
Differential Response of Müller Cells and Microglia in a Mouse Retinal Detachment Model and Its Implications in Detached and Non-Detached Regions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and RD Generation
2.2. ERG
2.3. Hematoxylin and Eosin (H&E) Staining
2.4. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
2.5. Immunohistochemistry
2.6. Statistical Analysis
3. Results
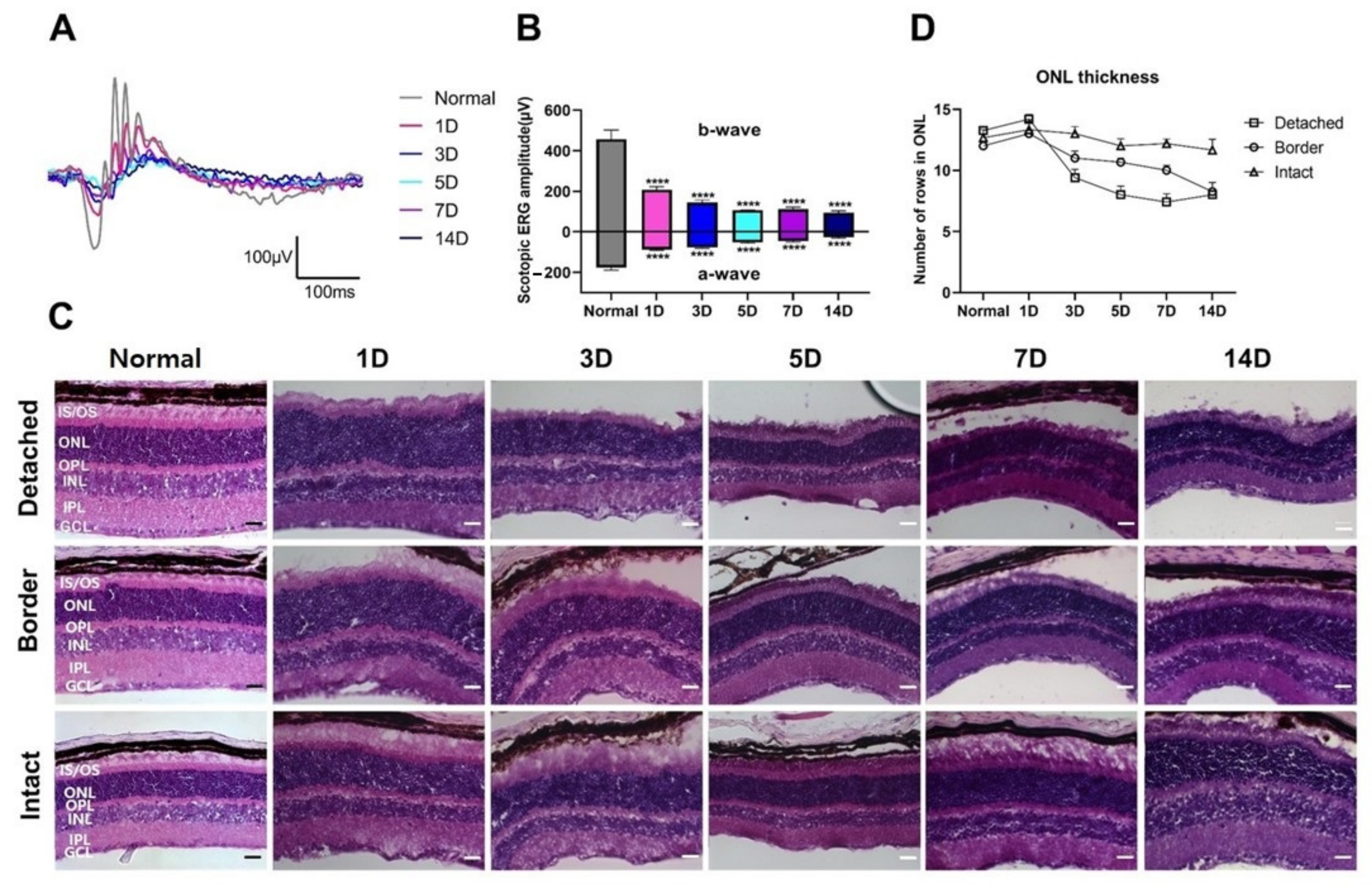
3.1. Functional and Histological Changes in Experimental RD
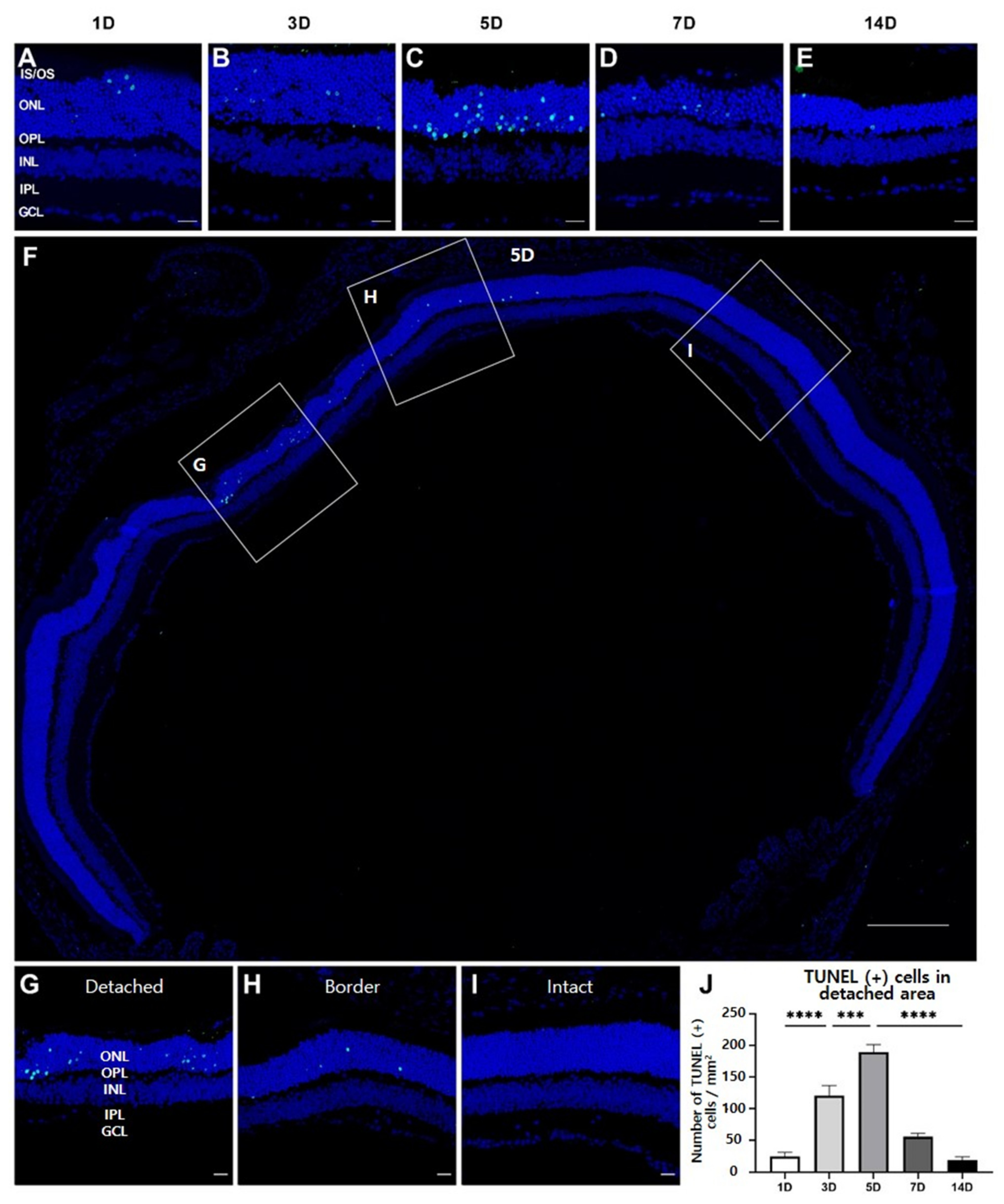
3.2. Photoreceptor Degeneration in Experimental RD
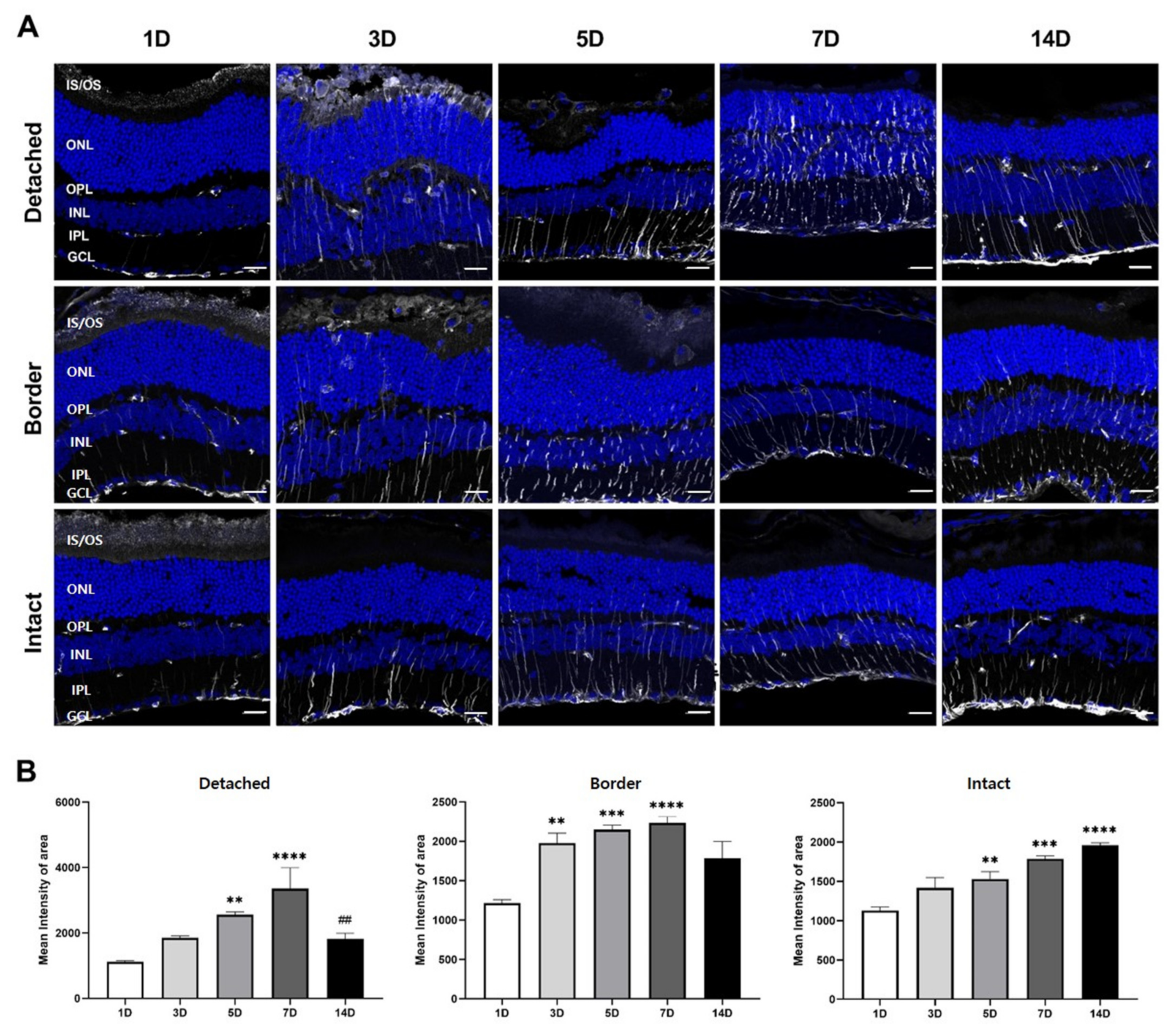
3.3. GFAP Expression in Müller Glial Cells in RD
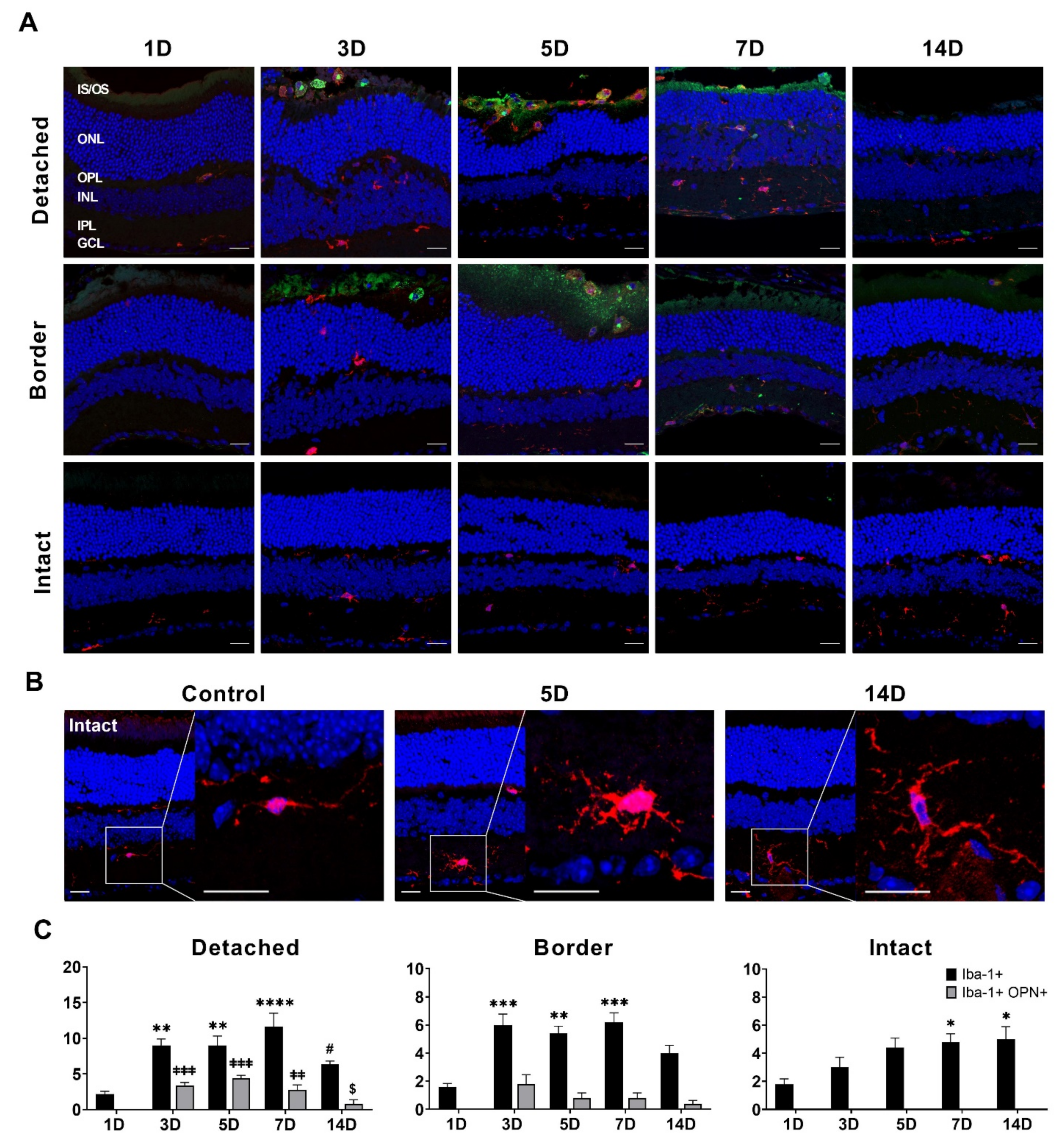
3.4. OPN Expression in Iba-1-Immunolabeled Microglial Cell in RD
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feltgen, N.; Walter, P. Rhegmatogenous retinal detachment--an ophthalmologic emergency. Dtsch Arztebl Int. 2014, 111, 12–21; quiz 22. [Google Scholar] [CrossRef] [Green Version]
- Dunaief, J.L.; Dentchev, T.; Ying, G.S.; Milam, A.H. The role of apoptosis in age-related macular degeneration. Cinical. Sci. 2002, 20, 1435–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J. Clin. Investig. 1998, 102, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G.P.; Sethi, C.S.; Linberg, K.A.; Charteris, D.G.; Fisher, S.K. Experimental retinal reattachment: A new perspective. Mol. Neurobiol. 2003, 8, 159–175. [Google Scholar] [CrossRef]
- Fisher, S.K.; Lewis, G.P.; Linberg, K.A.; Verardo, M.R. Cellular remodeling in mammalian retina: Results from studies of experimental retinal detachment. Prog. Retin. Eye Res. 2005, 24, 395–431. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.G.; Yang, L.; Bula, D.; Chen, D.F. Photoreceptor apoptosis in human retinal detachment. Am. J. Ophthalmol. 2005, 139, 605–610. [Google Scholar] [CrossRef]
- Cook, B.; Lewis, G.P.; Fisher, S.K.; Adler, R. Apoptotic photoreceptor degeneration in experimental retinal detachment. Investig. Ophthalmol. Vis. Sci. 1995, 36, 990–996. [Google Scholar]
- Dong, K.; Zhu, H.; Song, Z.; Gong, Y.; Wang, F.; Wang, W.; Zheng, Z.; Yu, Z.; Gu, Q.; Xu, X.; et al. Necrostatin-1 protects photoreceptors from cell death and improves functional outcome after experimental retinal detachment. Am. J. Pathol. 2012, 181, 1634–1641. [Google Scholar] [CrossRef]
- Lo, A.C.; Woo, T.T.; Wong, R.L.; Wong, D. Apoptosis and other cell death mechanisms after retinal detachment: Implications for photoreceptor rescue. Ophthalmologica 2011, 226 (Suppl. 1), 10–17. [Google Scholar] [CrossRef]
- Murakami, Y.; Notomi, S.; Hisatomi, T.; Nakazawa, T.; Ishibashi, T.; Miller, J.W.; Vavvas, D.G. Photoreceptor cell death and rescue in retinal detachment and degenerations. Prog. Retin. Eye Res. 2013, 37, 114–140. [Google Scholar] [CrossRef] [Green Version]
- Kunchithapautham, K.; Rohrer, B. Autophagy is one of the multiple mechanisms active in photoreceptor degeneration. Autophagy 2007, 3, 65–66. [Google Scholar] [CrossRef] [Green Version]
- Chinskey, N.D.; Zheng, Q.D.; Zacks, D.N. Control of photoreceptor autophagy after retinal detachment: The switch from survival to death. Investig. Ophthalmol. Vis. Sci. 2014, 55, 688–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iandiev, I.; Uckermann, O.; Pannicke, T.; Wurm, A.; Tenckhoff, S.; Pietsch, U.C.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Uhlmann, S. Glial cell reactivity in a porcine model of retinal detachment. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2161–2171. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.P.; Sethi, C.S.; Carter, K.M.; Charteris, D.G.; Fisher, S.K. Microglial cell activation following retinal detachment: A comparison between species. Mol. Vis. 2005, 11, 491–500. [Google Scholar]
- Jeong, E.; Paik, S.S.; Jung, S.W.; Chun, M.H.; Kim, I.B. Morphological and functional evaluation of an animal model for the retinal degeneration induced by N-methyl-N-nitrosourea. Anat. Cell Biol. 2011, 44, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.H.; Kim, H.I.; Paik, S.S.; Jung, S.W.; Kang, S.; Kim, I.B. Functional and morphological evaluation of blue light-emitting diode-induced retinal degeneration in mice. Graefe’s Arch. Clin. Exp. Ophthalmol. 2016, 254, 705–716. [Google Scholar] [CrossRef]
- Chang, S.W.; Kim, H.I.; Kim, G.H.; Park, S.J.; Kim, I.B. Increased Expression of Osteopontin in Retinal Degeneration Induced by Blue Light-Emitting Diode Exposure in Mice. Front. Mol. NeuroSci. 2016, 9, 58. [Google Scholar] [CrossRef]
- Kim, G.H.; Paik, S.S.; Park, Y.S.; Kim, H.G.; Kim, I.B. Amelioration of Mouse Retinal Degeneration After Blue LED Exposure by Glycyrrhizic Acid-Mediated Inhibition of Inflammation. Front. Cell NeuroSci. 2019, 13, 319. [Google Scholar] [CrossRef] [Green Version]
- Paik, S.S.; Jeong, E.; Jung, S.W.; Ha, T.J.; Kang, S.; Sim, S.; Jeon, J.H.; Chun, M.H.; Kim, I.B. Anthocyanins from the seed coat of black soybean reduce retinal degeneration induced by N-methyl-N-nitrosourea. Exp. Eye Res. 2012, 97, 55–62. [Google Scholar] [CrossRef]
- Fisher, S.K.; Lewis, G.P. Müller cell and neuronal remodeling in retinal detachment and reattachment and their potential consequences for visual recovery: A review and reconsideration of recent data. Vis. Res. 2003, 43, 887–897. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.B.; Kim, K.Y.; Joo, C.K.; Lee, M.Y.; Oh, S.J.; Chung, J.W.; Chun, M.H. Reaction of Müller cells after increased intraocular pressure in the rat retina. Exp. Brain Res. 1998, 121, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Ibata, I.; Ito, D.; Ohsawa, K.; Kohsaka, S. A novel gene iba1 in the major histocompatibility complex class III region encoding an EF hand protein expressed in a monocytic lineage. Biochem. Biophys. Res. Commun. 1996, 224, 855–862. [Google Scholar] [CrossRef]
- Hikita, S.T.; Vistica, B.P.; Jones, H.R.; Keswani, J.R.; Watson, M.M.; Ericson, V.R.; Ayoub, G.S.; Gery, I.; Clegg, D.O. Osteopontin is proinflammatory in experimental autoimmune uveitis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4435–4443. [Google Scholar] [CrossRef]
- Kim, M.D.; Cho, H.J.; Shin, T. Expression of osteopontin and its ligand, CD44, in the spinal cords of Lewis rats with experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2004, 151, 78–84. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, H.Y.; Cha, J.H.; Choi, J.Y.; Lee, M.Y. Transient microglial and prolonged astroglial upregulation of osteopontin following transient forebrain ischemia in rats. Brain Res. 2007, 1151, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, M.; Zickler, P.; Denhardt, D.T.; Hartung, H.P.; Jander, S. Increased thalamic neurodegeneration following ischaemic cortical stroke in osteopontin-deficient mice. Brain 2006, 129, 1426–1437. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhong, H.; Li, N.; Chen, K.; Chen, J.; Sun, J.; Xu, L.; Wang, J.; Zhang, M.; Liu, X.; et al. Osteopontin activates retinal microglia causing retinal ganglion cells loss via p38 MAPK signaling pathway in glaucoma. FASEB J. 2021, 35, e21405. [Google Scholar] [CrossRef]
- Heiss, W.D. The ischemic penumbra: Correlates in imaging and implications for treatment of ischemic stroke. The Johann Jacob Wepfer award 2011. Cereb. Dis. 2011, 32, 307–320. [Google Scholar] [CrossRef]
- Angermann, R.; Bechrakis, N.E.; Rauchegger, T.; Casazza, M.; Nowosielski, Y.; Zehetner, C. Effect of Timing on Visual Outcomes in Fovea-Involving Retinal Detachments Verified by SD-OCT. J. Ophthalmol. 2020, 2020, 2307935. [Google Scholar] [CrossRef]
- Kiang, L.; Ross, B.X.; Yao, J.; Shanmugam, S.; Andrews, C.A.; Hansen, S.; Besirli, C.G.; Zacks, D.N.; Abcouwer, S.F. Vitreous Cytokine Expression and a Murine Model Suggest a Key Role of Microglia in the Inflammatory Response to Retinal Detachment. Invest Ophthalmol. Vis. Sci. 2018, 59, 3767–3778. [Google Scholar] [CrossRef]
- Wurm, A.; Pannicke, T.; Iandiev, I.; Bühner, E.; Pietsch, U.C.; Reichenbach, A.; Wiedemann, P.; Uhlmann, S.; Bringmann, A. Changes in membrane conductance play a pathogenic role in osmotic glial cell swelling in detached retinas. Am. J. Pathol. 2006, 169, 1990–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollborn, M.; Francke, M.; Iandiev, I.; Bühner, E.; Foja, C.; Kohen, L.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Uhlmann, S. Early activation of inflammation- and immune response-related genes after experimental detachment of the porcine retina. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1262–1273. [Google Scholar] [CrossRef] [PubMed]
- Eastlake, K.; Banerjee, P.J.; Angbohang, A.; Charteris, D.G.; Khaw, P.T.; Limb, G.A. Müller glia as an important source of cytokines and inflammatory factors present in the gliotic retina during proliferative vitreoretinopathy. Glia 2016, 64, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Langmann, T. Microglia activation in retinal degeneration. J. Leukoc Biol. 2007, 81, 1345–1351. [Google Scholar] [CrossRef]
- Torbidoni, V.; Iribarne, M.; Ogawa, L.; Prasanna, G.; Suburo, A.M. Endothelin-1 and endothelin receptors in light-induced retinal degeneration. Exp. Eye Res. 2005, 81, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.P.; Fisher, S.K. Up-regulation of glial fibrillary acidic protein in response to retinal injury: Its potential role in glial remodeling and a comparison to vimentin expression. Int. Rev. Cytol. 2003, 230, 263–290. [Google Scholar] [CrossRef]
- Luna, G.; Lewis, G.P.; Banna, C.D.; Skalli, O.; Fisher, S.K. Expression profiles of nestin and synemin in reactive astrocytes and Müller cells following retinal injury: A comparison with glial fibrillar acidic protein and vimentin. Mol. Vis. 2010, 16, 2511–2523. [Google Scholar] [PubMed]
- Ma, W.; Zhang, Y.; Gao, C.; Fariss, R.N.; Tam, J.; Wong, W.T. Monocyte infiltration and proliferation reestablish myeloid cell homeostasis in the mouse retina following retinal pigment epithelial cell injury. Sci. Rep. 2017, 7, 8433. [Google Scholar] [CrossRef] [PubMed]
- Aredo, B.; Zhang, K.; Chen, X.; Wang, C.X.; Li, T.; Ufret-Vincenty, R.L. Differences in the distribution, phenotype and gene expression of subretinal microglia/macrophages in C57BL/6N (Crb1 rd8/rd8) versus C57BL6/J (Crb1 wt/wt) mice. J. Neuroinflamm. 2015, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Wirths, O.; Breyhan, H.; Marcello, A.; Cotel, M.C.; Brück, W.; Bayer, T.A. Inflammatory changes are tightly associated with neurodegeneration in the brain and spinal cord of the APP/PS1KI mouse model of Alzheimer’s disease. Neurobiol. Aging 2010, 31, 747–757. [Google Scholar] [CrossRef]
- Rojas, B.; Gallego, B.I.; Ramírez, A.I.; Salazar, J.J.; de Hoz, R.; Valiente-Soriano, F.J.; Avilés-Trigueros, M.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Triviño, A.; et al. Microglia in mouse retina contralateral to experimental glaucoma exhibit multiple signs of activation in all retinal layers. J. Neuroinflamm. 2014, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Ruiz, F.; Galindo-Romero, C.; Rodríguez-Ramírez, K.T.; Vidal-Sanz, M.; Agudo-Barriuso, M. Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells. Int. J. Mol. Sci. 2019, 20, 5733. [Google Scholar] [CrossRef] [Green Version]
- Jarrett, W.H., 2nd. Rhematogenous retinal detachment complicated by severe intraocular inflammation, hypotony, and choroidal detachment. Trans. Am. Ophthalmol. Soc. 1981, 79, 664–683. [Google Scholar] [PubMed]
- Shah, D.N.; Al-Moujahed, A.; Newcomb, C.W.; Kaçmaz, R.O.; Daniel, E.; Thorne, J.E.; Foster, C.S.; Jabs, D.A.; Levy-Clarke, G.A.; Nussenblatt, R.B.; et al. Exudative Retinal Detachment in Ocular Inflammatory Diseases: Risk and Predictive Factors. Am. J. Ophthalmol. 2020, 218, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Li, Y.; Wu, Y.; Qu, Y. Surgical complications of primary rhegmatogenous retinal detachment: A meta-analysis. PLoS ONE 2015, 10, e0116493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, J.A.; Velier, J.J.; Spera, P.; Jonak, Z.L.; Wang, X.; Barone, F.C.; Feuerstein, G.Z. Osteopontin and its integrin receptor alpha(v)beta3 are upregulated during formation of the glial scar after focal stroke. Stroke 1998, 29, 1698–1706; discussion 1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedtjärn, M.; Mallard, C.; Hagberg, H. Inflammatory gene profiling in the developing mouse brain after hypoxia-ischemia. J. Cereb. Blood Flow Metab. 2004, 24, 1333–1351. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Park, Y.-S.; Paik, S.-S.; Kim, I.-B. Differential Response of Müller Cells and Microglia in a Mouse Retinal Detachment Model and Its Implications in Detached and Non-Detached Regions. Cells 2021, 10, 1972. https://doi.org/10.3390/cells10081972
Lee S-H, Park Y-S, Paik S-S, Kim I-B. Differential Response of Müller Cells and Microglia in a Mouse Retinal Detachment Model and Its Implications in Detached and Non-Detached Regions. Cells. 2021; 10(8):1972. https://doi.org/10.3390/cells10081972
Chicago/Turabian StyleLee, Seung-Hee, Yong-Soo Park, Sun-Sook Paik, and In-Beom Kim. 2021. "Differential Response of Müller Cells and Microglia in a Mouse Retinal Detachment Model and Its Implications in Detached and Non-Detached Regions" Cells 10, no. 8: 1972. https://doi.org/10.3390/cells10081972