaneous Afferent DRG Neurons. J. Neurophysiol. 2000, 84, 710–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djouhri, L.; Lawson, S.N. Increased conduction velocity of nociceptive primary afferent neurons during unilateral hindlimb in-flammation in the anaesthetised guinea-pig. Neuroscience 2001, 102, 669–679. [Google Scholar] [CrossRef]
- Torsney, C. Inflammatory Pain Unmasks Heterosynaptic Facilitation in Lamina I Neurokinin 1 Receptor-Expressing Neurons in Rat Spinal Cord. J. Neurosci. 2011, 31, 5158–5168. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Gold, M.S. Inflammatory mediators potentiate high affinity GABA(A) currents in rat dorsal root ganglion neurons. Neurosci. Lett. 2012, 518, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-L.; Lee, K.-Y.; Priest, B.; Belfer, I.; Gold, M. Inflammatory mediator-induced modulation of GABAA currents in human sensory neurons. Neuroscience 2015, 310, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Dickie, A.C.; McCormick, B.; Lukito, V.; Wilson, K.L.; Torsney, C. Inflammatory Pain Reduces C Fiber Activity-Dependent Slowing in a Sex-Dependent Manner, Amplifying Nociceptive Input to the Spinal Cord. J. Neurosci. 2017, 37, 6488–6502. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.-Y.; Tang, X.-C.; Chang, Y.-C.; Xu, Z.-Z.; Chen, Q.-Y.; Cao, N.; Kong, L.-J.; Wang, Y.; Ma, K.-T.; Li, L.; et al. Improving NKCC1 Function Increases the Excitability of DRG Neurons Exacerbating Pain Induced After TRPV1 Activation of Primary Sensory Neurons. Front. Cell. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Laird, J.M.; García-Nicas, E.; Delpire, E.J.; Cervero, F. Presynaptic inhibition and spinal pain processing in mice: A possible role of the NKCC1 cation-chloride co-transporter in hyperalgesia. Neurosci. Lett. 2004, 361, 200–203. [Google Scholar] [CrossRef]
- Sung, K.W.; Kirby, M.; McDonald, M.P.; Lovinger, D.M.; Delpire, E. Abnormal GABAA receptor-mediated currents in dorsal root ganglion neurons isolated from Na-K-2Cl cotransporter null mice. J. Neurosci. 2000, 20, 7531–7538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudomin, P.; Schmidt, R.F. Presynaptic inhibition in the vertebrate spinal cord revisited. Exp. Brain Res. 1999, 129, 1–37. [Google Scholar] [CrossRef]
- Willis, W.D., Jr. Dorsal root potentials and dorsal root reflexes: A double-edged sword. Exp. Brain Res. 1999, 124, 395–421. [Google Scholar] [CrossRef]
- Sluka, K.A.; Rees, H.; Westlund, K.N.; Willis, W.D. Fiber types contributing to dorsal root reflexes induced by joint inflammation in cats and monkeys. J. Neurophysiol. 1995, 74, 981–989. [Google Scholar] [CrossRef]
- Reichling, D.B.; Kyrozis, A.; Wang, J.; MacDermott, A.B. Mechanisms of GABA and glycine depolarization-induced calcium transients in rat dorsal horn neurons. J. Physiol. 1994, 476, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Schöbel, N.; Radtke, D.; Lübbert, M.; Gisselmann, G.; Lehmann, R.; Cichy, A.; Schreiner, B.S.P.; Altmüller, J.; Spector, A.C.; Spehr, J.; et al. Trigeminal Ganglion Neurons of Mice Show Intracellular Chloride Accumulation and Chloride-Dependent Amplification of Capsaicin-Induced Responses. PLoS ONE 2012, 7, e48005. [Google Scholar] [CrossRef] [PubMed]
- Aptel, H.; Hilaire, C.; Pieraut, S.; Boukhaddaoui, H.; Mallié, S.; Valmier, J.; Scamps, F. The Cav3.2/alpha1H T-type Ca2+ current is a molecular determinant of excitatory effects of GABA in adult sensory neurons. Mol. Cell Neurosci. 2007, 36, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, J.L.; Yarnitsky, D. Mechanical hyperalgesias in neuropathic pain patients: Dynamic and static subtypes. Ann. Neurol. 1993, 33, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Sandkühler, J. Models and Mechanisms of Hyperalgesia and Allodynia. Physiol. Rev. 2009, 89, 707–758. [Google Scholar] [CrossRef]
- Cavanaugh, D.J.; Lee, H.; Lo, L.; Shields, S.D.; Zylka, M.J.; Basbaum, A.I.; Anderson, D.J. Distinct subsets of unmyelinated primary sensory fibers mediate behavioral responses to noxious thermal and mechanical stimuli. Proc. Natl. Acad. Sci. USA 2009, 106, 9075–9080. [Google Scholar] [CrossRef] [Green Version]
- Daou, I.; Beaudry, H.; Ase, A.R.; Wieskopf, J.S.; Ribeiro-Da-Silva, A.; Mogil, J.S.; Séguéla, P. Optogenetic Silencing of Nav1.8-Positive Afferents Alleviates Inflammatory and Neuropathic Pain. eNeuro 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Peirs, C.; Williams, S.-P.G.; Zhao, X.; Arokiaraj, C.M.; Ferreira, D.W.; Noh, M.-C.; Smith, K.M.; Halder, P.; Corrigan, K.A.; Gedeon, J.Y.; et al. Mechanical Allodynia Circuitry in the Dorsal Horn Is Defined by the Nature of the Injury. Neuron 2021, 109, 73–90.e7. [Google Scholar] [CrossRef]
- Petitjean, H.; Pawlowski, S.A.; Fraine, S.L.; Sharif, B.; Hamad, D.; Fatima, T.; Berg, J.; Brown, C.M.; Jan, L.-Y.; Ribeiro-Da-Silva, A.; et al. Dorsal Horn Parvalbumin Neurons Are Gate-Keepers of Touch-Evoked Pain after Nerve Injury. Cell Rep. 2015, 13, 1246–1257. [Google Scholar] [CrossRef] [Green Version]
- Peirs, C.; Williams, S.-P.G.; Zhao, X.; Walsh, C.E.; Gedeon, J.Y.; Cagle, N.E.; Goldring, A.C.; Hioki, H.; Liu, Z.; Marell, P.S.; et al. Dorsal Horn Circuits for Persistent Mechanical Pain. Neuron 2015, 87, 797–812. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Chen, J.; Wang, W.; Wang, W.; Koshimizu, Y.; Wei, Y.-Y.; Kaneko, T.; Li, Y.-Q.; Wu, S.-X. Neurochemical properties of enkephalinergic neurons in lumbar spinal dorsal horn revealed by preproenkephalin-green fluorescent protein transgenic mice. J. Neurochem. 2010, 113, 1555–1564. [Google Scholar] [CrossRef]
- Smith, K.M.; Browne, T.J.; Davis, O.C.; Coyle, A.; Boyle, K.A.; Watanabe, M.; Dickinson, S.A.; Iredale, J.A.; Gradwell, M.A.; Graham, B.A.; et al. Calretinin positive neurons form an excitatory amplifier network in the spinal cord dorsal horn. eLife 2019, 8, e49190. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Bonalume, V.; Gao, Q.; Chen, J.T.-C.; Rohr, K.; Hu, J.; Carr, R. Pre-Synaptic GABAA in NaV1.8+ Primary Afferents Is Required for the Development of Punctate but Not Dynamic Mechanical Allodynia following CFA Inflammation. Cells 2022, 11, 2390. https://doi.org/10.3390/cells11152390
Liu S, Bonalume V, Gao Q, Chen JT-C, Rohr K, Hu J, Carr R. Pre-Synaptic GABAA in NaV1.8+ Primary Afferents Is Required for the Development of Punctate but Not Dynamic Mechanical Allodynia following CFA Inflammation. Cells. 2022; 11(15):2390. https://doi.org/10.3390/cells11152390
Chicago/Turabian StyleLiu, Sheng, Veronica Bonalume, Qi Gao, Jeremy Tsung-Chieh Chen, Karl Rohr, Jing Hu, and Richard Carr. 2022. "Pre-Synaptic GABAA in NaV1.8+ Primary Afferents Is Required for the Development of Punctate but Not Dynamic Mechanical Allodynia following CFA Inflammation" Cells 11, no. 15: 2390. https://doi.org/10.3390/cells11152390