
Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues



1
Department of Obstetrics and Gynecology, David Geffen School of Medicine, University of California Los Angeles (UCLA), Los Angeles, CA 90095, USA
2
Division of Endocrinology and Metabolism, Charles R Drew University of Medicine and Science, Los Angeles, CA 90059, USA
3
Department of Endocrinology, Men’s Health: Aging and Metabolism, Brigham and Women’s Hospital, Boston, MA 02115, USA
4
Department of Biology, California State University, Dominguez Hills, Los Angeles, CA 90747, USA
*
Author to whom correspondence should be addressed.
Cells 2021, 10(11), 3030; https://doi.org/10.3390/cells10113030
Submission received: 30 September 2021
/
Revised: 1 November 2021
/
Accepted: 3 November 2021
/
Published: 5 November 2021
(This article belongs to the Special Issue Human Brown Adipose Tissues)
Abstract
:Obesity-associated metabolic abnormalities comprise a cluster of conditions including dyslipidemia, insulin resistance, diabetes and cardiovascular diseases that has affected more than 650 million people all over the globe. Obesity results from the accumulation of white adipose tissues mainly due to the chronic imbalance of energy intake and energy expenditure. A variety of approaches to treat or prevent obesity, including lifestyle interventions, surgical weight loss procedures and pharmacological approaches to reduce energy intake and increase energy expenditure have failed to substantially decrease the prevalence of obesity. Brown adipose tissue (BAT), the primary source of thermogenesis in infants and small mammals may represent a promising therapeutic target to treat obesity by promoting energy expenditure through non-shivering thermogenesis mediated by mitochondrial uncoupling protein 1 (UCP1). Since the confirmation of functional BAT in adult humans by several groups, approximately a decade ago, and its association with a favorable metabolic phenotype, intense interest on the significance of BAT in adult human physiology and metabolic health has emerged within the scientific community to explore its therapeutic potential for the treatment of obesity and metabolic diseases. A substantially decreased BAT activity in individuals with obesity indicates a role for BAT in the setting of human obesity. On the other hand, BAT mass and its prevalence correlate with lower body mass index (BMI), decreased age and lower glucose levels, leading to a lower incidence of cardio-metabolic diseases. The increased cold exposure in adult humans with undetectable BAT was associated with decreased body fat mass and increased insulin sensitivity. A deeper understanding of the role of BAT in human metabolic health and its interrelationship with body fat distribution and deciphering proper strategies to increase energy expenditure, by either increasing functional BAT mass or inducing white adipose browning, holds the promise for possible therapeutic avenues for the treatment of obesity and associated metabolic disorders.
1. Introduction
The prevalence of obesity and related metabolic complications have significantly increased worldwide. Obesity represents a major global challenge that increases the risk for several chronic diseases, including hypertension, fatty liver disease, diabetes, insulin resistance, dementia, osteoarthritis, sleep apnea and some types of cancer [1]. Thus, obesity presents a substantial burden, both at the individual and population levels, by significantly affecting the quality of life as well as socio-economic productivity. Typical therapeutic approaches targeting dietary intake and physical exercise aimed at reducing obesity and related metabolic complications have not provided long-term health benefits [2,3], suggesting that there remains an unmet need for the development of novel strategies for the treatment and prevention of obesity and associated metabolic complications. Since obesity mainly develops from surplus energy stored in the adipose tissues, therapeutic strategies directed against increasing the energy expenditure and reducing the energy intake, or both, provide attractive avenues to combat obesity-associated metabolic complications. BAT, by virtue of its ability to modulate the organism’s global energy expenditure in the form of heat, has been a subject of tremendous interest to counteract obesity-related diseases. This remarkable capacity of BAT to dissipate energy is mediated via the upregulation of the expression of brown-fat-specific mitochondrial uncoupling protein 1 (UCP1) [4]. In addition to its primary role in regulating thermogenesis, BAT is also involved in cross talk with several peripheral tissues, including the liver, skeletal muscle, gut, the central nervous system and immune cells to control the systemic energy balance and glucose homeostasis [5]. It was widely believed that BAT is present in significant amounts only in newborns, infants and patients with pheochromocytoma, and that it declines with age in adults. However, over the past few years, major technological advancements in clinical studies using 18fluoro-2-deoxyglucose positron emission tomography (18FDG-PET) scanning in combination with computed tomography (CT) identified functionally active BAT depots in adult humans, which are more frequent in women than in men [6,7,8]. A substantial 18F-FDG uptake into the adipose tissue in the supraclavicular and paraspinal regions was observed in adult humans following mild cold exposure compared to the subjects kept at a warm temperature (27 °C). The amount of BAT was found to be inversely correlated with the body-mass index, suggesting the importance of BAT in regulating adult human metabolism [6,7,8].
BAT is also known to secrete various metabolism-improving factors, collectively called BATokines, that target various cell types. Several BATokines, including fibroblast growth factor 21 (FGF21), interleukin 6 (IL-6), growth differentiation factor 15 (GDF15), neuregulin 4 (Nrg4), bone morphogenic protein 8b (Bmp8b), 12,13-diHOME, 9-HEPE and follistatin (Fst), amongst several others, have been reported to serve as BATokines [9,10,11,12,13,14]. Morphological and gene expression data obtained from BAT transplantation studies revealed a larger adipocyte size and a reduced thermogenic gene expression compared to the endogenous BAT, suggesting that the beneficial effects of BAT transplantation could be primarily due to the secreted factors from the transplant [15]. Human pluripotent stem cell-derived brown adipocytes are reported to significantly improve glucose and lipid metabolism and prevent obesity [16,17]. Recent studies demonstrate the association of human BAT with lower blood glucose and triglyceride levels, higher HDL levels, and improved cardio-metabolic health [18]. Future retrospective studies are needed to understand the role of BAT in metabolic health and delineate the clinical and physiological significance of human BAT in the context of obesity-related cardio-metabolic diseases. Besides the well-accepted view that BAT activation is beneficial in general, results of unfavorable outcomes from the hyper-activation of BAT have also been reported in promoting atherosclerosis in animal models [19], cancer cachexia [20] and breast cancer cells in promoting cancer progression [21]. These findings suggest, therefore, that therapeutic interventions for the treatment of obesity-associated metabolic conditions using BAT activation should be carefully controlled and monitored.
2. Imaging Studies for Human Brown Adipose Tissue Detection
In spite of early reports of the presence of BAT in adult humans in 1972 [22], its physiological and clinical relevance was mostly ignored by the scientific and medical community. Several simultaneous studies [6,7,8,23] reported the detection of adult human brown adipose tissue, using PET/CT scans, and their confirmation as BAT by molecular analyses of the biopsies from neck and supraclavicular regions in patients undergoing surgery. Considering the need to develop appropriate research protocols to identify depots of metabolic activities following BAT-activating interventions in human subjects, a variety of imaging techniques have been used over the years. Since biopsies of human BAT have been exclusively restricted for ethical reasons and are limited mostly to cadavers or excised tissues obtained for the analysis of oncogenic markers, several biomedical imaging techniques over the years have allowed us to study the functional and morphological characteristics of BAT in vivo.
18F-FDG-PET/CT is used as a gold standard for studying activated BAT in humans since 2009, despite its many limitations for identifying and quantifying inducible BAT [24]. The use of ionizing radiation is a major concern when applying PET imaging for BAT, especially in longitudinal studies with healthy human subjects. This technique relies on glucose uptake, but the stored energy in oxidative BAT generated from intracellular triglyceride hydrolysis serves as the main substrate for BAT oxidative metabolism [25]. In addition, evidence for small amounts of glucose uptake by acute stimulation of the β3-adrenergic receptor (β3-AR) has been reported [26]. Therefore, probing oxidative metabolism or fatty acid uptake would be a more sensitive method for molecular imaging of BAT and the assessment of its activity compared to 18F-FDG-PET. Nevertheless, from a historical perspective, 18F-FDG-PET/CT remains the most widely used imaging study. Studies from the supraclavicular FDG uptake in cancer patients were amongst the first to provide evidence for BAT prevalence in adult humans [27,28]. 18F-FDG-PET/CT imaging was capable of detecting higher iBAT levels associated with visceral adiposity. However, the insensitivity of this method to detect inactive BAT in obese and metabolically unhealthy subjects presents a significant limitation of this method in a clinical setting.
Molecular resonance imaging (MRI) and magnetic resonance spectroscopy (MRS) methods do not use radiation and take advantage of the morphological and functional information to differentiate BAT from white adipose tissues (WAT). MRI provides a greater tissue contrast and an adequate spatial resolution compared to other tomographic imaging techniques. The MRI protocol, mostly used to differentiate between WAT and BAT and analyze BAT morphology, is based on the quantitation of water and fat content in the tissue. BAT’s water–fat composition in humans is primarily measured by using chemical shift-encoding water–fat imaging present in the MRI scanners and reported to show a higher reproducibility in humans across all ages [29,30,31,32,33]. However, a proper identification of BAT exclusively based on MRI fat fraction measurements remains challenging, as cold exposure or adrenergic activation significantly change BAT’s lipid content and varies widely with BMI, age, external temperature and diet [22,34,35]. The heterogeneous mixture of BAT and WAT existing in the supraclavicular fossa of human BAT presents another limitation of the water–fat MRI technique. Relatively longer scan times often requiring sedation and use of anesthesia are another practical limitation to MRI. Recently, infrared tomography (IRT) has been used as a valid complementary method to standard 18FDG-PET/CT [36]. This method has the advantage of being non-invasive and quick, and could be used in larger studies in human subjects for the detection of BAT. Moreover, near infrared spectroscopy (NIRS) has recently been used for measuring changes in oxygen-dependent light absorption in the tissue in a radiation-free non-invasive manner [37]. Time-resolved NIRS (NIRTRS) could be used to measure the vascular density in the supraclavicular region by estimating hemoglobin (Hb) concentrations, and thereby could provide helpful information on BAT-density [37]. Numerous other ongoing advances in imaging techniques and research protocols using improved tracers with significant improvement in sensitivity and specificity could provide an accurate and reliable assessment of BAT depots as well their metabolic activation. Such advancement in imaging techniques would provide highly improved techniques to measure the mass and activity of human BAT in vivo and understand BAT physiology and its potential as a therapeutic target for the treatment of obesity-associated metabolic complications.
3. Sexual Dimorphisms in Rodent and Human Thermogenic Adipose Tissue
The prevalence of obesity and related metabolic diseases is rapidly increasing in both men and women. However, the clinical manifestations of these metabolic diseases significantly differ between the genders. Several retrospective studies over the years have indicated that the prevalence of BAT is higher in women than in men [6,38]. There is also some indication that the metabolic activity and BAT mass is higher in female subjects [38]. However, few studies found no significant difference in BAT activity and mass between men and women following cold exposure [26,39,40,41,42]. In another study, van Marken Lichtenbelt et al. reported that 95% of all young men possessed BAT, implying that it is less likely that women would have a higher prevalence of BAT [7]. The anatomical distribution of BAT is similar in both women and men and mostly in the cervical, supraclavicular, axillary, paraspinal, mediastinal and abdominal areas, of which the supraclavicular regions are the most common location of active BAT detection with 18F-FDG-PET/CT [43]. In a diet-induced obesity model, female mice display a higher vascularization of the perigonadal WAT compared to the male mice [44]. It is possible that such gender differences in vascularization could exist in humans, and the sex hormones could differentially mediate the extent of adrenergic activation between men and women. Rodriguez-Cuenca et al. reported that BAT from female rats are more sensitive to β-adrenergic stimulation [45]. Although elevated adrenergic responses to lipolysis are reported in women [46], the underlying differences in β-adrenergic-receptor-signaling between rats and humans could be significantly altered based on the presence of β1-AR in humans [47]. Female rats show an increased abundance of UCP1 expression compared to males and display a higher mitochondrial density [45]. Data from human studies reported that women display a higher cold sensation and may need to activate thermogenesis at higher temperatures compared to men [48]. In addition, female rats show higher protein expression levels of UCP1 and various other proteins involved in thermogenesis compared to male rats fed on a high fat diet [49], supporting the view that rodents display a higher BAT thermogenic capacity in females. In human studies, a similar upregulated UCP1 gene expression was found in the subcutaneous and perirenal WAT of females [50,51].
A systematic global gene expression profile of more than 100 inbred mice shows a higher WAT UCP1 expression and its association with healthier phenotype in female mice [52]. On the other hand, UCP1 expression positively correlated with increased fat mass and insulin resistance in male mice. Collectively, these reports support sexual differences in BAT prevalence and function.
4. Aging-Induced Changes in Beige and Brown Adipocytes
Adipose tissue undergoes profound changes with aging in terms of its composition and distribution, leading to metabolic alterations. This aging-related adipose tissue dysfunction has a severe impact on whole-body energy homeostasis and often results in the progressive development of metabolic complications. It has been established that human BAT mass and activity decline with age. BAT formation starts during gestation, where it is critical for thermogenesis in early phases of human life. Supraclavicular BAT is the major site of metabolically active BAT in childhood. BAT activity increases during adolescence and sexual maturation and declines with aging. Cold-stimulated BAT activity by 18F-FDG-PET/CT imaging is rarely detectable in living individuals over the age of sixty, although the BAT mass may not change [53]. A significant aging-related decline of BAT activity and a concomitant loss of UCP1 expression in rodents have also been reported in several studies, suggesting that a gradual decline of BAT activity is common in both rodents and humans [54].
Possible mechanisms involved during the aging-related impairment of BAT development and function include loss of mitochondrial function, reduced sensitivity to sympathetic tone, alterations in brown adipose progenitor/stem cell function and changes in the endocrine control of BAT formation [55,56]. A reduced capacity of beige adipocyte formation, resulting from aging-induced changes in trophic factors in the adipose tissue, also contributes to the impairment of the thermogenic function [57]. Mitochondrial dysfunction has long been associated with several aging-related disorders, including obesity, type-2 diabetes, tumorigenesis and neuronal diseases [58]. Increased mitochondrial DNA mutations and a progressive decline in mitochondrial biogenesis are important contributors to human aging-associated adipose tissue dysfunction [59]. The age-associated impairment of the regenerative potential of brown adipose stem/progenitor cells could contribute to dysfunctional BAT adipocytes [58]. The defective differentiation ability of the CD137- and TMEM26-expressing sub-population of WAT progenitor cells could also contribute to the loss of beige adipocyte during aging [60]. SirT1, a potent inducer of adipose browning, can significantly enhance the beige adipocyte differentiation capability in an elderly adipose-derived mesenchymal stem cell (AD-MSC) population [61]. The sympathetic nervous system (SNS) plays a central role in the recruitment of brown adipocytes and its thermogenic activity through β3-AR stimulation [62]. SNS stimulation and BAT activation were lower in older lean men compared to the younger and obese men [63]. In addition, Yoneshiro et al. reported that a single nucleotide polymorphism in β3-AR and UCP1 contributed to an aging-associated decline in BAT activity in humans [64], suggesting that the aging-related decrease in SNS stimulation and sensitivity in older human may result in a decreased ability to recruit and activate BAT.
Aging-associated hormonal changes also play a central role in regulating the thermogenic activity in human BAT. While sex hormones like estrogens and androgens positively regulate BAT activity and function [65,66], glucocorticoids such as dexamethasone reduce the catecholamine-induced expression of UCP1 [67]. Relatively higher levels of sex hormones in the young counteract the generally inhibitory effects of glucocorticoids in BAT. Because of aging, sex hormone levels significantly decline without a significant alteration in glucocorticoid levels, resulting in reduced energy expenditure and the loss of BAT activity. The aging-associated decrease in thyroid hormone (TH), known regulators of white adipose browning, has recently been demonstrated in murine WAT [68,69]. In light of these reports, future human studies are needed to develop effective pharmacological interventions aimed at preventing the aging-associated decline in BAT activity. Figure 1 summarizes the BAT distribution in infants, women and men.
5. Molecular Gene Signature of Brown and Beige Adipocytes in Rodents and Humans
Various cell-sorting and lineage-tracing studies in rodents have demonstrated that different types of adipose cells have distinct sets of molecular markers and arise from distinct pools of progenitors [60,70,71,72]. Classical brown adipocytes present in BAT originate from a population of dermomyotomes expressing Myf5, Pax7 and engrailed 1 [73,74]. Analyses of the global gene expression profile of interscapular BAT demonstrate that these brown adipocyte precursors resemble the gene signature, with skeletal muscle cells that also express a Myf5-positive precursor population [75]. Data from the proteomics analysis of interscapular BAT and skeletal muscle further confirmed a strong similarity in the protein levels [76]. Classical BAT is reported to selectively express high levels of Lhx8, Zic1, Eva1 (Mpzl2) and Epsti 1, although there are recent reports that suggest that Lhx8 could also be present in human beige adipocyte [77]. Several additional markers, including Ebf2, Pdk4, Hsbp7, Oplah, Fbxo3, Slc29a1 and Acot2, were found to be highly enriched in interscapular BAT obtained from 129SVE mice [60]. Recently, Prex1, a positive regulator of Ucp1 gene expression, was found to be another BAT marker present in the brown adipose precursor cells [78]. Since no published studies have reported the upregulation of Prex1 levels in beige adipocytes, it could be considered as a unique BAT marker.
Beige adipocytes, on the other hand, arise from a non-Myf5 lineage [79], although there are several recent reports that beige adipocytes in WAT depots may have multiple origins [50,72]. Beige adipocytes, in general, are selectively enriched in Tmem26, CD137, CD40 and Tbx1. These markers are expressed in high levels in epididymal adipose depots compared to the interscapular BAT following cold exposure or β-adrenergic stimulation [60]. Several additional beige-selective genes, including Klh113, Ear2, Sp100, and Slc27a, were reported in the same study following the analysis of gene expression in inguinal WAT and BAT. Gene expression and a histological analysis of human BAT that resemble beige adipocytes identified Cited1, Hoxc8, Hoxc9 and Shox2 as beige-selective genes [77,80]. Differences in the levels of microRNA (miRNA) between beige and BAT have also been identified. The Mir193b-365 cluster was found to be a key regulator of BAT development [81]. Additional miRNAs required for the maintenance and differentiation of BAT, including miRNA182 and miRNA 203, were identified following a comparative miRNA expression profiling of mouse and human BAT [82,83]. Some recently identified miRNA, including miRNA196b and miRNA26, are reported to positively and negatively regulate the brown adipogenesis of white fat progenitor cells, respectively [84,85].
Transcriptional analyses of human BAT isolated from multiple adipose depots display molecular signatures that closely resemble beige adipose-selective genes. The authors found that common beige adipose markers including HOXC8, HOXC9, CITED1, CD137, TMEM26 and TBX1 were highly expressed in all human BAT [77]. A molecular characterization of human perirenal BAT depot shows a higher expression of HMGCS2, CKMT1A/1B, KCNK3 and PGC1-α genes in BAT compared to the WAT [86]. They also identified high levels of BAT-associated genes, including ACOT11, PYGM and FABP3, in the perirenal adipose tissue, which display a high correlation with UCP1 and to each other. Recent studies in humans indicate high levels of UCP1-immunopositive adipocytes in the anatomically defined neck fat from the supraclavicular and spinal cord regions of adult human subjects and display molecular signatures of classical BAT, including high expression levels of genes involved in mitochondrial biogenesis and thermogenesis, including UCP1, PGC1α and DIO2 [87]. The degree of correlation of the genes assessed by hierarchical clustering in deep neck tissues with the highest UCP1 expression show increased levels of UCP1, ZIC1 and LHX8. Some genes previously designated as markers of BAT lineage in mice, including EBF3, MPZL2 and FBXO31, did not have an elevated expression in the deeper human neck depot or associate closely with UCP1 expression. On the other hand, the expression levels of CD137, TMEM26, TBX1, EVA1 and EBF3 does not differ between subcutaneous and deeper neck adipose depots. Since adipose tissue depots are highly heterogeneous and inter-subject differences could significantly affect the thermogenic potential, clonal cell lines from human neck fat were generated and characterized for their adipogenic and metabolic functions in vitro and in vivo after transplantation into immune-deficient nude mice [78]. Using the UCP1 reporter system, the authors were able to reveal two general categories of genes in preadipocytes with enhanced capability to regulate the late-stage thermogenic differentiation program. PREX1, CTTNBP2, ACTC1 and SSTR1 genes were identified in the first category as positive regulators with enhanced thermogenic capacity. On the other hand, in the second category, DMRTA1, EDNRB, FAT1 and PTPRB were reported to have a negative correlation with UCP1-luciferase levels [78]. Zuriaga et al., performed comparative gene analyses of browning gene expression profiles in the subcutaneous and visceral WAT of obese mice and human subjects and reported a pattern that is opposite to what is observed in mice [88]. Obese mice displayed a higher expression of typical browning markers UCP1, Cidea and Prdm16 and beige markers Tbx1 and P2rx5 compared to the epididymal WAT. However, in obese humans, visceral WAT show a higher expression of UCP1 and several other adipose browning (CIDEA, PRDM16, TBX1 and P2RX5) and mitochondrial (COX8B, PGC1α, ATP5A and NDUFA1) genes compared to the subcutaneous adipose depots [88]. These findings suggest an increased browning capacity of human visceral WAT compared to the subcutaneous WAT.
6. Cold Exposure and Activation of Sympathetic Nervous System
In earlier experiments, the appearance of brown-like adipocytes in the parametrial fat depots was reported in mice following severe cold exposure [89]. Subsequent studies in murine models over the years clearly established a role for cold-induced BAT activation as a physiological response to acute or mild cold exposure for maintaining thermal demands through non-shivering thermogenesis (NST) [90,91]. Cold acclimation led to the activation of PGC1α and the induction of a UCP1-PRDM16-signaling cascade in murine studies [92].
The higher prevalence of BAT during cold season and the reported seasonal variability in human subcutaneous and retroperitoneal WAT strongly suggest that cold exposure should promote adipose browning and the activation of BAT characteristics in humans [92,93]. Cold exposure is the most common method for BAT activation in humans, via the activation of NST [94], that is achieved either by reducing the ambient room temperature between 16 and 19 °C [95] or with cold water-infused jackets [96,97]. Cold acclimation increases the BAT’s oxidative capacity, which correlates with a reduction of shivering thermogenesis [94,98]. Cold adaptation in BAT is also associated with mitochondrial remodeling and vascularization for adaptive thermogenesis [91] and fatty acid oxidation through UCP1 during high metabolic demands [99].
A significant increase in BAT activity and NST was reported in humans following 10-day acclimation protocols [94]. Respiratory measurements in skeletal muscle suggested no significant contribution from mitochondrial uncoupling towards increased NST. The authors did not find any sex differences in BAT prevalence and activities between men and women. There was no significant increase in adipose browning markers in subcutaneous WAT, suggesting that a stronger cold stimulation or prolonged period of cold acclimation may be needed for the upregulation of general browning markers. In another human study, Ouellet et al. reported that BAT’s oxidative metabolism significantly contributed to an increased energy expenditure, glucose uptake and non-esterified fatty acid (NEFA) turnover in healthy men during acute cold exposure [96]. A similar increase in glucose uptake following the activation of BAT by cold exposure and insulin was reported in another study [100], suggesting that BAT activation leads to the clearance of plasma glucose. The analysis of glucose metabolism in individuals with (BAT+) or without (BAT−) BAT under cold exposure showed an increased whole-body glucose in the fasted and insulin- stimulated BAT+ individuals. This only suggests that BAT can indeed significantly influence systemic glucose metabolism in humans [101] and may represent a novel target to combat hyperglycemia. In addition, it was observed that a mild daily cold exposure (17 °C) for 6 weeks induces BAT recruitment in healthy human subjects with low BAT activity independently of age and fat-free mass [42], suggesting that the cold-induced recruitment of BAT could serve as a viable means to decrease fat mass in aging populations.
The sympathetic nervous system plays an important role in the physiological regulation of BAT activity, mediated mainly via the activation of the beta 3-adrenergic receptor (β3-AR) present in brown adipocytes. The chronic treatment of rats with selective β3-AR agonist CL316243 significantly increased the energy expenditure and body temperature, and these effects were associated with a 3–4 fold increase in Ucp1 expression in interscapular BAT [42,102]. In this study, CL316243 not only promoted BAT’s mitochondrial biogenesis, but it also inhibited the development of WAT hyperplasia during diet-induced obesity. The treatment of male mice with CL316243 led to a significant upregulation of the Ucp1 gene expression in retroperitoneal adipose tissue and correlated with weight loss [102,103]. These studies conducted in rodents support the view that the activation of β3-AR with CL316243 is an effective method to induce BAT activity and stimulate the beige cell phenotype in typical WAT. The long-term sympathetic regulation of thermogenic BAT activity and its association with reduced body weight in catecholamine-producing human pheochromocytoma patients is well known. The administration of β3-AR agonist in human subjects for 4–8 weeks significantly improved insulin sensitivity and lipid profiles without any change in body weight [104,105]. More recently, the treatment of human subjects with a single high dose of β3-AR agonist mirabegron increased BAT’s 18F-FDG uptake, and the energy expenditure [106], although β2-AR and β1-AR are more highly expressed in human BAT [107]. Mixed AR agonist ephedrine also increased the 18F-FDG uptake in BAT in lean but not obese subjects at room temperature [108]. However, the side-effects associated with increased heart rate and blood pressure induced by these compounds significantly limit the potential use of this therapeutic approach [106,108,109].
7. WAT Browning Factors
Due to the relatively lower amount of BAT present in adult humans, WAT has remained the major focus of targeting against obesity and related metabolic diseases. Even though WAT is known to have fewer numbers of mitochondria with larger lipid droplets, the manipulation of WAT to brown-like phenotype with increased thermogenic capacity through the process called “adipose browning” is being explored as a major therapeutic strategy. Several browning agents have been identified in recent years in animal studies, some of which are currently being investigated in humans [110,111]. The safe activation of adipose browning requires the identification of the specific molecular pathways involved in WAT browning. Fibroblast growth factor 21 (FGF21), expressed mainly in the liver [112], is also present in BAT [113] and regulates energy metabolism by promoting adipose browning and lipoprotein catabolism [114,115]. An increased FGF21 expression in adipose tissues was observed within 6 h of cold exposure without any change in its circulating levels [114]. Using differentiated brown and inguinal adipocytes derived from PGC-1α-knockout (KO) and PGC-1α Flox mice, Fisher et al. demonstrated that PGC-1α is required for FGF21-induced thermogenesis in adipose tissues [114]. The induction of PGC-1α by FGF21 was mostly regulated at the post-translational level. This induction of the PGC-1α protein content in brown and WAT was significantly attenuated in FGF21-KO mice, suggesting an important role for PGC-1α during FGF21-induced adipose browning [114]. The therapeutic potential of FGF21 analogs has recently been explored in obese human subjects with type-2 diabetes [116,117]. These analogs reduced body weight and improved circulating lipids but failed to improve glucose levels. These studies did not measure BAT activity or WAT browning. Additional endocrine factors including natriuretic peptides, prostaglandins and β-aminoisobutyric acid (BAIBA) and thyroid hormones (TH) are known to influence WAT differentiation and browning [118,119,120]. Several members of the transforming growth factor beta (TGF-β) superfamily, including bone morphogenic proteins (BMP) such as BMP7, BMP8b and BMP4, regulate various aspects of WAT browning and BAT differentiation in rodents, as well as in human adipocyte cell models [121,122,123]. Additional modulators of the TGF-β signaling pathway, including follistatin (Fst) and inhibitors of myostatin (Mst), have been recently reported to upregulate key thermogenic markers, increase mitochondrial biogenesis and promote WAT browning [124,125,126]. Improved insulin sensitivity and reduced lipogenic effects of Gleevec were associated with increased WAT browning and energy expenditure. These beneficial effects of Gleevec were blocked by the inhibition of PPARγ phosphorylation, suggesting a critical role for the PPARγ-mediated signaling pathway during the process [127]. Thiazolidinediones (TZDs), known agonists of PPARγ used as insulin sensitizing drugs for the treatment of T2D, promote adipose browning both in vivo and in vitro [128,129]. Another antidiabetic agent, liraglutide, that acts as an agonist of the glucagon-like peptide-1 (GLP-1) receptor, significantly reduced the weight gain in obese subjects [130]. Although the GLP-1 mediated weight loss results were mainly due to appetite suppression, increased energy expenditure is recently reported in liraglutide-treated obese human patients with type-2 diabetes [131]. The bile acid chenodeoxycholic acid (CDCA) increases BAT activity in both mice and humans [132,133]. Several plant-derived products influence adipocytes’ metabolic characteristics. Natural plant polyphenols such as resveratrol and butein increase the energy expenditure and promote thermogenesis by the induction of Prdm16, Ucp1 and Pgc-1α [134]. Another plant-based compound, capsaicin, stimulates the thermogenic Ucp1 and Bmp8b genes and triggers WAT browning by stimulating the SirT1-dependent acetylation of PPARγ and Prdm16 [135]. Another study in humans using a single dose of capsinoids demonstrated a selectively increased energy expenditure in people with metabolically active BAT, suggesting the beneficial effects of capsinoid-induced WAT browning in both humans and mice [136]. Other polyphenols, including tea catechin, have also been suggested as safe and effective therapeutic options to promote white adipose browning [137]. Lactate, an important metabolic intermediate, upregulates Ucp1 expression and induces WAT browning by regulating PPARγ signaling [138].
8. Exercise
Exercise is an effective way to prevent and treat obesity and related metabolic diseases. Although the adaptation to skeletal muscle is most widely studied, it also significantly influences the phenotype and functions of adipose tissues. Exercise increases whole-body energy expenditure and several metabolic adaptations to WAT through the process of adipose beiging mainly in subcutaneous WAT (scWAT). Exercise induces the upregulation of thermogenic genes such as Prdm16 and Ucp1 in scWAT and increases the presence of multilocular lipid droplets in adipocytes [139,140]. PGC1-α, a muscle-derived myokine, seems to be an important mediator of the exercise-induced browning process that influences myogenesis as well as mitochondrial oxidative phosphorylation [141]. Irisin, another browning agent that works through the PGC1-α pathway, is secreted from skeletal muscle and upregulates UCP1 expression in scWAT, increases energy expenditure and protects against diet-induced weight gain in mice [141]. The irisin-induced adipose browning of white adipocytes in vitro and its circulating levels are increased in humans following exercise and cold exposure [141,142]. However, human data obtained from several laboratories studying the effect of exercise on irisin levels are inconsistent and raise doubts about the role of irisin in mediating exercise effects [143,144]. Although the direct effects of irisin administration in humans have not yet been conducted, in some human studies plasma irisin levels were found to be higher in obese humans and its levels correlate positively with plasma FFA [145,146]. In light of these controversial reports, the beneficial role of irisin for treating or managing obesity has been questioned. Several other exercise-induced factors, including FGF21, IL-6, follistatin, myostatin, meteorin-like1 (Metrnl), lactate, BAIBA and brain-derived neurotrophic factor (BDNF) have been associated with WAT browning [119,147,148,149,150]. Exercise-induced changes in WAT include increased mitochondrial biogenesis and glucose uptake, decreased lipid content and inflammation [151,152]. Although such adaptations to exercise in multiple tissues are known in both rodents and humans, it is important to note that more investigations are needed to clearly establish exercise-induced adipose browning and whole-body metabolic health in human subjects.
9. MicroRNAs in Browning
Micro RNAs (miRNAs) are single stranded small non-coding RNAs (20–24 nucleotides) that regulate a wide spectrum of biological processes implicated in cellular metabolism and the pathology of a variety of diseases. In recent years, several miRNAs were identified that play important roles in regulating beige and brown adipocyte differentiation and function [153,154]. Delineating the regulatory mechanisms of miRNA action has great potential to provide therapeutic approaches to combat obesity and related metabolic diseases by increasing the overall energy expenditure. Several miRNAs, including miRNA328, miRNA193b-365, miRNA203, miRNA182, miRNA129, miRNA106b and miRNA93b, have been identified that specifically regulate brown adipogenesis either by promoting or inhibiting the BAT thermogenic program [153]. The inhibition of miRNA182 or miRNA203 results in a significant reduction of various BAT markers, including UCP1, PGC1-α and Cidea, as well as the downregulation of genes involved in the electron transport chain and oxidative phosphorylation [83]. However, there was no change in the levels of the classical adipogenic markers PPARγ and FABP4 in this study. Interestingly, the platelet-derived growth factor receptor alpha (Pdgfrα) and insulin-induced gene 1 (Insig1) were identified as targets of miRNA182 or miRNA203 [83]. The miRNA193b-365 cluster is highly expressed in BAT and is reported to promote brown fat differentiation by inhibiting the runt-related transcription factor 1 translocated to 1 (Runx1t1) [81]. The function of these miRNAs is still not confirmed, as in-vivo studies show no change in BAT morphology and function in the absence of this cluster. The levels of miRNA328 in mice on a high-fat diet were decreased in BAT and found to be upregulated during brown adipocyte differentiation [155]. On the other hand, a decreased expression of several thermogenic genes, including Ucp1, Prdm16, Pgc-1α and Cidea, was reported after the inhibition of miRNA328 in this study. In addition, the overexpression of miRNA328 in brown adipocytes significantly inhibited the Bace 1 protein, that decreases energy expenditure and UCP1 expression [156]. The MiRNA106b-93 cluster is identified, as negative regulators of BAT differentiation and their levels are found to be increased in obese mouse BAT [157]. The inhibition of miRNA 106-93 induced the expression of several BAT markers, including UCP1, PRDM16, PGC1-α and Cidea [157]. These findings suggest, therefore, an important role for the miRNA106b-93 cluster in BAT energy homeostasis. The circulating levels of liver-specific miRNA122 were increased in obese individuals and found to be negatively associated with BAT activity [158].
In addition, several miRNAs play a significant role in regulating beige adipocyte development and function. A better understanding of the key beige-specific miRNAs and their regulation would be critical for developing efficient therapeutic approaches to increase cellular energy expenditure and slow down the progression of obesity. miRNA196a plays an important role in the browning of white adipose progenitor cells, with increased expression of several brown adipogenic genes, such as C/ebpα, Prdm16, Ucp1 and Pgc1-α [84]. MiRNA196a induces WAT beiging in humans by the direct binding and suppression of homeobox C8 (Hoxc8), a known repressor of C/EBPβ. The overexpression of miRNA196a improved glucose metabolism and resistance to diet-induced obesity in mice [84]. The MiRNA26 family (miRNA26a and miRNA26b) has been identified as an important regulator of white and beige adipocyte differentiation in humans [159]. The MiRNA26 level is induced in WAT following cold exposure and mimics of miRNA26a/b promote adipose browning and the mitochondrial phenotype by upregulating the expression of UCP1, PGC1-α and aP2. Mechanistically, the adipose browning effects of the miRNA26 family are mediated by the direct targeting of ADAM17 [159]. MiRNA125-5p has been identified as a negative regulator of adipose browning. Its expression level is higher in WAT compared to BAT and decreases during browning [160]. Beige adipocyte formation and mitochondrial biogenesis is significantly inhibited following the injection of miRNA125-5p into WAT [161].
In addition, several miRNAs have been identified that play important roles in regulating both BAT and WAT browning. MiRNA30 family members promote the development of beige and thermogenesis [162]. The levels of miRNA30 are higher in BAT compared to WAT. The activation of β-AR signaling or cold exposure upregulates the expression of key thermogenic (UCP1 and Cidea) and mitochondrial biogenesis genes in brown adipocytes. The inhibition of miRNA30b/c levels in vivo results in the downregulation of UCP1 expression and the reduction of BAT mitochondrial respiration by its direct action on the targeting of receptor-interacting protein 140 (RIP140) [163], suggesting that miRNA30b/c positively regulates beige adipocyte development as well as BAT thermogenesis. MiRNA455 was identified as a key regulator of brown adipogenesis in multipotent mesenchymal cells. MiRNA455 levels are induced by cold exposure and BMP7 treatment [160]. The inhibition of miRNA455 results in the suppression of brown adipose differentiation. In addition, adipose-specific transgenic mice overexpressing miRNA455 display significant WAT browning, increased insulin sensitivity and glucose tolerance and resistance to a diet-induced increase in body weight [160]. MiRNA378 has been identified as the only miRNA that can regulate brown and white adipocytes in opposing fashion. The adipose-specific overexpression of miRNA378 increased the BAT mass and brown adipogenesis but showed a decreased WAT mass and cell size, and reduced diet-induced obesity in ob/ob mice [164]. These dual effects of miRNA378 in promoting brown adipogenesis but blunting WAT beige adipogenesis are caused by the direct targeting of Pde1b, a phosphodiesterase that causes a reduction in cAMP levels [165]. Table 1 briefly summarizes various miRNAs involved in brown/beige fat development and their targets [83,84,85,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169]. Further studies are needed to clearly understand the complex mechanism in order to develop suitable miRNA-based therapeutic approaches for the treatment of obesity and related metabolic disorders.
10. Challenges and Potential Concerns with BAT Activation as a Therapeutic Strategy to Combat Obesity
The therapeutic targeting of BAT for obesity was envisioned in late 1970s based on rodent studies, suggesting that BAT may be the major site for thermogenesis in response to both cold and diet [170,171]. The unequivocal discovery of active and functional BAT in healthy human subjects confirmed the expression of thermogenic UCP1 and its energy-dissipating capacity in humans. Increasing energy expenditure by cold exposure and BAT activation holds great potential to combat obesity-related metabolic complications; however, there are notable concerns with this approach that must be carefully evaluated. While there are many similarities between mouse and human BAT, notable morphological and anatomical differences also exist. In order to translate the significance of rodent studies in humans, it is important to generate proof of concept studies in humans for successful clinical outcomes. The activation of the β-AR signaling needed for BAT thermogenesis is mediated via β3-AR in rodents. Recent studies in humans revealed that β2-AR is the predominant adrenergic receptor [107], suggesting that a selective BAT activation in humans may be difficult to achieve and requires the activation of another β-AR subtype. Targeting thermogenic BAT activation could result in a significantly increased heart rate, as the increased energy expenditure requires an increased demand for oxygen. A chronic BAT activation from β3-AR agonists or thyroid hormone replacement may cause tachycardia and hypertension, resulting in unwanted cardiovascular side effects such as stroke or myocardial infarction [172,173]. Another potential problem during the pharmacological activation of BAT could result from the dissipation of heat. Weight loss medication 2,4-dinitrophenol (DNP), that works through the uncoupling of oxidative phosphorylation, causes hyperthermia and could be fatal at higher doses [174]. Although it is unlikely that selective pharmacological approaches to activate thermogenesis will have severe thermal stress at low doses, caution is warranted to avoid unwanted off-target effects. In addition, the impact of increased energy expenditure could lead to a compensatory hyperphagic response during cold exposure [175]. However, data accumulated over the past decade indicate that pharmacotherapy approaches to activate BAT via β3-AR agonists or the administration of various thermogenic food ingredients lead to increased food intake [176].
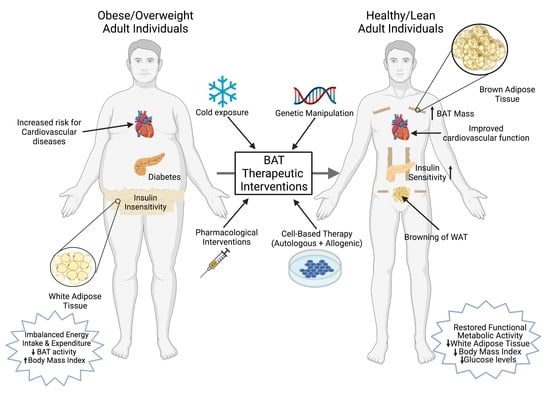
Several recent studies also suggest that BAT activation could be harmful. A cold-induced BAT hyper activation in ApoE-null mice resulted in the exacerbation of atherosclerotic plaques and their stability [19]. This cold-induced lipolysis in ApoE-null mice was food-intake-independent and significantly increased the LDL remnants. Increased levels of inflammatory cells and plaque-associated micro vessels were detected in cold-acclimated mice. The deletion of UCP1 in BAT protected these ApoE-null mice from the development of cold-induced atherosclerotic lesions. Adverse effects of BAT activation are also reported in the development of cancer cachexia [177]. Interleukin-6 (IL6) and growth and differentiation factor 15 (GDF15), two BAT-secreted factors, are key mediators of cancer-induced cachexia [177,178]. Using xenotransplantation models that use human breast cancer cells as well as fresh human tumor tissues transplanted into nude mice, Singh et al. provided evidence for the involvement of beige and brown adipose cells during the progression of breast cancer [21]. In this study, a selective enrichment of BAT-selective genes was observed in the tumor as well as the host cells, and a depletion of UCP1 and the Myf5-positive population resulted in a significantly reduced tumor size. It is not known whether BAT plays an important role in the progression of other cancers. These findings clearly indicate, therefore, that BAT-based therapeutic drug development for the treatment of obesity and related diseases should be carefully assessed to avoid unwanted adverse effects. Figure 2 summarizes a brief overview of several therapeutic interventions to increase BAT mass and activity.
11. Conclusions
Since the demonstration of active and functional BAT in adult human subjects, the activation of adipose browning characteristics to dissipate excess stored energy has shown great promise to combat obesity-associated comorbidities including type-2 diabetes, insulin resistance, hyperlipidemia and cardiovascular diseases. The identification of novel compounds that can promote adipose browning and understand the molecular signaling mechanism involved during the process will provide new insights for therapeutic drug development targeting obesity-related diseases. Recent studies have also provided ample evidence that BAT also plays an important role in the regulation of systemic and peripheral glucose homeostasis by functioning as a metabolic sink for glucose and triglycerides. A number of pharmacological agents have been tested to activate the adipose browning characteristics (Table 2). However, these attempts have mostly failed, partially due to their modest efficiency and adverse side effects, including elevated blood pressure [108]. It is also important to develop new techniques to assess the thermogenic capacity of novel compounds in adult humans, as 18F-FDG-PET/CT scans do not accurately measure the total BAT mass. Other currently available devices are not sensitive enough to accurately detect UCP1-expressing subcutaneous WAT and other adjacent tissues. Recent studies demonstrating the therapeutic potential of transplanted BAT in rodent studies to improve weight loss and improved insulin sensitivity have not been explored in human subjects. The effectiveness and clinical relevance of BAT activation and recruitment in human subjects needs to be further explored in detail.
Author Contributions
Conceptualizations, R.S. and S.P.; writing—original draft preparation, R.S., A.B. and G.D.; schematics and table creation, A.B. and G.D.; review and editing—R.S. and S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by TRDRP grant T31IP1551 (RS), NIH grants SC1CA232319 (SP), UHI NIMHD S21MD000103, and the Accelerating Excellence in Translational Sciences (AXIS) Center U54MD007598, at Charles R Drew University of Medicine and Science.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Calle, E.E.; Rodriguez, C.; Walker-Thurmond, K.; Thun, M.J. Overweight, Obesity, and Mortality from Cancer in a Prospectively Studied Cohort of U.S. Adults. N. Engl. J. Med. 2003, 348, 1625–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heymsfield, S.B.; Wadden, T.A. Mechanisms, Pathophysiology, and Management of Obesity. N. Engl. J. Med. 2017, 376, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Drug Therapy of the Metabolic Syndrome: Minimizing the Emerging Crisis in Polypharmacy. Nat. Rev. Drug. Discov. 2006, 5, 295–309. [Google Scholar] [CrossRef]
- Kajimura, S.; Saito, M. A New Era in Brown Adipose Tissue Biology: Molecular Control of Brown Fat Development and Energy Homeostasis. Annu. Rev. Physiol. 2014, 76, 225–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, A.; Becerril, S.; Ezquerro, S.; Méndez-Giménez, L.; Frühbeck, G. Crosstalk between Adipokines and Myokines in Fat Browning. Acta Physiol. 2017, 219, 362–381. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A.; et al. Identification and Importance of Brown Adipose Tissue in Adult Humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.A.F.L.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J.J. Cold-Activated Brown Adipose Tissue in Healthy Men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.-J.; Enerbäck, S.; et al. Functional Brown Adipose Tissue in Healthy Adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef]
- Hondares, E.; Iglesias, R.; Giralt, A.; Gonzalez, F.J.; Giralt, M.; Mampel, T.; Villarroya, F. Thermogenic Activation Induces FGF21 Expression and Release in Brown Adipose Tissue. J. Biol. Chem. 2011, 286, 12983–12990. [Google Scholar] [CrossRef] [Green Version]
- Chartoumpekis, D.V.; Habeos, I.G.; Ziros, P.G.; Psyrogiannis, A.I.; Kyriazopoulou, V.E.; Papavassiliou, A.G. Brown Adipose Tissue Responds to Cold and Adrenergic Stimulation by Induction of FGF21. Mol. Med. 2011, 17, 736–740. [Google Scholar] [CrossRef]
- Oka, M.; Kobayashi, N.; Matsumura, K.; Nishio, M.; Nakano, K.; Okamura, T.; Okochi, H.; Minamisawa, T.; Shiba, K.; Saeki, K. New Role for Growth/Differentiation Factor 15 in the Survival of Transplanted Brown Adipose Tissues in Cooperation with Interleukin-6. Cells 2020, 9, 1365. [Google Scholar] [CrossRef] [PubMed]
- Whittle, A.J.; Carobbio, S.; Martins, L.; Slawik, M.; Hondares, E.; Vázquez, M.J.; Morgan, D.; Csikasz, R.I.; Gallego, R.; Rodriguez-Cuenca, S.; et al. BMP8B Increases Brown Adipose Tissue Thermogenesis through Both Central and Peripheral Actions. Cell 2012, 149, 871–885. [Google Scholar] [CrossRef] [Green Version]
- Stanford, K.I.; Lynes, M.D.; Takahashi, H.; Baer, L.A.; Arts, P.J.; May, F.J.; Lehnig, A.C.; Middelbeek, R.J.W.; Richard, J.J.; So, K.; et al. 12,13-DiHOME: An Exercise-Induced Lipokine That Increases Skeletal Muscle Fatty Acid Uptake. Cell Metab. 2018, 27, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Leiria, L.O.; Wang, C.-H.; Lynes, M.D.; Yang, K.; Shamsi, F.; Sato, M.; Sugimoto, S.; Chen, E.Y.; Bussberg, V.; Narain, N.R.; et al. 12-Lipoxygenase Regulates Cold Adaptation and Glucose Metabolism by Producing the Omega-3 Lipid 12-HEPE from Brown Fat. Cell Metab. 2019, 30, 768–783. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, Z.; Zhu, X.; Meng, M.; Li, L.; Shen, Y.; Chi, Q.; Wang, D.; Zhang, Z.; Li, C.; et al. Brown Adipose Tissue Transplantation Improves Whole-Body Energy Metabolism. Cell Res. 2013, 23, 851–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanford, K.I.; Middelbeek, R.J.W.; Townsend, K.L.; An, D.; Nygaard, E.B.; Hitchcox, K.M.; Markan, K.R.; Nakano, K.; Hirshman, M.F.; Tseng, Y.-H.; et al. Brown Adipose Tissue Regulates Glucose Homeostasis and Insulin Sensitivity. J. Clin. Investig. 2013, 123, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishio, M.; Yoneshiro, T.; Nakahara, M.; Suzuki, S.; Saeki, K.; Hasegawa, M.; Kawai, Y.; Akutsu, H.; Umezawa, A.; Yasuda, K.; et al. Production of Functional Classical Brown Adipocytes from Human Pluripotent Stem Cells Using Specific Hemopoietin Cocktail without Gene Transfer. Cell Metab. 2012, 16, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becher, T.; Palanisamy, S.; Kramer, D.J.; Eljalby, M.; Marx, S.J.; Wibmer, A.G.; Butler, S.D.; Jiang, C.S.; Vaughan, R.; Schöder, H.; et al. Brown Adipose Tissue Is Associated with Cardiometabolic Health. Nat. Med. 2021, 27, 58–65. [Google Scholar] [CrossRef]
- Dong, M.; Yang, X.; Lim, S.; Cao, Z.; Honek, J.; Lu, H.; Zhang, C.; Seki, T.; Hosaka, K.; Wahlberg, E.; et al. Cold Exposure Promotes Atherosclerotic Plaque Growth and Instability via UCP1-Dependent Lipolysis. Cell Metab. 2013, 18, 118–129. [Google Scholar] [CrossRef] [Green Version]
- Petruzzelli, M.; Wagner, E.F. Mechanisms of Metabolic Dysfunction in Cancer-Associated Cachexia. Genes Dev. 2016, 30, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Parveen, M.; Basgen, J.M.; Fazel, S.; Meshesha, M.F.; Thames, E.C.; Moore, B.; Martinez, L.; Howard, C.B.; Vergnes, L.; et al. Increased Expression of Beige/Brown Adipose Markers from Host and Breast Cancer Cells Influence Xenograft Formation in Mice. Mol. Cancer Res. 2016, 14, 78–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, J.M. The Distribution of Brown Adipose Tissue in the Human. J. Anat. 1972, 112, 35–39. [Google Scholar]
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K.; et al. High Incidence of Metabolically Active Brown Adipose Tissue in Healthy Adult Humans: Effects of Cold Exposure and Adiposity. Diabetes 2009, 58, 1526–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, S.C.; Sampath, S.C.; Bredella, M.A.; Cypess, A.M.; Torriani, M. Imaging of Brown Adipose Tissue: State of the Art. Radiology 2016, 280, 4–19. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.W.Y.; Foster, D.O. Uptake of Glucose and Release of Fatty Acids and Glycerol by Rat Brown Adipose Tissue in Vivo. Can. J. Physiol. Pharmacol. 1986, 64, 609–614. [Google Scholar] [CrossRef]
- Olsen, J.M.; Sato, M.; Dallner, O.S.; Sandström, A.L.; Pisani, D.F.; Chambard, J.-C.; Amri, E.-Z.; Hutchinson, D.S.; Bengtsson, T. Glucose Uptake in Brown Fat Cells Is Dependent on MTOR Complex 2–Promoted GLUT1 Translocation. J. Cell Biol. 2014, 207, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohade, C.; Osman, M.; Pannu, H.K.; Wahl, R.L. Uptake in Supraclavicular Area Fat (“USA-Fat”): Description on 18F-FDG PET/CT. J. Nucl. Med. 2003, 44, 170–176. [Google Scholar]
- Hany, T.F.; Gharehpapagh, E.; Kamel, E.M.; Buck, A.; Himms-Hagen, J.; von Schulthess, G.K. Brown Adipose Tissue: A Factor to Consider in Symmetrical Tracer Uptake in the Neck and Upper Chest Region. Eur. J. Nucl. Med. 2002, 29, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.H.; Tovar, J.P.; Pavlova, Z.; Smith, M.L.; Gilsanz, V. Unequivocal Identification of Brown Adipose Tissue in a Human Infant. J. Magn. Reson. Imaging 2012, 35, 938–942. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.H.; Yin, L.; Aggabao, P.C.; Perkins, T.G.; Chia, J.M.; Gilsanz, V. Comparison of Brown and White Adipose Tissues in Infants and Children with Chemical-Shift-Encoded Water-Fat MRI: BAT and WAT in Infants and Children. J. Magn. Reson. Imaging 2013, 38, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.H.; Wu, T.-W.; Yin, L.; Kim, M.S.; Chia, J.M.; Perkins, T.G.; Gilsanz, V. MRI Detection of Brown Adipose Tissue with Low Fat Content in Newborns with Hypothermia. Magn. Reson. Imaging 2014, 32, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Iris Chen, Y.-C.; Cypess, A.M.; Chen, Y.-C.; Palmer, M.; Kolodny, G.; Kahn, C.R.; Kwong, K.K. Measurement of Human Brown Adipose Tissue Volume and Activity Using Anatomic MR Imaging and Functional MR Imaging. J. Nucl. Med. 2013, 54, 1584–1587. [Google Scholar] [CrossRef] [Green Version]
- Franz, D.; Weidlich, D.; Freitag, F.; Holzapfel, C.; Drabsch, T.; Baum, T.; Eggers, H.; Witte, A.; Rummeny, E.J.; Hauner, H.; et al. Association of Proton Density Fat Fraction in Adipose Tissue with Imaging-Based and Anthropometric Obesity Markers in Adults. Int. J. Obes. 2018, 42, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Muzik, O.; Mangner, T.J.; Leonard, W.R.; Kumar, A.; Janisse, J.; Granneman, J.G. 15O PET Measurement of Blood Flow and Oxygen Consumption in Cold-Activated Human Brown Fat. J. Nucl. Med. 2013, 54, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frontini, A.; Cinti, S. Distribution and Development of Brown Adipocytes in the Murine and Human Adipose Organ. Cell Metab. 2010, 11, 253–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, J.; Chalmers, J.; Morris, D.E.; Robinson, L.; Budge, H.; Symonds, M.E. The Use of Infrared Thermography in the Measurement and Characterization of Brown Adipose Tissue Activation. Temperature 2018, 5, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Hamaoka, T.; Nirengi, S.; Fuse, S.; Amagasa, S.; Kime, R.; Kuroiwa, M.; Endo, T.; Sakane, N.; Matsushita, M.; Saito, M.; et al. Near-Infrared Time-Resolved Spectroscopy for Assessing Brown Adipose Tissue Density in Humans: A Review. Front. Endocrinol. 2020, 11, 261. [Google Scholar] [CrossRef] [PubMed]
- Pfannenberg, C.; Werner, M.K.; Ripkens, S.; Stef, I.; Deckert, A.; Schmadl, M.; Reimold, M.; Haring, H.-U.; Claussen, C.D.; Stefan, N. Impact of Age on the Relationships of Brown Adipose Tissue With Sex and Adiposity in Humans. Diabetes 2010, 59, 1789–1793. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, V.; Routhier-Labadie, A.; Bellemare, W.; Lakhal-Chaieb, L.; Turcotte, E.; Carpentier, A.C.; Richard, D. Outdoor Temperature, Age, Sex, Body Mass Index, and Diabetic Status Determine the Prevalence, Mass, and Glucose-Uptake Activity of 18F-FDG-Detected BAT in Humans. J. Clin. Endocrinol. Metab. 2011, 96, 192–199. [Google Scholar] [CrossRef]
- Fletcher, L.A.; Kim, K.; Leitner, B.P.; Cassimatis, T.M.; O’Mara, A.E.; Johnson, J.W.; Halprin, M.S.; McGehee, S.M.; Brychta, R.J.; Cypess, A.M.; et al. Sexual Dimorphisms in Adult Human Brown Adipose Tissue. Obesity 2020, 28, 241–246. [Google Scholar] [CrossRef]
- Fuse, S.; Nirengi, S.; Amagasa, S.; Homma, T.; Kime, R.; Endo, T.; Sakane, N.; Matsushita, M.; Saito, M.; Yoneshiro, T.; et al. Brown Adipose Tissue Density Measured by Near-Infrared Time-Resolved Spectroscopy in Japanese, across a Wide Age Range. J. Biomed. Opt. 2018, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Yoneshiro, T.; Aita, S.; Matsushita, M.; Okamatsu-Ogura, Y.; Kameya, T.; Kawai, Y.; Miyagawa, M.; Tsujisaki, M.; Saito, M. Age-Related Decrease in Cold-Activated Brown Adipose Tissue and Accumulation of Body Fat in Healthy Humans. Obesity 2011, 19, 1755–1760. [Google Scholar] [CrossRef]
- Keuper, M.; Jastroch, M. The Good and the BAT of Metabolic Sex Differences in Thermogenic Human Adipose Tissue. Mol. Cell. Endocrinol. 2021, 533, 111337. [Google Scholar] [CrossRef] [PubMed]
- Rudnicki, M.; Abdifarkosh, G.; Rezvan, O.; Nwadozi, E.; Roudier, E.; Haas, T.L. Female Mice Have Higher Angiogenesis in Perigonadal Adipose Tissue Than Males in Response to High-Fat Diet. Front. Physiol. 2018, 9, 1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Cuenca, S.; Monjo, M.; Gianotti, M.; Proenza, A.M.; Roca, P. Expression of Mitochondrial Biogenesis-Signaling Factors in Brown Adipocytes Is Influenced Specifically by 17β-Estradiol, Testosterone, and Progesterone. Am. J. Physiol. -Endocrinol. Metab. 2007, 292, E340–E346. [Google Scholar] [CrossRef]
- Löfgren, P.; Hoffstedt, J.; Rydén, M.; Thörne, A.; Holm, C.; Wahrenberg, H.; Arner, P. Major Gender Differences in the Lipolytic Capacity of Abdominal Subcutaneous Fat Cells in Obesity Observed before and after Long-Term Weight Reduction. J. Clin. Endocrinol. Metab. 2002, 87, 764–771. [Google Scholar] [CrossRef] [Green Version]
- Riis-Vestergaard, M.J.; Richelsen, B.; Bruun, J.M.; Li, W.; Hansen, J.B.; Pedersen, S.B. Beta-1 and Not Beta-3 Adrenergic Receptors May Be the Primary Regulator of Human Brown Adipocyte Metabolism. J. Clin. Endocrinol. Metab. 2020, 105, e994–e1005. [Google Scholar] [CrossRef]
- Kaikaew, K.; van den Beukel, J.C.; Neggers, S.J.C.M.M.; Themmen, A.P.N.; Visser, J.A.; Grefhorst, A. Sex Difference in Cold Perception and Shivering Onset upon Gradual Cold Exposure. J. Therm. Biol. 2018, 77, 137–144. [Google Scholar] [CrossRef]
- Choi, D.K.; Oh, T.S.; Choi, J.-W.; Mukherjee, R.; Wang, X.; Liu, H.; Yun, J.W. Gender Difference in Proteome of Brown Adipose Tissues between Male and Female Rats Exposed to a High Fat Diet. Cell Physiol. Biochem. 2011, 28, 933–948. [Google Scholar] [CrossRef]
- Nookaew, I.; Svensson, P.-A.; Jacobson, P.; Jernås, M.; Taube, M.; Larsson, I.; Andersson-Assarsson, J.C.; Sjöström, L.; Froguel, P.; Walley, A.; et al. Adipose Tissue Resting Energy Expenditure and Expression of Genes Involved in Mitochondrial Function Are Higher in Women than in Men. J. Clin. Endocrinol. Metab. 2013, 98, E370–E378. [Google Scholar] [CrossRef]
- van den Beukel, J.C.; Grefhorst, A.; Hoogduijn, M.J.; Steenbergen, J.; Mastroberardino, P.G.; Dor, F.J.M.F.; Themmen, A.P.N. Women Have More Potential to Induce Browning of Perirenal Adipose Tissue than Men: Browning of Perirenal Fat in Women. Obesity 2015, 23, 1671–1679. [Google Scholar] [CrossRef]
- Norheim, F.; Hasin-Brumshtein, Y.; Vergnes, L.; Chella Krishnan, K.; Pan, C.; Seldin, M.M.; Hui, S.T.; Mehrabian, M.; Zhou, Z.; Gupta, S.; et al. Gene-by-Sex Interactions in Mitochondrial Functions and Cardio-Metabolic Traits. Cell Metab. 2019, 29, 932–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persichetti, A.; Sciuto, R.; Rea, S.; Basciani, S.; Lubrano, C.; Mariani, S.; Ulisse, S.; Nofroni, I.; Maini, C.L.; Gnessi, L. Prevalence, Mass, and Glucose-Uptake Activity of 18F-FDG-Detected Brown Adipose Tissue in Humans Living in a Temperate Zone of Italy. PLoS ONE 2013, 8, e63391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecoultre, V.; Ravussin, E. Brown Adipose Tissue and Aging. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Graja, A.; Schulz, T.J. Mechanisms of Aging-Related Impairment of Brown Adipocyte Development and Function. Gerontology 2014, 61, 211–217. [Google Scholar] [CrossRef]
- Rodgers, A.; Sferruzzi-Perri, A.N. Developmental Programming of Offspring Adipose Tissue Biology and Obesity Risk. Int. J. Obes. 2021, 45, 1170–1192. [Google Scholar] [CrossRef]
- Zoico, E.; Rubele, S.; De Caro, A.; Nori, N.; Mazzali, G.; Fantin, F.; Rossi, A.; Zamboni, M. Brown and Beige Adipose Tissue and Aging. Front. Endocrinol. 2019, 10, 368. [Google Scholar] [CrossRef] [Green Version]
- Lenaz, G. Role of Mitochondria in Oxidative Stress and Ageing. Biochim. Biophys. Acta (BBA)-Bioenerg. 1998, 1366, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Bratic, A.; Larsson, N.-G. The Role of Mitochondria in Aging. J. Clin. Investig. 2013, 123, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Khanh, V.C.; Zulkifli, A.F.; Tokunaga, C.; Yamashita, T.; Hiramatsu, Y.; Ohneda, O. Aging Impairs Beige Adipocyte Differentiation of Mesenchymal Stem Cells via the Reduced Expression of Sirtuin 1. Biochem. Biophys. Res. Commun. 2018, 500, 682–690. [Google Scholar] [CrossRef]
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected Evidence for Active Brown Adipose Tissue in Adult Humans. Am. J. Physiol. -Endocrinol. Metab. 2007, 293, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Bahler, L.; Verberne, H.J.; Admiraal, W.M.; Stok, W.J.; Soeters, M.R.; Hoekstra, J.B.; Holleman, F. Differences in Sympathetic Nervous Stimulation of Brown Adipose Tissue Between the Young and Old, and the Lean and Obese. J. Nucl. Med. 2016, 57, 372–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneshiro, T.; Ogawa, T.; Okamoto, N.; Matsushita, M.; Aita, S.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Impact of UCP1 and Β3AR Gene Polymorphisms on Age-Related Changes in Brown Adipose Tissue and Adiposity in Humans. Int. J. Obes. 2013, 37, 993–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sul, O.; Hyun, H.; Rajasekaran, M.; Suh, J.; Choi, H. Estrogen Enhances Browning in Adipose Tissue by M2 Macrophage Polarization via Heme Oxygenase-1. J. Cell. Physiol. 2021, 236, 1875–1888. [Google Scholar] [CrossRef]
- Fan, W.; Yanase, T.; Nomura, M.; Okabe, T.; Goto, K.; Sato, T.; Kawano, H.; Kato, S.; Nawata, H. Androgen Receptor Null Male Mice Develop Late-Onset Obesity Caused by Decreased Energy Expenditure and Lipolytic Activity but Show Normal Insulin Sensitivity With High Adiponectin Secretion. Diabetes 2005, 54, 1000–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soumano, K. Glucocorticoids Inhibit the Transcriptional Response of the Uncoupling Protein-1 Gene to Adrenergic Stimulation in a Brown Adipose Cell Line. Mol. Cell. Endocrinol. 2000, 165, 7–15. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Takahashi, N.; Yasubuchi, M.; Kim, Y.-I.; Hashizaki, H.; Kim, M.-J.; Sakamoto, T.; Goto, T.; Kawada, T. Triiodothyronine Induces UCP-1 Expression and Mitochondrial Biogenesis in Human Adipocytes. Am. J. Physiol.-Cell Physiol. 2012, 302, C463–C472. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Sánchez, N.; Moreno-Navarrete, J.M.; Contreras, C.; Rial-Pensado, E.; Fernø, J.; Nogueiras, R.; Diéguez, C.; Fernández-Real, J.-M.; López, M. Thyroid Hormones Induce Browning of White Fat. J. Endocrinol. 2017, 232, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Schulz, T.J.; Huang, T.L.; Tran, T.T.; Zhang, H.; Townsend, K.L.; Shadrach, J.L.; Cerletti, M.; McDougall, L.E.; Giorgadze, N.; Tchkonia, T.; et al. Identification of Inducible Brown Adipocyte Progenitors Residing in Skeletal Muscle and White Fat. Proc. Natl. Acad. Sci. USA 2011, 108, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-H.; Petkova, A.P.; Mottillo, E.P.; Granneman, J.G. In Vivo Identification of Bipotential Adipocyte Progenitors Recruited by Β3-Adrenoceptor Activation and High-Fat Feeding. Cell Metab. 2012, 15, 480–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, R.; Rodeheffer, M.S. Characterization of the Adipocyte Cellular Lineage in Vivo. Nat. Cell Biol. 2013, 15, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Holzman, M.A.; Ryckman, A.; Finkelstein, T.M.; Landry-Truchon, K.; Schindler, K.A.; Bergmann, J.M.; Jeannotte, L.; Mansfield, J.H. HOXA5 Participates in Brown Adipose Tissue and Epaxial Skeletal Muscle Patterning and in Brown Adipocyte Differentiation. Front. Cell Dev. Biol. 2021, 9, 632303. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Wang, G.; Diao, Y.; Long, Y.; Fu, X.; Weng, M.; Zhou, L.; Sun, K.; Cheung, T.H.; Ip, N.Y.; et al. A Molecular Switch Regulating Cell Fate Choice between Muscle Progenitor Cells and Brown Adipocytes. Dev. Cell 2017, 41, 382–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmons, J.A.; Wennmalm, K.; Larsson, O.; Walden, T.B.; Lassmann, T.; Petrovic, N.; Hamilton, D.L.; Gimeno, R.E.; Wahlestedt, C.; Baar, K.; et al. Myogenic Gene Expression Signature Establishes That Brown and White Adipocytes Originate from Distinct Cell Lineages. Proc. Natl. Acad. Sci. USA 2007, 104, 4401–4406. [Google Scholar] [CrossRef] [Green Version]
- Forner, F.; Kumar, C.; Luber, C.A.; Fromme, T.; Klingenspor, M.; Mann, M. Proteome Differences between Brown and White Fat Mitochondria Reveal Specialized Metabolic Functions. Cell Metab. 2009, 10, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, L.Z.; Shinoda, K.; Ohno, H.; Scheel, D.W.; Tomoda, E.; Ruiz, L.; Hu, H.; Wang, L.; Pavlova, Z.; Gilsanz, V.; et al. Human BAT Possesses Molecular Signatures That Resemble Beige/Brite Cells. PLoS ONE 2012, 7, e49452. [Google Scholar] [CrossRef]
- Xue, R.; Lynes, M.D.; Dreyfuss, J.M.; Shamsi, F.; Schulz, T.J.; Zhang, H.; Huang, T.L.; Townsend, K.L.; Li, Y.; Takahashi, H.; et al. Clonal Analyses and Gene Profiling Identify Genetic Biomarkers of the Thermogenic Potential of Human Brown and White Preadipocytes. Nat. Med. 2015, 21, 760–768. [Google Scholar] [CrossRef]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scimè, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 Controls a Brown Fat/Skeletal Muscle Switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Lidell, M.E.; Betz, M.J.; Leinhard, O.D.; Heglind, M.; Elander, L.; Slawik, M.; Mussack, T.; Nilsson, D.; Romu, T.; Nuutila, P.; et al. Evidence for Two Types of Brown Adipose Tissue in Humans. Nat. Med. 2013, 19, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Xie, H.; Mori, M.A.; Alexander, R.; Yuan, B.; Hattangadi, S.M.; Liu, Q.; Kahn, C.R.; Lodish, H.F. Mir193b–365 Is Essential for Brown Fat Differentiation. Nat. Cell Biol. 2011, 13, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güller, I.; McNaughton, S.; Crowley, T.; Gilsanz, V.; Kajimura, S.; Watt, M.; Russell, A.P. Comparative Analysis of MicroRNA Expression in Mouse and Human Brown Adipose Tissue. BMC Genom. 2015, 16, 820. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Cho, H.; Alexander, R.; Patterson, H.C.; Gu, M.; Lo, K.A.; Xu, D.; Goh, V.J.; Nguyen, L.N.; Chai, X.; et al. MicroRNAs Are Required for the Feature Maintenance and Differentiation of Brown Adipocytes. Diabetes 2014, 63, 4045–4056. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Nakagami, H.; Rodriguez-Araujo, G.; Nimura, K.; Kaneda, Y. Essential Role for MiR-196a in Brown Adipogenesis of White Fat Progenitor Cells. PLoS Biol. 2012, 10, e1001314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Bi, P.; Shan, T.; Yang, X.; Yin, H.; Wang, Y.-X.; Liu, N.; Rudnicki, M.A.; Kuang, S. MiR-133a Regulates Adipocyte Browning In Vivo. PLoS Genet. 2013, 9, e1003626. [Google Scholar] [CrossRef] [Green Version]
- Svensson, P.-A.; Lindberg, K.; Hoffmann, J.M.; Taube, M.; Pereira, M.J.; Mohsen-Kanson, T.; Hafner, A.-L.; Rizell, M.; Palming, J.; Dani, C.; et al. Characterization of Brown Adipose Tissue in the Human Perirenal Depot: BAT in the Human Perirenal Depot. Obesity 2014, 22, 1830–1837. [Google Scholar] [CrossRef]
- Cypess, A.M.; White, A.P.; Vernochet, C.; Schulz, T.J.; Xue, R.; Sass, C.A.; Huang, T.L.; Roberts-Toler, C.; Weiner, L.S.; Sze, C.; et al. Anatomical Localization, Gene Expression Profiling and Functional Characterization of Adult Human Neck Brown Fat. Nat. Med. 2013, 19, 635–639. [Google Scholar] [CrossRef]
- Zuriaga, M.A.; Fuster, J.J.; Gokce, N.; Walsh, K. Humans and Mice Display Opposing Patterns of “Browning” Gene Expression in Visceral and Subcutaneous White Adipose Tissue Depots. Front. Cardiovasc. Med. 2017, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, P.; Arch, J.R.S.; Ashwell, M. Brown Adipose Tissue in the Parametrial Fat Pad of the Mouse. FEBS Lett. 1984, 167, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Haman, F.; Mantha, O.L.; Cheung, S.S.; DuCharme, M.B.; Taber, M.; Blondin, D.P.; McGarr, G.W.; Hartley, G.L.; Hynes, Z.; Basset, F.A. Oxidative Fuel Selection and Shivering Thermogenesis during a 12- and 24-h Cold-Survival Simulation. J. Appl. Physiol. 2016, 120, 640–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bal, N.C.; Maurya, S.K.; Pani, S.; Sethy, C.; Banerjee, A.; Das, S.; Patnaik, S.; Kundu, C.N. Mild Cold Induced Thermogenesis: Are BAT and Skeletal Muscle Synergistic Partners? Biosci. Rep. 2017, 37, BSR20171087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, P.A.; Finlin, B.S.; Zhu, B.; Rasouli, N.; McGehee, R.E.; Westgate, P.M.; Dupont-Versteegden, E.E. The Effects of Temperature and Seasons on Subcutaneous White Adipose Tissue in Humans: Evidence for Thermogenic Gene Induction. J. Clin. Endocrinol. Metab. 2014, 99, E2772–E2779. [Google Scholar] [CrossRef] [PubMed]
- Betz, M.J.; Slawik, M.; Lidell, M.E.; Osswald, A.; Heglind, M.; Nilsson, D.; Lichtenauer, U.D.; Mauracher, B.; Mussack, T.; Beuschlein, F.; et al. Presence of Brown Adipocytes in Retroperitoneal Fat From Patients With Benign Adrenal Tumors: Relationship With Outdoor Temperature. J. Clin. Endocrinol. Metab. 2013, 98, 4097–4104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Lans, A.A.J.J.; Hoeks, J.; Brans, B.; Vijgen, G.H.E.J.; Visser, M.G.W.; Vosselman, M.J.; Hansen, J.; Jörgensen, J.A.; Wu, J.; Mottaghy, F.M.; et al. Cold Acclimation Recruits Human Brown Fat and Increases Nonshivering Thermogenesis. J. Clin. Investig. 2013, 123, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Weir, G.; Ramage, L.E.; Akyol, M.; Rhodes, J.K.; Kyle, C.J.; Fletcher, A.M.; Craven, T.H.; Wakelin, S.J.; Drake, A.J.; Gregoriades, M.-L.; et al. Substantial Metabolic Activity of Human Brown Adipose Tissue during Warm Conditions and Cold-Induced Lipolysis of Local Triglycerides. Cell Metab. 2018, 27, 1348–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellet, V.; Labbé, S.M.; Blondin, D.P.; Phoenix, S.; Guérin, B.; Haman, F.; Turcotte, E.E.; Richard, D.; Carpentier, A.C. Brown Adipose Tissue Oxidative Metabolism Contributes to Energy Expenditure during Acute Cold Exposure in Humans. J. Clin. Investig. 2012, 122, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Blondin, D.P.; Labbé, S.M.; Noll, C.; Kunach, M.; Phoenix, S.; Guérin, B.; Turcotte, É.E.; Haman, F.; Richard, D.; Carpentier, A.C. Selective Impairment of Glucose but Not Fatty Acid or Oxidative Metabolism in Brown Adipose Tissue of Subjects with Type 2 Diabetes. Diabetes 2015, 64, 2388–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondin, D.P.; Carpentier, A.C. The Role of BAT in Cardiometabolic Disorders and Aging. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 497–513. [Google Scholar] [CrossRef]
- Suchacki, K.J.; Stimson, R.H. Nutritional Regulation of Human Brown Adipose Tissue. Nutrients 2021, 13, 1748. [Google Scholar] [CrossRef]
- Orava, J.; Nuutila, P.; Lidell, M.E.; Oikonen, V.; Noponen, T.; Viljanen, T.; Scheinin, M.; Taittonen, M.; Niemi, T.; Enerbäck, S.; et al. Different Metabolic Responses of Human Brown Adipose Tissue to Activation by Cold and Insulin. Cell Metab. 2011, 14, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Annamalai, P.; Enerbäck, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M.; et al. Brown Adipose Tissue Improves Whole-Body Glucose Homeostasis and Insulin Sensitivity in Humans. Diabetes 2014, 63, 4089–4099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himms-Hagen, J.; Cui, J.; Danforth, E.; Taatjes, D.J.; Lang, S.S.; Waters, B.L.; Claus, T.H. Effect of CL-316,243, a Thermogenic Beta 3-Agonist, on Energy Balance and Brown and White Adipose Tissues in Rats. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 1994, 266, R1371–R1382. [Google Scholar] [CrossRef] [PubMed]
- Granneman, J.G.; Li, P.; Zhu, Z.; Lu, Y. Metabolic and Cellular Plasticity in White Adipose Tissue I: Effects of β 3 -Adrenergic Receptor Activation. Am. J. Physiol. -Endocrinol. Metab. 2005, 289, E608–E616. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.M.; Toubro, S.; van Baak, M.A.; Gottesdiener, K.M.; Larson, P.; Saris, W.H.; Astrup, A. Effect of a 28-d Treatment with L-796568, a Novel Β3-Adrenergic Receptor Agonist, on Energy Expenditure and Body Composition in Obese Men. Am. J. Clin. Nutr. 2002, 76, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyer, C.; Tataranni, P.A.; Snitker, S.; Danforth, E.; Ravussin, E. Increase in Insulin Action and Fat Oxidation after Treatment with CL 316,243, a Highly Selective Beta3-Adrenoceptor Agonist in Humans. Diabetes 1998, 47, 1555–1561. [Google Scholar] [CrossRef]
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Elía, E.F.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of Human Brown Adipose Tissue by a Β3-Adrenergic Receptor Agonist. Cell Metab. 2015, 21, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Blondin, D.P.; Nielsen, S.; Kuipers, E.N.; Severinsen, M.C.; Jensen, V.H.; Miard, S.; Jespersen, N.Z.; Kooijman, S.; Boon, M.R.; Fortin, M.; et al. Human Brown Adipocyte Thermogenesis Is Driven by Β2-AR Stimulation. Cell Metab. 2020, 32, 287–300. [Google Scholar] [CrossRef]
- Carey, A.L.; Formosa, M.F.; Van Every, B.; Bertovic, D.; Eikelis, N.; Lambert, G.W.; Kalff, V.; Duffy, S.J.; Cherk, M.H.; Kingwell, B.A. Ephedrine Activates Brown Adipose Tissue in Lean but Not Obese Humans. Diabetologia 2013, 56, 147–155. [Google Scholar] [CrossRef]
- O’Mara, A.E.; Johnson, J.W.; Linderman, J.D.; Brychta, R.J.; McGehee, S.; Fletcher, L.A.; Fink, Y.A.; Kapuria, D.; Cassimatis, T.M.; Kelsey, N.; et al. Chronic Mirabegron Treatment Increases Human Brown Fat, HDL Cholesterol, and Insulin Sensitivity. J. Clin. Investig. 2020, 130, 2209–2219. [Google Scholar] [CrossRef]
- Warner, A.; Mittag, J. Breaking BAT: Can Browning Create a Better White? J. Endocrinol. 2016, 228, R19–R29. [Google Scholar] [CrossRef] [Green Version]
- Kaisanlahti, A.; Glumoff, T. Browning of White Fat: Agents and Implications for Beige Adipose Tissue to Type 2 Diabetes. J. Physiol. Biochem. 2019, 75, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Markan, K.R.; Naber, M.C.; Ameka, M.K.; Anderegg, M.D.; Mangelsdorf, D.J.; Kliewer, S.A.; Mohammadi, M.; Potthoff, M.J. Circulating FGF21 Is Liver Derived and Enhances Glucose Uptake During Refeeding and Overfeeding. Diabetes 2014, 63, 4057–4063. [Google Scholar] [CrossRef] [Green Version]
- Hondares, E.; Gallego-Escuredo, J.M.; Flachs, P.; Frontini, A.; Cereijo, R.; Goday, A.; Perugini, J.; Kopecky, P.; Giralt, M.; Cinti, S.; et al. Fibroblast Growth Factor-21 Is Expressed in Neonatal and Pheochromocytoma-Induced Adult Human Brown Adipose Tissue. Metabolism 2014, 63, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Fisher, f.M.; Kleiner, S.; Douris, N.; Fox, E.C.; Mepani, R.J.; Verdeguer, F.; Wu, J.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E.; et al. FGF21 Regulates PGC-1 and Browning of White Adipose Tissues in Adaptive Thermogenesis. Genes Dev. 2012, 26, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlein, C.; Talukdar, S.; Heine, M.; Fischer, A.W.; Krott, L.M.; Nilsson, S.K.; Brenner, M.B.; Heeren, J.; Scheja, L. FGF21 Lowers Plasma Triglycerides by Accelerating Lipoprotein Catabolism in White and Brown Adipose Tissues. Cell Metab. 2016, 23, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, S.; Zhou, Y.; Li, D.; Rossulek, M.; Dong, J.; Somayaji, V.; Weng, Y.; Clark, R.; Lanba, A.; Owen, B.M.; et al. A Long-Acting FGF21 Molecule, PF-05231023, Decreases Body Weight and Improves Lipid Profile in Non-Human Primates and Type 2 Diabetic Subjects. Cell Metab. 2016, 23, 427–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaich, G.; Chien, J.Y.; Fu, H.; Glass, L.C.; Deeg, M.A.; Holland, W.L.; Kharitonenkov, A.; Bumol, T.; Schilske, H.K.; Moller, D.E. The Effects of LY2405319, an FGF21 Analog, in Obese Human Subjects with Type 2 Diabetes. Cell Metab. 2013, 18, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Kelton, C.M.L.; Guo, J.J.; Bian, B.; Heaton, P.C. Treatment of Obesity: Pharmacotherapy Trends in the United States from 1999 to 2010: Pharmacotherapy Treatment of Obesity. Obesity 2015, 23, 1721–1728. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.D.; Boström, P.; O’Sullivan, J.F.; Schinzel, R.T.; Lewis, G.D.; Dejam, A.; Lee, Y.-K.; Palma, M.J.; Calhoun, S.; Georgiadi, A.; et al. β-Aminoisobutyric Acid Induces Browning of White Fat and Hepatic β-Oxidation and Is Inversely Correlated with Cardiometabolic Risk Factors. Cell Metab. 2014, 19, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vegiopoulos, A.; Muller-Decker, K.; Strzoda, D.; Schmitt, I.; Chichelnitskiy, E.; Ostertag, A.; Diaz, M.B.; Rozman, J.; Hrabe de Angelis, M.; Nusing, R.M.; et al. Cyclooxygenase-2 Controls Energy Homeostasis in Mice by de Novo Recruitment of Brown Adipocytes. Science 2010, 328, 1158–1161. [Google Scholar] [CrossRef]
- Qian, S.-W.; Tang, Y.; Li, X.; Liu, Y.; Zhang, Y.-Y.; Huang, H.-Y.; Xue, R.-D.; Yu, H.-Y.; Guo, L.; Gao, H.-D.; et al. BMP4-Mediated Brown Fat-like Changes in White Adipose Tissue Alter Glucose and Energy Homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, E798–E807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafson, B.; Hammarstedt, A.; Hedjazifar, S.; Hoffmann, J.M.; Svensson, P.-A.; Grimsby, J.; Rondinone, C.; Smith, U. BMP4 and BMP Antagonists Regulate Human White and Beige Adipogenesis. Diabetes 2015, 64, 1670–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsen, M.; Raschke, S.; Tennagels, N.; Schwahn, U.; Jelenik, T.; Roden, M.; Romacho, T.; Eckel, J. BMP4 and BMP7 Induce the White-to-Brown Transition of Primary Human Adipose Stem Cells. Am. J. Physiol.-Cell Physiol. 2014, 306, C431–C440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, M.; Reddy, S.T.; Vergnes, L.; Pervin, S.; Grijalva, V.; Stout, D.; David, J.; Li, X.; Tomasian, V.; Reid, C.B.; et al. Follistatin Promotes Adipocyte Differentiation, Browning, and Energy Metabolism. J. Lipid. Res. 2014, 55, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Braga, M.; Reddy, S.T.; Lee, S.-J.; Parveen, M.; Grijalva, V.; Vergnes, L.; Pervin, S. Follistatin Targets Distinct Pathways To Promote Brown Adipocyte Characteristics in Brown and White Adipose Tissues. Endocrinology 2017, 158, 1217–1230. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.; Pervin, S.; Norris, K.; Bhasin, S.; Singh, R. Inhibition of in Vitro and in Vivo Brown Fat Differentiation Program by Myostatin: Brown Fat and Myostatin. Obesity 2013, 21, 1180–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-S.; Kim, E.-S.; Jung, J.-E.; Marciano, D.P.; Jo, A.; Koo, J.Y.; Choi, S.Y.; Yang, Y.R.; Jang, H.-J.; Kim, E.-K.; et al. PPARγ Antagonist Gleevec Improves Insulin Sensitivity and Promotes the Browning of White Adipose Tissue. Diabetes 2016, 65, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Ohno, H.; Shinoda, K.; Spiegelman, B.M.; Kajimura, S. PPARγ Agonists Induce a White-to-Brown Fat Conversion through Stabilization of PRDM16 Protein. Cell Metab. 2012, 15, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Loh, R.K.C.; Formosa, M.F.; Eikelis, N.; Bertovic, D.A.; Anderson, M.J.; Barwood, S.A.; Nanayakkara, S.; Cohen, N.D.; La Gerche, A.; Reutens, A.T.; et al. Pioglitazone Reduces Cold-Induced Brown Fat Glucose Uptake despite Induction of Browning in Cultured Human Adipocytes: A Randomised, Controlled Trial in Humans. Diabetologia 2018, 61, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Pi-Sunyer, X.; Astrup, A.; Fujioka, K.; Greenway, F.; Halpern, A.; Krempf, M.; Lau, D.C.W.; le Roux, C.W.; Violante Ortiz, R.; Jensen, C.B.; et al. A Randomized, Controlled Trial of 3.0 Mg of Liraglutide in Weight Management. N. Engl. J. Med. 2015, 373, 11–22. [Google Scholar] [CrossRef]
- Beiroa, D.; Imbernon, M.; Gallego, R.; Senra, A.; Herranz, D.; Villarroya, F.; Serrano, M.; Ferno, J.; Salvador, J.; Escalada, J.; et al. GLP-1 Agonism Stimulates Brown Adipose Tissue Thermogenesis and Browning Through Hypothalamic AMPK. Diabetes 2014, 63, 3346–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teodoro, J.S.; Zouhar, P.; Flachs, P.; Bardova, K.; Janovska, P.; Gomes, A.P.; Duarte, F.V.; Varela, A.T.; Rolo, A.P.; Palmeira, C.M.; et al. Enhancement of Brown Fat Thermogenesis Using Chenodeoxycholic Acid in Mice. Int. J. Obes. 2014, 38, 1027–1034. [Google Scholar] [CrossRef]
- Broeders, E.P.M.; Nascimento, E.B.M.; Havekes, B.; Brans, B.; Roumans, K.H.M.; Tailleux, A.; Schaart, G.; Kouach, M.; Charton, J.; Deprez, B.; et al. The Bile Acid Chenodeoxycholic Acid Increases Human Brown Adipose Tissue Activity. Cell Metab. 2015, 22, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Song, N.-J.; Choi, S.; Rajbhandari, P.; Chang, S.-H.; Kim, S.; Vergnes, L.; Kwon, S.-M.; Yoon, J.-H.; Lee, S.; Ku, J.-M.; et al. Prdm4 Induction by the Small Molecule Butein Promotes White Adipose Tissue Browning. Nat. Chem. Biol. 2016, 12, 479–481. [Google Scholar] [CrossRef]
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin Induces Browning of White Adipose Tissue and Counters Obesity by Activating TRPV1 Channel-Dependent Mechanisms: TRPV1 Activates Browning of WAT to Counter Obesity. Br. J. Pharmacol. 2016, 173, 2369–2389. [Google Scholar] [CrossRef]
- Yoneshiro, T.; Aita, S.; Matsushita, M.; Kayahara, T.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Recruited Brown Adipose Tissue as an Antiobesity Agent in Humans. J. Clin. Investig. 2013, 123, 3404–3408. [Google Scholar] [CrossRef] [Green Version]
- Yoneshiro, T.; Matsushita, M.; Hibi, M.; Tone, H.; Takeshita, M.; Yasunaga, K.; Katsuragi, Y.; Kameya, T.; Sugie, H.; Saito, M. Tea Catechin and Caffeine Activate Brown Adipose Tissue and Increase Cold-Induced Thermogenic Capacity in Humans. Am. J. Clin. Nutr. 2017, 105, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Carriere, A.; Jeanson, Y.; Berger-Muller, S.; Andre, M.; Chenouard, V.; Arnaud, E.; Barreau, C.; Walther, R.; Galinier, A.; Wdziekonski, B.; et al. Browning of White Adipose Cells by Intermediate Metabolites: An Adaptive Mechanism to Alleviate Redox Pressure. Diabetes 2014, 63, 3253–3265. [Google Scholar] [CrossRef] [Green Version]
- Stanford, K.I.; Middelbeek, R.J.W.; Goodyear, L.J. Exercise Effects on White Adipose Tissue: Beiging and Metabolic Adaptations. Diabetes 2015, 64, 2361–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevellin, E.; Scorzeto, M.; Olivieri, M.; Granzotto, M.; Valerio, A.; Tedesco, L.; Fabris, R.; Serra, R.; Quarta, M.; Reggiani, C.; et al. Exercise Training Induces Mitochondrial Biogenesis and Glucose Uptake in Subcutaneous Adipose Tissue Through ENOS-Dependent Mechanisms. Diabetes 2014, 63, 2800–2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Lee, P.; Linderman, J.D.; Smith, S.; Brychta, R.J.; Wang, J.; Idelson, C.; Perron, R.M.; Werner, C.D.; Phan, G.Q.; Kammula, U.S.; et al. Irisin and FGF21 Are Cold-Induced Endocrine Activators of Brown Fat Function in Humans. Cell Metab. 2014, 19, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otero-Díaz, B.; Rodríguez-Flores, M.; Sánchez-Muñoz, V.; Monraz-Preciado, F.; Ordoñez-Ortega, S.; Becerril-Elias, V.; Baay-Guzmán, G.; Obando-Monge, R.; García-García, E.; Palacios-González, B.; et al. Exercise Induces White Adipose Tissue Browning Across the Weight Spectrum in Humans. Front. Physiol. 2018, 9, 1781. [Google Scholar] [CrossRef] [PubMed]
- Hecksteden, A.; Wegmann, M.; Steffen, A.; Kraushaar, J.; Morsch, A.; Ruppenthal, S.; Kaestner, L.; Meyer, T. Irisin and Exercise Training in Humans—Results from a Randomized Controlled Training Trial. BMC Med. 2013, 11, 235. [Google Scholar] [CrossRef] [Green Version]
- Pardo, M.; Crujeiras, A.B.; Amil, M.; Aguera, Z.; Jiménez-Murcia, S.; Baños, R.; Botella, C.; de la Torre, R.; Estivill, X.; Fagundo, A.B.; et al. Association of Irisin with Fat Mass, Resting Energy Expenditure, and Daily Activity in Conditions of Extreme Body Mass Index. Int. J. Endocrinol. 2014, 2014, 857270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viitasalo, A.; Ågren, J.; Venäläinen, T.; Pihlajamäki, J.; Jääskeläinen, J.; Korkmaz, A.; Atalay, M.; Lakka, T.A. Association of Plasma Fatty Acid Composition with Plasma Irisin Levels in Normal Weight and Overweight/Obese Children. Pediatric Obes. 2016, 11, 299–305. [Google Scholar] [CrossRef]
- Hansen, J.S.; Pedersen, B.K.; Xu, G.; Lehmann, R.; Weigert, C.; Plomgaard, P. Exercise-Induced Secretion of FGF21 and Follistatin Are Blocked by Pancreatic Clamp and Impaired in Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2016, 101, 2816–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, J.G.; Murholm, M.; Carey, A.L.; Biensø, R.S.; Basse, A.L.; Allen, T.L.; Hidalgo, J.; Kingwell, B.A.; Febbraio, M.A.; Hansen, J.B.; et al. Role of IL-6 in Exercise Training- and Cold-Induced UCP1 Expression in Subcutaneous White Adipose Tissue. PLoS ONE 2014, 9, e84910. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.R.; Long, J.Z.; White, J.P.; Svensson, K.J.; Lou, J.; Lokurkar, I.; Jedrychowski, M.P.; Ruas, J.L.; Wrann, C.D.; Lo, J.C.; et al. Meteorin-like Is a Hormone That Regulates Immune-Adipose Interactions to Increase Beige Fat Thermogenesis. Cell 2014, 157, 1279–1291. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Choi, E.Y.; Liu, X.; Martin, A.; Wang, C.; Xu, X.; During, M.J. White to Brown Fat Phenotypic Switch Induced by Genetic and Environmental Activation of a Hypothalamic-Adipocyte Axis. Cell Metab. 2011, 14, 324–338. [Google Scholar] [CrossRef] [Green Version]
- Vidal, P.; Stanford, K.I. Exercise-Induced Adaptations to Adipose Tissue Thermogenesis. Front. Endocrinol. 2020, 11, 270. [Google Scholar] [CrossRef]
- Dewal, R.S.; Stanford, K.I. Effects of Exercise on Brown and Beige Adipocytes. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Gharanei, S.; Shabir, K.; Brown, J.E.; Weickert, M.O.; Barber, T.M.; Kyrou, I.; Randeva, H.S. Regulatory MicroRNAs in Brown, Brite and White Adipose Tissue. Cells 2020, 9, 2489. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pan, R.; Pfeifer, A. Regulation of Brown and Beige Fat by MicroRNAs. Pharmacol. Ther. 2017, 170, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Oliverio, M.; Schmidt, E.; Mauer, J.; Baitzel, C.; Hansmeier, N.; Khani, S.; Konieczka, S.; Pradas-Juni, M.; Brodesser, S.; Van, T.-M.; et al. Dicer1–MiR-328–Bace1 Signalling Controls Brown Adipose Tissue Differentiation and Function. Nat. Cell Biol. 2016, 18, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Meakin, P.J.; Harper, A.J.; Hamilton, D.L.; Gallagher, J.; McNeilly, A.D.; Burgess, L.A.; Vaanholt, L.M.; Bannon, K.A.; Latcham, J.; Hussain, I.; et al. Reduction in BACE1 Decreases Body Weight, Protects against Diet-Induced Obesity and Enhances Insulin Sensitivity in Mice. Biochem. J. 2012, 441, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zuo, J.; Zhang, Y.; Xie, Y.; Hu, F.; Chen, L.; Liu, B.; Liu, F. Identification of MiR-106b-93 as a Negative Regulator of Brown Adipocyte Differentiation. Biochem. Biophys. Res. Commun. 2013, 438, 575–580. [Google Scholar] [CrossRef]
- Okamatsu-Ogura, Y.; Matsushita, M.; Bariuan, J.V.; Nagaya, K.; Tsubota, A.; Saito, M. Association of Circulating Exosomal MiR-122 Levels with BAT Activity in Healthy Humans. Sci. Rep. 2019, 9, 13243. [Google Scholar] [CrossRef] [Green Version]
- Karbiener, M.; Pisani, D.F.; Frontini, A.; Oberreiter, L.M.; Lang, E.; Vegiopoulos, A.; Mössenböck, K.; Bernhardt, G.A.; Mayr, T.; Hildner, F.; et al. MicroRNA-26 Family Is Required for Human Adipogenesis and Drives Characteristics of Brown Adipocytes: MicroRNA-26 Family in Human Adipogenesis. Stem. Cells 2014, 32, 1578–1590. [Google Scholar] [CrossRef]
- Zhang, H.; Guan, M.; Townsend, K.L.; Huang, T.L.; An, D.; Yan, X.; Xue, R.; Schulz, T.J.; Winnay, J.; Mori, M.; et al. MicroRNA-455 Regulates Brown Adipogenesis via a Novel HIF1an-AMPK -PGC1α Signaling Network. EMBO Rep. 2015, 16, 1378–1393. [Google Scholar] [CrossRef] [Green Version]
- Giroud, M.; Pisani, D.F.; Karbiener, M.; Barquissau, V.; Ghandour, R.A.; Tews, D.; Fischer-Posovszky, P.; Chambard, J.-C.; Knippschild, U.; Niemi, T.; et al. MiR-125b Affects Mitochondrial Biogenesis and Impairs Brite Adipocyte Formation and Function. Mol. Metab. 2016, 5, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wang, M.; Xiao, T.; Yin, B.; He, L.; Meng, W.; Dong, M.; Liu, F. MiR-30 Promotes Thermogenesis and the Development of Beige Fat by Targeting RIP140. Diabetes 2015, 64, 2056–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritah, A.; Christian, M.; Parker, M.G. The Metabolic Coregulator RIP140: An Update. Am. J. Physiol. -Endocrinol. Metab. 2010, 299, E335–E340. [Google Scholar] [CrossRef]
- Pan, D.; Mao, C.; Quattrochi, B.; Friedline, R.H.; Zhu, L.J.; Jung, D.Y.; Kim, J.K.; Lewis, B.; Wang, Y.-X. MicroRNA-378 Controls Classical Brown Fat Expansion to Counteract Obesity. Nat. Commun. 2014, 5, 4725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, S.H.; Blount, M.A.; Corbin, J.D. Mammalian Cyclic Nucleotide Phosphodiesterases: Molecular Mechanisms and Physiological Functions. Physiol. Rev. 2011, 91, 651–690. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Jin, L.; Han, L.; Yuan, Y.; Mu, Q.; Wang, H.; Yang, J.; Ning, G.; Zhou, D.; Zhang, Z. MiR-129-5p Inhibits Adipogenesis through Autophagy and May Be a Potential Biomarker for Obesity. Int. J. Endocrinol. 2019, 2019, 5069578. [Google Scholar] [CrossRef]
- Ge, X.; Sathiakumar, D.; Lua, B.J.G.; Kukreti, H.; Lee, M.; McFarlane, C. Myostatin Signals through MiR-34a to Regulate Fndc5 Expression and Browning of White Adipocytes. Int. J. Obes. 2017, 41, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Trajkovski, M. MiR-27 Orchestrates the Transcriptional Regulation of Brown Adipogenesis. Metabolism 2014, 63, 272–282. [Google Scholar] [CrossRef]
- Karkeni, E.; Astier, J.; Tourniaire, F.; El Abed, M.; Romier, B.; Gouranton, E.; Wan, L.; Borel, P.; Salles, J.; Walrand, S.; et al. Obesity-Associated Inflammation Induces MicroRNA-155 Expression in Adipocytes and Adipose Tissue: Outcome on Adipocyte Function. J. Clin. Endocrinol. Metab. 2016, 101, 1615–1626. [Google Scholar] [CrossRef]
- Trayhurn, P.; Douglas, J.B.; McGuckin, M.M. Brown Adipose Tissue Thermogenesis Is ‘Suppressed’ during Lactation in Mice. Nature 1982, 298, 59–60. [Google Scholar] [CrossRef]
- Rothwell, N.J.; Stock, M.J. A Role for Brown Adipose Tissue in Diet-Induced Thermogenesis. Nature 1979, 281, 31–35. [Google Scholar] [CrossRef]
- Osman, F.; Franklyn, J.A.; Daykin, J.; Chowdhary, S.; Holder, R.L.; Sheppard, M.C.; Gammage, M.D. Heart Rate Variability and Turbulence in Hyperthyroidism before, during, and after Treatment. Am. J. Cardiol. 2004, 94, 465–469. [Google Scholar] [CrossRef]
- Scheen, A.J. Sibutramine on Cardiovascular Outcome. Diabetes Care 2011, 34, S114–S119. [Google Scholar] [CrossRef] [Green Version]
- Grundlingh, J.; Dargan, P.I.; El-Zanfaly, M.; Wood, D.M. 2,4-Dinitrophenol (DNP): A Weight Loss Agent with Significant Acute Toxicity and Risk of Death. J. Med. Toxicol. 2011, 7, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langeveld, M.; Tan, C.Y.; Soeters, M.R.; Virtue, S.; Ambler, G.K.; Watson, L.P.E.; Murgatroyd, P.R.; Chatterjee, V.K.; Vidal-Puig, A. Mild Cold Effects on Hunger, Food Intake, Satiety and Skin Temperature in Humans. Endocr. Connect. 2016, 5, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Dulloo, A.G. The Search for Compounds That Stimulate Thermogenesis in Obesity Management: From Pharmaceuticals to Functional Food Ingredients: Thermogenic Bioactive Food Ingredients. Obes. Rev. 2011, 12, 866–883. [Google Scholar] [CrossRef] [PubMed]
- Tsai, V.W.W.; Husaini, Y.; Sainsbury, A.; Brown, D.A.; Breit, S.N. The MIC-1/GDF15-GFRAL Pathway in Energy Homeostasis: Implications for Obesity, Cachexia, and Other Associated Diseases. Cell Metab. 2018, 28, 353–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmers, T.A.; Fishel, M.L.; Bonetto, A. STAT3 in the Systemic Inflammation of Cancer Cachexia. Semin. Cell Dev. Biol. 2016, 54, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Ozguven, S.; Ones, T.; Yilmaz, Y.; Turoglu, H.T.; Imeryuz, N. The Role of Active Brown Adipose Tissue in Human Metabolism. Eur. J. Nucl. Med. Mol. Imaging 2016, 43, 355–361. [Google Scholar] [CrossRef]
- Blondin, D.P.; Labbé, S.M.; Tingelstad, H.C.; Noll, C.; Kunach, M.; Phoenix, S.; Guérin, B.; Turcotte, É.E.; Carpentier, A.C.; Richard, D.; et al. Increased Brown Adipose Tissue Oxidative Capacity in Cold-Acclimated Humans. J. Clin. Endocrinol. Metab. 2014, 99, E438–E446. [Google Scholar] [CrossRef] [PubMed]
- Ahfeldt, T.; Schinzel, R.T.; Lee, Y.-K.; Hendrickson, D.; Kaplan, A.; Lum, D.H.; Camahort, R.; Xia, F.; Shay, J.; Rhee, E.P.; et al. Programming Human Pluripotent Stem Cells into White and Brown Adipocytes. Nat. Cell Biol. 2012, 14, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Shankar, K.; Kumar, D.; Gupta, S.; Varshney, S.; Rajan, S.; Srivastava, A.; Gupta, A.; Gupta, A.P.; Vishwakarma, A.L.; Gayen, J.R.; et al. Role of Brown Adipose Tissue in Modulating Adipose Tissue Inflammation and Insulin Resistance in High-Fat Diet Fed Mice. Eur. J. Pharmacol. 2019, 854, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.H.; Kim, J.I.; Jeon, Y.G.; Park, J.; Kim, J.B. Effects of Three Thiazolidinediones on Metabolic Regulation and Cold-Induced Thermogenesis. Mol. Cells 2018, 41, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, C.; Liu, J.; Xie, W.; Xu, W.; Liang, F.; Huang, K.; He, X. Intraperitoneal Administration of Follistatin Promotes Adipocyte Browning in High-Fat Diet-Induced Obese Mice. PLoS ONE 2019, 14, e0220310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
BAT distribution in infants, women and men. BAT is stored in a separate interscapular depot in infants, and they lose their brown adipose tissue as they age. In adult humans, most brown adipocytes can be found in the supraclavicular BAT depots in the neck region. Lesser amounts of BAT are found across the aorta, vertebrae, axillary and kidney areas. There is a similar distribution of brown adipose tissue in both women and men. However, women have a greater amount of BAT mass and activity.
Figure 1.
BAT distribution in infants, women and men. BAT is stored in a separate interscapular depot in infants, and they lose their brown adipose tissue as they age. In adult humans, most brown adipocytes can be found in the supraclavicular BAT depots in the neck region. Lesser amounts of BAT are found across the aorta, vertebrae, axillary and kidney areas. There is a similar distribution of brown adipose tissue in both women and men. However, women have a greater amount of BAT mass and activity.

Figure 2.
Therapeutic interventions to increase BAT mass and activity. Acute or chronic cold exposure activates BAT metabolism or induces BAT browning. 3-AR blockers and mTOR inhibitors inhibit the browning of WAT and the activation of BAT. Exercise activates BAT and induces WAT browning via the upregulation of irisin, follistatin (Fst) and FGF21 (fibroblast growth factor 21). Intravenous (IV) administration of proteins, i.e., FGF21, growth and differentiation factor 15 (GDF15), bone morphogenic protein 7 (BMP7) and follistatin (FST), or anti-Activin receptor type 2B (ActRIIB), anti-Myostatin (Mst) antibodies and various pharmaceutical agents, including Liraglutide and thiazolidinediones, upregulate thermogenic genes like UCP1 to induce the browning of WAT. WAT browning is also induced through the use of Gleevec, lactate and Butein. Certain cytokines, catecholamines and metabolites were used for the browning of WAT. IL-6 receptor inhibitors block the browning of WAT. BAT transplantation increases BAT mass and activity. MiRNAs target regulatory mechanisms that regulate thermogenesis and the browning of WAT either positively or negatively. Trans-differentiation of induced pluripotent stem cells and mesenchymal progenitor cells into brown adipocytes by targeting regulatory mechanisms to overexpress thermogenic and adipogenic genes. Diet or consumption of oral supplements known to activate BAT, like capsinoids, tea catechin and BAIBA can be extracted and utilized as an IV injection or as an oral supplement.
Figure 2.
Therapeutic interventions to increase BAT mass and activity. Acute or chronic cold exposure activates BAT metabolism or induces BAT browning. 3-AR blockers and mTOR inhibitors inhibit the browning of WAT and the activation of BAT. Exercise activates BAT and induces WAT browning via the upregulation of irisin, follistatin (Fst) and FGF21 (fibroblast growth factor 21). Intravenous (IV) administration of proteins, i.e., FGF21, growth and differentiation factor 15 (GDF15), bone morphogenic protein 7 (BMP7) and follistatin (FST), or anti-Activin receptor type 2B (ActRIIB), anti-Myostatin (Mst) antibodies and various pharmaceutical agents, including Liraglutide and thiazolidinediones, upregulate thermogenic genes like UCP1 to induce the browning of WAT. WAT browning is also induced through the use of Gleevec, lactate and Butein. Certain cytokines, catecholamines and metabolites were used for the browning of WAT. IL-6 receptor inhibitors block the browning of WAT. BAT transplantation increases BAT mass and activity. MiRNAs target regulatory mechanisms that regulate thermogenesis and the browning of WAT either positively or negatively. Trans-differentiation of induced pluripotent stem cells and mesenchymal progenitor cells into brown adipocytes by targeting regulatory mechanisms to overexpress thermogenic and adipogenic genes. Diet or consumption of oral supplements known to activate BAT, like capsinoids, tea catechin and BAIBA can be extracted and utilized as an IV injection or as an oral supplement.


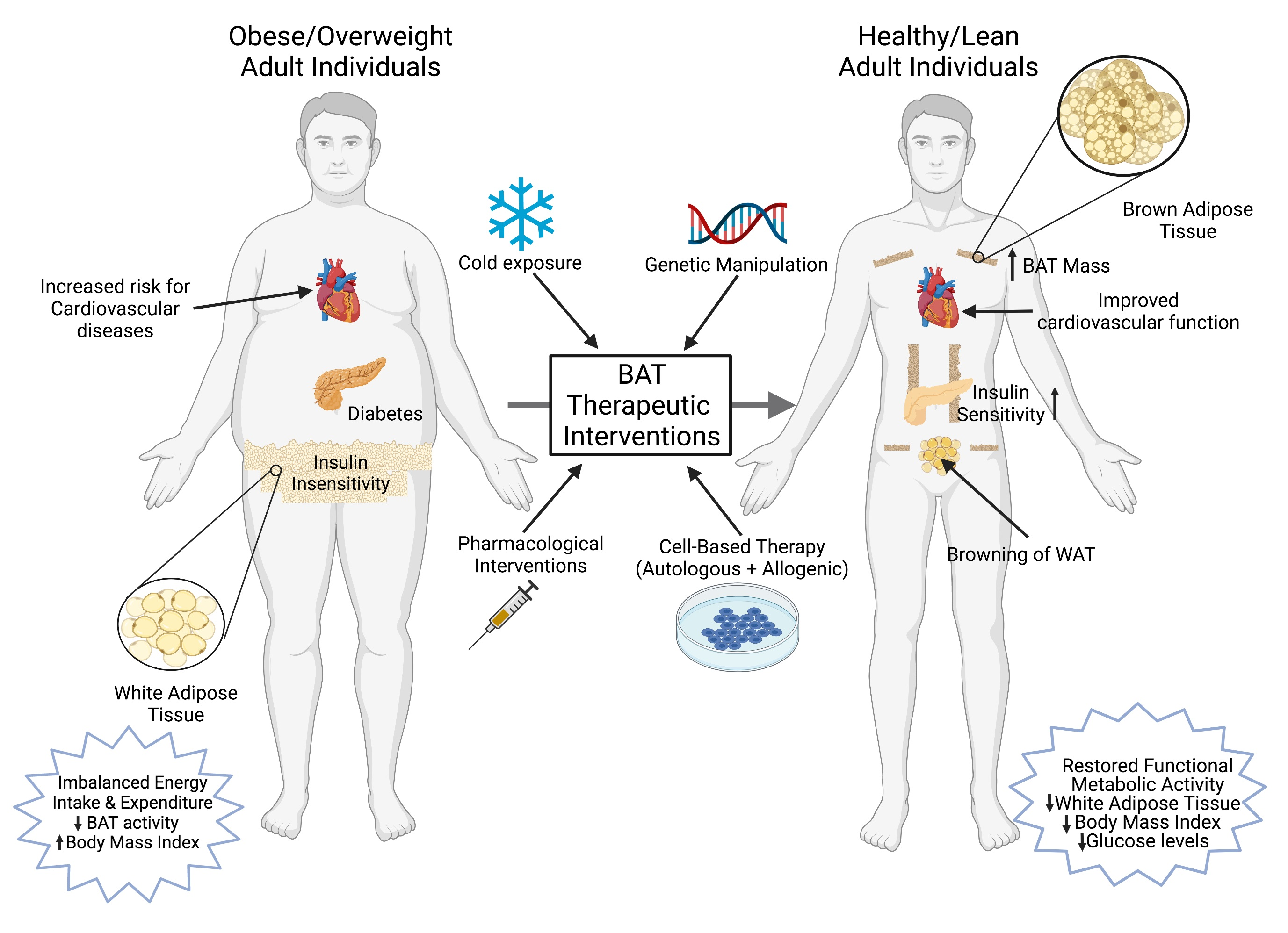{kind=link}
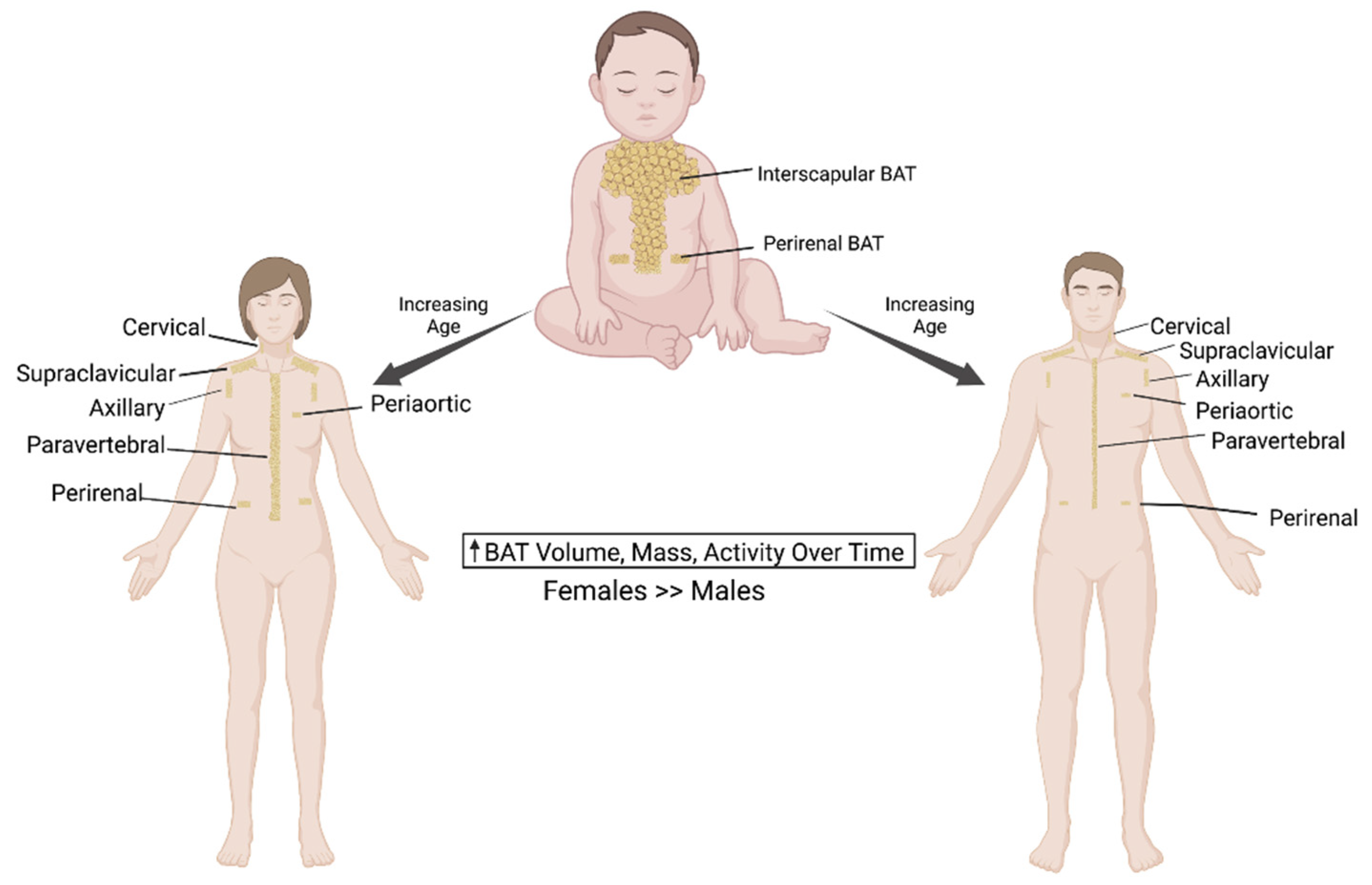{kind=link}
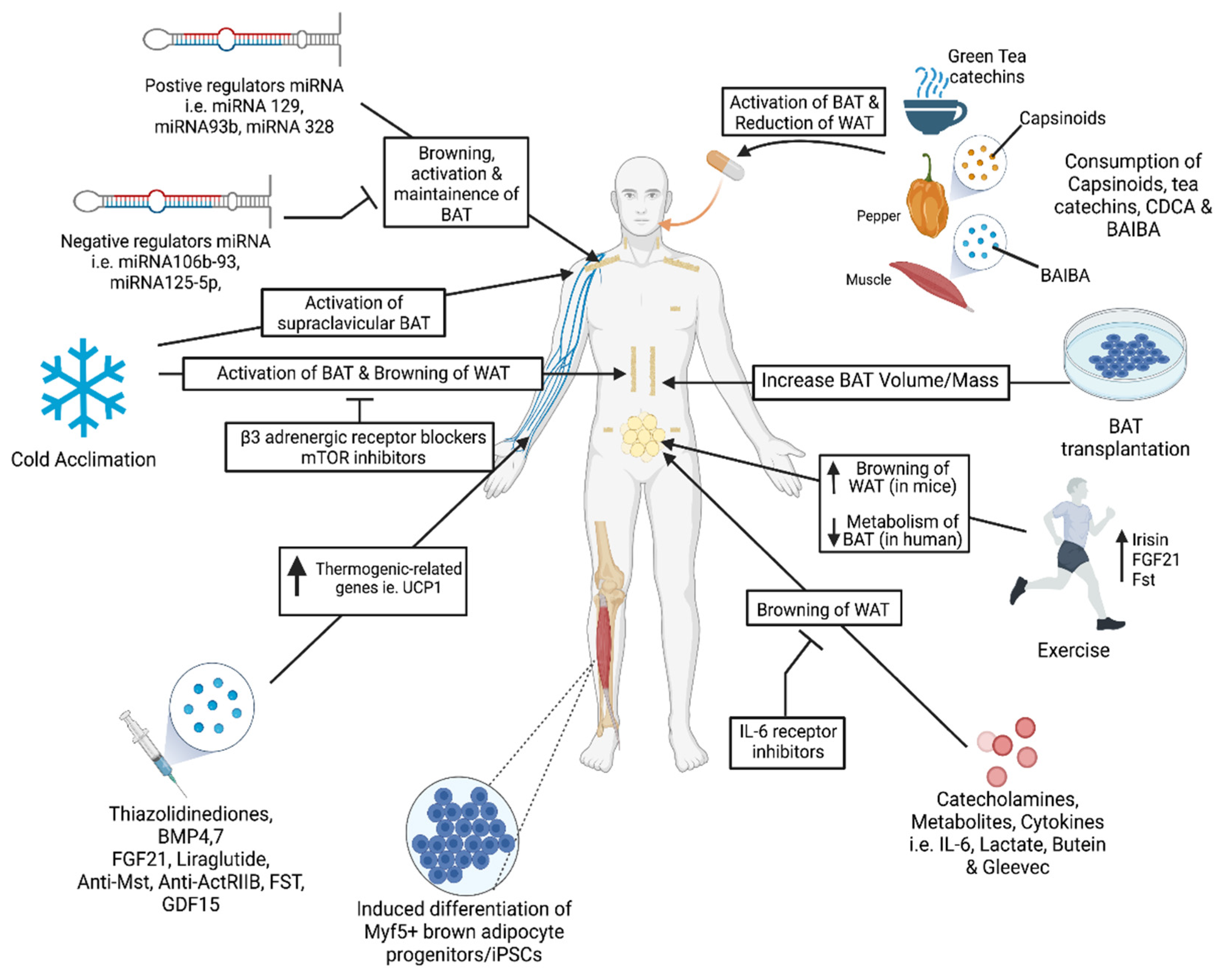{kind=link}
Table 1.
MicroRNAs involved in brown/beige fat development and their targets.
| miRNA | Role in Brown/Beige Fat Development | Target(s) | Species/Reference |
|---|---|---|---|
| miRNA328 | Positive regulation of BAT differentiation | Bace1 | Mouse [155,156] |
| miRNA 193b-365 | Regulation of BAT differentiation | Runx1t1 | Mouse [81] |
| miRNA203 | Promotes BAT adipogenesis | Insig 1, Pdgfr 2 | Mouse [83] |
| miRNA182 | Promotes BAT adipogenesis | Insig 1, Pdgfr 2 | Mouse [83] |
| miRNA129 | Positive regulator of BAT function | Igf2, Egr1 | Human, Mouse [166] |
| miRNA106b | Negative regulator of BAT differentiation | Ppara | Mouse [157] |
| miRNA34a | Negative regulator of BAT and beige adipogenesis | Fgfr1 | Mouse [167] |
| miRNA27b | Negative regulator of BAT and beige adipogenesis | Prdm16, Ppara, Pparg, Creb | Mouse [168] |
| miRNA93b | Negative regulator of BAT differentiation | Ppara | Mouse [157] |
| miRNA196a | Promotes adipose browning | Hoxc8 | Mouse [84] |
| miRNA26a/b | Promotes white and beige adipocyte differentiation | ADAM17, Fbx19 | Human, Mouse [159] |
| miRNA125-5p | Negative regulator of adipose browning and mitochondrial biogenesis | MMP11 | Human [161] |
| miRNA30b/c | Positive regulator of BAT and beige adipocyte development | Rip140 | Mouse [162,163] |
| miRNA455 | Positive regulator of BAT and beige adipose browning | Runx1t1, Necdin, Hif1an | Human, Mouse [160] |
| miRNA378 | Promotes BAT mass and brown adipogenesis but negatively regulates beige adipogenesis | Pde1b | Mouse [164,165] |
| miRNA122 | Negatively regulates BAT activity | CD320, AldoA, BCKDK | Human [158] |
| miRNA133 | Negative regulator of BAT differentiation | Prdm16 | Mouse [85] |
| miRNA155 | Negative regulator of BAT differentiation | C/ebpβ | Mouse, Human [169] |
Table 2.
Adipose browning studies in human and rodent studies.
| Model | Design | Factors Measured | Findings |
|---|---|---|---|
| Human | Tested 17 healthy subjects in 15–16 °C with increasing time to 6 h/day for 10 days with 18F-FDG-PET & abdominal subcutaneous fat biopsy [94] | Glucose uptake in BAT & WAT, energy expenditure, RMR, and NST. | Both females & males in cold exposure: |
| NST , Detectable BAT volume . | |||
| BAT activity , Energy expenditure . | |||
| Human | Single-blinded, placebo-controlled clinical trial. Exposed to 17 °C for 2 h/day for 6 weeks. Subjects with undetectable BAT were placed in cold (n = 12) and control (n = 10) groups. Capsinoid capsules with 9 or 0 mg/day given for 6 weeks. [136] | Body fat content; metabolic activity; EE; 18F-FDG-PET utilized. | After cold exposure: BAT activity in BAT + than in BAT− groups. |
| Beige adipocytes recruited by BAT via cold & capsinoid treatment. | |||
| Capsinoids BAT thermogenesis & recruited BAT. | |||
| Human | A study between BAT & adiposity in 162 healthy volunteers (103 males & 59 females). 18F-FDG-PET/CT scan after 2 h at 19 °C. [42] | BMI, serum leptin, areas of visceral & subcutaneous fat. | 41% of subjects were found with cold-activated BAT. |
| Detectable BAT with age, with BAT found in 50% of 20-year-olds and >10% in 50-year-olds. | |||
| Human | Clinical trial with 15 subjects underwent cold exposure for 120 min at 14 °C. Treated with 200 mg/os mirabegron or placebo in a double blinded trial. [106] | Detected & quantified mirabegron via Agilent 6460 LC-MS/MS triple quadrupole mass spectrometer. | Cold exposure Resting metabolic rate. |
| Mirabegron BAT glucose uptake. | |||
| No correlation between drug- & cold-stimulation to measure BAT mass or activity. | |||
| Human | rospective study analyzed 5907 patients with 18F-FDG PET/CT scanning, but stringent standards analysis of only 100 patients. [179] | Focal FDG uptake, blood glucose, liver fat content, lipid panel (Total, HDL, and LDL cholesterol; ALT; AST; triglycerides). | 8 men and 17 women brown fat uptake [ABAT (+) group]. |
| Total & LDL cholesterol in ABAT (+) vs. ABAT (−). | |||
| Prevalence of NAFLD in ABAT (−) vs. ABAT (+). | |||
| ABAT serum transaminases (ALT & AST). | |||
| Human | 10 lean & 14 overweight men on cold exposure for 2 h. 18F-FDG-PET scan for BAT activity. [7] | BAT activity & volume, skin & core temperature, BMI | BAT activity in obese subjects vs. lean subjects. |
| BAT volume in obese subjects vs. lean subjects. | |||
| Human | 6 non-acclimated men placed in a 10 °C environment for 2 h daily for 4 weeks. Subjected to electromyography and PET with [11C] acetate & [18F] FDG. [180] | Shivering intensity; BAT metabolism; fractional glucose uptake; insulin; triglycerides; T3; T4; ACTH; leptin levels | 1.9-fold in thermogenic rate in cold exposure. |
| Total BAT volume and activity by 45% after 4 weeks. | |||
| Fractional glucose uptake in supraclavicular BAT. | |||
| Mouse | C57Bl/6 mice with FGF21-KO phenotype subjected 5 °C for 72-h period. Extracted brown fat pads and subjected for RNA isolation, immunohistochemistry, and western blot analysis. [114] | Temperature, confluence of BAT in culture, PGC-1a mRNA expression. | Single-dose recombinant FGF21 browning of WAT. |
| Cold exposure & β3-AR stimulation FGF21 in BAT & thermogenically competent WAT depots. | |||
| FGF21 absence impaired adaptive thermogenesis. | |||
| PGC1-α required for FGF21’s function. | |||
| Mouse | Treated hESCs & hiPSCs to induce embryoid bodies and adipogenic differentiation for 21 days. Subcutaneously transplanted adipocytes in Rag2−/−; Il2yc mice. Infused FGF21 or saline pumps transplanted to the interscapular region. [181] | Lipolysis activity; adiponectin & leptin expression; glucose uptake; oxygen consumption | Converted hPSCs MPCs BAT & WAT |
| Doxycycline treatment PPARG2 WAT phenotype. PPARG2-CEBPB-PRDM16 axis programmed BAT. Transplantation of hPSC-derived adipocytes mature functionality. | |||
| Mouse | 6–8 week-old mice fed with HFD for 8 weeks prior to BAT transplantation; no cold exposure. [182] | Body weight, basal glucose levels, and insulin tolerance. | BAT transplantation HFD-induced obesity; Glucose tolerance; Insulin resistance. BAT removal diet-induced obesity & insulin resistance. |
| Mouse | Study with strain-, sex-, and age-matched donor mice, whose BAT transplanted to other mice. Recipient mice fed on HFD, post transplantation for 20 weeks; cold exposure utilized. [15] | Body weight, total body fat (%), body temperature, O2 consumption, mRNA | BAT transplantation HFD-induced weight gain; |
| Total fat; core body temperature; mRNA of FA oxidation-related genes; HFD-induced insulin resistance; HFD-induced hepatic steatosis & obesity. | |||
| Mouse | 12-week-old male C57BL/6 mice as recipient mice fed standard food and were transplanted into the visceral cavity with 0.1 or 0.4 g BAT, or 0.1 g WAT from epididymal fat pad. [16] | Glucose concentration & uptake; food & water intake; CO2 & heat production; plasma lipids, hormones & proteins. | BAT transplantation glucose tolerance & insulin sensitivity; BAT mass metabolic homeostasis. |
| BAT transplantation more favorable circulating lipid and hormonal profile; circulating norepinephrine concentration & 5-fold increase in serum FGF21 levels. | |||
| Mouse | TZDs (Lobe, Rosi, and Pio) tested on cold acclimated 10-week-old C57BLKS/J-Leprdb/Leprdb male mice for 4 weeks. Raw264.7 macrophages & 3T3-cells treated with TZDs. [183] | Blood glucose; Glucose uptake; body weight; serum triglycerides, cholesterol, and FFA. | TZDs promoted adipocyte differentiation. |
| Lobe stimulation >> two TZDs (Rosi & Pio) effect. | |||
| Lobe beige adipocyte formation & proinflammatory signals in leukocytes & adipocytes. | |||
| Mouse | After acclimation, 4- to 6-week-old C57BL/6J male mice put on high a fat diet for 8 weeks. Post-8 weeks, intraperitoneal injection of 8.5 ug/kg of recombinant follistatin (once/day for one week). [184] | Rectal temperature, mice activity; O2 consumption; CO2 production; glucose tolerance. | HFD-induced obesity; insulin sensitivity. |
| subcutaneous fat browning via AMPK-PGC1a-irisin signaling pathway. | |||
| metabolism via insulin pathway in beige adipocytes. | |||
| Mouse | Male and female heterozygous mice used for mouse embryonic fibroblasts. C57BL/6 mice exposed to cold environment for 8 h. BAT harvested and compared to thermoneutral mice. 0.5 ug/uL of FST on brown preadipocytes for BAT differentiation. [124] | Genotyped embryos, gene and protein expressions of browning & metabolism-related genes; Oxygen consumption. | Follistatin cellular respiration. |
| Follistatin secreted by AT in a paracrine manner. | |||
| Follistatin brown adipocyte and thermogenic gene expression in differentiated brown adipocytes and MEF cultures from both WT and FST KO mice. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Singh, R.; Barrios, A.; Dirakvand, G.; Pervin, S. Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues. Cells 2021, 10, 3030. https://doi.org/10.3390/cells10113030
AMA Style
Singh R, Barrios A, Dirakvand G, Pervin S. Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues. Cells. 2021; 10(11):3030. https://doi.org/10.3390/cells10113030
Chicago/Turabian StyleSingh, Rajan, Albert Barrios, Golnaz Dirakvand, and Shehla Pervin. 2021. "Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues" Cells 10, no. 11: 3030. https://doi.org/10.3390/cells10113030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.