
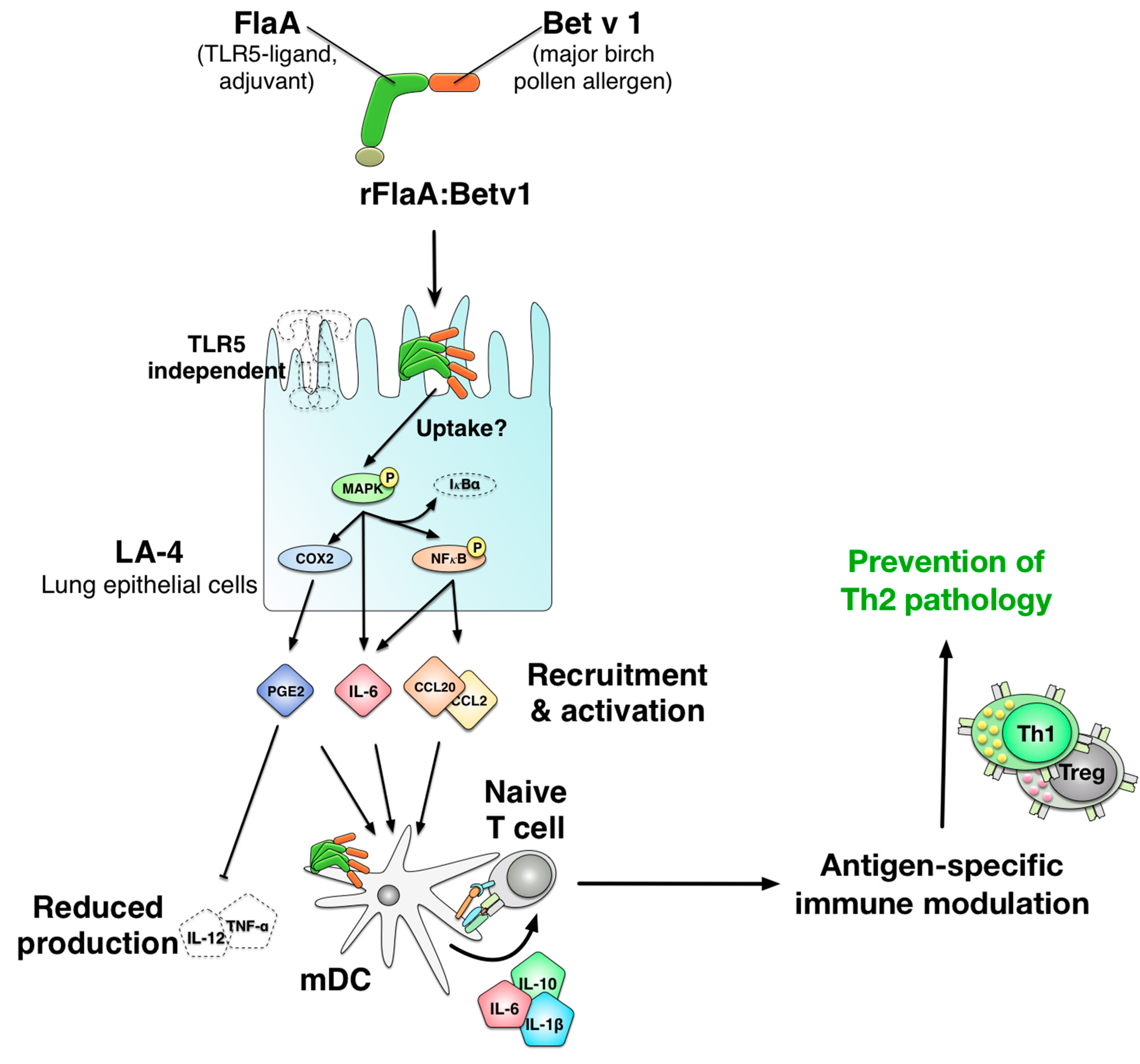
The Fusion Protein rFlaA:Betv1 Modulates DC Responses by a p38-MAPK and COX2-Dependent Secretion of PGE2 from Epithelial Cells

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Flow Cytometry
2.2. Generation of Recombinant Proteins
2.3. Culture, Stimulation, Viability of LA-4 Cells
2.4. ELISA
2.5. Inhibitors
2.6. Western Blot
2.7. Fluorescence Labeling, Uptake, and Microscopy
2.8. Mice
2.9. Preparation of LA-4:DC Co-Cultures
2.10. Statistical Analysis
3. Results
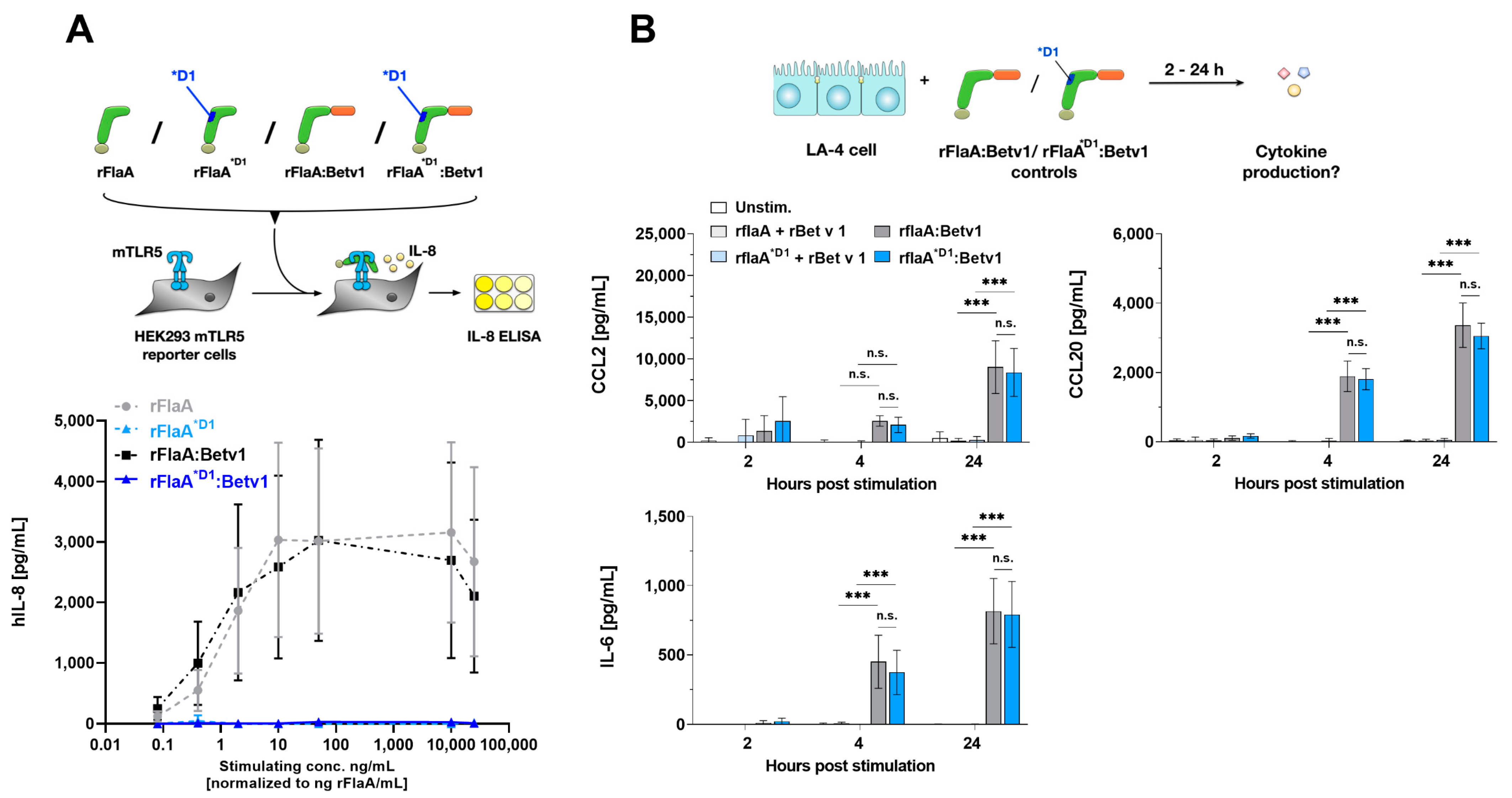
3.1. rFlaA:Betv1 Induces Chemokine and IL-6 Secretion from LA-4 Epithelial Cells
3.2. rFlaA:Betv1-Mediated LA-4 Cell Activation Is TLR5-Independent
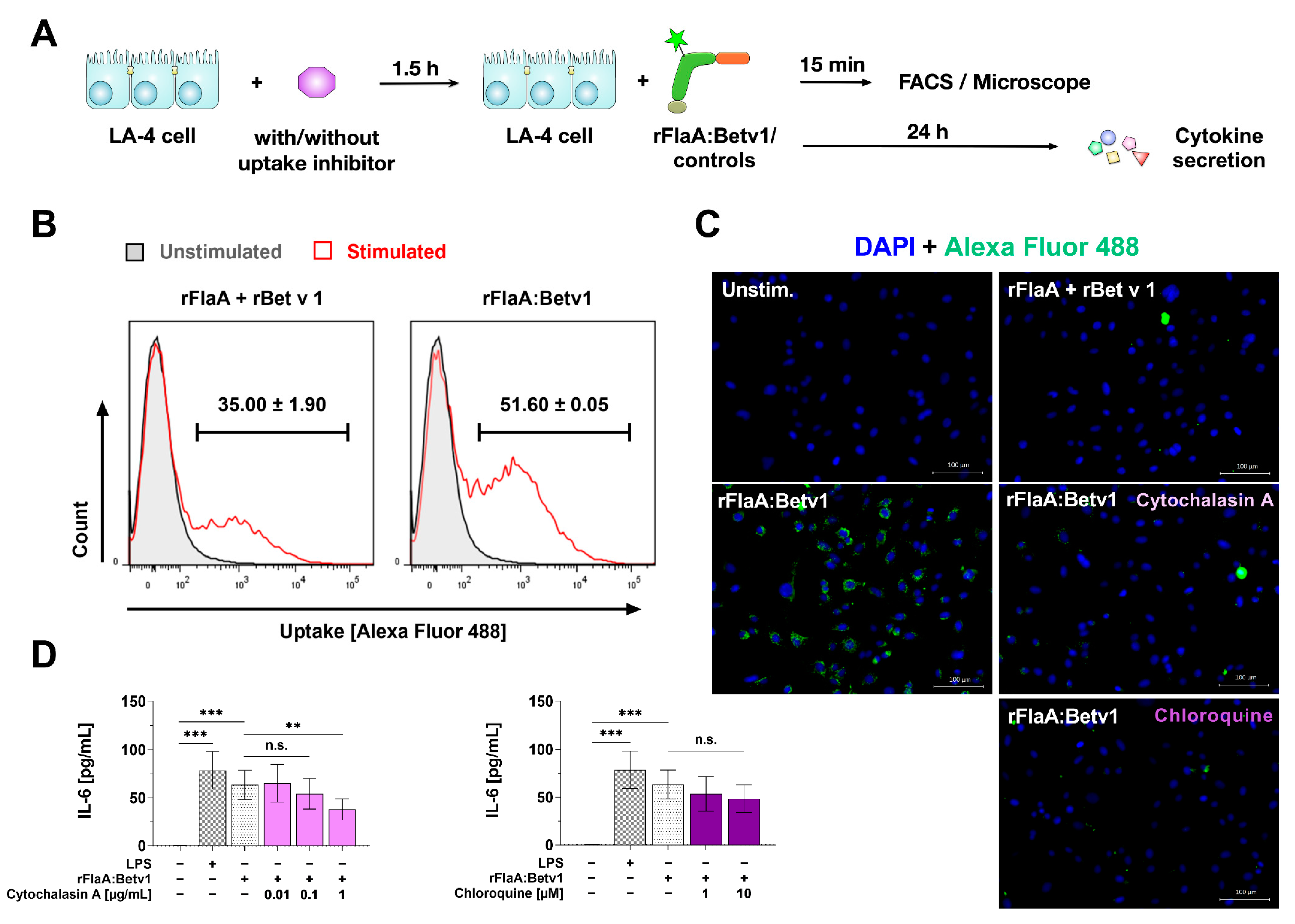
3.3. rFlaA:Betv1 Is Taken up More Strongly by LA-4 Cells
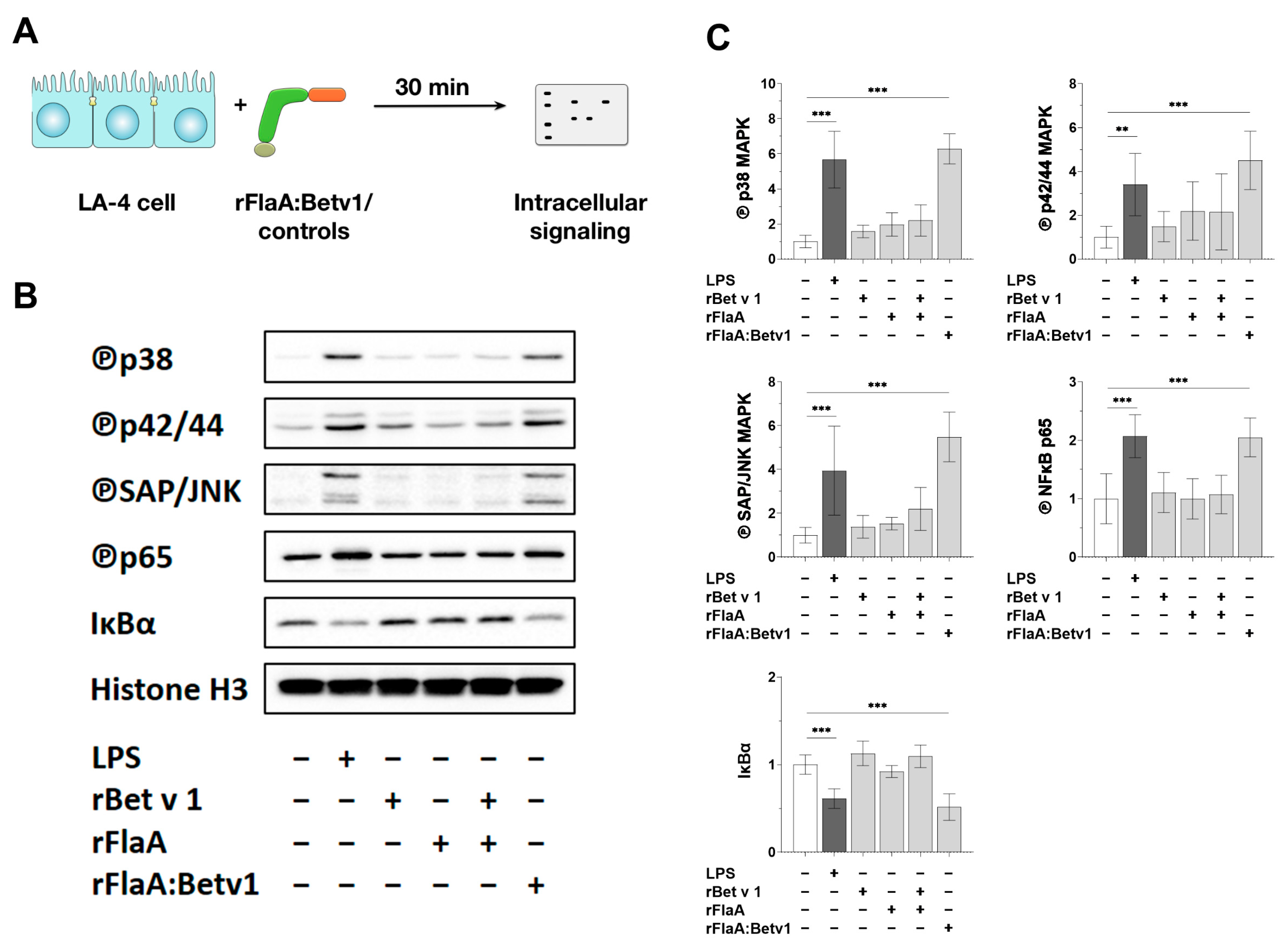
3.4. rFlaA:Betv1 Induces MAPK- and NFκB-Phosphorylation in LA-4 Epithelial Cells
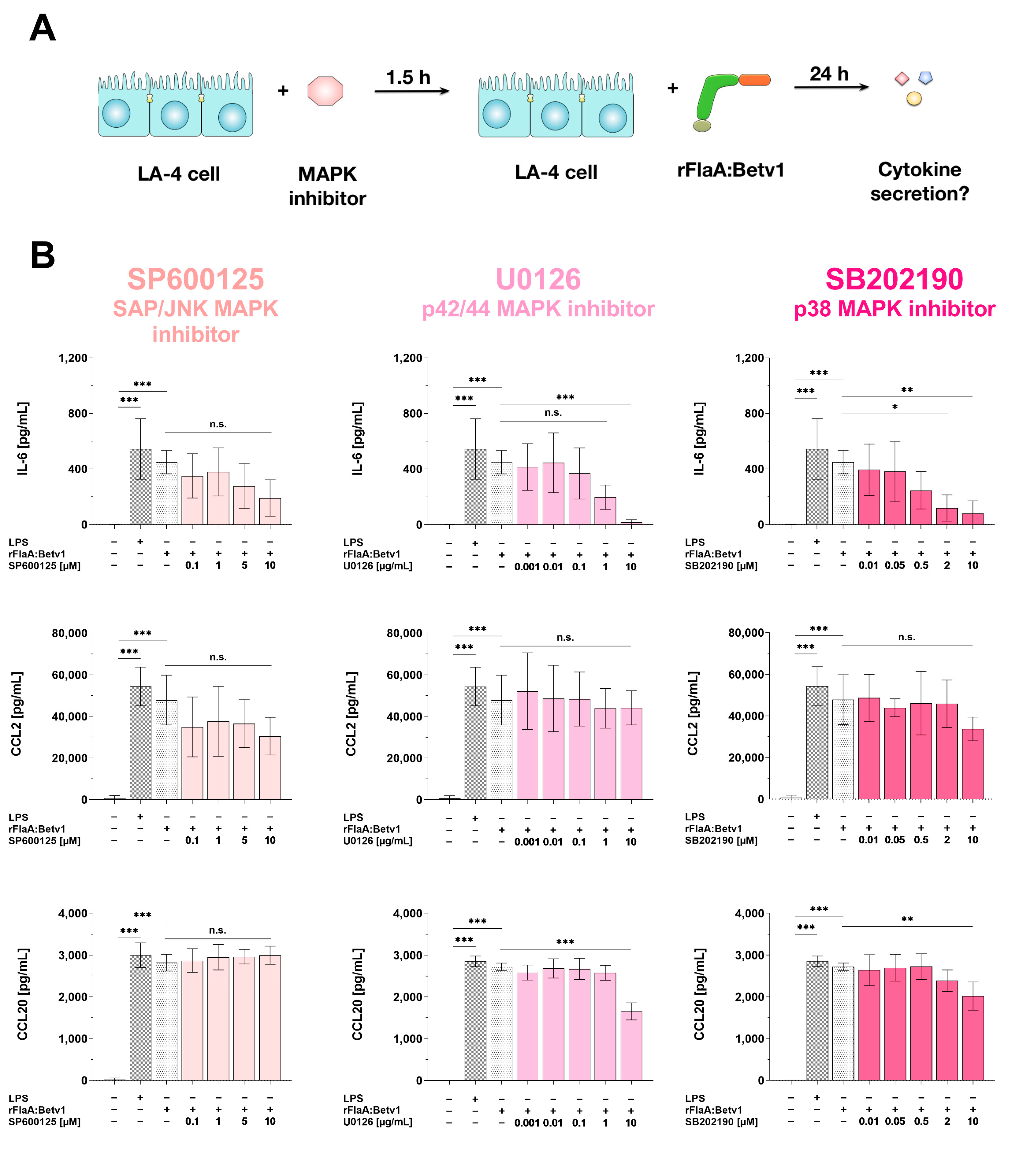
3.5. MAP Kinase-Signaling Contributes to Both rFlaA:Betv1-Induced Pro-Inflammatory IL-6 and CCL20 Secretion from LA-4 Cells
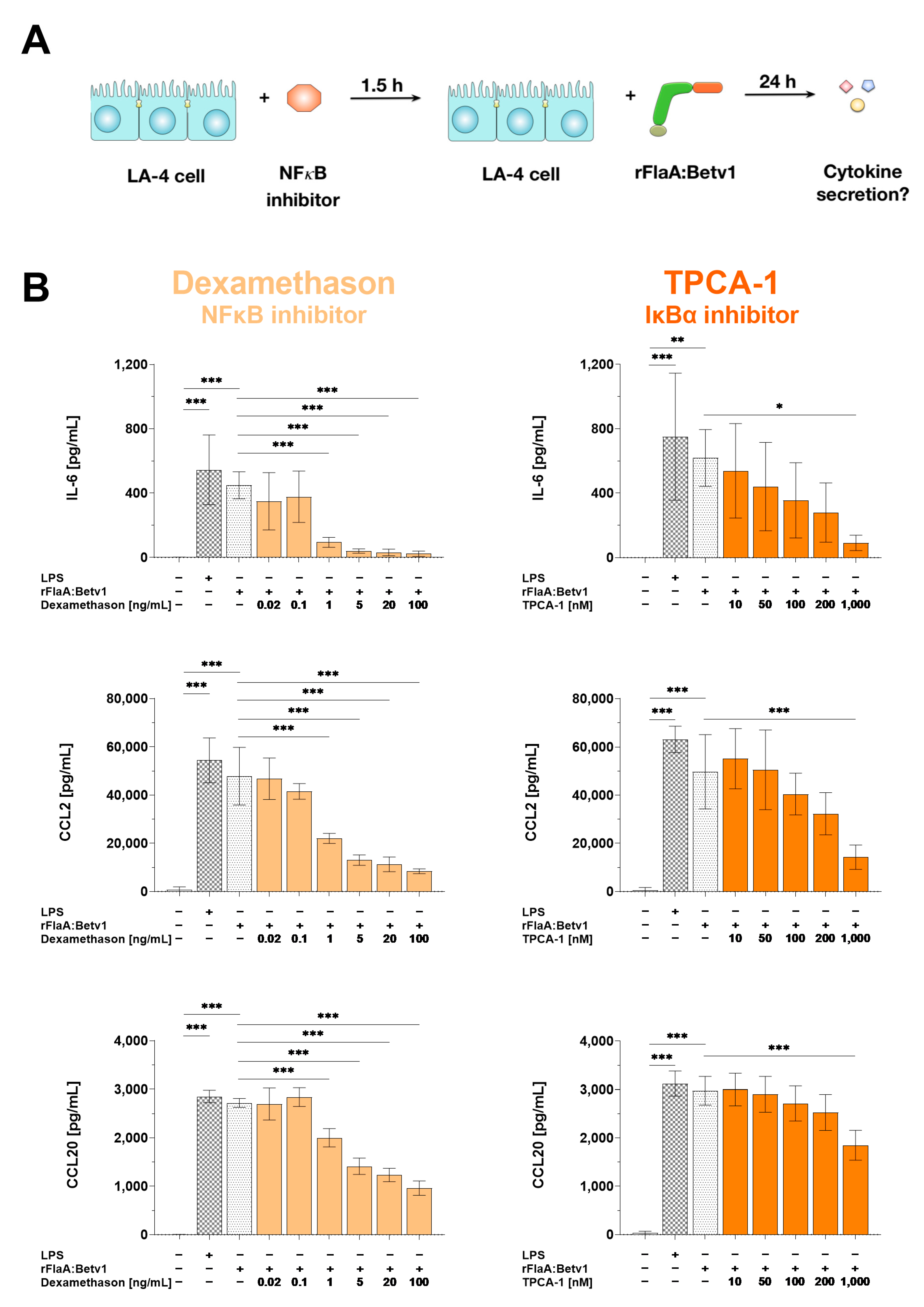
3.6. NFκB-Signaling Contributes to Both rFlaA:Betv1-Induced Pro-Inflammatory IL-6 and CCL2/CCL20 Chemokine Secretion in LA-4 Cells
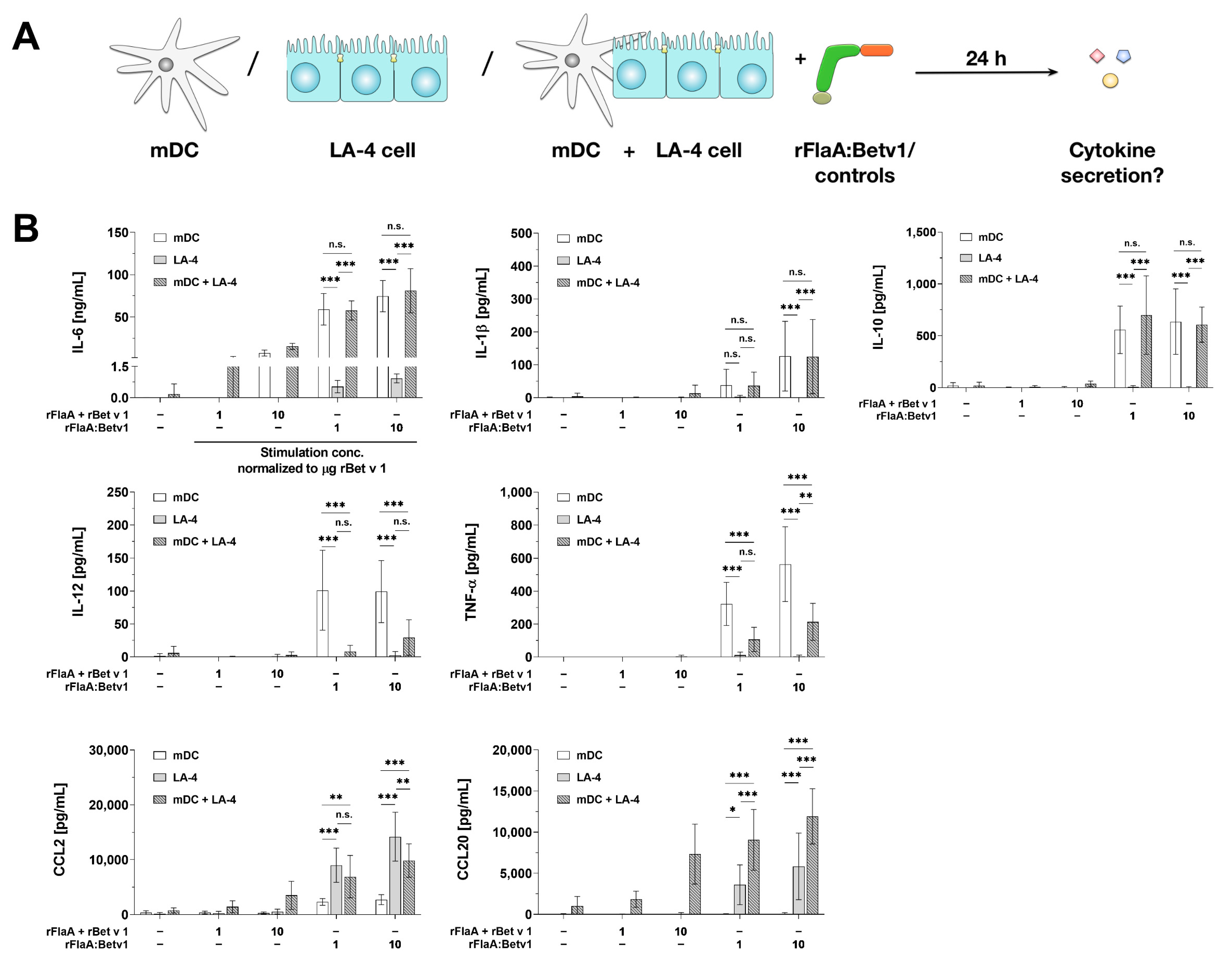
3.7. In Co-Cultures with BALB/c mDCs LA-4 Cells Modulate mDC-Derived, rFlaA:Betv1-Induced Cytokine Secretion
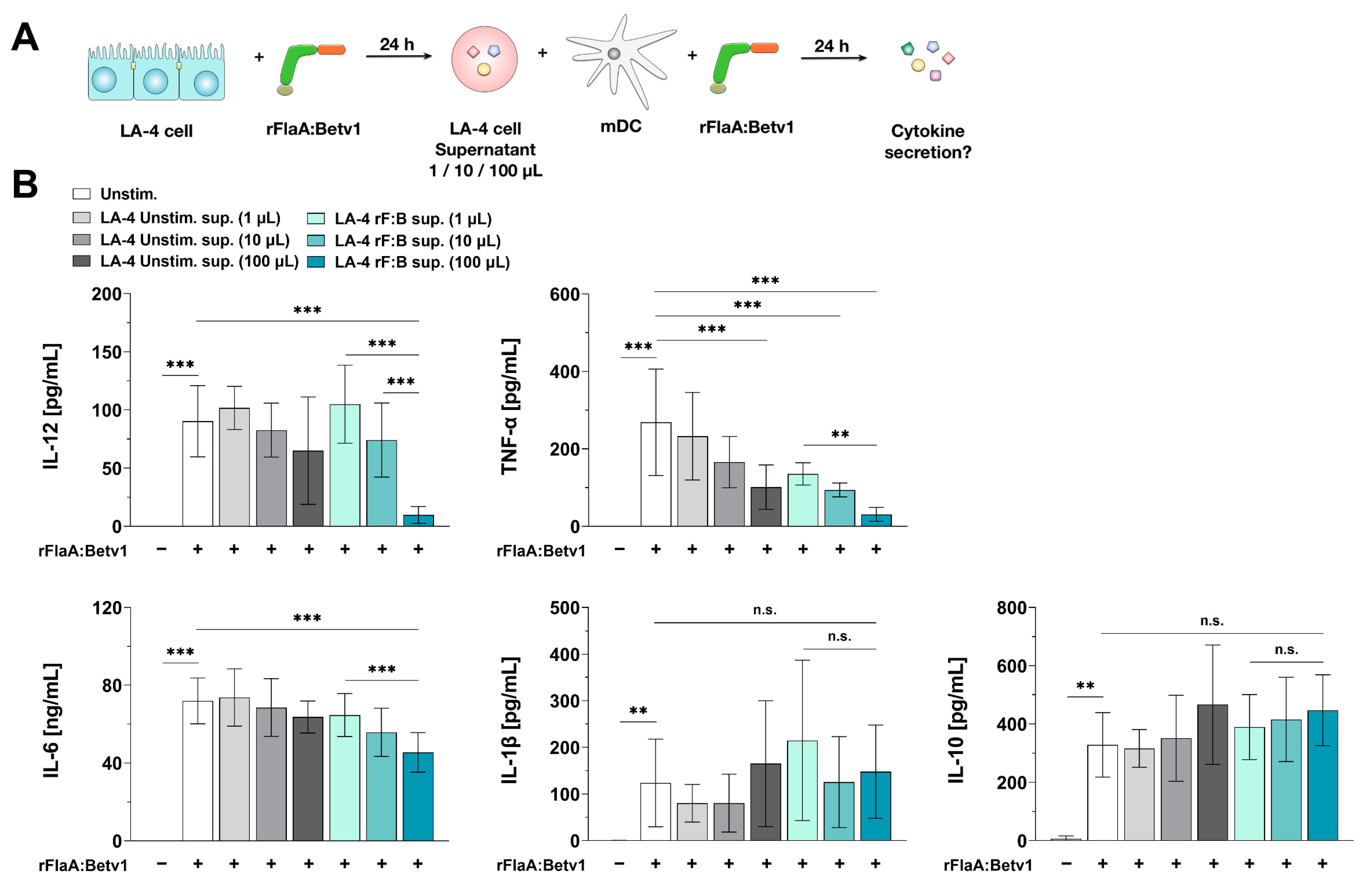
3.8. Epithelial Cell-Derived Soluble Factors Modulate mDC-Responses to rFlaA:Betv1
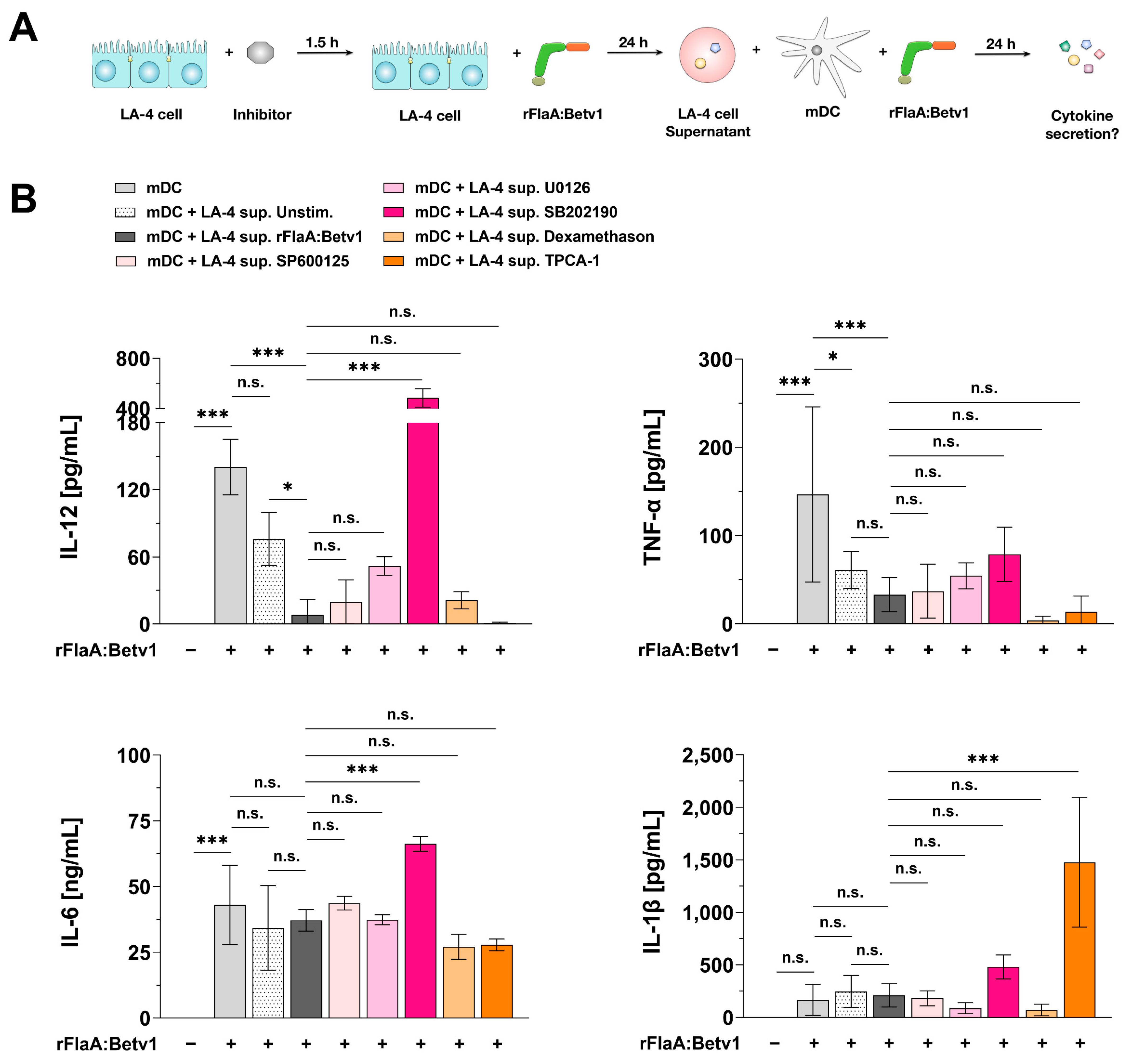
3.9. The Modulation of mDC Responses by rFlaA:Betv1-Stimulated LA-4 Cells Is Dependent on the p38-MAPK-Signaling Pathway
3.10. Modulation of mDC Responses by rFlaA:Betv1-Stimulated LA-4 Cells Is Dependent on the COX2/PGE2 Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| cDC2s | Type 2 conventional dendritic cells |
| COX2 | Cyclooxygenase-2 |
| FliC | Flagellin type C |
| ILC2s | Group 2 innate lymphoid cells |
| MAPK | Mitogen-activated protein kinase |
| mDCs | Myeloid dendritic cells |
| MPLA | Monophosphoryl lipid A |
| mTOR | Mammalian target of rapamycin |
| NLRC4 | NOD-like receptor 4 |
| PAMPs | Pathogen-associated molecular patterns |
| PGE2 | Prostaglandin E2 |
| PRRs | Pattern recognition receptors |
| rBetv1 | Recombinant birch pollen allergen Betv1 |
| rFlaA | Recombinant flagellin A from Listeria monocytogenes |
| rFlaA:Betv1 | Fusion protein consisting of flagellin A from Listeria monocytogenes and the major birch pollen allergen Betv1 |
| RLU | Relative light unit |
| TLR | Toll-like receptor |
References
- Kundi, M. New Hepatitis B Vaccine Formulated with an Improved Adjuvant System. Expert Rev. Vaccines 2007, 6, 133–140. [Google Scholar] [CrossRef]
- Agnandji, S.T.; Lell, B.; Soulanoudjingar, S.S.; Fernandes, J.F.; Abossolo, B.P.; Conzelmann, C.; Methogo, B.G.N.O.; Doucka, Y.; Flamen, A.; Mordmüller, B.; et al. First Results of Phase 3 Trial of RTS,S/AS01 Malaria Vaccine in African Children. N. Engl. J. Med. 2011, 365, 1863–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GlaxoSmithKline Vaccine HPV-007 Study Group; Romanowski, B.; de Borba, P.C.; Naud, P.S.; Roteli-Martins, C.M.; De Carvalho, N.S.; Teixeira, J.C.; Aoki, F.; Ramjattan, B.; Shier, R.M.; et al. Sustained Efficacy and Immunogenicity of the Human Papillomavirus (HPV)-16/18 AS04-Adjuvanted Vaccine: Analysis of a Randomised Placebo-Controlled Trial up to 6.4 Years. Lancet 2009, 374, 1975–1985. [Google Scholar] [CrossRef]
- Toussi, D.; Massari, P. Immune Adjuvant Effect of Molecularly-Defined Toll-Like Receptor Ligands. Vaccines 2014, 2, 323–353. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The Innate Immune Response to Bacterial Flagellin Is Mediated by Toll-like Receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Amer, A.; Body-Malapel, M.; Kanneganti, T.-D.; Ozören, N.; Jagirdar, R.; Inohara, N.; Vandenabeele, P.; Bertin, J.; Coyle, A.; et al. Cytosolic Flagellin Requires Ipaf for Activation of Caspase-1 and Interleukin 1beta in Salmonella-Infected Macrophages. Nat. Immunol. 2006, 7, 576–582. [Google Scholar] [CrossRef]
- Lee, S.E.; Kim, S.Y.; Jeong, B.C.; Kim, Y.R.; Bae, S.J.; Ahn, O.S.; Lee, J.; Song, H.; Kim, J.M.; Choy, H.E.; et al. A Bacterial Flagellin, Vibrio Vulnificus FlaB, Has a Strong Mucosal Adjuvant Activity to Induce Protective Immunity. Infect. Immun. 2006, 74, 694–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honko, A.N.; Sriranganathan, N.; Lees, C.J.; Mizel, S.B. Flagellin Is an Effective Adjuvant for Immunization against Lethal Respiratory Challenge with Yersinia Pestis. Infect. Immun. 2006, 74, 1113–1120. [Google Scholar] [CrossRef] [Green Version]
- Huleatt, J.W.; Jacobs, A.R.; Tang, J.; Desai, P.; Kopp, E.B.; Huang, Y.; Song, L.; Nakaar, V.; Powell, T.J. Vaccination with Recombinant Fusion Proteins Incorporating Toll-like Receptor Ligands Induces Rapid Cellular and Humoral Immunity. Vaccine 2007, 25, 763–775. [Google Scholar] [CrossRef]
- Song, L.; Xiong, D.; Kang, X.; Yang, Y.; Wang, J.; Guo, Y.; Xu, H.; Chen, S.; Peng, D.; Pan, Z.; et al. An Avian Influenza A (H7N9) Virus Vaccine Candidate Based on the Fusion Protein of Hemagglutinin Globular Head and Salmonella Typhimurium Flagellin. BMC Biotechnol. 2015, 15, 79. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, L.A.; Kotlyarov, R.Y.; Kovaleva, A.A.; Potapchuk, M.V.; Korotkov, A.V.; Sergeeva, M.V.; Kasianenko, M.A.; Kuprianov, V.V.; Ravin, N.V.; Tsybalova, L.M.; et al. Protection against Multiple Influenza A Virus Strains Induced by Candidate Recombinant Vaccine Based on Heterologous M2e Peptides Linked to Flagellin. PLoS ONE 2015, 10, e0119520. [Google Scholar] [CrossRef]
- Wang, B.-Z.; Gill, H.S.; He, C.; Ou, C.; Wang, L.; Wang, Y.-C.; Feng, H.; Zhang, H.; Prausnitz, M.R.; Compans, R.W. Microneedle Delivery of an M2e-TLR5 Ligand Fusion Protein to Skin Confers Broadly Cross-Protective Influenza Immunity. J. Control. Release 2014, 178, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, K.N.; Phipps, J.P.; Johnson, J.B.; Mizel, S.B. A Recombinant Flagellin-Poxvirus Fusion Protein Vaccine Elicits Complement-Dependent Protection against Respiratory Challenge with Vaccinia Virus in Mice. Viral Immunol. 2010, 23, 201–210. [Google Scholar] [CrossRef] [PubMed]
- McDonald, W.F.; Huleatt, J.W.; Foellmer, H.G.; Hewitt, D.; Tang, J.; Desai, P.; Price, A.; Jacobs, A.; Takahashi, V.N.; Huang, Y.; et al. A West Nile Virus Recombinant Protein Vaccine That Coactivates Innate and Adaptive Immunity. J. Infect. Dis. 2007, 195, 1607–1617. [Google Scholar] [CrossRef]
- Lee, S.E.; Nguyen, C.T.; Kim, S.Y.; Thi, T.N.; Rhee, J.H. Tetanus Toxin Fragment C Fused to Flagellin Makes a Potent Mucosal Vaccine. Clin. Exp. Vaccine Res. 2015, 4, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weimer, E.T.; Lu, H.; Kock, N.D.; Wozniak, D.J.; Mizel, S.B. A Fusion Protein Vaccine Containing OprF Epitope 8, OprI, and Type A and B Flagellins Promotes Enhanced Clearance of Nonmucoid Pseudomonas Aeruginosa. Infect. Immun. 2009, 77, 2356–2366. [Google Scholar] [CrossRef] [Green Version]
- Kitzmüller, C.; Kalser, J.; Mutschlechner, S.; Hauser, M.; Zlabinger, G.J.; Ferreira, F.; Bohle, B. Fusion Proteins of Flagellin and the Major Birch Pollen Allergen Betv1 Show Enhanced Immunogenicity, Reduced Allergenicity, and Intrinsic Adjuvanticity. J. Allergy Clin. Immunol. 2018, 141, 293–299.e6. [Google Scholar] [CrossRef] [Green Version]
- Schülke, S.; Burggraf, M.; Waibler, Z.; Wangorsch, A.; Wolfheimer, S.; Kalinke, U.; Vieths, S.; Toda, M.; Scheurer, S. A Fusion Protein of Flagellin and Ovalbumin Suppresses the TH2 Response and Prevents Murine Intestinal Allergy. J. Allergy Clin. Immunol. 2011, 128, 1340–1348.e12. [Google Scholar] [CrossRef]
- Schülke, S.; Waibler, Z.; Mende, M.-S.; Zoccatelli, G.; Vieths, S.; Toda, M.; Scheurer, S. Fusion Protein of TLR5-Ligand and Allergen Potentiates Activation and IL-10 Secretion in Murine Myeloid DC. Mol. Immunol. 2010, 48, 341–350. [Google Scholar] [CrossRef]
- Schülke, S.; Wolfheimer, S.; Gadermaier, G.; Wangorsch, A.; Siebeneicher, S.; Briza, P.; Spreitzer, I.; Schiller, D.; Loeschner, B.; Uematsu, S.; et al. Prevention of Intestinal Allergy in Mice by RflaA:Ova Is Associated with Enforced Antigen Processing and TLR5-Dependent IL-10 Secretion by MDC. PLoS ONE 2014, 9, e87822. [Google Scholar] [CrossRef]
- Stefan, S.; Kirsten, K.; Sonja, W.; Nadine, D.; Andrea, W.; Andreas, R.; Peter, B.; Isabel, P.; Gabriele, G.; Fatima, F.; et al. Conjugation of Wildtype and Hypoallergenic Mugwort Allergen Art v 1 to Flagellin Induces IL-10-DC and Suppresses Allergen-Specific TH2-Responses in Vivo. Sci. Rep. 2017, 7, 11782. [Google Scholar] [CrossRef] [Green Version]
- Schülke, S.; Fiedler, A.-H.; Junker, A.-C.; Flaczyk, A.; Wolfheimer, S.; Wangorsch, A.; Heinz, A.; Beckert, H.; Nagl, B.; Bohle, B.; et al. Critical Role of Mammalian Target of Rapamycin for IL-10 Dendritic Cell Induction by a Flagellin a Conjugate in Preventing Allergic Sensitization. J. Allergy Clin. Immunol. 2018, 141, 1786–1798.e11. [Google Scholar] [CrossRef] [Green Version]
- Moeller, T.; Wolfheimer, S.; Goretzki, A.; Scheurer, S.; Schülke, S. NFκB- and MAP-Kinase Signaling Contribute to the Activation of Murine Myeloid Dendritic Cells by a Flagellin A:Allergen Fusion Protein. Cells 2019, 8, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turley, C.B.; Rupp, R.E.; Johnson, C.; Taylor, D.N.; Wolfson, J.; Tussey, L.; Kavita, U.; Stanberry, L.; Shaw, A. Safety and Immunogenicity of a Recombinant M2e-Flagellin Influenza Vaccine (STF2.4xM2e) in Healthy Adults. Vaccine 2011, 29, 5145–5152. [Google Scholar] [CrossRef]
- Treanor, J.J.; Taylor, D.N.; Tussey, L.; Hay, C.; Nolan, C.; Fitzgerald, T.; Liu, G.; Kavita, U.; Song, L.; Dark, I.; et al. Safety and Immunogenicity of a Recombinant Hemagglutinin Influenza-Flagellin Fusion Vaccine (VAX125) in Healthy Young Adults. Vaccine 2010, 28, 8268–8274. [Google Scholar] [CrossRef]
- Khan, S.; Bijker, M.S.; Weterings, J.J.; Tanke, H.J.; Adema, G.J.; van Hall, T.; Drijfhout, J.W.; Melief, C.J.M.; Overkleeft, H.S.; van der Marel, G.A.; et al. Distinct Uptake Mechanisms but Similar Intracellular Processing of Two Different Toll-like Receptor Ligand-Peptide Conjugates in Dendritic Cells. J. Biol. Chem. 2007, 282, 21145–21159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, A.J.; Huffnagle, G.B. The Microbiome and Regulation of Mucosal Immunity. Immunology 2014, 142, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Hammad, H.; Lambrecht, B.N. Barrier Epithelial Cells and the Control of Type 2 Immunity. Immunity 2015, 43, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, R.; Takeda, K. Roles of Intestinal Epithelial Cells in the Maintenance of Gut Homeostasis. Exp. Mol. Med. 2017, 49, e338. [Google Scholar] [CrossRef] [Green Version]
- Roan, F.; Obata-Ninomiya, K.; Ziegler, S.F. Epithelial Cell-Derived Cytokines: More than just Signaling the Alarm. J. Clin. Investig. 2019, 129, 1441–1451. [Google Scholar] [CrossRef] [Green Version]
- Maffía, P.C.; Guerrieri, D.; Villalonga, X.; Caro, F.; Gómez, S.; Tateosian, N.; Bogado, B.P.; Sánchez, M.L.; Ambrosi, N.; Chuluyan, E. Cementoin-SLPI Fusion Protein Binds to Human Monocytes and Epithelial Cells and Shows Higher Biological Activity than SLPI. Sci. Rep. 2018, 8, 5332. [Google Scholar] [CrossRef] [Green Version]
- Savar, N.S.; Jahanian-Najafabadi, A.; Mahdavi, M.; Shokrgozar, M.A.; Jafari, A.; Bouzari, S. In Silico and In Vivo Studies of Truncated Forms of Flagellin (FliC) of Enteroaggregative Escherichia coli Fused to FimH from Uropathogenic Escherichia coli as a Vaccine Candidate against Urinary Tract Infections. J. Biotechnol. 2014, 175, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Siebeneicher, S.; Reuter, S.; Krause, M.; Wangorsch, A.; Maxeiner, J.; Wolfheimer, S.; Schülke, S.; Naito, S.; Heinz, A.; Taube, C.; et al. Epicutaneous Immune Modulation with Betv1 plus R848 Suppresses Allergic Asthma in a Murine Model. Allergy 2014, 69, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Andersen-Nissen, E.; Smith, K.D.; Strobe, K.L.; Barrett, S.L.R.; Cookson, B.T.; Logan, S.M.; Aderem, A.; Andersen-Nissen, E.; Smith, K.D.; Strobe, K.L.; et al. Evasion of Toll-like Receptor 5 by Flagellated Bacteria. Proc. Natl. Acad. Sci. USA 2005, 102, 9247–9252. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-J.; Papp, G.; Miskey, C.; Fiedler, A.; Goretzki, A.; Wolfheimer, S.; Zimmermann, J.; Crauwels, P.; Ivics, Z.; van Zandbergen, G.; et al. The Flagellin:Allergen Fusion Protein RFlaA:Betv1 Induces a MyD88- and MAPK-Dependent Activation of Glucose Metabolism in Macrophages. Cells 2021, 10, 2614. [Google Scholar] [CrossRef]
- Cha, Y.I.; Solnica-Krezel, L.; DuBois, R.N. Fishing for Prostanoids: Deciphering the Developmental Functions of Cyclooxygenase-Derived Prostaglandins. Dev. Biol. 2006, 289, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Kalinski, P. Regulation of Immune Responses by Prostaglandin E2. J. Immunol. 2012, 188, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Speth, J.M.; Bourdonnay, E.; Penke, L.R.K.; Mancuso, P.; Moore, B.B.; Weinberg, J.B.; Peters-Golden, M. Alveolar Epithelial Cell-Derived Prostaglandin E2 Serves as a Request Signal for Macrophage Secretion of Suppressor of Cytokine Signaling 3 during Innate Inflammation. J. Immunol. 2016, 196, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, L.M.; Belvisi, M.G.; Bode, K.A.; Bauer, J.; Schmidt, C.; Suchy, M.-T.; Tsikas, D.; Scheuerer, J.; Lasitschka, F.; Gröne, H.-J.; et al. Bronchial Epithelial Cell-Derived Prostaglandin E2 Dampens the Reactivity of Dendritic Cells. J. Immunol. 2011, 186, 2095–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- N’Guessan, P.D.; Hippenstiel, S.; Etouem, M.O.; Zahlten, J.; Beermann, W.; Lindner, D.; Opitz, B.; Witzenrath, M.; Rosseau, S.; Suttorp, N.; et al. Streptococcus Pneumoniae Induced P38 MAPK- and NF-KappaB-Dependent COX-2 Expression in Human Lung Epithelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L1131–L1138. [Google Scholar] [CrossRef] [Green Version]
- Vassiliou, E.; Jing, H.; Ganea, D. Prostaglandin E2 Inhibits TNF Production in Murine Bone Marrow-Derived Dendritic Cells. Cell. Immunol. 2003, 223, 120–132. [Google Scholar] [CrossRef]
- Hubbard, L.L.N.; Ballinger, M.N.; Thomas, P.E.; Wilke, C.A.; Standiford, T.J.; Kobayashi, K.S.; Flavell, R.A.; Moore, B.B. A Role for IL-1 Receptor-Associated Kinase-M in Prostaglandin E2-Induced Immunosuppression Post-Bone Marrow Transplantation. J. Immunol. 2010, 184, 6299–6308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, T.S.; Nataro, J.P.; Poteet-Smith, C.E.; Smith, J.A.; Guerrant, R.L. Enteroaggregative Escherichia Coli Expresses a Novel Flagellin That Causes IL-8 Release from Intestinal Epithelial Cells. J. Clin. Investig. 2000, 105, 1769–1777. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, D.; Sun, T. Flagellin of Pseudomonas Aeruginosa Induces Transforming Growth Factor Beta 1 Expression in Normal Bronchial Epithelial Cells through Mitogen Activated Protein Kinase Cascades. Chin. Med. J. 2011, 124, 599–605. [Google Scholar] [PubMed]
- Kondo, Y.; Higa-Nakamine, S.; Maeda, N.; Toku, S.; Kakinohana, M.; Sugahara, K.; Kukita, I.; Yamamoto, H. Stimulation of Cell Migration by Flagellin through the P38 MAP Kinase Pathway in Cultured Intestinal Epithelial Cells. J. Cell Biochem. 2016, 117, 247–258. [Google Scholar] [CrossRef]
- Iwasa, S.; Ota, H.; Nishio, K.; Ohtsu, M.; Kusunoki, M.; Gojoubori, T.; Shirakawa, T.; Asano, M. Functional Expression of TLR5 in Murine Salivary Gland Epithelial Cells. J. Oral Sci. 2016, 58, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, A.; Van Maele, L.; Fougeron, D.; Cayet, D.; Sirard, J.-C. The GM-CSF Released by Airway Epithelial Cells Orchestrates the Mucosal Adjuvant Activity of Flagellin. J. Immunol. 2020, 205, 2873–2882. [Google Scholar] [CrossRef]
- Ramirez-Moral, I.; Yu, X.; Butler, J.M.; van Weeghel, M.; Otto, N.A.; Ferreira, B.L.; Maele, L.V.; Sirard, J.C.; de Vos, A.F.; de Jong, M.D.; et al. MTOR-Driven Glycolysis Governs Induction of Innate Immune Responses by Bronchial Epithelial Cells Exposed to the Bacterial Component Flagellin. Mucosal Immunol. 2021, 14, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Brands, X.; van’t Veer, C.; de Vos, A.F.; Scicluna, B.P.; van der Poll, T. Flagellin Induces Innate Immune Genes in Bronchial Epithelial Cells In Vivo: Role of TET2. Scand. J. Immunol. 2021, 94, e13046. [Google Scholar] [CrossRef]
- Nakamoto, K.; Watanabe, M.; Sada, M.; Inui, T.; Nakamura, M.; Honda, K.; Wada, H.; Ishii, H.; Takizawa, H. Pseudomonas Aeruginosa-Derived Flagellin Stimulates IL-6 and IL-8 Production in Human Bronchial Epithelial Cells: A Potential Mechanism for Progression and Exacerbation of COPD. Exp. Lung Res. 2019, 45, 255–266. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, K.; Ambati, B.; Yu, F.-S.X. Toll-like Receptor 5-Mediated Corneal Epithelial Inflammatory Responses to Pseudomonas Aeruginosa Flagellin. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4247–4254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial Flagellin-a Potent Immunomodulatory Agent. Exp. Mol. Med. 2017, 49, e373. [Google Scholar] [CrossRef] [PubMed]
- Tallant, T.; Deb, A.; Kar, N.; Lupica, J.; de Veer, M.J.; DiDonato, J.A. Flagellin Acting via TLR5 Is the Major Activator of Key Signaling Pathways Leading to NF-Kappa B and Proinflammatory Gene Program Activation in Intestinal Epithelial Cells. BMC Microbiol. 2004, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.S.; Kang, J.; Steiner, T.S. Enteroaggregative Escherichia Coli Flagellin-Induced Interleukin-8 Secretion Requires Toll-like Receptor 5-Dependent P38 MAP Kinase Activation. Immunology 2004, 112, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Van Maele, L.; Fougeron, D.; Janot, L.; Didierlaurent, A.; Cayet, D.; Tabareau, J.; Rumbo, M.; Corvo-Chamaillard, S.; Boulenouar, S.; Jeffs, S.; et al. Airway Structural Cells Regulate TLR5-Mediated Mucosal Adjuvant Activity. Mucosal Immunol. 2014, 7, 489–500. [Google Scholar] [CrossRef]
- Cai, Z.; Shi, Z.; Sanchez, A.; Zhang, T.; Liu, M.; Yang, J.; Wang, F.; Zhang, D. Transcriptional Regulation of Tlr11 Gene Expression in Epithelial Cells. J. Biol. Chem. 2009, 284, 33088–33096. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Zhang, G.; Hayden, M.S.; Greenblatt, M.B.; Bussey, C.; Flavell, R.A.; Ghosh, S. A Toll-like Receptor That Prevents Infection by Uropathogenic Bacteria. Science 2004, 303, 1522–1526. [Google Scholar] [CrossRef]
- Mathur, R.; Oh, H.; Zhang, D.; Park, S.-G.; Seo, J.; Koblansky, A.; Hayden, M.S.; Ghosh, S. A Mouse Model of Salmonella Typhi Infection. Cell 2012, 151, 590–602. [Google Scholar] [CrossRef] [Green Version]
- Hatai, H.; Lepelley, A.; Zeng, W.; Hayden, M.S.; Ghosh, S. Toll-Like Receptor 11 (TLR11) Interacts with Flagellin and Profilin through Disparate Mechanisms. PLoS ONE 2016, 11, e0148987. [Google Scholar] [CrossRef] [Green Version]
- Gschwandtner, M.; Derler, R.; Midwood, K.S. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior beyond Chemotaxis. Front. Immunol. 2019, 10, 2759. [Google Scholar] [CrossRef] [Green Version]
- Schutyser, E.; Struyf, S.; Van Damme, J. The CC Chemokine CCL20 and Its Receptor CCR6. Cytokine Growth Factor Rev. 2003, 14, 409–426. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, W.; Zhao, C.; Wang, Y.; Wu, H.; Sun, X.; Guan, Y.; Zhang, Y. Prostaglandin E2 Inhibits Group 2 Innate Lymphoid Cell Activation and Allergic Airway Inflammation Through E-Prostanoid 4-Cyclic Adenosine Monophosphate Signaling. Front. Immunol. 2018, 9, 501. [Google Scholar] [CrossRef] [PubMed]
- Zasłona, Z.; Okunishi, K.; Bourdonnay, E.; Domingo-Gonzalez, R.; Moore, B.B.; Lukacs, N.W.; Aronoff, D.M.; Peters-Golden, M. Prostaglandin E2 Suppresses Allergic Sensitization and Lung Inflammation by Targeting the E Prostanoid 2 Receptor on T Cells. J. Allergy Clin. Immunol. 2014, 133, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grishin, A.V.; Wang, J.; Potoka, D.A.; Hackam, D.J.; Upperman, J.S.; Boyle, P.; Zamora, R.; Ford, H.R. Lipopolysaccharide Induces Cyclooxygenase-2 in Intestinal Epithelium via a Noncanonical P38 MAPK Pathway. J. Immunol. 2006, 176, 580–588. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-J.; Flaczyk, A.; Wolfheimer, S.; Goretzki, A.; Jamin, A.; Wangorsch, A.; Vieths, S.; Scheurer, S.; Schülke, S. The Fusion Protein rFlaA:Betv1 Modulates DC Responses by a p38-MAPK and COX2-Dependent Secretion of PGE2 from Epithelial Cells. Cells 2021, 10, 3415. https://doi.org/10.3390/cells10123415
Lin Y-J, Flaczyk A, Wolfheimer S, Goretzki A, Jamin A, Wangorsch A, Vieths S, Scheurer S, Schülke S. The Fusion Protein rFlaA:Betv1 Modulates DC Responses by a p38-MAPK and COX2-Dependent Secretion of PGE2 from Epithelial Cells. Cells. 2021; 10(12):3415. https://doi.org/10.3390/cells10123415
Chicago/Turabian StyleLin, Yen-Ju, Adam Flaczyk, Sonja Wolfheimer, Alexandra Goretzki, Annette Jamin, Andrea Wangorsch, Stefan Vieths, Stephan Scheurer, and Stefan Schülke. 2021. "The Fusion Protein rFlaA:Betv1 Modulates DC Responses by a p38-MAPK and COX2-Dependent Secretion of PGE2 from Epithelial Cells" Cells 10, no. 12: 3415. https://doi.org/10.3390/cells10123415