
Dorsal Root Injury—A Model for Exploring Pathophysiology and Therapeutic Strategies in Spinal Cord Injury


Laboratory of Regenertive Neurobiology, Biomedical Center, Department of Neuroscience, Uppsala University, 75124 Uppsala, Sweden
*
Author to whom correspondence should be addressed.
Cells 2021, 10(9), 2185; https://doi.org/10.3390/cells10092185
Submission received: 28 July 2021
/
Revised: 19 August 2021
/
Accepted: 20 August 2021
/
Published: 25 August 2021
(This article belongs to the Special Issue Cellular and Molecular Mechanisms of Dorsal Root Injury and Injured Spinal Cord)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Unraveling the cellular and molecular mechanisms of spinal cord injury is fundamental for our possibility to develop successful therapeutic approaches. These approaches need to address the issues of the emergence of a non-permissive environment for axonal growth in the spinal cord, in combination with a failure of injured neurons to mount an effective regeneration program. Experimental in vivo models are of critical importance for exploring the potential clinical relevance of mechanistic findings and therapeutic innovations. However, the highly complex organization of the spinal cord, comprising multiple types of neurons, which form local neural networks, as well as short and long-ranging ascending or descending pathways, complicates detailed dissection of mechanistic processes, as well as identification/verification of therapeutic targets. Inducing different types of dorsal root injury at specific proximo-distal locations provide opportunities to distinguish key components underlying spinal cord regeneration failure. Crushing or cutting the dorsal root allows detailed analysis of the regeneration program of the sensory neurons, as well as of the glial response at the dorsal root-spinal cord interface without direct trauma to the spinal cord. At the same time, a lesion at this interface creates a localized injury of the spinal cord itself, but with an initial neuronal injury affecting only the axons of dorsal root ganglion neurons, and still a glial cell response closely resembling the one seen after direct spinal cord injury. In this review, we provide examples of previous research on dorsal root injury models and how these models can help future exploration of mechanisms and potential therapies for spinal cord injury repair.
1. Introduction
Dorsal root ganglion (DRG) neurons are pseudounipolar cells with a single process, bifurcating in a peripherally and a centrally directed branch. The latter initially shares the typical features of peripheral nerve tissue with Schwann cells and basement lamina. Just outside the spinal cord, at the dorsal root transitional/entry zone (DRTZ/DREZ), dorsal root axons encounter central glia and course through channels between astrocytes [1,2,3]. Myelinated axons that enter the spinal cord are myelinated by Schwann cells peripherally and by oligodendrocytes centrally, forming a so-called transitional node at the DREZ [4]. Since myelination in the central nervous system (CNS) is triggered at axonal diameters smaller than in the peripheral nervous system (PNS) [5], large non-myelinated sensory axons in the PNS are likely to become myelinated after their entry into the spinal cord [6]. DRG harbors a range of neuronal subtypes, reflecting multiple tactile, proprioceptive and visceral, and nociceptive sensory functions [7,8,9,10,11]. Thus, dorsal root injury models allow the analysis of regenerative potential related to structural and/or functional properties of the affected neurons.
Disorders that damage dorsal roots include compression, (e.g., from herniated disc), infections, autoimmune mechanisms, metabolic disorder, neurotoxic agents, and trauma. Clinically, the most important type of traumatic injury of the dorsal root is spinal root avulsion, a forceful, longitudinal traction of spinal nerve roots, interrupting their connections with the spinal cord. This condition may occur during complicated delivery, from a fall from a high height, or in a traffic accident, and most commonly affects spinal nerve roots innervating the upper extremity, i.e., brachial plexus avulsion.
Root avulsion injuries in experimental animals are typically performed by a surgical approach after exposure of the spinal cord and appropriate root entry regions to achieve standardized and reproducible lesions. However, in clinical root avulsion injuries, the forces that pull the roots from the spinal cord are usually exercised from outside the vertebral canal. The lesion is variable in location and extent of injury, and with possible differences in, for example, the consequences for the regional vascular supply. In terms of location, clinical avulsion injuries can occur distal (postganglionic injury) or proximal (preganglionic injury) to the dorsal root ganglia [12]. The postganglionic avulsion injury has the character of a peripheral nerve injury with the possibility for functional recovery, whereas a preganglionic avulsion injury is, in principle, a longitudinal injury of the spinal cord at the entry of spinal nerve roots, and thus provides similar therapeutic challenges as other spinal cord injuries [13].
Spinal root avulsion injuries typically affect ventral and well as dorsal roots, causing paralysis of denervated muscles, loss of sensory and autonomic function, and, most often, neuropathic pain. Cauda equine syndrome, which encompasses avulsion of lumbosacral roots, also leads to weakness or paralysis of muscles controlling micturition, defecation, and sexual functions. Surgical treatment of spinal root avulsion offers a possibility for partial functional restoration of motor function by reinnervation of proximally located muscles, but clinically efficient treatment for restoring sensory function is thus far not available [13,14]. Thus, dorsal root avulsion exhibits therapeutic challenges specific for this condition but also features clearly relevant for other forms of spinal cord injury. Mechanisms that are implicated in spinal cord pathophysiology and repair are covered in numerous recent reviews, e.g., reference [15].
Here, we discuss injury (axotomy) of dorsal roots as an attractive experimental model to address key aspects of spinal cord injury repair in vivo. Already Ramon y Cajal made the conclusion from his experiment that whereas injury to the peripheral branch of dorsal root ganglia is followed by axonal regeneration and reconnection with peripheral targets, injury to the central branch results in abortive axonal growth [16]. A detailed description of the different dorsal root injury approaches, their advantages and disadvantages were provided previously [17]. Restoration of functions that are lost following dorsal root injury requires that injured dorsal root axons are able to extend axons through a non-permissive environment, which is rapidly building up at the DREZ. In addition, the regenerative response of the central process of DRG neurons is markedly weaker than what follows after an injury to their peripheral process. These two factors contribute to the failure of injured dorsal root axons to re-enter the spinal cord in a similar way a non-permissive environment and attenuated growth response obstruct functional regeneration in the injured spinal cord.
Thus, axotomy (crushing or cutting) of the dorsal root in its peripheral compartment allows investigation of how axonal extension can be promoted, first along with Schwann cells, and thereafter when encountering central glia, which reacts to the injury at a distance, i.e., without damage to the spinal cord itself. On the other hand, the most centrally located dorsal root injury, dorsal root avulsion, will damage the DREZ, including its central glial component, and thereby constitute a true CNS injury, but isolated to the dorsal root axons and their immediate surroundings [13]. In this brief review, we describe how DREZ becomes a barrier for axonal ingrowth, give examples of previous and current research exploring how to overcome this barrier in the adult nervous system, and how future spinal cord injury research can benefit from using dorsal root injury models.
2. Development of the DREZ—From Entry to No-Entry
During development, dorsal root sensory axons enter the spinal cord in temporally successive waves [18]. A transient population of neural crest-derived cells, boundary cap cells, located at spinal cord entry/exit points [19], allow sensory axons to penetrate the spinal cord [20,21] while preventing central glia [22,23] and motor neurons [24,25] from migrating peripherally. Friedrich’s ataxia is a rare, recessive autosomal neurological disorder causing severe impairment in muscle coordination. It is characterized pathologically by abnormalities of myelinated axons in the peripheral nervous system, and spinal cord dorsal column, and of extensive peripheral migration of astrocytes into the dorsal roots [26].
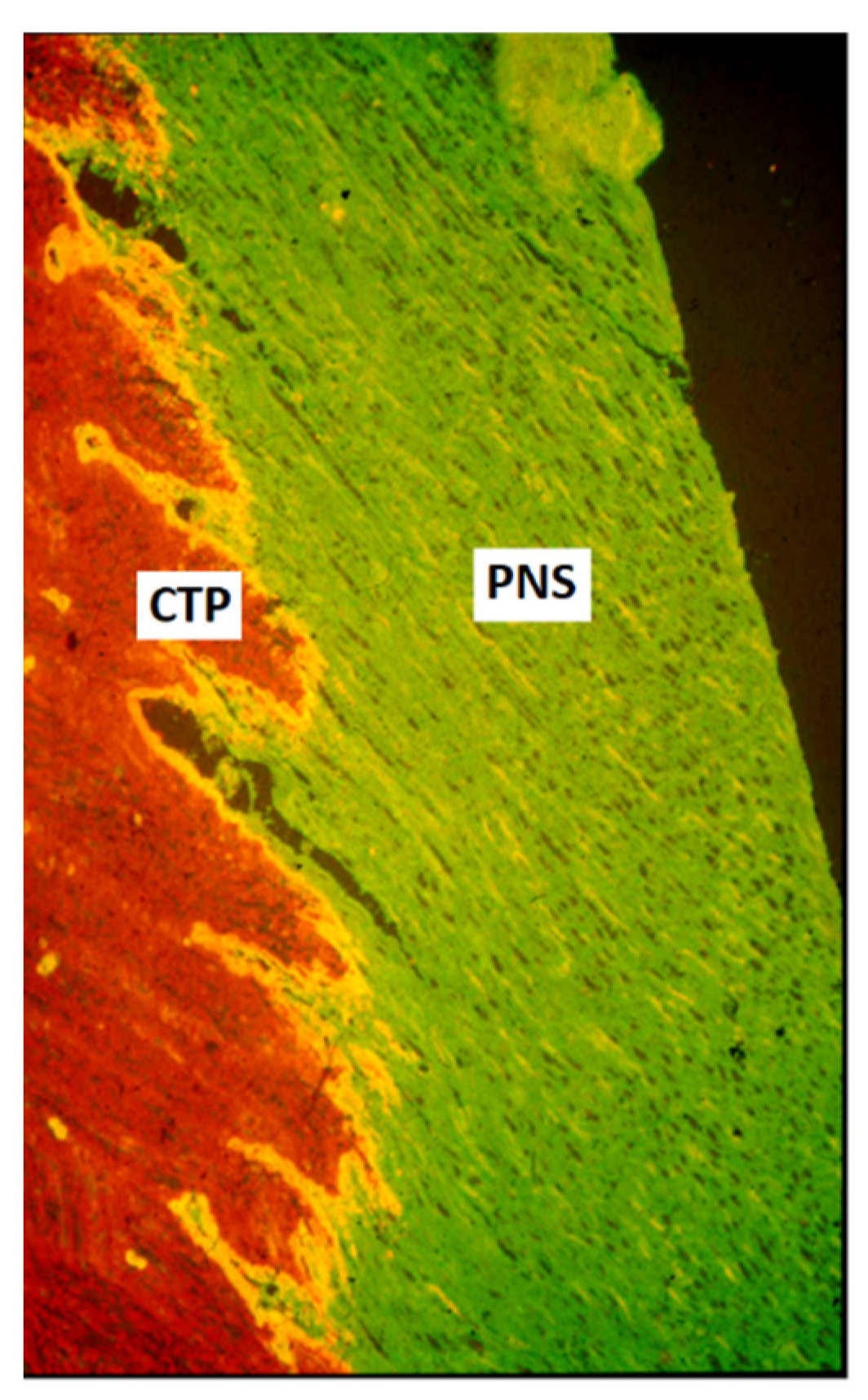
The regulated sensory axon ingrowth appears to be dependent on semaphorin [25,27] and netrin [28,29] signaling. During the late fetal or early postnatal period, the boundary between PNS and CNS becomes a barrier for axonal entry [2,3,30,31]. This barrier is located some distance outside the spinal cord surface, and the central part, containing astrocytes and oligodendrocytes forms, at most levels, dome-shaped central tissue projections (Figure 1). The emergence of this barrier correlates with the deposition of extracellular matrix molecules, such as cytotactin/tenascin and chondroitin 6-sulfate-containing proteoglycans (CPSG) [32,33,34]. The PNS-CNS interface is easily visualized, e.g., by immunolabeling with anti-laminin antibodies for the PNS and anti-glial fibrillary acidic protein for the CNS compartment (Figure 1).
3. Overcoming the Limited Regenerative Competence after Dorsal Root Injury
An appropriate cell body response, comprising a set of so-called regeneration associated genes (RAGs) [36], combined with the emergence of growth supportive axon-glial interactions at the injury site and during axonal elongation, are fundamental for successful nerve regeneration. Axotomy of dorsal root axons results in a much weaker regenerative response by the sensory neurons than what follows after injury to their peripheral branch. Rodent regenerating dorsal root axons grow with a rate of ca 1 mm/24 h compared to ca 3 mm/24 h for injured peripheral nerve axons [37] and fail to display the molecular modifications that is typical after peripheral nerve injury [38,39,40]. Furthermore, injured dorsal root axons have limited ability to grow for long distances even in a peripheral nerve environment [41,42] unless actions to enhance their intrinsic regenerative potential are taken.
Although DRG is composed of phenotypically diverse neurons, they appear to undergo a uniform transcriptional program after peripheral nerve injury, induced by cyclic AMP-dependent transcription factor (ATF-3) [43]. Whether the attenuated regenerative response after dorsal root injury is also mediated by a uniform transcriptional program is unknown. Following dorsal root injury, a neuronal size-related differential expression was shown for mRNAs of the guidance molecules semaphorins and their receptors (neuropilins), as well as for vascular endothelial growth factor (VEGF) [44], suggesting possible differences in the long-term regenerative response to axotomy of central sensory axons.
Interventions, e.g., the use of growth factors or cell-based approaches to promote regeneration of dorsal root axons, are likely to result in differential effects depending on neuronal subtype-specific growth factor receptor pattern and cell-matrix interactions signaling systems (see Section 3.3).
3.1. The Preconditioning Paradigm
The potential of DRG neurons to mount an efficient regenerative response after dorsal root injury is shown by a so-called conditioning lesion, i.e., an injury to the peripheral sensory axons. This intervention, which is typically performed one to two weeks prior to injury to the central processes of the same sensory neurons induces RAGs in DRG neurons and stimulates growth supportive interactions between dorsal root axons and associated non-neuronal cells, thereby promoting elongation of injured dorsal root axons [45]. The conditioning lesion paradigm has made a significant contribution to our understanding of regeneration failure after injury to primary sensory axons.
A conditioning peripheral nerve lesion by itself may allow spinal cord entry of a limited number of injured dorsal root axons [46], which appear to originate from specific DRG subpopulations [47]. An effect of a conditioning injury is also seen on axonal growth after an injury to the central sensory processes in the dorsal column [48], even when the peripheral injury is performed after the spinal cord lesion [49,50]. Since the conditioning lesion approach is not suitable for clinical application, a range of alternative strategies have been adopted to achieve spinal cord ingrowth from injured dorsal root axons. Here, we discuss a few of the most promising of these strategies.
3.2. Axonal Protein Synthesis and Axon Regeneration
Although much of the regeneration-associated neuronal changes occur in the cell bodies of the affected neurons, the importance of a local regeneration supporting metabolic system, proposed already long ago [51,52], is now well established [53,54,55,56,57]. Such metabolic machinery is highly relevant for the repair of human nerve injury since the distance between the site of axotomy and the affected nerve cell body is often considerable. Axonally transported ribosomes and mRNA, together with axonal uptake of ribosomes and mRNA from Schwann cell-derived exosomes, enables initiation of axonal elongation without the early support from the nerve cell body. The starting point and time course of this process may be critical for the possibility of injured axons to negotiate a reactive CNS environment. Transfer of microvesicles from Schwann-like cells has shown promising beneficial effects on axonal growth [58]. To speed up the initiation of the local axonal regenerative machinery might offer a window of opportunity for spinal cord ingrowth of injured dorsal root axons before the fully non-permissive barrier at the DREZ is formed.
3.3. Growth Factors for Promoting Regeneration of Injured Dorsal Root Axons
Extracellular [59] or gene transfer mediated [60] administration of selected growth factors has led to a marked improvement in the regenerative capacity of injured dorsal root axons. The most striking effects have been shown with intrathecal delivery of neurotrophin (NT)3, or glial cell line-derived neurotrophic factor (GDNF) [61,62], or systemic administration of Artemin [63,64,65], a member of the GDNF family. The combined release of peptide mimetics of ciliary neurotrophic factor (CNTF) and GDNF from mesoporous silica nanoparticles implanted on the avulsed DREZ supported partial sensory axon ingrowth [66]. Since these effects are mediated after binding to specific receptors, which are expressed by subpopulations of DRG neurons, a combination of multiple growth factors would be required to achieve the broadest possible impact. Treatment with platelet-rich plasma, a likely source of an abundance of growth factors, was shown to partially restore sensorimotor function after dorsal root injury and may meet the requirement for multiple growth factor administration [67].
Growth factor-mediated growth of dorsal root axons appears to act on intracellular pathways, which at least in part share those implicated in the conditioning lesion response [68], although simultaneous effects on non-neuronal cells at the DREZ may also occur [69]. Importantly, there appears to be a limited time window for neurotrophin-mediated entry of regenerating dorsal root axons since just a short delay of treatment fails to support spinal cord ingrowth [70]. This emphasizes the desire to initiate growth supportive therapy as soon as possible. Dorsal root injury is quite favorable for addressing this aspect.
3.4. Modification of the mTOR Pathway
PTEN (phosphatase and tensin homolog deleted on chromosome 10) has emerged as promising target to recover the intrinsic growth capacity of adult neurons. PTEN inhibits phosphoinositide 3-kinase (PI3-K)/Akt/mTOR pathway, which is critical for sustained axonal elongation. Activation of PTEN also appears to contribute to myelin-associated growth inhibition [71]. Blocking PTEN expression by gene deletion, RNA silencing, or pharmacological inhibitors significantly promotes sustained regenerative growth in vivo by CNS [72,73,74,75,76] and PNS [77,78] neurons without long-term adverse effects [79]. Treatment with a retinoic acid receptor β (RARβ) agonist was shown to modify PTEN activity, promote the growth of sensory axons, counteract the growth repelling environment at the DREZ, and support sensorimotor recovery after spinal root avulsion [80]. Further studies revealed that retinoic acid synthesis by NG2 glia is an important mechanism in providing a growth permissive CNS environment [81].
3.5. Targeting Microtubules
To allow elongation of regenerating axons, microtubules behind the leading edge need to be stabilized [82]. Studies in non-mammalian species even demonstrated efficient microtubule-based axon regeneration without the formation of growth cones [83]. Treatment with the microtubule-stabilizing agent taxol promoted axon regeneration in the mammalian spinal cord [84]. Furthermore, inhibition of kinesin-5, a motor protein that acts as a “brake” on microtubule advancement, promotes sensory neurite extension on a growth inhibitory substrate in vitro [85]. Down-regulation of fidgetin, a microtubule severing molecule, mobilizes an increased mass of labile microtubules, associated with the growth of dorsal root axons into the spinal cord, although evidence of functional repair was not observed [86,87]. Ingrowth of injured dorsal root axons was also shown by daily stimulation for 12 weeks of DRG neurons expressing the receptor hM3Dq, a so-called designer receptor exclusively activated by designer drugs (DREADDs), with its ligand clozapine-N-oxide (CNO), in combination with proteoglycan degrading enzyme treatment [88].
4. Regulating Gene Expression after Axon Injury
The mechanisms for induction and maintenance of a neuronal regeneration program, including its regulation at the gene level, were extensively reviewed [36]. From having previously focused on and clarified the expression of regeneration-associated genes, studies in recent years have begun to unravel how these genes are regulated by non-coding RNAs, and by epigenetic DNA and histone modifications. Non-coding RNAs have distinct roles in transcriptional and posttranscriptional processes and cooperate in a regulatory network for gene expression [36]. Non-coding RNAs which have been implicated in the regulation of peripheral nerve regeneration include long non-coding RNA (lncRNA) [89], micro-RNA (miRNA) [90], and circular RNA (circRNA) [91].
Following spinal cord injury, there are marked changes in the expression of non-coding RNAs, which appear to be associated with the pathophysiological events and the subsequent repair mechanisms [92]. MiR-155 resulted in improved regeneration and functional recovery after spinal cord dorsal column injury, effects that appear to be mediated through the reduced inflammatory response as well as induction of regeneration-associated genes [93]. miRNA-26a primed mesenchymal stem cells were shown to promote spinal cord repair through the mTOR pathway [94], and miR-20a and miR 155-5p promotes the growth of injured sensory axons in the spinal cord dorsal column via the cAMP/PKA [95] and PDZ-RhoGEF/RhoA/GAP43 [96] pathways, respectively. Non-coding RNAs have also been implicated in neuropathic pain [97,98], a common, severely disabling sequelae of spinal root or spinal cord lesion.
Epigenetic modification has recently emerged as a significant factor influencing the outcome after neural injury [99]. Successful peripheral nerve regeneration requires active DNA demethylation [100]. Injury of peripheral but not central sensory axons results in increased chromatin accessibility and histone acetylation. These epigenetic changes are accompanied by the induction or regeneration-associated gene expression [101]. Along the same line, DNA methylation of sequences of the Myc proto-oncogene preceded the up-regulation of a set of RAGs after a conditioning lesion of sensory neurons [102]. Interestingly, environmental enrichment prior to a spinal cord injury was shown to promote functional recovery, mediated by Creb-binding protein histone acetylation [103]. By combining environmental enrichment with a preconditioning lesion, a further enhancement of the growth capacity of injured spinal cord sensory axons has been achieved [104]. Further exploration of these findings, e.g., in dorsal root injury models, may help to develop combinatorial therapies based on interventions with bioactive agents and rehabilitative measures [105].
5. Growth Inhibition at the DREZ
5.1. Manipulating Growth Inhibitory Factors
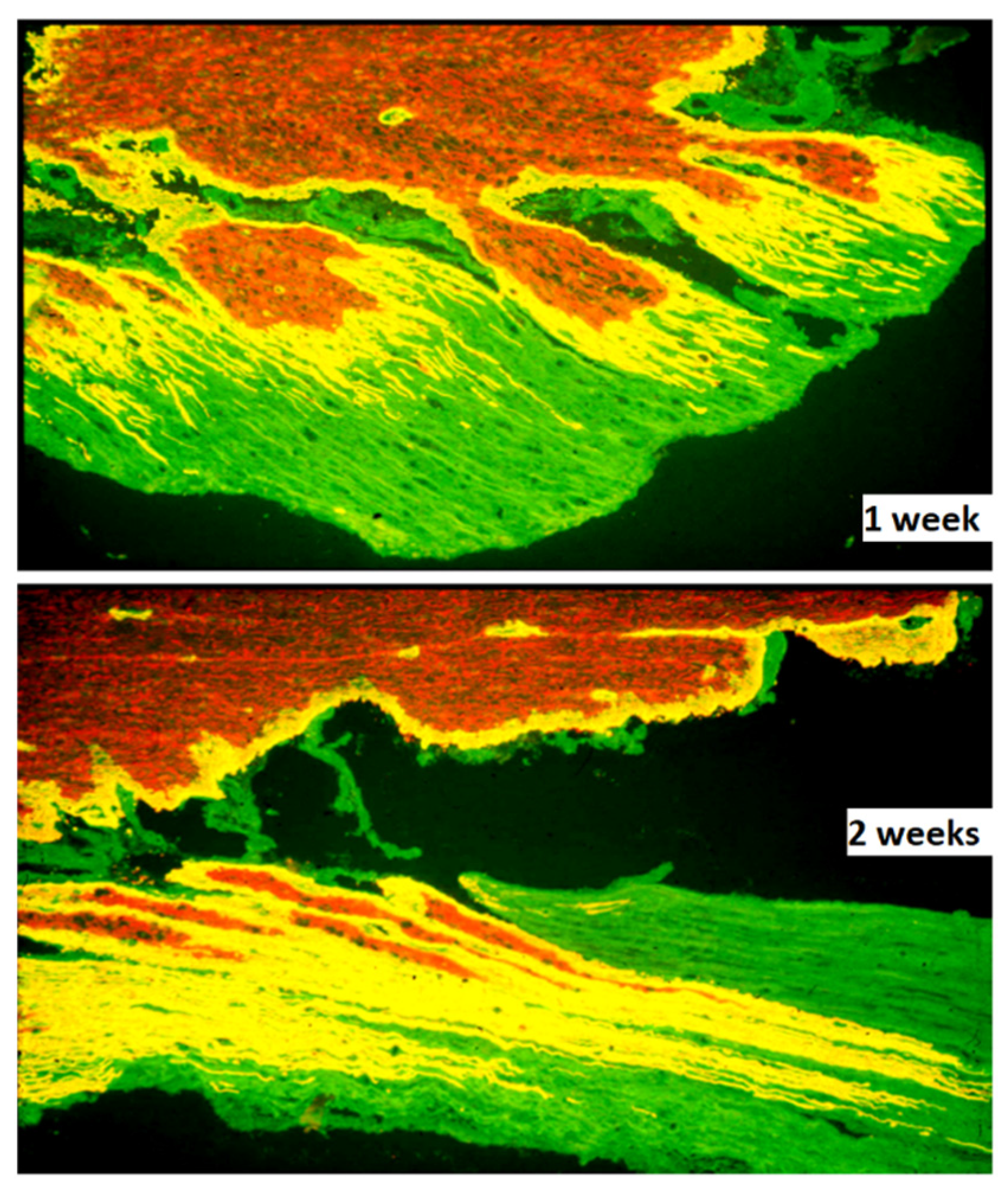
Microglia and astrocytes play critical roles in the pathophysiology and repair after spinal cord injury and are, therefore, potential targets for therapy [106]. Reactive CNS glia creates an efficient barrier for spinal cord ingrowth of injured axons at the DREZ. Astrocytes at the DREZ and along the sensory pathways in the spinal cord proliferate [107,108], and undergo marked hypertrophy, as evidenced, for example, by the peripheral extension of long processes [35,109] (Figure 2). The growth-inhibitory influence at the DREZ is considered to be formed mainly by the deposition of proteoglycans [31,32,33] and the presence of myelin-associated growth inhibitors [110] Up-regulation of astrocyte expressed calcium-binding protein S100A4/Mts1 [111], shown to be a growth inhibitor in vitro [112,113] and in vivo after implantation of bNCSCs to the avulsed spinal cord [114], may also contribute.
Previous studies reported that a conditioning lesion combined with enzymatic degradation of chondroitinase sulfate proteoglycans with chondroitinase ABC [115], their removal through gene deletion [116], blocking Nogo-receptor function [117,118], or integrin activation [119] allow regeneration of injured dorsal root axons into the spinal cord. However, recent experiments indicate that a conditioning lesion in combination with proteoglycan degradation or blocking myelin-associated inhibitors is not sufficient in this context, and that specific growth factor stimulation, in this specific case with GDNF, is required for sensory axon entry and functional recovery [120]. This finding indicates that the mechanisms which prevent spinal cord ingrowth by injured dorsal root axons are still far from clear and need further investigations.
The limited effect on sensory axon spinal cord ingrowth by pharmacological interventions targeting the DREZ may be related to the observation that regenerating sensory axons form synapse-like contacts with non-neuronal cells, possibly NG2-positive glia, rather than traversing the PNS-CNS boundary and enter the spinal cord [121,122,123,124]. This process resembles the appearance of synapse-like connections between injured sensory axons in the dorsal column and NG2 positive glia [125]. Thus, NG2+ glia may provide cues for establishing contacts with regenerating dorsal root axons, perhaps originating from specific sensory neurons. To identify these cues in detail and apply counteracting interventions may contribute to functional dorsal root axon regeneration.
5.2. Clearance of Axonal and Myelin Debris
During Wallerian degeneration of peripheral nerves, Schwann cells in cooperation with fibroblasts and immune cells, clear disintegrating axons and myelin in preparation for a growth permissive pathway for axon regeneration. In contrast, myelin debris arising from Wallerian degeneration in the central nervous system remains for long periods of time [126,127]. In humans, residues of degenerating myelin have been observed decades after spinal cord injury. These residues and the associated cellular reactions appear to hamper axonal growth. To speed up myelin clearance in the injured spinal cord is thus a relevant objective for promoting spinal cord injury repair. This issue can be favorably explored after dorsal root injury, which allows examination of myelin degradation and elimination in the spinal cord dorsal column without direct damage to the spinal cord, leaving its vascular supply and tissue architecture essentially unaffected.
The long-term lingering of myelin debris after spinal cord injury is probably the result of an insufficient activation of the macrophage/microglial program required for rapid and complete myelin degradation [128,129,130]. Toll-like receptor signaling mediates myelin phagocytosis [131,132], whereas beta-amyloid precursor clearing enzyme (BACE)1 [133,134], and signal regulatory protein-α (SIRPα) [135] delays this process. These findings provide options for approaches for rapid elimination of myelin debris and, hence, possibly minimize one obstacle for sensory axon extension across the DREZ.
Small rodents have dominated experimental studies on spinal cord injury for decades, but, as often recognized, translation from these studies to humans is frequently difficult. Early studies of Wallerian degeneration in the spinal cord or animals with considerably larger myelinated axons than what is the case in the small rodents have shown the emergence and long-term presence of abnormal profiles, such as the “myelinoclasts” in monkeys [136]. Similar profiles have also been observed in the cat [137,138] but not in the rat [126,127]. These structures are formed of layers of myelin wrapped around non-neuronal cells, most likely microglia, which subsequently appear to undergo degeneration and become phagocytosed by other populations of microglia [138]. The mechanisms underlying these distinct differences in how Wallerian degeneration proceeds in larger organisms compared to rats or mice may be important to consider in translational research on spinal cord injury repair. Further analysis also needs to consider the presence of microglia/macrophage subpopulations and their properties in response to Wallerian degeneration in the spinal cord. Recent studies using single-cell RNA analysis have shown the existence of multiple macrophage populations in degenerating peripheral nerve [139] and a marked heterogeneity of microglia in neural disorders [140].
6. Secondary Neurodegeneration in the Spinal Cord
Dorsal root avulsion has consequences not only for spinal cord glia and immune cells but also results in degeneration of postsynaptic neurons in the dorsal as well as ventral horn [141]. Loss of neurons after spinal cord injury may be a consequence of the vascular and immune pathophysiology and thus becomes an important part of the secondary injury area [141,142]. In addition, neurons may be lost at a significant distance from both the primary and secondary lesion area as a result of diminished input to postsynaptic neurons (anterograde transneuronal degeneration).
Conversely, there is a loss of previously existing postsynaptic targets, causing presynaptic neurons to degenerate (retrograde transneuronal degeneration). These processes effectively impair the possibility for potentially regenerating axons to restore functionally useful connections and for replacement of lost connections by circuit rewiring [143]. Dorsal root avulsion provides a clinically relevant condition for exploring mechanisms underlying anterograde transneuronal degeneration in particular and means to counteract this process.
7. Bridges for Dorsal Root Injury Repair
Cell-based bridges, acellular bridges, and their combinations are attractive strategies for the repair of the spinal cord and dorsal root injury. The overall purpose of these strategies is to promote functional recovery by (i) survival/growth support for intrinsic cells,(ii) disease modification to facilitate endogenous repair, and (iii) replacement of lost cells or introduction of relay cells. The implementation of these strategies for spinal cord injury is discussed by Guo [144]. Lin et al. analyze how stem cells and biomimetic material for spinal cord repair interact with the human immune system [145], and Rodríguez-Barrera addresses the potential of recruitment from endogenous stem cells of cells for spinal cord injury repair [146]. Here, we discuss bridges aimed to promote functional growth of injured dorsal root axons into the spinal cord.
7.1. Cell-Based Bridges
Early studies aimed to provide a bridge for regenerating dorsal root axons into the spinal cord by exploiting, for example, Schwann cells [147], or activated macrophages [148], both types of cells which cooperate in peripheral nerve regeneration. These attempts were not successful, however; Schwann cells do not readily integrate into a CNS environment [32,149,150], and activated macrophages in a CNS environment tend to induce axonal retraction rather than extension [151].
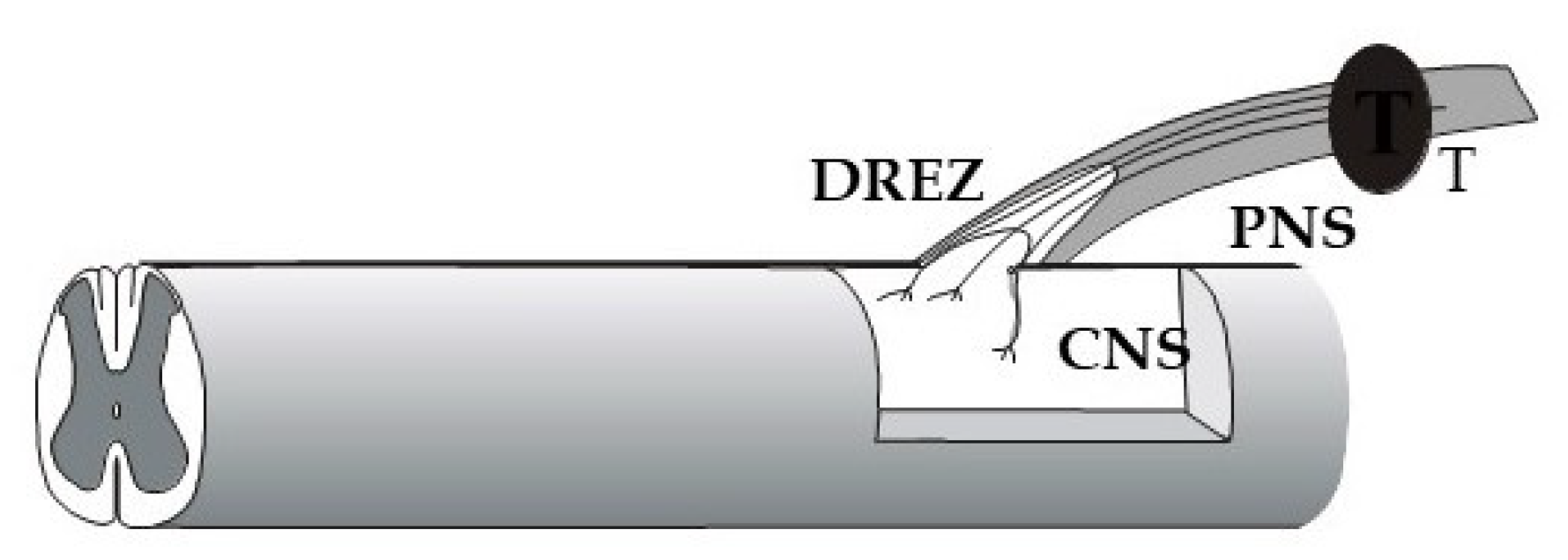
With the purpose of probing the greater capacity for immature neurons to regenerate after injury, human embryonic/fetal dorsal root ganglion cells were implanted into the cavity of extirpated rodent DRGs (Figure 3). Sensory axons from the implant entered the host spinal cord along blood vessels, made functional synaptic connections there, and projected axons into the peripheral nerve [152,153]. These findings provided proof of principle that the barrier between the peripheral and central nervous systems can be overcome in adulthood. This sensory conduit from the periphery to the spinal cord circuitry was possible to create with highly growth competent sensory neurons, even without interfering with the non-permissive DREZ environment, through their ability to circumvent the glial barrier by extending along blood vessels [152].
Olfactory ensheathing cells (OECs) are CNS glia, ensheathing non-myelinated olfactory nerve axons in the intact nervous system, and able to serve as a cellular substrate for regenerating axons. OEC implants readily integrate into non-olfactory CNS regions and have been shown to be able to provide a pathway for sensory axon growth into the spinal cord and restore sensorimotor function after spinal root avulsion [154].
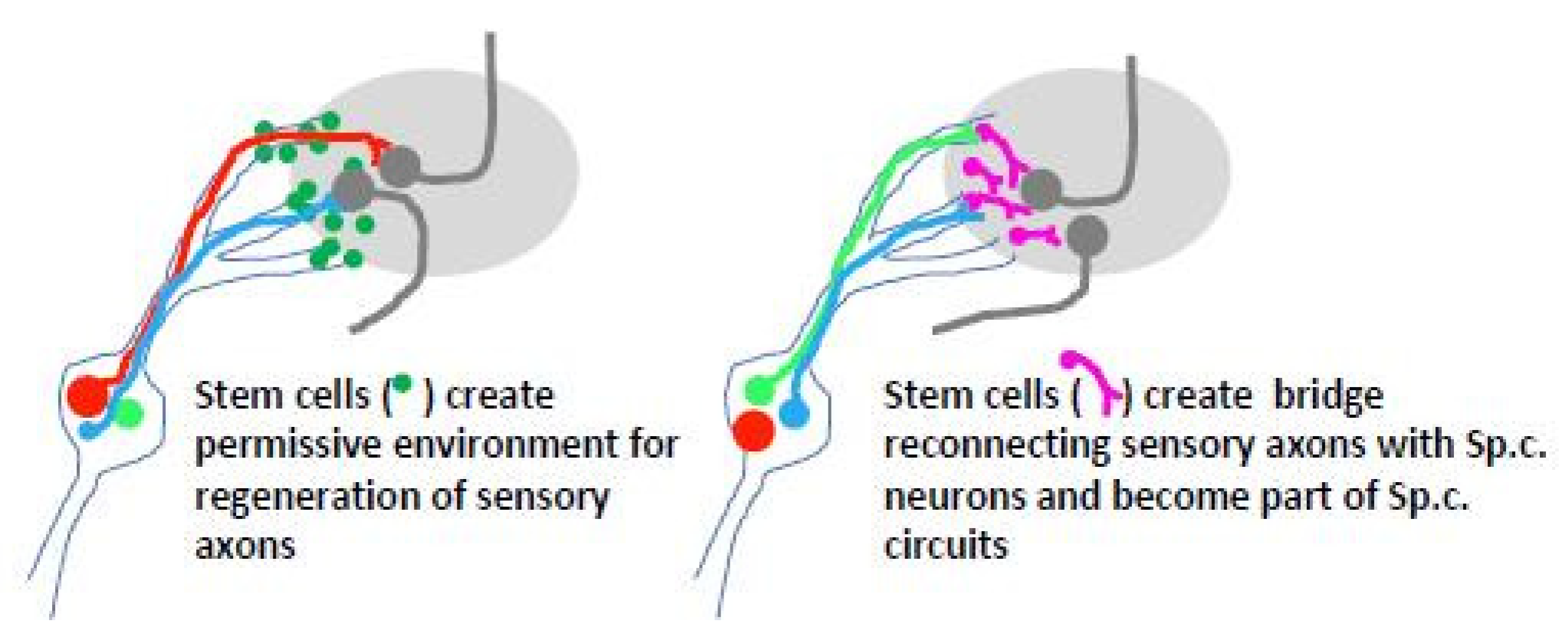
Stem cell-based transplants at the DREZ could serve as facilitators for entry of dorsal root axons or as neuronal relays to interconnect dorsal root axons in the PNS compartment with spinal cord neurons (Figure 4). Since boundary cap cells allow sensory axons to enter the spinal cord during development (see Section 2), stem cells generated in vitro from this location were transplanted to the site of dorsal root avulsion in the mouse [155,156]. Transplanted cells survived, formed cellular bridges in the peripheral dorsal root compartment, partially migrated into the spinal cord, and underwent diverse neural differentiation but failed to support axonal ingrowth into the spinal cord. A possible explanation for this failure is that transplanted boundary cap neural crest stem cells contribute to a population of cells that express the growth repelling protein Mts1/S100A4 [114].
Human neural progenitor cell implants were shown to support sensory axon ingrowth and partial recovery of sensory function after dorsal root avulsion, seemingly by creating channels, i.e., acting as growth facilitators [157]. When such transplants were combined with nanoparticle-mediated delivery of growth factor peptide mimetics to the DREZ, sensory axon ingrowth was aborted [64]. Further studies indicated that growth factor administration accelerated progenitor cell maturation, probably causing a decline in their growth supportive properties [64]. This finding indicates that a combinatorial treatment strategy does not necessarily provide the anticipated additive or synergistic effects. However, combining platelet-rich plasma gel, a likely source of multiple growth factors, and human embryonic stem cell transplants allowed sensory axon ingrowth into the spinal cord [158].
In addition, to provide growth-supporting substrate, OEC or stem cell implants are likely to release growth supportive agents and presumably modify immune responses in a way that does not antagonize sensory axon growth across the DREZ. Clarifying the mechanisms underlying these approaches and explore the way to improve their efficiency is thus likely to contribute to the exploitation of cell-based therapies in direct spinal cord injury, e.g., by the administration of exosomes containing defined growth supporting and/or disease-modifying components [144,159].
7.2. The Dorsal Root as Pathway for Bridging Spinal Cord Injury
Pioneering studies by Aguayo and collaborators demonstrated that injured CNS neurons are able to elongate for long distances within a peripheral nerve graft [160,161,162]. As an extension of these findings, it became clear that lack of a growth-supporting environment is a key factor underlying the regeneration failure of injured axons in the CNS. In line with this, injured CNS axons fail to leave a peripheral nerve graft and re-enter the CNS and make functional connections due to the establishment of a non-permissive environment at the interface between the graft and the CNS tissue [163].
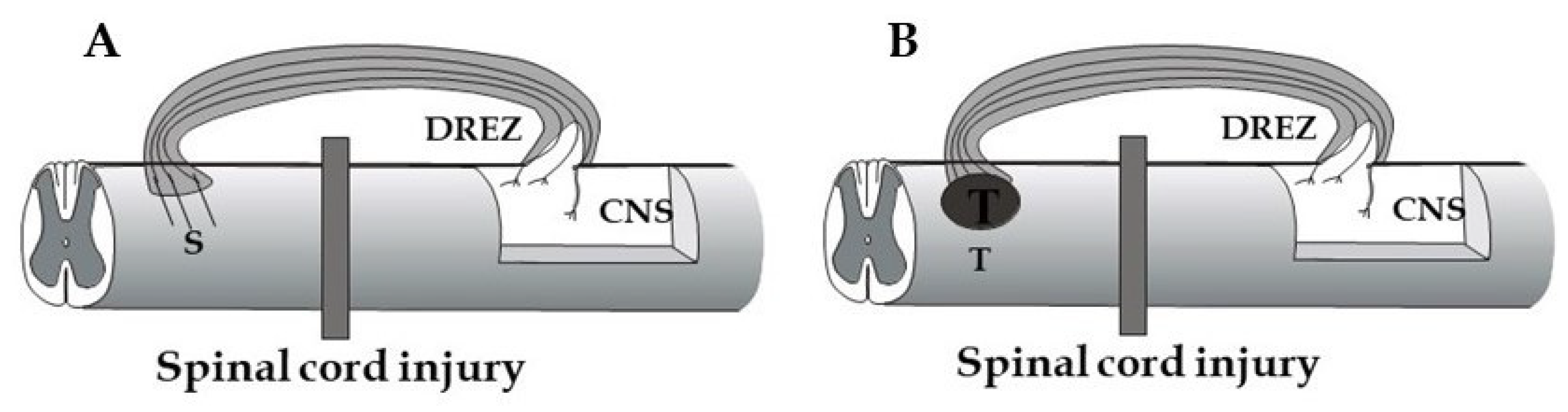
The application of bridges that circumvent a severely damaged spinal cord area would be a relevant option for restoring descending control of specific spinal cord functions. To make the axons elongating in the bridge enter the spinal cord, the graft-cord barrier issue needs to be resolved. This situation has similarities to the growth repelling barrier facing injured dorsal root axons at the DREZ. Approaches, which are discussed above to promote dorsal root regeneration into the spinal cord, are thus likely to be relevant also for achieving entry of axons, which have grown through a bridging graft. One additional, interesting aspect in this context is that the intact DREZ does not appear to exhibit a growth repellent barrier [164]. Figure 5 shows two possible approaches exploiting the modified or intact DREZ as an entry site for bridging a spinal cord injury, either by host axons growing through a peripheral nerve or biomimetic graft (A) or by axons from an intraspinal, stem cell-derived neuronal transplant (B).
7.3. Bioprinting and Dorsal Root Injury Repair
3D bioprinting has rapidly developed as a highly attractive technology for biomimetic implants that is able to support neuronal survival, counteract disease driving processes and support functional repair following spinal cord injury [144]. Based on detailed information on the underlying pathophysiological processes and conditions of the patient, 3D bioprinted implants may be designed to match patient-specific conditions [165]. Recent studies have demonstrated 3D bioprinted constructs that support neurite outgrowth from dorsal root ganglion neurons in vitro [166,167,168]. Dorsal root injury would be a useful in vivo model for exploring these developments and their further translation to the injured spinal cord.
8. Conclusions
Injuries that interrupt dorsal root axons result in loss of sensorimotor functions and often neuropathic pain, symptoms which are usually most serious following root avulsion injury. Injured dorsal root axons are unable to restore lost connections due to insufficient regeneration performance in combination with the encounter of a growth repelling CNS environment, i.e., factors which are closely similar to those that underlie the regeneration failure after direct spinal cord injury. Dorsal root injury offers attractive experimental models to determine key mechanisms in these processes in vivo, their exploitation for the development of novel therapeutic strategies, and their translation to a clinical setting for dorsal root injuries.
Dorsal root avulsion, which constitutes a longitudinal spinal cord injury, is of particular relevance in the broader context of spinal cord injury A successful strategy to tackle dorsal root avulsion injury most likely requires a combinatorial approach, applied in an appropriate spatial and temporal manner. We envision such a strategy might include the following actions. (i) Cell-based approaches, e.g., via exosomes derived from primed stem cells or as bioprinted constructs, may be suitable for promoting tissue repair at the injury site., as well as for creating pathways across the DREZ. (ii) Intrathecal administration of agents that specifically counteract non-neuronal axon growth inhibitors at the DREZ, as well as along ascending sensory pathways, are likely to be needed as additive measures. (iii) Application of gene regulatory tools to initiate and achieve sustained axonal growth by all subpopulations of dorsal root ganglion neurons. The dorsal root-spinal cord unit provides a favorable platform for assessing the outcome of these interventions, individually as well as in various combinations, initially in small rodent models and subsequently translated to larger species prior to clinical trials in humans.
Funding
The author’s cited work was supported by the European Commission BMH4-CT97-2586, the Swedish Research Council projects 5420, 6555, 9917 and 20716, Olle Engkvist Foundation and Signhild Engkvist’s Foundation.
Data Availability Statement
The authors’ findings and data referred to in the article are available from the corresponding author on reasonable request.
Acknowledgments
We are grateful for the cooperative work and technical support which have contributed to the authors research referred to in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Berthold, C.H.; Carlstedt, T. Observations on the morphology at the transition between the peripheral and the central nervous system in the cat. II. General organization of the transitional region in S1 dorsal rootlets. Acta Physiol. Scand. Suppl. 1977, 446, 23–42. [Google Scholar]
- Fraher, J.P.; Sheehan, M.M. The CNS-PNS transitional zone of rat cervical dorsal roots during development and at maturity. A morphological and morphometric study. J. Anat. 1987, 152, 189–203. [Google Scholar]
- Fraher, J. Axons and glial interfaces: Ultrastructural studies. J. Anat. 2002, 200, 415–430. [Google Scholar] [CrossRef]
- Berthold, C.H.; Carlstedt, T. Observations on the morphology at the transition between the peripheral and the central nervous system in the cat. III. Myelinated fibres in S1 dorsal rootlets. Acta Physiol. Scand. Suppl. 1977, 446, 43–60. [Google Scholar]
- Hildebrand, C.; Remahl, S.; Persson, H.; Bjartmar, C. Myelinated nerve fibres in the CNS. Prog. Neurobiol. 1993, 40, 319–384. [Google Scholar] [CrossRef]
- Carlstedt, T. Observations on the morphology at the transition between the peripheral and the central nervous system in the cat. IV. Unmyelinated fibres in S1 dorsal rootlets. Acta Physiol. Scand. Suppl. 1977, 446, 61–72. [Google Scholar] [PubMed]
- Jänig, W. Neurobiology of visceral afferent neurons: Neuroanatomy, functions, organ regulations and sensations. Biol. Psychol. 1996, 42, 29–51. [Google Scholar] [CrossRef]
- Lawson, S.N. Phenotype and function of somatic primary afferent nociceptive neurones with C-, Adelta- or Aalpha/beta-fibres. Exp Physiol. 2002, 872, 39–44. [Google Scholar] [CrossRef]
- Le Pichon, C.E.; Chesler, A.T. The functional and anatomical dissection of somatosensory subpopulations using mouse genetics. Front Neuroanat. 2014, 8, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, A.J. Identifying functional populations among the interneurons in laminae I-III of the spinal dorsal horn. Mol. Pain. 2017, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Kupari, J.; Usoskin, D.; Parisien, M.; Lou, D.; Hu, Y.; Fatt, M.; Lönnerberg, P.; Spångberg, M.; Eriksson, B.; Barkas, N.; et al. Single cell transcriptomics of primate sensory neurons identifies cell types associated with chronic pain. Nat. Commun. 2021, 12, 1510. [Google Scholar] [CrossRef] [PubMed]
- Wade, R.G.; Takwoingi, Y.; Wormald, J.C.R.; Ridgway, J.P.; Tanner, S.; Rankine, J.J.; Bourke, G. Magnetic resonance imaging for detecting root avulsions in traumatic adult brachial plexus injuries: Protocol for a systematic review of diagnostic accuracy. Syst. Rev. 2018, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Carlstedt, T.; Havton, L. The longitudinal spinal cord injury: Lessons from intraspinal plexus, cauda equina and medullary conus lesions. Handbook Clin. Neurol. 2012, 109, 337–354. [Google Scholar] [CrossRef]
- Carlstedt, T. New Treatments for Spinal Nerve Root Avulsion Injury. Front. Neurol. 2016, 7, 135, Erratum in: Front. Neurol. 2017, 8, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hachem, L.D.; Fehlings, M.G. Pathophysiology of Spinal Cord Injury. Neurosurg Clin N Am. 2021, 32, 305–313. [Google Scholar] [CrossRef]
- Ramon y Cajal, S.; DeFelipe, J.; Jones, E.G. Cajal’s Degeneration and Regeneration of the Nervous System; Oxford University Press: Oxford, UK, 1991; ISBN 13:9780195065169. [Google Scholar] [CrossRef]
- Aldskogius, H.; Kozlova, E.N. Dorsal root injury for the study of spinal cord injury repair. Animal Models of Spinal Cord Repair; Humana Press: Totowa, NJ, USA, 2013; pp. 109–129. [Google Scholar] [CrossRef]
- Masuda, T.; Shiga, T. Chemorepulsion and cell adhesion molecules in patterning initial trajectories of sensory axons. Neurosci. Res. 2005, 51, 337–347. [Google Scholar] [CrossRef]
- Niederländer, C.; Lumsden, A. Late emigrating neural crest cells migrate specifically to the exit points of cranial branchiomotor nerves. Development 1996, 122, 2367–2374. [Google Scholar] [CrossRef]
- Golding, J.P.; Cohen, J. Border controls at the mammalian spinal cord: Late-surviving neural crest boundary cap cells at dorsal root entry sites may regulate sensory afferent ingrowth and entry zone morphogenesis. Mol. Cell. Neurosci. 1997, 9, 381–396. [Google Scholar] [CrossRef] [Green Version]
- Golding, J.; Shewan, D.; Cohen, J. Maturation of the mammalian dorsal root entry zone--from entry to no entry. Trends Neurosci. 1997, 20, 303–308. [Google Scholar] [CrossRef]
- Kucenas, S.; Wang, W.D.; Knapik, E.W.; Appel, B. A selective glial barrier at motor axon exit points prevents oligodendrocyte migration from the spinal cord. J. Neurosci. 2009, 29, 15187–15194. [Google Scholar] [CrossRef]
- Coulpier, F.; Decker, L.; Funalot, B.; Vallat, J.M.; Garcia-Bragado, F.; Charnay, P.; Topilko, P. CNS/PNS boundary transgression by central glia in the absence of Schwann cells or Krox20/Egr2 function. J. Neurosci. 2010, 30, 5958–5967. [Google Scholar] [CrossRef]
- Bron, R.; Vermeren, M.; Kokot, N.; Andrews, W.; Little, G.E.; Mitchell, K.J.; Cohen, J. Boundary cap cells constrain spinal motor neuron somal migration at motor exit points by a semaphorin-plexin mechanism. Neural Dev. 2007, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Mauti, O.; Domanitskaya, E.; Andermatt, I.; Sadhu, R.; Stoeckli, E.T. Semaphorin6A acts as a gate keeper between the central and the peripheral nervous system. Neural Dev. 2007, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Koeppen, A.H.; Becker, A.B.; Qian, J.; Gelman, B.B.; Mazurkiewicz, J.E. Friedreich Ataxia: Developmental Failure of the Dorsal Root Entry Zone. J. Neuropathol. Exp. Neurol. 2017, 76, 969–977. [Google Scholar] [CrossRef]
- Liu, R.Q.; Wang, W.; Legg, A.; Abramyan, J.; O’Connor, T.P. Semaphorin 5B is a repellent cue for sensory afferents projecting into the developing spinal cord. Development 2014, 141, 1940–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, T.; Sakuma, C.; Yaginuma, H. Role for netrin-1 in sensory axonal guidance in higher vertebrates. Fukushima, J. Med. Sci. 2009, 55, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, A.M.; Jucius, T.J.; Sigaud, L.P.; Tang, F.L.; Xiong, W.C.; Ackerman, S.L.; Burgess, R.W. Analysis of Expression Pattern and Genetic Deletion of Netrin5 in the Developing Mouse. Front. Mol. Neurosci. 2016, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Carlstedt, T.; Dalsgaard, C.J.; Molander, C. Regrowth of lesioned dorsal root nerve fibers into the spinal cord of neonatal rats. Neurosci. Lett. 1987, 74, 14–18. [Google Scholar] [CrossRef]
- Carlstedt, T. Reinnervation of the mammalian spinal cord after neonatal dorsal root crush. J. Neurocytol. 1988, 17, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Grimpe, B.; Pressman, Y.; Lupa, M.D.; Horn, K.P.; Bunge, M.B.; Silver, J. The role of proteoglycans in Schwann cell/astrocyte interactions and in regeneration failure at PNS/CNS interfaces. Mol. Cell. Neurosci. 2005, 28, 18–29. [Google Scholar] [CrossRef]
- Beggah, A.T.; Dours-Zimmermann, M.T.; Barras, F.M.; Brosius, A.; Zimmermann, D.R.; Zurn, A.D. Lesion-induced differential expression and cell association of Neurocan, Brevican, Versican V1 and V2 in the mouse dorsal root entry zone. Neuroscience 2005, 133, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Waselle, L.; Quaglia, X.; Zurn, A.D. Differential proteoglycan expression in two spinal cord regions after dorsal root injury. Mol. Cell. Neurosci. 2009, 42, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Aldskogius, H.; Kozlova, E.N. Strategies for repair of the deafferented spinal cord. Brain Res. Brain Res. Rev. 2002, 40, 301–308. [Google Scholar] [CrossRef]
- Mahar, M.; Cavalli, V. Intrinsic mechanisms of neuronal axon regeneration. Nat. Rev. Neurosci. 2018, 19, 323–337. [Google Scholar] [CrossRef]
- Richardson, P.M.; Verge, V.M. Axonal regeneration in dorsal spinal roots is accelerated by peripheral axonal transection. Brain Res. 1987, 411, 406–408. [Google Scholar] [CrossRef]
- Broude, E.; McAtee, M.; Kelley, M.S.; Bregman, B.S. c-Jun expression in adult rat dorsal root ganglion neurons: Differential response after central or peripheral axotomy. Exp. Neurol. 1997, 148, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaiger, F.W.; Hager, G.; Schmitt, A.B.; Horvat, A.; Hager, G.; Streif, R.; Spitzer, C.; Gamal, S.; Breuer, S.; Brook, G.A.; et al. Peripheral but not central axotomy induces changes in Janus kinases (JAK) and signal transducers and activators of transcription (STAT). Eur. J. Neurosci. 2000, 12, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.; Zhou, L.; Serger, E.; Palmisano, I.; De Virgiliis, F.; Hutson, T.H.; Mclachlan, E.; Freiwald, A.; La Montanara, P.; Shkura, K.; et al. AMPK controls the axonal regenerative ability of dorsal root ganglia sensory neurons after spinal cord injury. Nat. Metab. 2020, 2, 918–933. [Google Scholar] [CrossRef]
- Richardson, P.M.; Issa, V.M. Peripheral injury enhances central regeneration of primary sensory neurones. Nature 1984, 309, 791–793. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.M.; Verge, V.M. The induction of a regenerative propensity in sensory neurons following peripheral axonal injury. J. Neurocytol. 1986, 15, 585–594. [Google Scholar] [CrossRef]
- Renthal, W.; Tochitsky, I.; Yang, L.; Cheng, Y.C.; Li, E.; Kawaguchi, R.; Geschwind, D.H.; Woolf, C.J. Transcriptional Reprogramming of Distinct Peripheral Sensory Neuron Subtypes after Axonal Injury. Neuron 2020, 108, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, T.; Risling, M.; Carlstedt, T.; Hammarberg, H.; Wallquist, W.; Cullheim, S.; Sköld, M.K. Expression of Semaphorins, Neuropilins, VEGF, and Tenascins in Rat and Human Primary Sensory Neurons after a Dorsal Root Injury. Front. Neurol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Liu, R.; Xu, Y.; Ma, X.; Zhou, B. The Mechanisms of Peripheral Nerve Preconditioning Injury on Promoting Axonal Regeneration. Neural Plast. 2021, 2021, 6648004. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.S.; Woolf, C.J.; Haque, N.S.; Anderson, P.N. Axonal regeneration from injured dorsal roots into the spinal cord of adult rats. J. Comp. Neurol. 1999, 410, 42–54. [Google Scholar] [CrossRef]
- Kalous, A.; Keast, J.R. Conditioning lesions enhance growth state only in sensory neurons lacking calcitonin gene-related peptide and isolectin B4-binding. Neuroscience 2010, 166, 107–121. [Google Scholar] [CrossRef]
- Neumann, S.; Woolf, C.J. Regeneration of dorsal column fibers into and beyond the lesion site following adult spinal cord injury. Neuron 1999, 23, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Ylera, B.; Ertürk, A.; Hellal, F.; Nadrigny, F.; Hurtado, A.; Tahirovic, S.; Oudega, M.; Kirchhoff, F.; Bradke, F. Chronically CNS-injured adult sensory neurons gain regenerative competence upon a lesion of their peripheral axon. Curr. Biol. 2009, 19, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Blesch, A.; Lu, P.; Tsukada, S.; Alto, L.T.; Roet, K.; Coppola, G.; Geschwind, D.; Tuszynski, M.H. Conditioning lesions before or after spinal cord injury recruit broad genetic mechanisms that sustain axonal regeneration: Superiority to camp-mediated effects. Exp. Neurol. 2012, 235, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Tobias, G.S.; Koenig, E. Axonal protein synthesizing activity during the early outgrowth period following neurotomy. Exp. Neurol. 1975, 49, 221–234. [Google Scholar] [CrossRef]
- Frankel, R.D.; Koenig, E. Identification of locally synthesized proteins in proximal stump axons of the neurotomized hypoglossal nerve. Brain Res. 1978, 141, 67–76. [Google Scholar] [CrossRef]
- Lopez-Leal, R.; Court, F.A. Schwann Cell Exosomes Mediate Neuron-Glia Communication and Enhance Axonal Regeneration. Cell. Mol. Neurobiol. 2016, 36, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Schnatz, A.; Schillner, M.; Woertge, S.; Müller, C.; von Graevenitz, I.; Waisman, A.; van Minnen, J.; Vogelaar, C.F. A predominantly glial origin of axonal ribosomes after nerve injury. Glia 2018, 66, 1591–1610. [Google Scholar] [CrossRef]
- Hervera, A.; De Virgiliis, F.; Palmisano, I.; Zhou, L.; Tantardini, E.; Kong, G.; Hutson, T.; Danzi, M.C.; Perry, R.B.; Santos, C.X.C.; et al. Reactive oxygen species regulate axonal regeneration through the release of exosomal NADPH oxidase 2 complexes into injured axons. Nat. Cell Biol. 2018, 20, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Terenzio, M.; Koley, S.; Samra, N.; Rishal, I.; Zhao, Q.; Sahoo, P.K.; Urisman, A.; Marvaldi, L.; Oses-Prieto, J.A.; Forester, C.; et al. Locally translated mTOR controls axonal local translation in nerve injury. Science 2018, 359, 1416–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koley, S.; Rozenbaum, M.; Fainzilber, M.; Terenzio, M. Translating regeneration: Local protein synthesis in the neuronal injury response. Neurosci. Res. 2019, 139, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Ye, K.; Yu, J.; Li, L.; Wang, H.; Tang, B.; Ni, W.; Zhou, J.; Ling, Y.; Lu, X.; Niu, D.; et al. Microvesicles from Schwann-Like Cells as a New Biomaterial Promote Axonal Growth. J. Biomed. Nanotechnol. 2021, 17, 291–302. [Google Scholar] [CrossRef]
- Smith, G.M.; Falone, A.E.; Frank, E. Sensory axon regeneration: Rebuilding functional connections in the spinal cord. Trends Neurosci. 2012, 35, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bo, X.; Wu, D.; Yeh, J.; Zhang, Y. Gene therapy approaches for neuroprotection and axonal regeneration after spinal cord and spinal root injury. Curr. Gene Ther. 2011, 11, 101–115. [Google Scholar] [CrossRef]
- Ramer, M.S.; Priestley, J.V.; McMahon, S.B. Functional regeneration of sensory axons into the adult spinal cord. Nature 2000, 403, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Ramer, M.S.; Bishop, T.; Dockery, P.; Mobarak, M.S.; O’Leary, D.; Fraher, J.P.; Priestley, J.V.; McMahon, S.B. Neurotrophin-3-mediated regeneration and recovery of proprioception following dorsal rhizotomy. Mol. Cell. Neurosci. 2002, 19, 239–249. [Google Scholar] [CrossRef]
- Wang, R.; King, T.; Ossipov, M.H.; Rossomando, A.J.; Vanderah, T.W.; Harvey, P.; Cariani, P.; Frank, E.; Sah, D.W.Y.; Porreca, F. Persistent restoration of sensory function by immediate or delayed systemic artemin after dorsal root injury. Nat. Neurosci. 2008, 11, 488–496. [Google Scholar] [CrossRef] [Green Version]
- Harvey, P.; Gong, B.; Rossomando, A.J.; Frank, E. Topographically specific regeneration of sensory axons in the spinal cord. Proc. Natl Acad. Sci. USA 2010, 107, 11585–11590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, L.E.; Gibson, M.E.; Arnold, H.M.; Pepinsky, B.; Frank, E. Artemin promotes functional long-distance axonal regeneration to the brainstem after dorsal root crush. Proc. Natl Acad. Sci. USA 2015, 112, 6170–6175. [Google Scholar] [CrossRef] [Green Version]
- Hoeber, J.; König, N.; Trolle, C.; Lekholm, E.; Zhou, C.; Pankratova, S.; Åkesson, E.; Fredriksson, R.; Aldskogius, H.; Kozlova, E.N. A Combinatorial Approach to Induce Sensory Axon Regeneration into the Dorsal Root Avulsed Spinal Cord. Stem Cells Dev. 2017, 26, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.V.; Silva, M.V.R.D.; Chiarotto, G.B.; Volpe, B.B.; Santana, M.H.; Malheiros Luzo, Â.C.; Oliveira, A.L.R. Reflex arc recovery after spinal cord dorsal root repair with platelet rich plasma (PRP). Brain Res. Bull. 2019, 152, 212–224. [Google Scholar] [CrossRef]
- Teng, F.Y.; Tang, B.L. Axonal regeneration in adult CNS neurons—signaling molecules and pathways. J. Neurochem. 2006, 96, 1501–1508. [Google Scholar] [CrossRef]
- Hanna-Mitchell, A.T.; O’Leary, D.; Mobarak, M.S.; Ramer, M.S.; McMahon, S.B.; Priestley, J.V.; Kozlova, E.N.; Aldskogius, H.; Dockery, P.; Fraher, J.P. The impact of neurotrophin-3 on the dorsal root transitional zone following injury. Spinal Cord 2008, 46, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Ramer, M.S.; Duraisingam, I.; Priestley, J.V.; McMahon, S.B. Two-tiered inhibition of axon regeneration at the dorsal root entry zone. J. Neurosci. 2001, 21, 2651–2660. [Google Scholar] [CrossRef]
- Park, K.K.; Liu, K.; Hu, Y.; Smith, P.D.; Wang, C.; Cai, B.; Xu, B.; Connolly, L.; Kramvis, I.; Sahin, M.; et al. Promoting axon regeneration in the adult CNS by modulation of the PTEN/mTOR pathway. Science 2008, 322, 963–966. [Google Scholar] [CrossRef] [Green Version]
- Park, K.K.; Liu, K.; Hu, Y.; Kanter, J.L.; He, Z. PTEN/mTOR and axon regeneration. Exp Neurol. 2010, 223, 45–50. [Google Scholar] [CrossRef]
- Ohtake, Y.; Hayat, U.; Li, S. PTEN inhibition and axon regeneration and neural repair. Neural Regen. Res. 2015, 10, 1363–1368. [Google Scholar] [CrossRef]
- Bhowmick, S.; Abdul-Muneer, P.M. PTEN Blocking Stimulates Corticospinal and Raphespinal Axonal Regeneration and Promotes Functional Recovery After Spinal Cord Injury. J. Neuropathol. Exp. Neurol. 2021, 80, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Luan, L.; Li, J.; Yang, L. MiR-212-3p improves rat functional recovery and inhibits neurocyte apoptosis in spinal cord injury models via PTEN downregulation-mediated activation of AKT/mTOR pathway. Brain Res. 2021, 1768, 147576. [Google Scholar] [CrossRef]
- Kar, A.N.; Lee, S.J.; Sahoo, P.K.; Thames, E.; Yoo, S.; Houle, J.D.; Twiss, J.L. MicroRNAs 21 and 199a-3p Regulate Axon Growth Potential through Modulation of Pten and mTor mRNAs. eNeuro 2021, 8, ENEURO.0155–21.2021. [Google Scholar] [CrossRef]
- Christie, K.J.; Zochodne, D. Peripheral axon regrowth: New molecular approaches. Neuroscience 2013, 240, 310–324. [Google Scholar] [CrossRef]
- Cong, M.; Shen, M.; Wu, X.; Li, Y.; Wang, L.; He, Q.; Shi, H.; Ding, F. Improvement of sensory neuron growth and survival via negatively regulating PTEN by miR-21-5p-contained small extracellular vesicles from skin precursor-derived Schwann cells. Stem Cell Res. Ther. 2021, 12, 80. [Google Scholar] [CrossRef] [PubMed]
- Gutilla, E.A.; Steward, O. Selective neuronal PTEN deletion: Can we take the brakes off of growth without losing control? Neural Regen. Res. 2016, 11, 1201–1203. [Google Scholar] [CrossRef]
- Goncalves, M.B.; Malmqvist, T.; Clarke, E.; Hubens, C.J.; Grist, J.; Hobbs, C.; Trigo, D.; Risling, M.; Angeria, M.; Damberg, P.; et al. Neuronal RARβ Signaling Modulates PTEN Activity Directly in Neurons and via Exosome Transfer in Astrocytes to Prevent Glial Scar Formation and Induce Spinal Cord Regeneration. J. Neurosci. 2015, 35, 15731–15745. [Google Scholar] [CrossRef]
- Goncalves, M.B.; Wu, Y.; Trigo, D.; Clarke, E.; Malmqvist, T.; Grist, J.; Hobbs, C.; Carlstedt, T.P.; Corcoran, J.P.T. Retinoic acid synthesis by NG2 expressing cells promotes a permissive environment for axonal outgrowth. Neurobiol. Dis. 2018, 111, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Hur, E.M.; Zhou, F.Q. Growing the growth cone: Remodeling the cytoskeleton to promote axon regeneration. Trends Neurosci. 2012, 35, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.Q.; Zhang, G.; Jamison, C., Jr.; Takano, H.; Haydon, P.G.; Selzer, M.E. Axon regeneration in the absence of growth cones: Acceleration by cyclic AMP. J. Comp. Neurol. 2009, 515, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Hellal, F.; Hurtado, A.; Ruschel, J.; Flynn, K.C.; Laskowski, C.J.; Umlauf, M.; Kapitein, L.C.; Strikis, D.; Lemmon, V.; Bixby, J.; et al. Microtubule stabilization reduces scarring and causes axon regeneration after spinal cord injury. Science 2011, 331, 928–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Liu, M.; Son, Y.J.; Timothy Himes, B.; Snow, D.M.; Yu, W.; Baas, P.W. Inhibition of Kinesin-5, a microtubule-based motor protein, as a strategy for enhancing regeneration of adult axons. Traffic 2011, 12, 269–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, T.O.; Matamoros, A.J.; Friedman, J.M.; Friedman, A.J.; Nacharaju, P.; Yu, W.; Sharp, D.J.; Baas, P.W. Nanoparticle Delivery of Fidgetin siRNA as a Microtubule-based Therapy to Augment Nerve Regeneration. Sci. Rep. 2017, 7, 9675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamoros, A.J.; Tom, V.J.; Wu, D.; Rao, Y.; Sharp, D.J.; Baas, P.W. Knockdown of Fidgetin Improves Regeneration of Injured Axons by a Microtubule-Based Mechanism. J. Neurosci. 2019, 39, 2011–2024. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Jin, Y.; Shapiro, T.M.; Hinduja, A.; Baas, P.W.; Tom, V.J. Chronic neuronal activation increases dynamic microtubules to enhance functional axon regeneration after dorsal root crush injury. Nat. Commun. 2020, 11, 6131. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Zhang, M.; Li, P.; Tang, W.; Liu, Y.; Hu, Y.; Cui, Q.; Liu, M.; Jiang, J. Bioinformatics analysis of long non-coding RNAs involved in nerve regeneration following sciatic nerve injury. Mol. Pain. 2020, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, S.; He, J.H. Transcriptomic analysis reveals essential microRNAs after peripheral nerve injury. Neural Regen. Res. 2021, 16, 1865–1870. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Huang, T.; Chen, Y.; Shen, L.; Zhou, S.; Zhang, S.; Yu, B. Circ-Spidr enhances axon regeneration after peripheral nerve injury. Cell Death Dis. 2019, 10, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Jia, Y.; Tang, W.; Cui, Q.; Liu, M.; Jiang, J. Roles of Non-coding RNAs in Central Nervous System Axon Regeneration. Front. Neurosci. 2021, 15, 630633. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, A.D.; Mandrekar-Colucci, S.; Hall, J.C.; Sweet, D.R.; Schmitt, P.J.; Xu, X.; Guan, Z.; Mo, X.; Guerau-de-Arellano, M.; Popovich, P.G. miR-155 Deletion in Mice Overcomes Neuron-Intrinsic and Neuron-Extrinsic Barriers to Spinal Cord Repair. J. Neurosci. 2016, 36, 8516–8532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Tian, Z.; He, L.; Liu, C.; Wang, N.; Rong, L.; Liu, B. Exosomes derived from miR-26a-modified MSCs promote axonal regeneration via the PTEN/AKT/mTOR pathway following spinal cord injury. Stem Cell Res. Ther. 2021, 12, 224. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, B.; Wang, Z.; Yuan, X.; Chen, C.; Zhang, Y.; Xia, Z.; Wang, X.; Yu, M.; Tao, W.; et al. miR-155-5p Promotes Dorsal Root Ganglion Neuron Axonal Growth in an Inhibitory Microenvironment via the cAMP/PKA Pathway. Int. J. Biol. Sci. 2019, 15, 1557–1570. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Li, B.; Yuan, X.; Cui, L.; Wang, Z.; Zhang, Y.; Yu, M.; Xiu, Y.; Zhang, Z.; Li, W.; et al. MiR-20a Plays a Key Regulatory Role in the Repair of Spinal Cord Dorsal Column Lesion via PDZ-RhoGEF/RhoA/GAP43 Axis in Rat. Cell. Mol. Neurobiol. 2019, 39, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Suzuki, H. Emerging roles of microRN9As in chronic pain. Neurochem. Int. 2014, 77, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Bono, J.; Tao, Y.X. Long noncoding RNA (lncRNA): A target in neuropathic pain. Expert Opin. Ther. Targets. 2019, 23, 15–20. [Google Scholar] [CrossRef]
- Tang, B.L. Axon regeneration induced by environmental enrichment- epigenetic mechanisms. Neural Regen. Res. 2020, 15, 10–15. [Google Scholar] [CrossRef]
- Weng, Y.L.; An, R.; Cassin, J.; Joseph, J.; Mi, R.; Wang, C.; Zhong, C.; Jin, S.G.; Pfeifer, G.; Bellacosa, A.; et al. An Intrinsic Epigenetic Barrier for Functional Axon Regeneration. Neuron 2017, 94, 37–346.e6. [Google Scholar] [CrossRef] [Green Version]
- Palmisano, I.; Danzi, M.C.; Hutson, T.H.; Zhou, L.; McLachlan, E.; Serger, E.; Shkura, K.; Srivastava, P.K.; Hervera, A.; Neill, N.O.; et al. Epigenomic signatures underpin the axonal regenerative ability of dorsal root ganglia sensory neurons. Nat. Neurosci. 2019, 22, 1913–1924. [Google Scholar] [CrossRef]
- Shin, H.Y.; Kwon, M.J.; Lee, E.M.; Kim, K.; Oh, Y.J.; Kim, H.S.; Hwang, D.H.; Kim, B.G. Role of Myc Proto-Oncogene as a Transcriptional Hub to Regulate the Expression of Regeneration-Associated Genes following Preconditioning Peripheral Nerve Injury. J. Neurosci. 2021, 41, 446–460. [Google Scholar] [CrossRef]
- Hutson, T.H.; Kathe, C.; Palmisano, I.; Bartholdi, K.; Hervera, A.; De Virgiliis, F.; McLachlan, E.; Zhou, L.; Kong, G.; Barraud, Q.; et al. Cbp-dependent histone acetylation mediates axon regeneration induced by environmental enrichment in rodent spinal cord injury models. Sci. Transl. Med. 2019, 11, eaaw2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Virgiliis, F.; Hutson, T.H.; Palmisano, I.; Amachree, S.; Miao, J.; Zhou, L.; Todorova, R.; Thompson, R.; Danzi, M.C.; Lemmon, V.P.; et al. Enriched conditioning expands the regenerative ability of sensory neurons after spinal cord injury via neuronal intrinsic redox signaling. Nat. Commun. 2020, 11, 6425. [Google Scholar] [CrossRef] [PubMed]
- Hutson, T.H.; Di Giovanni, S. The translational landscape in spinal cord injury: Focus on neuroplasticity and regeneration. Nat. Rev. Neurol. 2019, 15, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Kisucká, A.; Bimbová, K.; Bačová, M.; Gálik, J.; Lukáčová, N. Microglial/astroglial interactions—An important therapeutic target in treatment of spinal cord injury. Cells 2021, 10, 1943. [Google Scholar] [CrossRef]
- Liu, L.; Rudin, M.; Kozlova, E.N. Glial cell proliferation in the spinal cord after dorsal rhizotomy or sciatic nerve transection in the adult rat. Exp. Brain Res. 2000, 131, 64–73. [Google Scholar] [CrossRef]
- Kozlova, E.N. Differentiation and migration of astrocytes in the spinal cord following dorsal root injury in the adult rat. Eur. J. Neurosci. 2003, 17, 782–790. [Google Scholar] [CrossRef]
- Fraher, J.P. The transitional zone and CNS regeneration. J. Anat. 1999, 194 (Pt 2), 161–182, Corrected and republished in: J. Anat. 2000, 196 (Pt 1), 137–158. [Google Scholar] [CrossRef]
- Domeniconi, M.; Filbin, M.T. Overcoming inhibitors in myelin to promote axonal regeneration. J Neurol Sci. 2005, 233, 43–47. [Google Scholar] [CrossRef]
- Kozlova, E.N.; Lukanidin, E. Metastasis-associated Mts1 (S100A4) protein is selectively expressed in white matter astrocytes and upregulated after dorsal root or peripheral nerve injury. Glia 1999, 27, 249–258. [Google Scholar] [CrossRef]
- Fang, Z.; Forslund, N.; Takenaga, K.; Lukanidin, E.; Kozlova, E.N. Sensory neurite outgrowth on white matter astrocytes is influenced by intracellular and extracellular S100A4 protein. J. Neurosci. Res. 2006, 83, 619–626. [Google Scholar] [CrossRef]
- D’Ambrosi, N.; Milani, M.; Apolloni, S. S100A4 in the Physiology and Pathology of the Central and Peripheral Nervous System. Cells 2021, 10, 798. [Google Scholar] [CrossRef]
- Trolle, C.; Ivert, P.; Hoeber, J.; Rocamonde-Lago, I.; Vasylovska, S.; Lukanidin, E.; Kozlova, E.N. Boundary cap neural crest stem cell transplants contribute Mts1/S100A4-expressing cells in the glial scar. Regen. Med. 2017, 12, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, M.P.; Horn, K.P.; Tom, V.J.; Miller, J.H.; Busch, S.A.; Nair, D.; Silver, D.J.; Silver, J. Chronic enhancement of the intrinsic growth capacity of sensory neurons combined with the degradation of inhibitory proteoglycans allows functional regeneration of sensory axons through the dorsal root entry zone in the mammalian spinal cord. J. Neurosci. 2005, 25, 8066–8076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaglia, X.; Beggah, A.T.; Seidenbecher, C.; Zurn, A.D. Delayed priming promotes CNS regeneration post-rhizotomy in Neurocan and Brevican-deficient mice. Brain 2008, 131, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Harvey, P.A.; Lee, D.H.; Qian, F.; Weinreb, P.H.; Frank, E. Blockade of Nogo receptor ligands promotes functional regeneration of sensory axons after dorsal root crush. J. Neurosci. 2009, 29, 6285–6295. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Zhou, Z.; Hu, J.; Fink, D.J.; Mata, M. Soluble nogo receptor down-regulates expression of neuronal Nogo-A to enhance axonal regeneration. J. Biol. Chem. 2010, 285, 2783–2795. [Google Scholar] [CrossRef] [Green Version]
- Cheah, M.; Andrews, M.R.; Chew, D.J.; Moloney, E.B.; Verhaagen, J.; Fässler, R.; Fawcett, J.W. Expression of an activated integrin promotes Long-Distance sensory axon regeneration in the spinal cord. J. Neurosci. 2016, 36, 7283–7297. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Kim, H.; Han, S.B.; Manire, M.; Yoo, R.; Pang, S.; Smith, G.M.; Son, Y.J. Co-targeting myelin inhibitors and CSPGs markedly enhances regeneration of GDNF-stimulated, but not conditioning-lesioned, sensory axons into the spinal cord. Elife 2021, 10, e63050. [Google Scholar] [CrossRef]
- Carlstedt, T. Regenerating axons form nerve terminals at astrocytes. Brain Res. 1985, 347, 188–191. [Google Scholar] [CrossRef]
- Liuzzi, F.J.; Lasek, R.J. Astrocytes block axonal regeneration in mammals by activating the physiological stop pathway. Science 1987, 237, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, A.; Skuba, A.; Himes, B.T.; Bhagat, S.L.; Hyun, J.K.; Tessler, A.; Bishop, D.; Son, Y.-J. In vivo imaging of dorsal root regeneration: Rapid immobilization and presynaptic differentiation at the CNS/PNS border. J. Neurosci. 2011, 31, 4569–4582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.B.; Kim, H.; Skuba, A.; Tessler, A.; Ferguson, T.; Son, Y.J. Sensory Axon Regeneration: A Review from an in vivo Imaging Perspective. Exp Neurobiol. 2012, 21, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Filous, A.R.; Tran, A.; Howell, C.J.; Busch, S.A.; Evans, T.A.; Stallcup, W.B.; Kang, S.H.; Bergles, D.E.; Lee, S.I.; Levine, J.M.; et al. Entrapment via synaptic-like connections between NG2 proteoglycan+ cells and dystrophic axons in the lesion plays a role in regeneration failure after spinal cord injury. J. Neurosci. 2014, 34, 16369–16384. [Google Scholar] [CrossRef] [PubMed]
- Bignami, A.; Dahl, D.; Nguyen, B.T.; Crosby, C.J. The fate of axonal debris in Wallerian degeneration of rat optic and sciatic nerves. Electron microscopy and immunofluorescence studies with neurofilament antisera. J. Neuropathol. Exp. Neurol. 1981, 40, 537–550. [Google Scholar] [CrossRef]
- George, R.; Griffin, J.W. Delayed macrophage responses and myelin clearance during Wallerian degeneration in the central nervous system: The dorsal radiculotomy model. Exp. Neurol. 1994, 129, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Persson, J.K.; Svensson, M.; Aldskogius, H. Glial cell responses, complement, and clusterin in the central nervous system following dorsal root transection. Glia 1998, 23, 221–238. [Google Scholar] [CrossRef]
- Rotshenker, S. Wallerian degeneration: The innate-immune response to traumatic nerve injury. J. Neuroinflammation 2011, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- McPhail, L.T.; Borisoff, J.F.; Tsang, B.; Hwi, L.P.; Kwiecien, J.M.; Ramer, M.S. Protracted myelin clearance hinders central primary afferent regeneration following dorsal rhizotomy and delayed neurotrophin-3 treatment. Neurosci Lett. 2007, 411, 206–211. [Google Scholar] [CrossRef]
- Hosmane, S.; Tegenge, M.A.; Rajbhandari, L.; Uapinyoying, P.; Kumar, N.G.; Thakor, N.; Venkatesan, A. Toll/Interleukin-1 Receptor Domain-Containing Adapter Inducing Interferon-β Mediates Microglial Phagocytosis of Degenerating Axons. J. Neurosci. 2012, 32, 7745–7757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajbhandari, L.; Tegenge, M.A.; Shrestha, S.; Ganesh Kumar, N.; Malik, A.; Mithal, A.; Hosmane, S.; Venkatesan, A. Toll-like receptor 4 deficiency impairs microglial phagocytosis of degenerating axons. Glia 2014, 62, 1982–1991. [Google Scholar] [CrossRef]
- Farah, M.H.; Pan, B.H.; Hoffman, P.N.; Ferraris, D.; Tsukamoto, T.; Nguyen, T.; Wong, P.C.; Price, D.L.; Slusher, B.S.; Griffin, J.W. Reduced BACE (beta-amyloid precursor cleaving enzyme) 1 activity enhances clearance of myelin debris and regeneration of axons in the injured peripheral nervous system. J. Neurosci. 2011, 31, 5744–5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallon, C.; Rockenstein, E.; Masliah, E.; Farah, M.H. Increased BACE1 activity inhibits peripheral nerve regeneration after injury. Neurobiol Dis. 2017, 106, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Elberg, G.; Liraz-Zaltsman, S.; Reichert, F.; Matozaki, T.; Tal, M.; Rotshenker, S. Deletion of SIRPα (signal regulatory protein-α) promotes phagocytic clearance of myelin debris in Wallerian degeneration, axon regeneration, and recovery from nerve injury. J. Neuroinflammation 2019, 16, 277. [Google Scholar] [CrossRef] [PubMed]
- Daniel, P.M.; Strich, S.J. Histological observations on Wallerian degeneration in the spinal cord of the baboon, Papio papio. Acta Neuropathol. 1969, 12, 314–328. [Google Scholar] [CrossRef]
- Bignami, A.; Ralston, H.J., 3rd. The cellular reaction to Wallerian degeneration in the central nervous system of the cat. Brain Res. 1969, 13, 444–461. [Google Scholar] [CrossRef]
- Aldskogius, H. Indirect and direct Wallerian degeneration in the intramedullary root fibres of the hypoglossal nerve. An electron microscopical study in the kitten. Adv. Anat. Embryol. Cell Biol. 1974, 50, 7–78. [Google Scholar]
- Amann, L.; Prinz, M. The origin, fate and function of macrophages in the peripheral nervous system-an update. Int. Immunol. 2020, 32, 709–717. [Google Scholar] [CrossRef]
- Ochocka, N.; Kaminska, B. Microglia Diversity in Healthy and Diseased Brain: Insights from Single-Cell Omics. Int. J. Mol. Sci. 2021, 22, 3027. [Google Scholar] [CrossRef] [PubMed]
- Chew, D.J.; Carlstedt, T.; Shortland, P.J. A comparative histological analysis of two models of nerve root avulsion injury in the adult rat. Neuropathol. Appl. Neurobiol. 2011, 37, 613–632. [Google Scholar] [CrossRef] [PubMed]
- Anjum, A.; Yazid, M.D.; Fauzi Daud, M.; Idris, J.; Ng, A.M.H.; Selvi Naicker, A.; Ismail, O.H.R.; Athi Kumar, R.K.; Lokanathan, Y. Spinal Cord Injury: Pathophysiology, Multimolecular Interactions, and Underlying Recovery Mechanisms. Int. J. Mol. Sci. 2020, 21, 7533. [Google Scholar] [CrossRef]
- Guerout, N. Plasticity of the injured spinal cord. Cells 2021, 10, 1886. [Google Scholar] [CrossRef]
- Guo, S.; Redenski, I.; Levenberg, S. Spinal Cord Repair: From Cells and Tissue Engineering to Extracellular Vesicles. Cells 2021, 10, 1872. [Google Scholar] [CrossRef]
- Lin, C.; Ekblad-Nordberg, Å.; Michaëlsson, J.; Götherström, C.; Hsu, C.-C.; Ye, H.; Johansson, J.; Rising, A.; Sundström, E.; Åkesson, E. In Vitro Study of Human Immune Responses to Hyaluronic Acid Hydrogels, Recombinant Spidroins and Human Neural Progenitor Cells of Relevance to Spinal Cord Injury Repair. Cells 2021, 10, 1713. [Google Scholar] [CrossRef]
- Rodríguez-Barrera, R.; Rivas-González, M.; García-Sánchez, J.; Mojica-Torres, D.; Ibarra, A. Neurogenesis after Spinal Cord Injury: State of the Art. Cells 2021, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Sims, T.J.; Gilmore, S.A. Regeneration of dorsal root axons into experimentally altered glial environments in the rat spinal cord. Exp. Brain Res. 1994, 99, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Prewitt, C.M.; Niesman, I.R.; Kane, C.J.; Houlé, J.D. Activated macrophage/microglial cells can promote the regeneration of sensory axons into the injured spinal cord. Exp. Neurol. 1997, 148, 433–443. [Google Scholar] [CrossRef]
- Afshari, F.T.; Kwok, J.C.; Fawcett, J.W. Astrocyte-produced ephrins inhibit schwann cell migration via VAV2 signaling. J Neurosci. 2010, 30, 4246–4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afshari, F.T.; Kwok, J.C.; White, L.; Fawcett, J.W. Schwann cell migration is integrin-dependent and inhibited by astrocyte-produced aggrecan. Glia 2010, 58, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Horn, K.P.; Busch, S.A.; Hawthorne, A.L.; van Rooijen, N.; Silver, J. Another barrier to regeneration in the CNS: Activated macrophages induce extensive retraction of dystrophic axons through direct physical interactions. J. Neurosci. 2008, 28, 9330–9341. [Google Scholar] [CrossRef]
- Kozlova, E.N.; Seiger, A.; Aldskogius, H. Human dorsal root ganglion neurons from embryonic donors extend axons into the host rat spinal cord along laminin peripheral surroundings of the dorsal root transitional zone. J. Neurocytol. 1997, 26, 811–822. [Google Scholar] [CrossRef]
- Levinsson, A.; Holmberg, H.; Schouenborg, J.; Seiger, A.; Aldskogius, H.; Kozlova, E.N. Functional connections are established in the deafferented rat spinal cord by peripherally transplanted human embryonic sensory neurons. Eur. J. Neurosci. 2000, 12, 3589–3595. [Google Scholar] [CrossRef] [PubMed]
- Minkelyte, K.; Collins, A.; Liadi, M.; Ibrahim, A.; Li, D.; Li, Y. High-Yield Mucosal Olfactory Ensheathing Cells Restore Loss of Function in Rat Dorsal Root Injury. Cells 2021, 10, 1186. [Google Scholar] [CrossRef] [PubMed]
- Trolle, C.; Konig, N.; Abrahamsson, N.; Vasylovska, S.; Kozlova, E.N. Boundary cap neural crest stem cells homotopically implanted to the injured dorsal root transitional zone give rise to different types of neurons and glia in adult rodents. BMC Neurosci. 2014, 15, 60. [Google Scholar] [CrossRef] [Green Version]
- König, N.; Trolle, C.; Kapuralin, K.; Adameyko, I.; Mitrecic, D.; Aldskogius, H.; Shortland, P.J.; Kozlova, E.N. Murine neural crest stem cells and embryonic stem cell-derived neuron precursors survive and differentiate after transplantation in a model of dorsal root avulsion. J. Tissue Eng. Regen. Med. 2017, 11, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Hoeber, J.; Trolle, C.; König, N.; Du, Z.; Gallo, A.; Hermans, E.; Aldskogius, H.; Shortland, P.; Zhang, S.C.; Deumens, R.; et al. Human Embryonic Stem Cell-Derived Progenitors Assist Functional Sensory Axon Regeneration after Dorsal Root Avulsion Injury. Sci. Rep. 2015, 5, 10666. [Google Scholar] [CrossRef] [Green Version]
- de Castro, M.V.; da Silva, M.V.R.; Chiarotto, G.B.; Santana, M.H.A.; Luzo, Â.C.M.; Kyrylenko, S.; de Oliveira, A.L.R. Spinal Reflex Recovery after Dorsal Rhizotomy and Repair with Platelet-Rich Plasma (PRP) Gel Combined with Bioengineered Human Embryonic Stem Cells (hESCs). Stem Cells Int. 2020, 2020, 8834360. [Google Scholar] [CrossRef]
- Andjus, P.; Kosanović, M.; Milićević, K.; Gautam, M.; Vainio, S.J.; Jagečić, D.; Kozlova, E.N.; Pivoriūnas, A.; Chachques, J.C.; Sakaj, M.; et al. Extracellular Vesicles as Innovative Tool for Diagnosis, Regeneration and Protection against Neurological Damage. Int. J. Mol. Sci. 2020, 21, 6859. [Google Scholar] [CrossRef]
- Richardson, P.M.; McGuinness, U.M.; Aguayo, A.J. Axons from CNS neurons regenerate into PNS grafts. Nature 1980, 284, 264–265. [Google Scholar] [CrossRef]
- David, S.; Aguayo, A.J. Axonal elongation into peripheral nervous system “bridges” after central nervous system injury in adult rats. Science 1981, 214, 931–933. [Google Scholar] [CrossRef]
- Benfey, M.; Aguayo, A.J. Extensive elongation of axons from rat brain into peripheral nerve grafts. Nature 1982, 296, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.M.; McGuinness, U.M.; Aguayo, A.J. Peripheral nerve autografts to the rat spinal cord: Studies with axonal tracing methods. Brain Res. 1982, 237, 147–612. [Google Scholar] [CrossRef]
- McPhail, L.T.; Plunet, W.T.; Das, P.; Ramer, M.S. The astrocytic barrier to axonal regeneration at the dorsal root entry zone is induced by rhizotomy. Eur. J. Neurosci. 2005, 21, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Selim, O.; Lakhani, S.; Midha, S.; Mosahebi, A.; Kalaskar, D.M. Three-Dimensional Engineered Peripheral Nerve: Toward a New Era of Patient-Specific Nerve Repair Solutions. Tissue Eng. Part B Rev. 2021. [Google Scholar] [CrossRef]
- Behbehani, M.; Glen, A.; Taylor, C.S.; Schuhmacher, A.; Claeyssens, F.; Haycock, J.W. Pre-clinical evaluation of advanced nerve guide conduits using a novel 3D in vitro testing model. Int. J. Bioprint. 2017, 4, 123. [Google Scholar] [CrossRef]
- Jeffries, G.D.M.; Xu, S.; Lobovkina, T.; Kirejev, V.; Tusseau, F.; Gyllensten, C.; Singh, A.K.; Karila, P.; Moll, L.; Orwar, O. 3D micro-organisation printing of mammalian cells to generate biological tissues. Sci Rep. 2020, 10, 19529. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.B.; Spearman, B.S.; Hlavac, N.; Schmidt, C.E. Three-Dimensional Bioprinted Hyaluronic Acid Hydrogel Test Beds for Assessing Neural Cell Responses to Competitive Growth Stimuli. ACS Biomater. Sci. Eng. 2020, 6, 6819–6830. [Google Scholar] [CrossRef]
Figure 1.
Intact, the lumbar dorsal root of the adult rat. Horizontal section through several dorsal rootlets, immunostained with antibodies to laminin (green) and glial fibrillary acidic protein (GFAP; red). The dorsal root transitional zone is formed by cone-shaped peripheral projections of central nervous tissue, so-called central tissue projections (CTP; red), which interdigitate with the peripheral nervous compartment of the root (green). ×200. Reproduced with permission from Aldskogius, H., Kozlova,
E.N., Brain Research Reviews; published by Elsevier, 2002 [35].
Figure 1.
Intact, the lumbar dorsal root of the adult rat. Horizontal section through several dorsal rootlets, immunostained with antibodies to laminin (green) and glial fibrillary acidic protein (GFAP; red). The dorsal root transitional zone is formed by cone-shaped peripheral projections of central nervous tissue, so-called central tissue projections (CTP; red), which interdigitate with the peripheral nervous compartment of the root (green). ×200. Reproduced with permission from Aldskogius, H., Kozlova,
E.N., Brain Research Reviews; published by Elsevier, 2002 [35].

Figure 2.
Lumbar dorsal roots following ipsilateral dorsal root injury. Horizontal sections, immunostained with antibodies to laminin (green) and GFAP (red). The extensive outgrowth of astroglial processes (red and yellow) into the peripheral compartment is shown at one week (above) and two weeks (below) following injury. ×200. Reproduced with permission from Aldskogius, H., Kozlova,
E.N., Brain Research Reviews; published by Elsevier, 2002 [35].
Figure 2.
Lumbar dorsal roots following ipsilateral dorsal root injury. Horizontal sections, immunostained with antibodies to laminin (green) and GFAP (red). The extensive outgrowth of astroglial processes (red and yellow) into the peripheral compartment is shown at one week (above) and two weeks (below) following injury. ×200. Reproduced with permission from Aldskogius, H., Kozlova,
E.N., Brain Research Reviews; published by Elsevier, 2002 [35].

Figure 3.
Scheme of experiment with human embryonic/fetal dorsal root ganglion (DRG) transplant (T) in the cavity of the extirpated native DRG of an adult rat recipient. Sensory axons from the transplant grow through the host DREZ and make synaptic contacts in the host spinal cord. For further information. Reproduced with permission from Kozlova, E.N., Seiger, A.,
Aldskogius, H., Journal of Neurocytology; published by Springer, 1997 [152,153].
Figure 3.
Scheme of experiment with human embryonic/fetal dorsal root ganglion (DRG) transplant (T) in the cavity of the extirpated native DRG of an adult rat recipient. Sensory axons from the transplant grow through the host DREZ and make synaptic contacts in the host spinal cord. For further information. Reproduced with permission from Kozlova, E.N., Seiger, A.,
Aldskogius, H., Journal of Neurocytology; published by Springer, 1997 [152,153].

Figure 4.
Two options based on the use of stem cell-based transplants at the DREZ, to facilitate entry of injured dorsal root axons (left) or to create a neuronal relay between dorsal root axons (forming synapses with the relay) and the host spinal cord circuitry (right).
Figure 4.
Two options based on the use of stem cell-based transplants at the DREZ, to facilitate entry of injured dorsal root axons (left) or to create a neuronal relay between dorsal root axons (forming synapses with the relay) and the host spinal cord circuitry (right).

Figure 5.
Principle scheme to use the DREZ as an entry point for axons growing in a bridge circumventing a spinal cord injury. (A) Axons from injured descending pathways (S) grow into and within a peripheral nerve graft (or biomimetic conduit) and may be able to enter the spinal cord through the intact or drug/cell modified DREZ. (B) Axons from an intraspinal transplant (T) grow through the bridge and may, because of their immaturity, be able to grow through the intact or reactive DREZ.
Figure 5.
Principle scheme to use the DREZ as an entry point for axons growing in a bridge circumventing a spinal cord injury. (A) Axons from injured descending pathways (S) grow into and within a peripheral nerve graft (or biomimetic conduit) and may be able to enter the spinal cord through the intact or drug/cell modified DREZ. (B) Axons from an intraspinal transplant (T) grow through the bridge and may, because of their immaturity, be able to grow through the intact or reactive DREZ.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aldskogius, H.; Kozlova, E.N. Dorsal Root Injury—A Model for Exploring Pathophysiology and Therapeutic Strategies in Spinal Cord Injury. Cells 2021, 10, 2185. https://doi.org/10.3390/cells10092185
AMA Style
Aldskogius H, Kozlova EN. Dorsal Root Injury—A Model for Exploring Pathophysiology and Therapeutic Strategies in Spinal Cord Injury. Cells. 2021; 10(9):2185. https://doi.org/10.3390/cells10092185
Chicago/Turabian StyleAldskogius, Håkan, and Elena N. Kozlova. 2021. "Dorsal Root Injury—A Model for Exploring Pathophysiology and Therapeutic Strategies in Spinal Cord Injury" Cells 10, no. 9: 2185. https://doi.org/10.3390/cells10092185
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.