
Regional Development of Glioblastoma: The Anatomical Conundrum of Cancer Biology and Its Surgical Implication

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Neural Stem Cells
3.2. Embryological Development
3.3. Astrocytes and the Spatiotemporal Diversity
3.4. Myelin-Associated Signaling
3.5. Neurons and Glutamate Transporters
3.6. Extracellular Matrix and Peritumoral Environment
3.7. Atlas of GBM and the Lateralization Puzzle
3.8. Surgical Implications
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Wrensch, M.; Minn, Y.; Chew, T.; Bondy, M.; Berger, M.S. Epidemiology of primary brain tumors: Current concepts and review of the literature. Neuro-Oncology 2002, 4, 278–299. [Google Scholar] [CrossRef]
- Iwadate, Y.; Fukuda, K.; Matsutani, T.; Saeki, N. Intrinsic protective mechanisms of the neuron-glia network against glioma invasion. J. Clin. Neurosci. 2016, 26, 19–25. [Google Scholar] [CrossRef]
- Altieri, R.; Zenga, F.; Ducati, A.; Melcarne, A.; Cofano, F.; Mammi, M.; Di Perna, G.; Savastano, R.; Garbossa, D. Tumor location and patient age predict biological signatures of high-grade gliomas. Neurosurg. Rev. 2018, 41, 599–604. [Google Scholar] [CrossRef]
- Sampetrean, O.; Saya, H. Modeling phenotypes of malignant gliomas. Cancer Sci. 2018, 109, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Lathia, J.D.; Mack, S.C.; Mulkearns-Hubert, E.E.; Valentim, C.L.L.; Rich, J.N. Cancer stem cells in glioblastoma. Genes Dev. 2015, 29, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, A.; García-Verdugo, J.M.; Tramontin, A.D. A unified hypothesis on the lineage of neural stem cells. Nat. Rev. Neurosci. 2001, 2, 287–293. [Google Scholar] [CrossRef]
- Zheng, H.; Ying, H.; Yan, H.; Kimmelman, A.C.; Hiller, D.J.; Chen, A.-J.; Perry, S.R.; Tonon, G.; Chu, G.C.; Ding, Z.; et al. p53 and Pten control neural and glioma stem/progenitor cell renewal and differentiation. Nature 2008, 455, 1129–1133. [Google Scholar] [CrossRef]
- Duan, S.; Yuan, G.; Liu, X.; Ren, R.; Li, J.; Zhang, W.; Wu, J.; Xu, X.; Fu, L.; Li, Y.; et al. PTEN deficiency reprogrammes human neural stem cells towards a glioblastoma stem cell-like phenotype. Nat. Commun. 2015, 6, 10068. [Google Scholar] [CrossRef]
- Fontán-Lozano, Á.; Morcuende, S.; Davis-López de Carrizosa, M.A.; Benítez-Temiño, B.; Mejías, R.; Matarredona, E.R. To Become or Not to Become Tumorigenic: Subventricular Zone Versus Hippocampal Neural Stem Cells. Front. Oncol. 2020, 10, 2644. [Google Scholar] [CrossRef]
- Gimple, R.C.; Bhargava, S.; Dixit, D.; Rich, J.N. Glioblastoma stem cells: Lessons from the tumor hierarchy in a lethal cancer. Genes Dev. 2019, 33, 591–609. [Google Scholar] [CrossRef]
- Sidaway, P. CNS cancer: Glioblastoma subtypes revisited. Nat. Rev. Clin. Oncol. 2017, 14, 587. [Google Scholar] [CrossRef]
- Wu, C.; Song, H.; Fu, X.; Li, S.; Jiang, T. Transcriptomic Analysis of Glioma Based on IDH Status Identifies ACAA2 as a Prognostic Factor in Lower Grade Glioma. Biomed. Res. Int. 2020, 2020, 1086792. [Google Scholar] [CrossRef] [Green Version]
- Bruzzone, M.G.; Eoli, M.; Cuccarini, V.; Grisoli, M.; Valletta, L.; Finocchiaro, G. Genetic signature of adult gliomas and correlation with MRI features. Expert Rev. Mol. Diagn. 2009, 9, 709–720. [Google Scholar] [CrossRef]
- McLendon, R.; Friedman, A.; Bigner, D.; Van Meir, E.G.; Brat, D.J.; Mastrogianakis, G.M.; Olson, J.J.; Mikkelsen, T.; Lehman, N.; Aldape, K.; et al. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef]
- Ellingson, B.M.; Lai, A.; Harris, R.J.; Selfridge, J.M.; Yong, W.H.; Das, K.; Pope, W.B.; Nghiemphu, P.L.; Vinters, H.V.; Liau, L.M.; et al. Probabilistic Radiographic Atlas of Glioblastoma Phenotypes. Am. J. Neuroradiol. 2013, 34, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Skjulsvik, A.J.; Bø, H.K.; Jakola, A.S.; Berntsen, E.M.; Bø, L.E.; Reinertsen, I.; Myrmel, K.S.; Sjåvik, K.; Åberg, K.; Berg, T.; et al. Is the anatomical distribution of low-grade gliomas linked to regions of gliogenesis? J. Neurooncol. 2020, 147, 147–157. [Google Scholar] [CrossRef] [Green Version]
- De Luca, C.; Colangelo, A.M.; Virtuoso, A.; Alberghina, L.; Papa, M. Neurons, Glia, Extracellular Matrix and Neurovascular Unit: A Systems Biology Approach to the Complexity of Synaptic Plasticity in Health and Disease. Int. J. Mol. Sci. 2020, 21, 1539. [Google Scholar] [CrossRef] [Green Version]
- Ghashghaei, H.T.; Lai, C.; Anton, E.S. Neuronal migration in the adult brain: Are we there yet? Nat. Rev. Neurosci. 2007, 8, 141–151. [Google Scholar] [CrossRef]
- Cope, E.C.; Gould, E. Adult Neurogenesis, Glia, and the Extracellular Matrix. Cell Stem Cell 2019, 24, 690–705. [Google Scholar] [CrossRef]
- Fuentealba, L.C.; Rompani, S.B.; Parraguez, J.I.; Obernier, K.; Romero, R.; Cepko, C.L.; Alvarez-Buylla, A. Embryonic origin of postnatal neural stem cells. Cell 2015, 161, 1644–1655. [Google Scholar] [CrossRef] [Green Version]
- Doetsch, F.; García-Verdugo, J.M.; Alvarez-Buylla, A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J. Neurosci. 1997, 17, 5046–5061. [Google Scholar] [CrossRef]
- Joel, M.; Sandberg, C.J.; Boulland, J.-L.; Vik-Mo, E.O.; Langmoen, I.A.; Glover, J.C. Inhibition of Tumor Formation and Redirected Differentiation of Glioblastoma Cells in a Xenotypic Embryonic Environment. Available online: https://anatomypubs.onlinelibrary.wiley.com/doi/abs/10.1002/dvdy.24001 (accessed on 18 February 2021).
- Stoll, E.A.; Horner, P.J.; Rostomily, R.C. The impact of age on oncogenic potential: Tumor-initiating cells and the brain microenvironment. Aging Cell 2013, 12, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Glass, R.; Synowitz, M.; Kronenberg, G.; Walzlein, J.-H.; Markovic, D.S.; Wang, L.-P.; Gast, D.; Kiwit, J.; Kempermann, G.; Kettenmann, H. Glioblastoma-Induced Attraction of Endogenous Neural Precursor Cells Is Associated with Improved Survival. J. Neurosci. 2005, 25, 2637–2646. [Google Scholar] [CrossRef] [Green Version]
- Walzlein, J.-H.; Synowitz, M.; Engels, B.; Markovic, D.S.; Gabrusiewicz, K.; Nikolaev, E.; Yoshikawa, K.; Kaminska, B.; Kempermann, G.; Uckert, W.; et al. The Antitumorigenic Response of Neural Precursors Depends on Subventricular Proliferation and Age. Stem Cells 2008, 26, 2945–2954. [Google Scholar] [CrossRef] [Green Version]
- Friedmann-Morvinski, D.; Bushong, E.A.; Ke, E.; Soda, Y.; Marumoto, T.; Singer, O.; Ellisman, M.H.; Verma, I.M. Dedifferentiation of neurons and astrocytes by oncogenes can induce gliomas in mice. Science 2012, 338, 1080–1084. [Google Scholar] [CrossRef] [Green Version]
- Piccirillo, S.G.M.; Reynolds, B.A.; Zanetti, N.; Lamorte, G.; Binda, E.; Broggi, G.; Brem, H.; Olivi, A.; Dimeco, F.; Vescovi, A.L. Bone morphogenetic proteins inhibit the tumorigenic potential of human brain tumour-initiating cells. Nature 2006, 444, 761–765. [Google Scholar] [CrossRef]
- Chirasani, S.R.; Sternjak, A.; Wend, P.; Momma, S.; Campos, B.; Herrmann, I.M.; Graf, D.; Mitsiadis, T.; Herold-Mende, C.; Besser, D.; et al. Bone morphogenetic protein-7 release from endogenous neural precursor cells suppresses the tumourigenicity of stem-like glioblastoma cells. Brain 2010, 133, 1961–1972. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Daniels, S.B.; Lennington, J.B.; Notti, R.Q.; Conover, J.C. The aging neurogenic subventricular zone. Aging Cell 2006, 5, 139–152. [Google Scholar] [CrossRef]
- Stoll, E.A.; Habibi, B.A.; Mikheev, A.M.; Lasiene, J.; Massey, S.C.; Swanson, K.R.; Rostomily, R.C.; Horner, P.J. Increased re-entry into cell cycle mitigates age-related neurogenic decline in the murine subventricular zone. Stem Cells 2011, 29, 2005–2017. [Google Scholar] [CrossRef] [Green Version]
- Yaşargil, M.G.; Kadri, P.A.S.; Yasargil, D.C.H. Microsurgery for malignant gliomas. J. Neurooncol. 2004, 69, 67–81. [Google Scholar] [CrossRef]
- Lojewski, X.; Wegner, F.; Wegner, F.; Hallmeyer-Elgner, S.; Kirsch, M.; Schwarz, J.; Schwarz, J.; Schwarz, J. Human Adult White Matter Progenitor Cells Are Multipotent Neuroprogenitors Similar to Adult Hippocampal Progenitors. Stem Cells Transl. Med. 2014, 3, 458–469. [Google Scholar] [CrossRef]
- Oliver-De La Cruz, J.; Carrión-Navarro, J.; García-Romero, N.; Gutiérrez-Martín, A.; Lázaro-Ibáñez, E.; Escobedo-Lucea, C.; Perona, R.; Belda-Iniesta, C.; Ayuso-Sacido, A. SOX2+ Cell Population from Normal Human Brain White Matter Is Able to Generate Mature Oligodendrocytes. PLoS ONE 2014, 9, e99253. [Google Scholar] [CrossRef] [Green Version]
- Sanai, N.; Alvarez-Buylla, A.; Berger, M.S. Neural stem cells and the origin of gliomas. N. Engl. J. Med. 2005, 353, 811–822. [Google Scholar] [CrossRef]
- Anthony, T.E.; Klein, C.; Fishell, G.; Heintz, N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron 2004, 41, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Muzio, L.; Mallamaci, A. Foxg1 Confines Cajal-Retzius Neuronogenesis and Hippocampal Morphogenesis to the Dorsomedial Pallium. J. Neurosci. 2005, 25, 4435–4441. [Google Scholar] [CrossRef] [Green Version]
- Greig, L.C.; Woodworth, M.B.; Galazo, M.J.; Padmanabhan, H.; Macklis, J.D. Molecular logic of neocortical projection neuron specification, development and diversity. Nat. Rev. Neurosci. 2013, 14, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Wang, Y.; Dimos, J.T.; Fasano, C.A.; Phoenix, T.N.; Lemischka, I.R.; Ivanova, N.B.; Stifani, S.; Morrisey, E.E.; Temple, S. The timing of cortical neurogenesis is encoded within lineages of individual progenitor cells. Nat. Neurosci. 2006, 9, 743–751. [Google Scholar] [CrossRef]
- Molofsky, A.V.; Deneen, B. Astrocyte development: A Guide for the Perplexed. Glia 2015, 63, 1320–1329. [Google Scholar] [CrossRef]
- Merkle, F.T.; Fuentealba, L.C.; Sanders, T.A.; Magno, L.; Kessaris, N.; Alvarez-Buylla, A. Adult neural stem cells in distinct microdomains generate previously unknown interneuron types. Nat. Neurosci. 2014, 17, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Perez, O.; Romero-Rodriguez, R.; Soriano-Navarro, M.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Epidermal growth factor induces the progeny of subventricular zone type B cells to migrate and differentiate into oligodendrocytes. Stem Cells 2009, 27, 2032–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.A.; Cha, S.; Mayo, M.C.; Chen, M.-H.; Keles, E.; VandenBerg, S.; Berger, M.S. Relationship of glioblastoma multiforme to neural stem cell regions predicts invasive and multifocal tumor phenotype. Neuro-Oncology 2007, 9, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.R. Specification and Morph.hogenesis of Astrocytes. Science 2010, 330, 774–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haim, L.B.; Rowitch, D.H. Functional diversity of astrocytes in neural circuit regulation. Nat. Rev. Neurosci. 2017, 18, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Emsley, J.G.; Macklis, J.D. Astroglial heterogeneity closely reflects the neuronal-defined anatomy of the adult murine CNS. Neuron Glia Biol. 2006, 2, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Papa, M.; De Luca, C.; Petta, F.; Alberghina, L.; Cirillo, G. Astrocyte-neuron interplay in maladaptive plasticity. Neurosci. Biobehav. Rev. 2014, 42, 35–54. [Google Scholar] [CrossRef]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.C.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G.; et al. Uniquely hominid features of adult human astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef]
- Lin, C.-C.J.; Yu, K.; Hatcher, A.; Huang, T.-W.; Lee, H.K.; Carlson, J.; Weston, M.C.; Chen, F.; Zhang, Y.; Zhu, W.; et al. Identification of diverse astrocyte populations and their malignant analogs. Nat. Neurosci. 2017, 20, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Sala, L.; Cirillo, G.; Riva, G.; Romano, G.; Giussani, C.; Cialdella, A.; Todisco, A.; Virtuoso, A.; Cerrito, M.G.; Bentivegna, A.; et al. Specific Expression of a New Bruton Tyrosine Kinase Isoform (p65BTK) in the Glioblastoma Gemistocytic Histotype. Front. Mol. Neurosci. 2019, 12, 2. [Google Scholar] [CrossRef]
- Laviv, Y.; Berkowitz, S.; Kanner, A.K.; Fichman, S.; Benouaich-Amiel, A.; Siegal, T.; Yust-Katz, S. Gemistocytes in newly diagnosed glioblastoma multiforme: Clinical significance and practical implications in the modern era. J. Clin. Neurosci. 2021, 88, 120–127. [Google Scholar] [CrossRef]
- Hong, J.-H.; Kang, S.; Sa, J.K.; Park, G.; Oh, Y.T.; Kim, T.H.; Yin, J.; Kim, S.S.; D’Angelo, F.; Koo, H.; et al. Modulation of Nogo receptor 1 expression orchestrates myelin-associated infiltration of glioblastoma. Brain 2021, 144, 636–654. [Google Scholar] [CrossRef] [PubMed]
- Curinga, G.; Smith, G.M. Molecular/genetic manipulation of extrinsic axon guidance factors for CNS repair and regeneration. Exp. Neurol. 2008, 209, 333–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braisted, J.E.; Catalano, S.M.; Stimac, R.; Kennedy, T.E.; Tessier-Lavigne, M.; Shatz, C.J.; O’Leary, D.D.M. Netrin-1 Promotes Thalamic Axon Growth and Is Required for Proper Development of the Thalamocortical Projection. J. Neurosci. 2000, 20, 5792–5801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrivée, B.; Freitas, C.; Trombe, M.; Lv, X.; DeLafarge, B.; Yuan, L.; Bouvrée, K.; Bréant, C.; Toro, R.D.; Bréchot, N.; et al. Activation of the UNC5B receptor by Netrin-1 inhibits sprouting angiogenesis. Genes Dev. 2007, 21, 2433–2447. [Google Scholar] [CrossRef] [Green Version]
- Jarjour, A.A.; Durko, M.; Luk, T.L.; Marçal, N.; Shekarabi, M.; Kennedy, T.E. Autocrine Netrin Function Inhibits Glioma Cell Motility and Promotes Focal Adhesion Formation. PLoS ONE 2011, 6, e25408. [Google Scholar] [CrossRef]
- Yiin, J.-J.; Hu, B.; Jarzynka, M.J.; Feng, H.; Liu, K.-W.; Wu, J.Y.; Ma, H.-I.; Cheng, S.-Y. Slit2 inhibits glioma cell invasion in the brain by suppression of Cdc42 activity. Neuro-Oncology 2009, 11, 779–789. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, X.; Georgakilas, A.G.; Chen, P.; Hu, H.; Yang, Y.; Tian, S.; Xia, L.; Zhang, J.; Cai, X.; et al. Tetramethylpyrazine (TMP) protects cerebral neurocytes and inhibits glioma by down regulating chemokine receptor CXCR4 expression. Cancer Lett. 2013, 336, 281–289. [Google Scholar] [CrossRef]
- Liu, Y.; Carlsson, R.; Ambjørn, M.; Hasan, M.; Badn, W.; Darabi, A.; Siesjö, P.; Issazadeh-Navikas, S. PD-L1 Expression by Neurons Nearby Tumors Indicates Better Prognosis in Glioblastoma Patients. J. Neurosci. 2013, 33, 14231–14245. [Google Scholar] [CrossRef]
- Nieto-Sampedro, M.; Valle-Argos, B.; Gómez-Nicola, D.; Fernández-Mayoralas, A.; Nieto-Díaz, M. Inhibitors of Glioma Growth that Reveal the Tumour to the Immune System. Clin. Med. Insights Oncol. 2011, 5, CMO-S7685. [Google Scholar] [CrossRef]
- Ye, Z.-C.; Sontheimer, H. Glioma Cells Release Excitotoxic Concentrations of Glutamate. Cancer Res. 1999, 59, 4383–4391. [Google Scholar]
- Martinez-Lozada, Z.; Robinson, M.B. Reciprocal communication between astrocytes and endothelial cells is required for astrocytic glutamate transporter 1 (GLT-1) expression. Neurochem. Int. 2020, 139, 104787. [Google Scholar] [CrossRef] [PubMed]
- Lehre, K.P.; Danbolt, N.C. The Number of Glutamate Transporter Subtype Molecules at Glutamatergic Synapses: Chemical and Stereological Quantification in Young Adult Rat Brain. J. Neurosci. 1998, 18, 8751–8757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlyle, B.C.; Kitchen, R.R.; Kanyo, J.E.; Voss, E.Z.; Pletikos, M.; Sousa, A.M.M.; Lam, T.T.; Gerstein, M.B.; Sestan, N.; Nairn, A.C. A multiregional proteomic survey of the postnatal human brain. Nat. Neurosci. 2017, 20, 1787–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Sanes, J.R. Neuronal cell-type classification: Challenges, opportunities and the path forward. Nat. Rev. Neurosci. 2017, 18, 530–546. [Google Scholar] [CrossRef]
- Zilles, K. Brodmann: A pioneer of human brain mapping—his impact on concepts of cortical organization. Brain 2018, 141, 3262–3278. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.B.; Walsh, C.A. Cerebral cortical neuron diversity and development at single-cell resolution. Curr. Opin. Neurobiol. 2017, 42, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Scherer, H.J. Structural Development in Gliomas. Am. J. Cancer 1938, 34, 333–351. [Google Scholar] [CrossRef]
- Mughal, A.A.; Zhang, L.; Fayzullin, A.; Server, A.; Li, Y.; Wu, Y.; Glass, R.; Meling, T.; Langmoen, I.A.; Leergaard, T.B.; et al. Patterns of Invasive Growth in Malignant Gliomas—The Hippocampus Emerges as an Invasion-Spared Brain Region. Neoplasia 2018, 20, 643–656. [Google Scholar] [CrossRef]
- Esmaeili, M.; Stensjøen, A.L.; Berntsen, E.M.; Solheim, O.; Reinertsen, I. The Direction of Tumour Growth in Glioblastoma Patients. Sci. Rep. 2018, 8, 1199. [Google Scholar] [CrossRef]
- Virtuoso, A.; Giovannoni, R.; De Luca, C.; Gargano, F.; Cerasuolo, M.; Maggio, N.; Lavitrano, M.; Papa, M. The Glioblastoma Microenvironment: Morphology, Metabolism, and Molecular Signature of Glial Dynamics to Discover Metabolic Rewiring Sequence. Int. J. Mol. Sci. 2021, 22, 3301. [Google Scholar] [CrossRef]
- Wade, A.; Robinson, A.E.; Engler, J.R.; Petritsch, C.; James, C.D.; Phillips, J.J. Proteoglycans and their roles in brain cancer. FEBS J. 2013, 280, 2399–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirillo, G.; Colangelo, A.M.; Bianco, M.R.; Cavaliere, C.; Zaccaro, L.; Sarmientos, P.; Alberghina, L.; Papa, M. BB14, a Nerve Growth Factor (NGF)-like peptide shown to be effective in reducing reactive astrogliosis and restoring synaptic homeostasis in a rat model of peripheral nerve injury. Biotechnol. Adv. 2012, 30, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Silver, D.J.; Siebzehnrubl, F.A.; Schildts, M.J.; Yachnis, A.T.; Smith, G.M.; Smith, A.A.; Scheffler, B.; Reynolds, B.A.; Silver, J.; Steindler, D.A. Chondroitin Sulfate Proteoglycans Potently Inhibit Invasion and Serve as a Central Organizer of the Brain Tumor Microenvironment. J. Neurosci. 2013, 33, 15603–15617. [Google Scholar] [CrossRef] [Green Version]
- Sim, H.; Hu, B.; Viapiano, M.S. Reduced Expression of the Hyaluronan and Proteoglycan Link Proteins in Malignant Gliomas*. J. Biol. Chem. 2009, 284, 26547–26556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chen, X.; Sun, L.; Bi, X.; He, H.; Chen, L.; Pang, J. MicroRNA-34a inhibits cell growth and migration in human glioma cells via MMP-9. Mol. Med. Rep. 2019, 20, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Xue, W.; Zhao, W.; Schachner, M. Expression and function of chondroitin 4-sulfate and chondroitin 6-sulfate in human glioma. FASEB J. 2020, 34, 2853–2868. [Google Scholar] [CrossRef] [Green Version]
- De Luca, C.; Papa, M. Looking Inside the Matrix: Perineuronal Nets in Plasticity, Maladaptive Plasticity and Neurological Disorders. Neurochem. Res. 2016, 41, 1507–1515. [Google Scholar] [CrossRef]
- Dauth, S.; Grevesse, T.; Pantazopoulos, H.; Campbell, P.H.; Maoz, B.M.; Berretta, S.; Parker, K.K. Extracellular matrix protein expression is brain region dependent. J. Comp. Neurol. 2016, 524, 1309–1336. [Google Scholar] [CrossRef]
- Altieri, R.; Barbagallo, D.; Certo, F.; Broggi, G.; Ragusa, M.; Di Pietro, C.; Caltabiano, R.; Magro, G.; Peschillo, S.; Purrello, M.; et al. Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia-Glioma Cross-Talk. Brain Sci. 2021, 11, 200. [Google Scholar] [CrossRef]
- De Luca, C.; Papa, M. Matrix Metalloproteinases, Neural Extracellular Matrix, and Central Nervous System Pathology. Prog. Mol. Biol. Transl. Sci. 2017, 148, 167–202. [Google Scholar] [CrossRef]
- Penfield, W. Microglia and the Process of Phagocytosis in Gliomas. Am. J. Pathol. 1925, 1, 77–90.15. [Google Scholar] [PubMed]
- Altieri, R.; Melcarne, A.; Junemann, C.; Zeppa, P.; Zenga, F.; Garbossa, D.; Certo, F.; Barbagallo, G. Inferior Fronto-Occipital fascicle anatomy in brain tumor surgeries: From anatomy lab to surgical theater. J. Clin. Neurosci. 2019, 68, 290–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altieri, R.; Raimondo, S.; Tiddia, C.; Sammarco, D.; Cofano, F.; Zeppa, P.; Monticelli, M.; Melcarne, A.; Junemann, C.; Zenga, F.; et al. Glioma surgery: From preservation of motor skills to conservation of cognitive functions. J. Clin. Neurosci. 2019, 70, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Cui, Y.; Liu, X.; Ren, X.; Li, M.; Lin, S. Proliferation-dominant high-grade astrocytoma: Survival benefit associated with extensive resection of FLAIR abnormality region. J. Neurosurg. 2019, 132, 998–1005. [Google Scholar] [CrossRef]
- Fyllingen, E.H.; Bø, L.E.; Reinertsen, I.; Jakola, A.S.; Sagberg, L.M.; Berntsen, E.M.; Salvesen, Ø.; Solheim, O. Survival of glioblastoma in relation to tumor location: A statistical tumor atlas of a population-based cohort. Acta Neurochir. (Wien.) 2021, 163, 1895–1905. [Google Scholar] [CrossRef]
- Lai, A.; Kharbanda, S.; Pope, W.B.; Tran, A.; Solis, O.E.; Peale, F.; Forrest, W.F.; Pujara, K.; Carrillo, J.A.; Pandita, A.; et al. Evidence for sequenced molecular evolution of IDH1 mutant glioblastoma from a distinct cell of origin. J. Clin. Oncol. 2011, 29, 4482–4490. [Google Scholar] [CrossRef] [Green Version]
- Al-Holou, W.N.; Hodges, T.R.; Everson, R.G.; Freeman, J.; Zhou, S.; Suki, D.; Rao, G.; Ferguson, S.D.; Heimberger, A.B.; McCutcheon, I.E.; et al. Perilesional Resection of Glioblastoma Is Independently Associated With Improved Outcomes. Neurosurgery 2020, 86, 112–121. [Google Scholar] [CrossRef]
- Sun, T.; Patoine, C.; Abu-Khalil, A.; Visvader, J.; Sum, E.; Cherry, T.J.; Orkin, S.H.; Geschwind, D.H.; Walsh, C.A. Early Asymmetry of Gene Transcription in Embryonic Human Left and Right Cerebral Cortex. Science 2005, 308, 1794–1798. [Google Scholar] [CrossRef] [Green Version]
- Ferronha, T.; Rabadán, M.A.; Gil-Guiñon, E.; Le Dréau, G.; de Torres, C.; Martí, E. LMO4 is an essential cofactor in the Snail2-mediated epithelial-to-mesenchymal transition of neuroblastoma and neural crest cells. J. Neurosci. 2013, 33, 2773–2783. [Google Scholar] [CrossRef] [Green Version]
- Al-Holou, W.N.; Suki, D.; Hodges, T.R.; Everson, R.G.; Freeman, J.; Ferguson, S.D.; McCutcheon, I.E.; Prabhu, S.S.; Weinberg, J.S.; Sawaya, R.; et al. Circumferential sulcus-guided resection technique for improved outcomes of low-grade gliomas. J. Neurosurg. 2022, 1, 1–11. [Google Scholar] [CrossRef]
- Bohman, L.-E.; Swanson, K.R.; Moore, J.L.; Rockne, R.; Mandigo, C.; Hankinson, T.; Assanah, M.; Canoll, P.; Bruce, J.N. Magnetic resonance imaging characteristics of glioblastoma multiforme: Implications for understanding glioma ontogeny. Neurosurgery 2010, 67, 1319–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffau, H.; Capelle, L. Preferential brain locations of low-grade gliomas. Cancer 2004, 100, 2622–2626. [Google Scholar] [CrossRef] [PubMed]






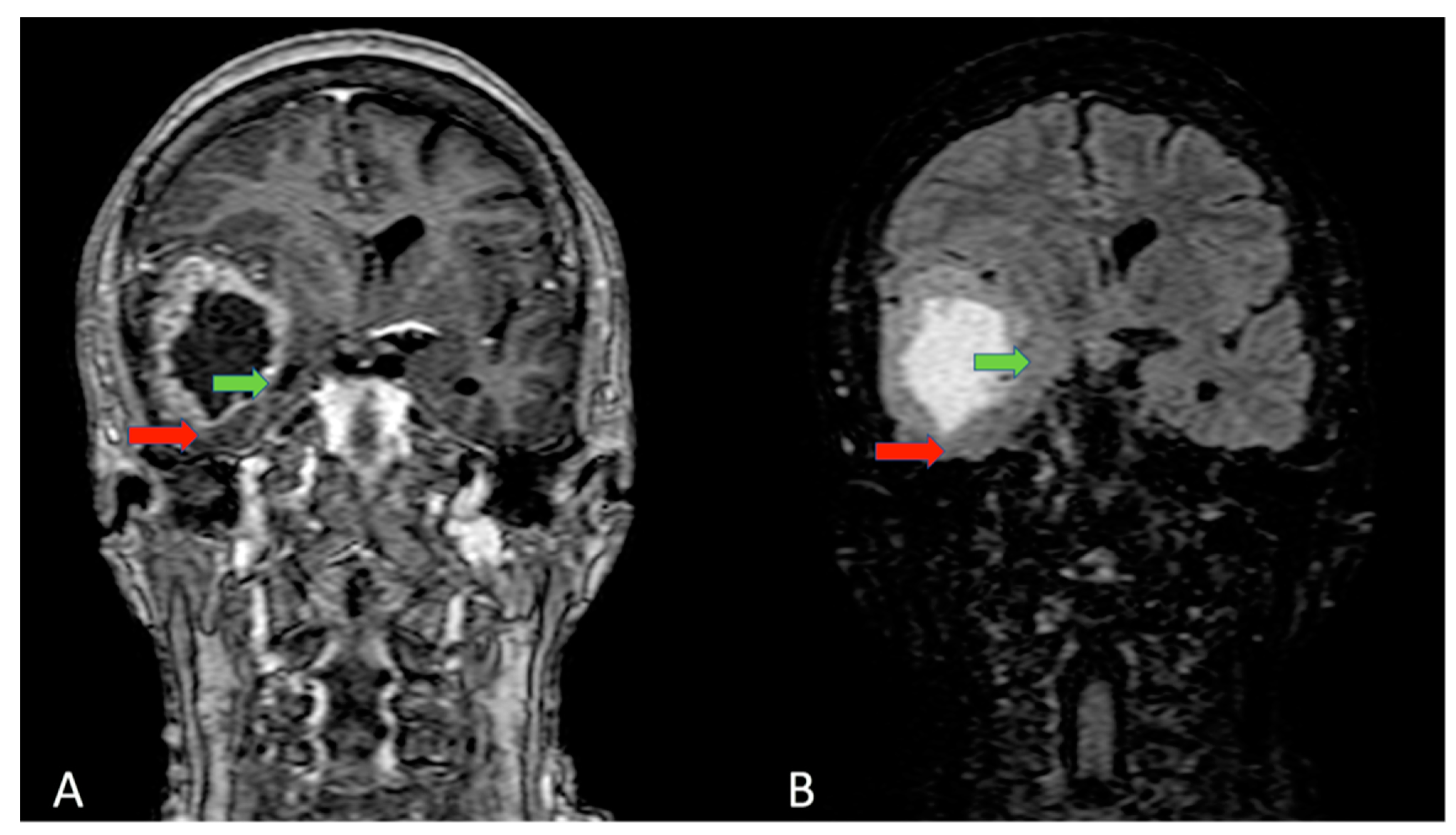{kind=link}
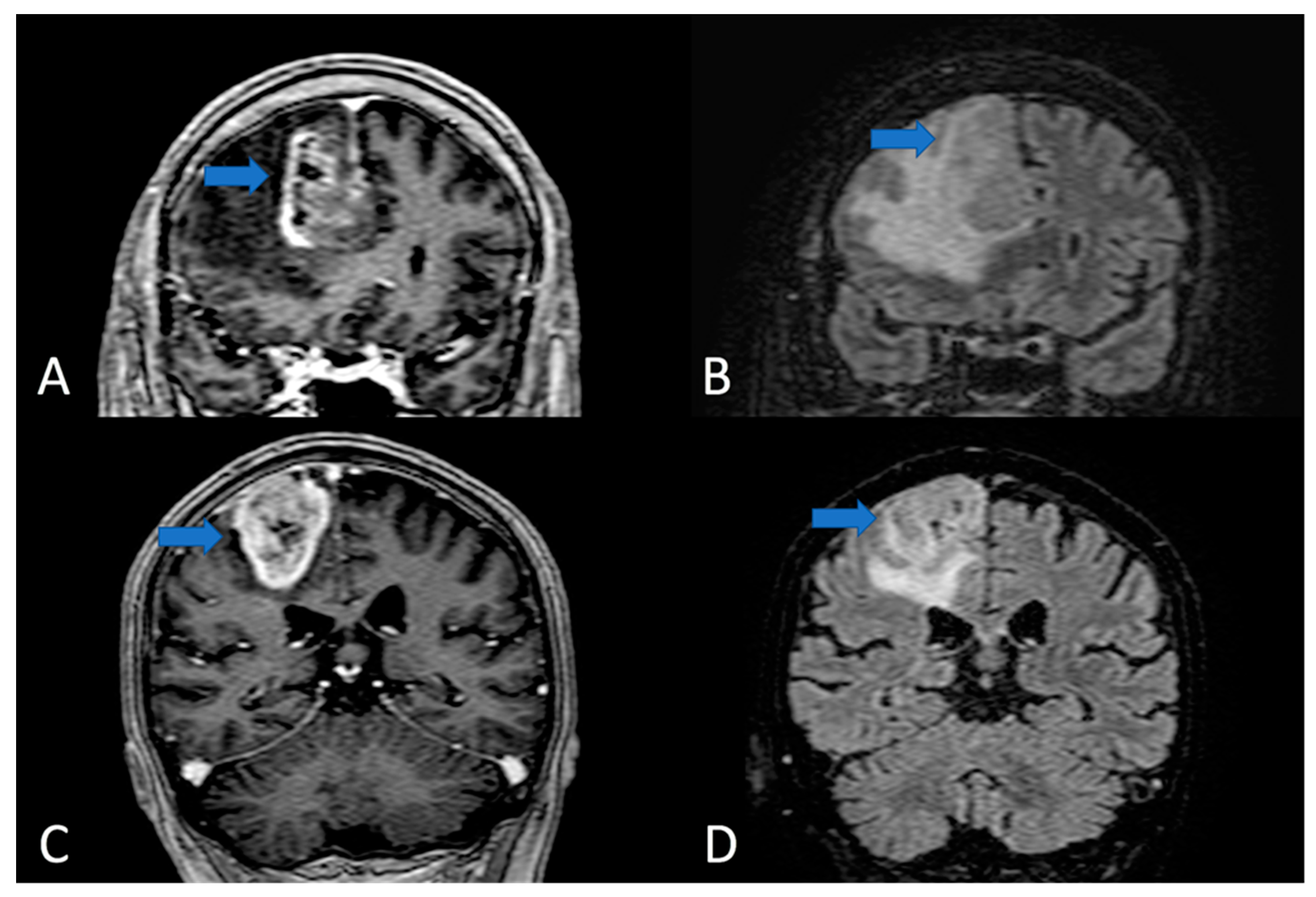{kind=link}
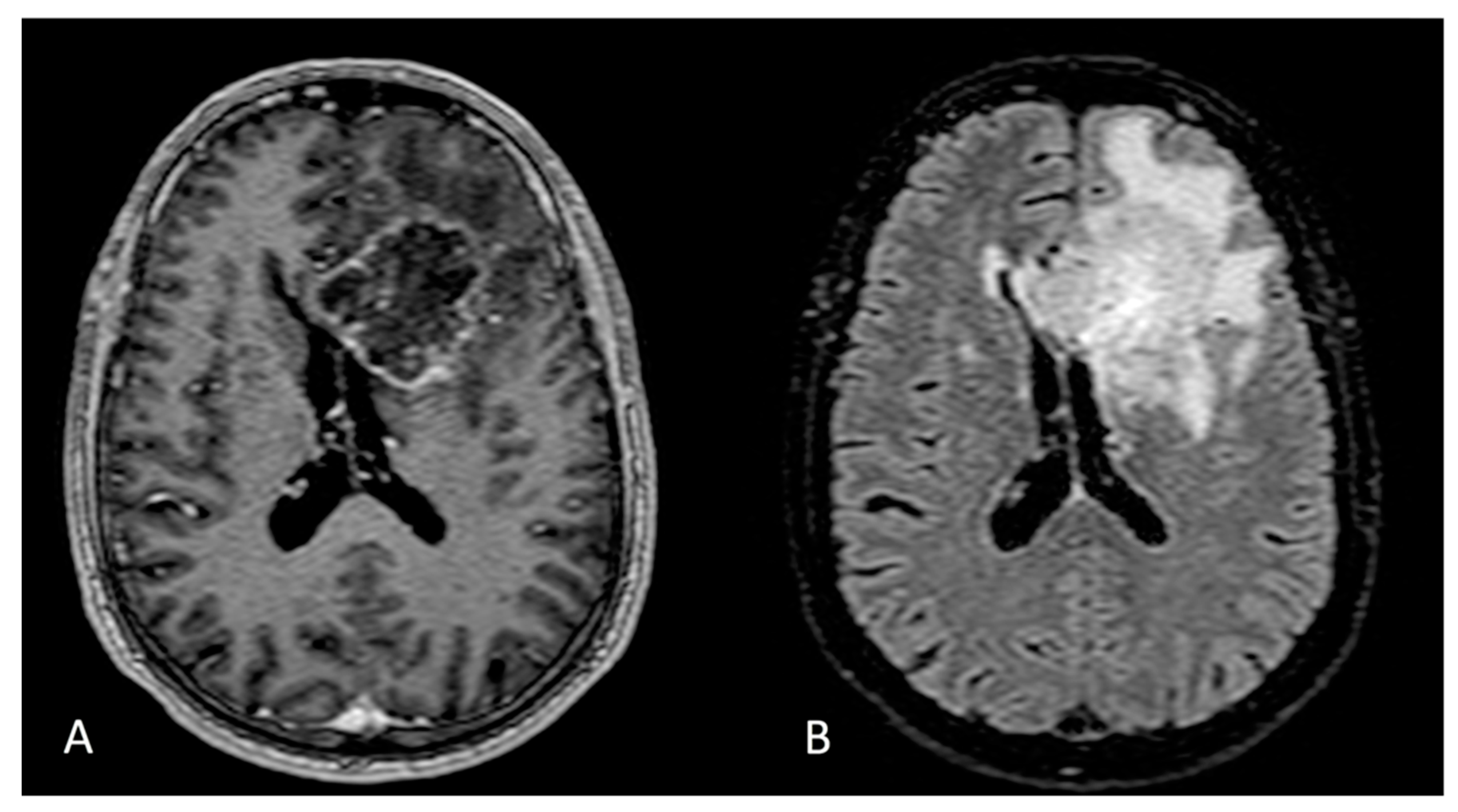{kind=link}
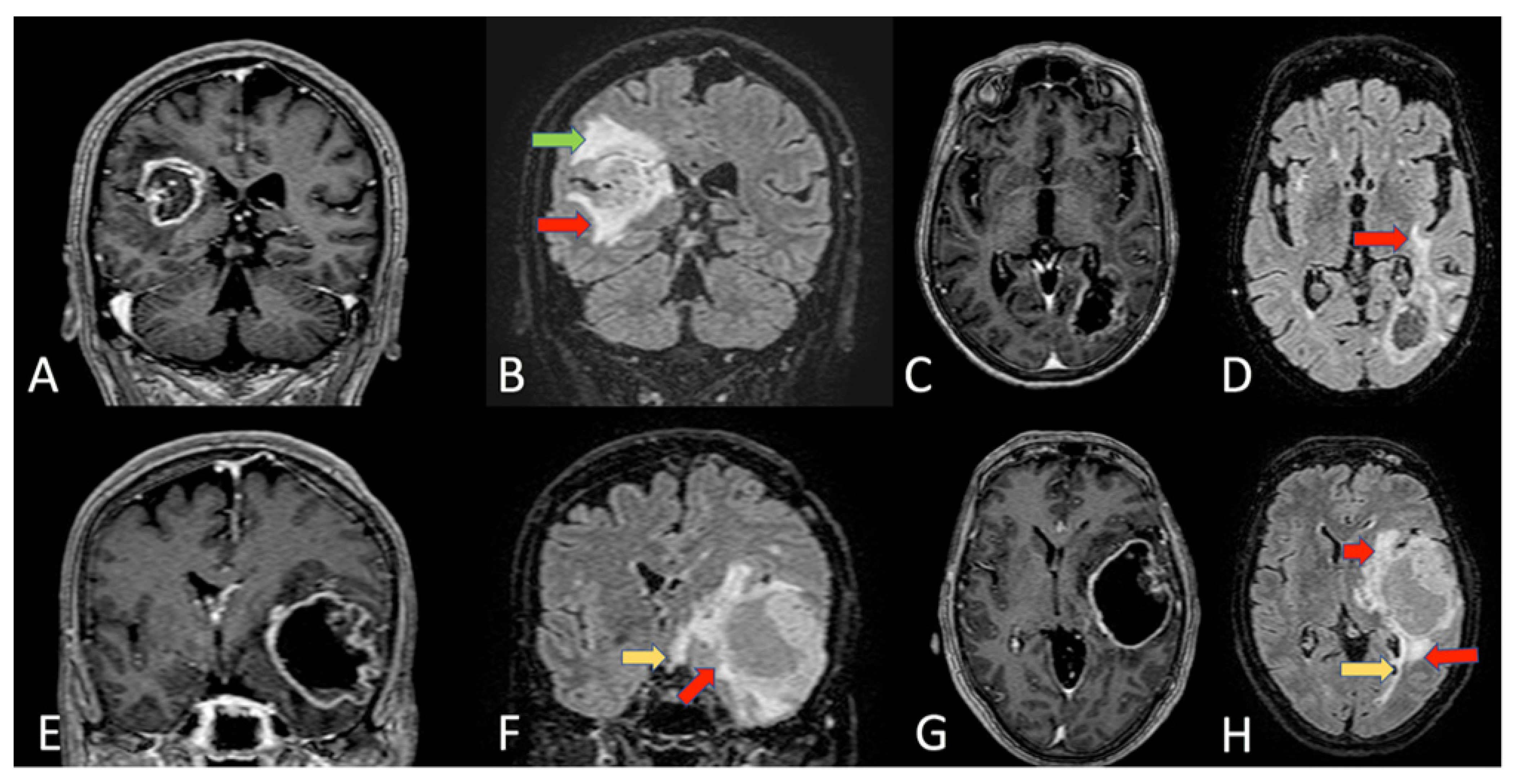{kind=link}
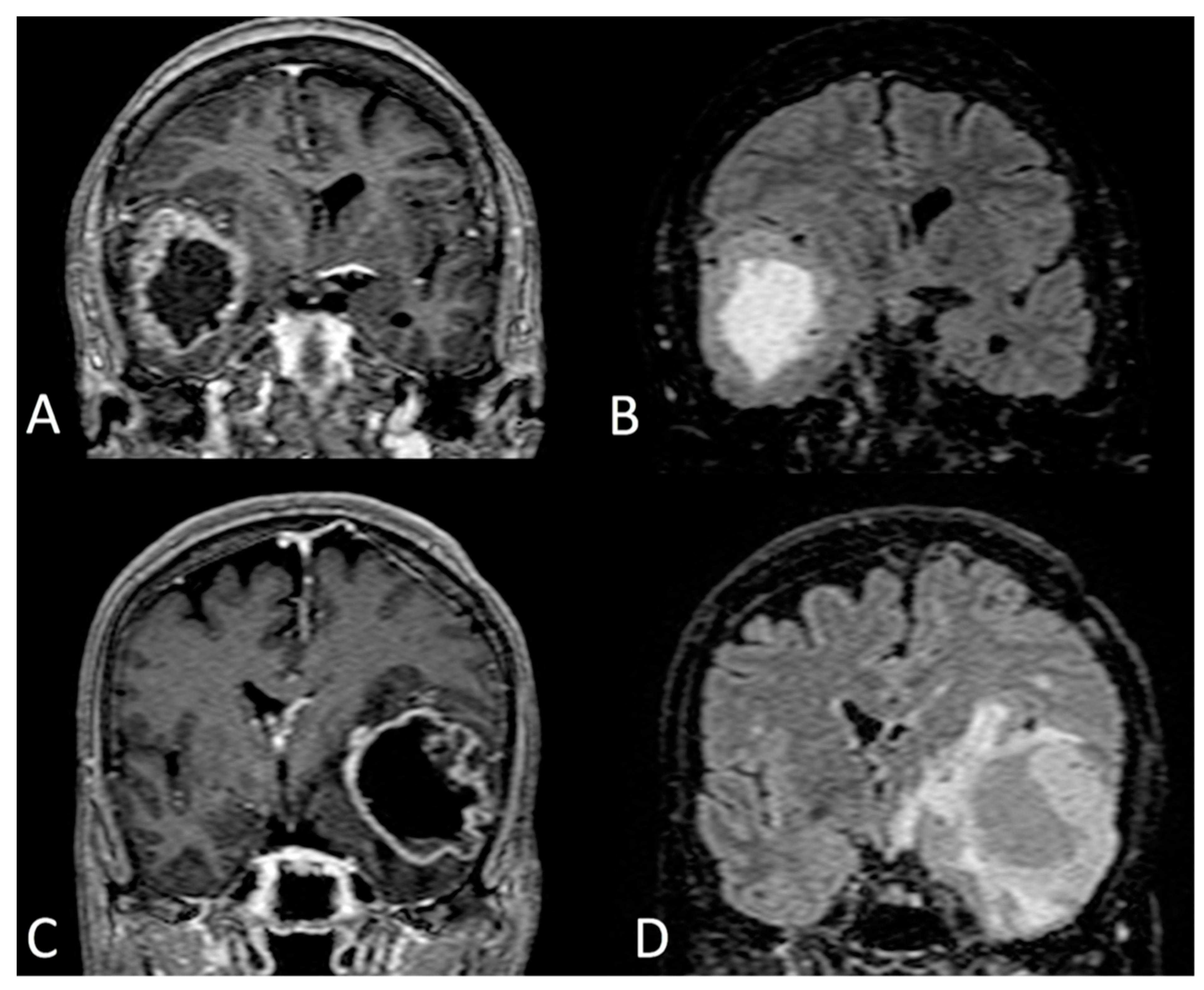{kind=link}
| Cellular and Extracellular Elements | Anatomical Localization | Putative Mechanism | Ref. | GBM Biological Signature |
|---|---|---|---|---|
| NSC | SVG, SGZ | Age-related decline of the tumor-suppressor BMP7 | [24,28,29,30] | GBM growth |
| NSC or progenitor cells | Subcortical white matter | Reactivation of migratory genes (EGF, Lck) | [35,92] | GBM invasiveness |
| Human ectodermic progenitors | Cortex | Differentially expressed genes (i.e., NEUROD6, ID2, LMO4) | [89,90] | GBM lateralization |
| Developing Astrocytes (B1 cells) | SVZ, Cortex | EGF overexpression | [42,43] | GBM invasiveness |
| Astrocytic subpopulations | Thalamus, Cortex, Brainstem | Mesenchymal signature (cluster B); Epilepsy-associated genes enrichment (cluster C) | [49] | GBM invasiveness |
| Astrocytes | Cerebellum, primary visual and dorsal prefrontal cortices | GLAST/GLT-1 overexpression | [16,63,64,93] | GBM growth (spared regions) |
| Neurons | Cerebellum | PD-L1 overexpression. Neurostatin release. | [59,60] | GBM growth (spared regions) |
| Oligodendrocytes | White matter | Nogo, semaphorin, ephrins downregulation | [3,52] | GBM invasiveness |
| Extracellular matrix | Hippocampal inlet, amygdala, and hypothalamus | Particular asset of aggrecan expression in contrast to Tenascin-R. | [69,79] | GBM growth (spared regions) |
| GSC | Left temporal lobe | MGMT methylated promoter; EGFR amplification | [16,86] | Bulky phenotype (short overall survivor) |
| GSC | Right temporal lobe | MGMT unmethylated promoter; IDH1 WT; Mesenchymal signature | [16] | Diffusive phenotype (short overall survival) |
| GSC | Frontal lobe | Focal PTEN loss; IDH1R132mut;p53mut; Focal EGFR amplification; Proneural signature | [16,87] | Diffusive phenotype |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, C.; Virtuoso, A.; Papa, M.; Certo, F.; Barbagallo, G.M.V.; Altieri, R. Regional Development of Glioblastoma: The Anatomical Conundrum of Cancer Biology and Its Surgical Implication. Cells 2022, 11, 1349. https://doi.org/10.3390/cells11081349
De Luca C, Virtuoso A, Papa M, Certo F, Barbagallo GMV, Altieri R. Regional Development of Glioblastoma: The Anatomical Conundrum of Cancer Biology and Its Surgical Implication. Cells. 2022; 11(8):1349. https://doi.org/10.3390/cells11081349
Chicago/Turabian StyleDe Luca, Ciro, Assunta Virtuoso, Michele Papa, Francesco Certo, Giuseppe Maria Vincenzo Barbagallo, and Roberto Altieri. 2022. "Regional Development of Glioblastoma: The Anatomical Conundrum of Cancer Biology and Its Surgical Implication" Cells 11, no. 8: 1349. https://doi.org/10.3390/cells11081349