
Berberine Improves Cognitive Impairment by Simultaneously Impacting Cerebral Blood Flow and β-Amyloid Accumulation in an APP/tau/PS1 Mouse Model of Alzheimer’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Reagents
2.2. Animals and Treatment
2.3. Magnetic Resonance Imaging
2.3.1. Scanning Parameters
2.3.2. 3D Arterial Spin Labeling Technique
2.4. Morris Water Maze Task
2.4.1. Spatial Learning Test
2.4.2. Probe Trial
2.5. Immunofluorescence Staining
2.6. TUNEL Assay
2.7. Laser Speckle Contrast Imaging
2.8. Western Blot Assay
2.9. Statistical Analysis
3. Results
3.1. BBR Accelerates the Recovery of CBF in the Brain of 3×Tg AD Mice
3.2. BBR Relieves Cognitive Impairments in 3×Tg AD Mice
3.3. BBR Reduces the Production of Aβ and Inhibits Apoptosis in the Brains of 3×Tg AD Mice
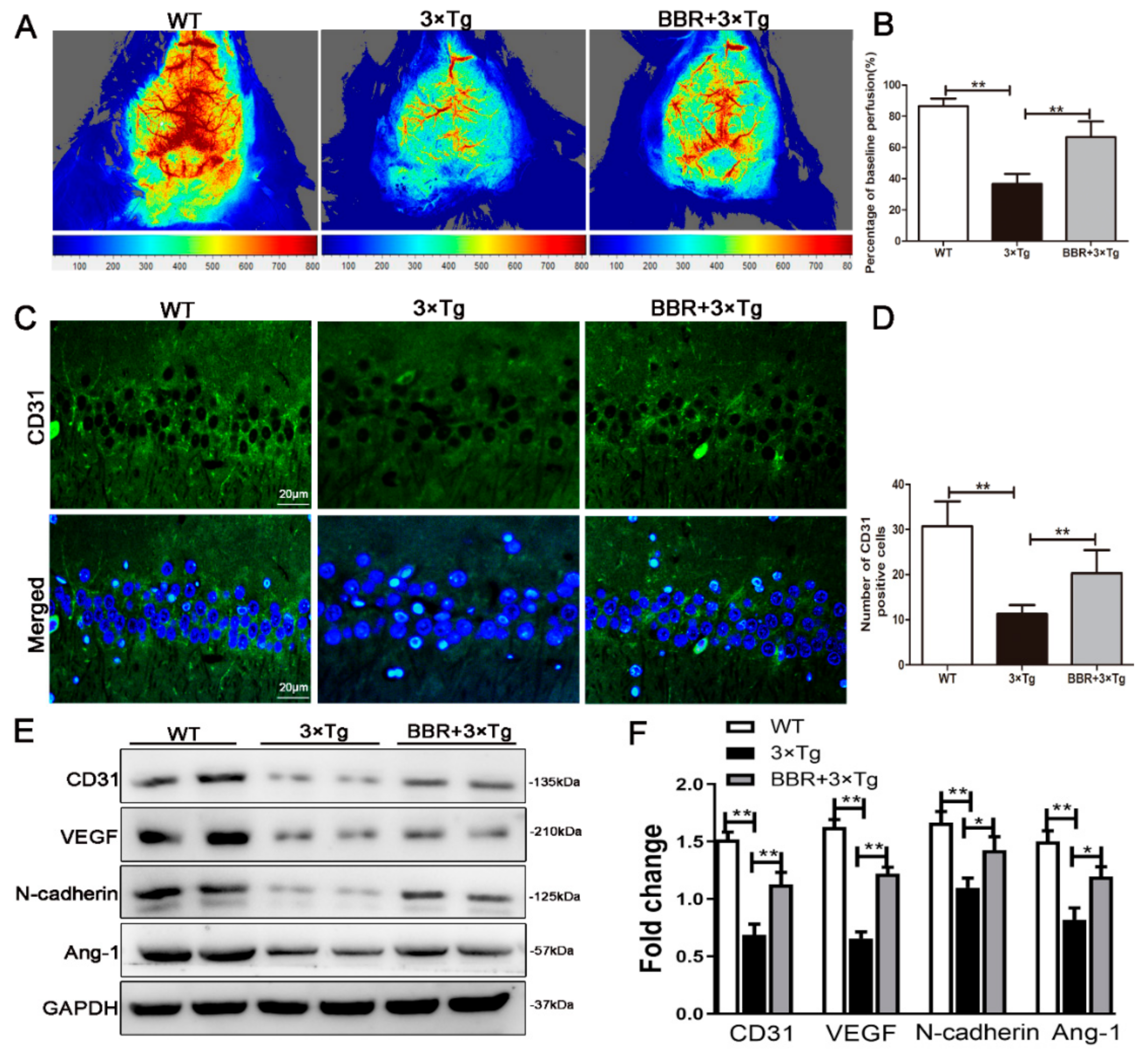
3.4. BBR Increases the Cerebral CBF Perfusion and Proangiogenic Factors in the Brains of 3×Tg AD Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hampel, H.; Vergallo, A.; Afshar, M.; Akman-Anderson, L.; Arenas, J.; Benda, N.; Batrla, R.; Broich, K.; Caraci, F.; Cuello, A.C.; et al. Blood-based systems biology biomarkers for next-generation clinical trials in Alzheimer’s disease. Dialogues Clin. Neurosci. 2019, 21, 177–191. [Google Scholar] [PubMed]
- Adav, S.S.; Sze, S.K. Hypoxia-Induced Degenerative Protein Modifications Associated with Aging and Age-Associated Disorders. Aging Dis. 2020, 11, 341–364. [Google Scholar] [CrossRef] [Green Version]
- Kalaria, R.N. Neuropathological diagnosis of vascular cognitive impairment and vascular dementia with implications for Alzheimer’s disease. Acta Neuropathol. 2016, 131, 659–685. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, M.; Anusuyadevi, M.; Aigner, K.M.; Unger, M.S.; Kniewallner, K.M.; de Sousa, D.M.B.; Altendorfer, B.; Mrowetz, H.; Bogdahn, U.; Aigner, L. TGF-β Signaling: A Therapeutic Target to Reinstate Regenerative Plasticity in Vascular Dementia? Aging Dis. 2020, 11, 828–850. [Google Scholar] [CrossRef] [PubMed]
- Archer, H.A.; Smailagic, N.; John, C.; Holmes, R.B.; Takwoingi, Y.; Coulthard, E.J.; Cullum, S. Regional cerebral blood flow single photon emission computed tomography for detection of Frontotemporal dementia in people with suspected dementia. Cochrane Database Syst. Rev. 2015, 23, CD010896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, C.H.; Wang, Y.; Fung, M.-L.; Zhang, C.; Lim, L.W. Rodent Models of Amyloid-Beta Feature of Alzheimer’s Disease: Development and Potential Treatment Implications. Aging Dis. 2020, 11, 1235–1259. [Google Scholar] [CrossRef]
- Bailey, T.L.; Rivara, C.B.; Rocher, A.B.; Hof, P.R. The nature and effects of cortical microvascular pathology in aging and Alzheimer’s disease. Neurol. Res. 2004, 26, 573–578. [Google Scholar] [CrossRef]
- Lorius, N.; Locascio, J.J.; Rentz, D.M.; Johnson, K.A.; Sperling, R.A.; Viswanathan, A.; Marshall, G.A.; Alzheimer’s Disease Neuroimaging Initiative. Vascular disease and risk factors are associated with cognitive decline in the alzheimer disease spectrum. Alzheimer Dis. Assoc. Disord. 2015, 29, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Prohovnik, I.; Mayeux, R.; Sackeim, H.A.; Smith, G.; Stern, Y.; Alderson, P.O. Cerebral perfusion as a diagnostic marker of early Alzheimer’s disease. Neurology 1988, 38, 931–937. [Google Scholar] [CrossRef]
- Pluta, R.; Furmaga-Jabłońska, W.; Maciejewski, R.; Ułamek-Kozioł, M.; Jabłoński, M. Brain ischemia activates β- and γ-secretase cleavage of amyloid precursor protein: Significancein sporadic Alzheimer’s disease. Mol. Neurobiol. 2013, 47, 425–434. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Perkins, A.; Cisternas, P.; Muñoz, B.; Taylor, X.; You, Y.; Garringer, H.J.; Oblak, A.L.; Atwood, B.K.; Vidal, R.; et al. Tau as a mediator of neurotoxicity associated to cerebral amyloid angiopathy. Acta Neuropathol. Commun. 2019, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, K. A Microcirculatory Theory of Aging. Aging Dis. 2019, 10, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Ochalek, A.; Mihalik, B.; Avci, H.X.; Chandrasekaran, A.; Téglási, A.; Bock, I.; Giudice, M.L.; Táncos, Z.; Molnár, K.; László, L.; et al. Neurons derived from sporadic Alzheimer’s disease iPSCs reveal elevated TAU hyperphosphorylation, increased amyloid levels, and GSK3B activation. Alzheimers Res. Ther. 2017, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Barrera-Ocampo, A.; Lopera, F. Amyloid-beta immunotherapy: The hope for Alzheimer disease? Colomb. Med. 2016, 47, 203–212. [Google Scholar] [CrossRef]
- Kulkarni, S.K.; Dhir, A. Berberine: A plant alkaloid with therapeutic potential for central nervous system disorders. Phytother. Res. 2010, 24, 317–324. [Google Scholar] [CrossRef]
- Huang, M.; Chen, S.; Liang, Y.; Guo, Y. The role of berberine in the multi-target treatment of senile dementia. Curr. Top. Med. Chem. 2016, 16, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; et al. Triple-transgenic model of Alzheimer’s disease with plaques and tangles: Intracellular Abeta and synaptic dysfunction. Neuron 2003, 39, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef]
- Jing, Z.; Shi, C.; Zhu, L.; Xiang, Y.; Chen, P.; Xiong, Z.; Li, W.; Ruan, Y.; Huang, L. Chroniccerebral hypoperfusion induces vascular plasticity and hemodynamics but also neuronal degeneration and cognitive impairment. J. Cereb. Blood Flow Metab. 2015, 35, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.C.; Temple, V.; Oh, E.; VanRyzin, C.; Williams, A.; Cornwell, B.; Grillon, C.; Pine, D.S.; Ernst, M.; Merke, D.P. Early and rogen exposure modulates spatial cognition in congenital adrenal hyperplasia (CAH). Psychoneuroendocrinology 2008, 33, 973–980. [Google Scholar] [CrossRef] [Green Version]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, J.C. Is Alzheimer’s disease a neurodegenerative or a vascular disorder? Data, dogma, and dialectics. Lancet Neurol. 2004, 3, 184–190. [Google Scholar] [CrossRef]
- Ni, Y.-Q.; Lin, X.; Zhan, J.-K.; Liu, Y.-S. Roles and Functions of Exosomal Non-coding RNAs in Vascular Aging. Aging Dis. 2020, 11, 164–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogure, D.; Matsuda, H.; Ohnishi, T.; Asada, T.; Uno, M.; Kunihiro, T.; Nakano, S.; Takasaki, M. Longitudinal evaluation of early Alzheimer’s disease using brain perfusion SPECT. J. Nucl. Med. 2000, 41, 1155–1162. [Google Scholar] [PubMed]
- Rodriguez, G.; Vitali, P.; Calvini, P.; Bordoni, C.; Girtler, N.; Taddei, G.; Mariani, G.; Nobili, F. Hippocampal perfusion in mild Alzheimer’s disease. Psychiatry Res. 2000, 100, 65–74. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Petersen, R.C.; Xu, Y.C.; O’Brien, P.C.; Smith, G.E.; Ivnik, R.J.; Boeve, B.F.; Waring, S.C.; Tangalos, E.G.; Kokmen, E. Prediction of AD with MRI-based hippocampal volume in mild cognitive impairment. Neurology 1999, 52, 1397–1403. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Liu, Z.; Xiao, M.; Wang, C.; Tian, F. Chronic Cerebral Hypoperfusion Promotes Amyloid-Beta Pathogenesis via Activating β/γ-Secretases. Neurochem. Res. 2017, 42, 3446–3455. [Google Scholar] [CrossRef]
- Aski, M.L.; Rezvani, M.E.; Khaksari, M.; Hafizi, Z.; Pirmoradi, Z.; Niknazar, S.; Mehrjerdi, F.Z. Neuroprotective effect of berberine chloride on cognitive impairment and hippocampal damage in experimental model of vascular dementia. Iran J. Basic Med. Sci. 2018, 21, 53–58. [Google Scholar]
- Jia, Y.; Wang, N.; Zhang, Y.; Xue, D.; Lou, H.; Liu, X. Alteration in the Function and Expression of SLC and ABC Transporters in the Neurovascular Unit in Alzheimer’s Disease and the Clinical Significance. Aging Dis. 2020, 11, 390–404. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Kui, L.; Demetrios, T.; Gong, X.; Tang, M. A Glimmer of Hope: Maintain Mitochondrial Homeostasis to Mitigate Alzheimer’s Disease. Aging Dis. 2020, 11, 1260–1275. [Google Scholar] [CrossRef]
- Do, T.M.; Alata, W.; Dodacki, A.; Traversy, M.-T.; Chacun, H.; Pradier, L.; Scherrmann, J.-M.; Farinotti, R.; Calon, F.; Bourasset, F. Altered cerebral vascular volumes and solute transport at the blood-brain barriers of two transgenic mouse models of Alzheimer’s disease. Neuropharmacology 2014, 81, 311–317. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. Hypoxia/ischemia activate processing of Amyloid Precursor Protein: Impact of vascular dysfunction in the pathogenesis of Alzheimer’s disease. J. Neurochem. 2017, 140, 536–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, E.E.; Doetsch, F. FACS isolation of endothelial cells and pericytes from mouse brain microregions. Nat. Protoc. 2018, 13, 738–751. [Google Scholar] [CrossRef] [PubMed]
- Solerte, S.B.; Ferrari, E.; Cuzzoni, G.; Locatelli, E.; Giustina, A.; Zamboni, M.; Schifino, N.; Rondanelli, M.; Gazzaruso, C.; Fioravanti, M. Decreased release of the angiogenic peptide vascular endothelial growth factor in Alzheimer’sdisease: Recovering effect with insulin and DHEA sulfate. Dement. Geriatr. Cogn. Disord. 2005, 19, 1–10. [Google Scholar] [CrossRef]
- Chakraborty, A.; Chatterjee, M.; Twaalfhoven, H.; Del Campo Milan, M.; Teunissen, C.E.; Scheltens, P.; Fontijn, R.D.; van Der Flier, W.M.; de Vries, H.E. Vascular Endothelial Growth Factor remains unchanged in cerebrospinal fluid of patients with Alzheimer’s disease and vascular dementia. Alzheimers Res. Ther. 2018, 10, 58. [Google Scholar] [CrossRef]
- Hozzein, W.N.; Badr, G.; Badr, B.M.; Allam, A.; Ghamdi, A.A.; Al-Wadaan, M.A.; Al-Waili, N.S. Bee venom improves diabetic wound healing by protecting functional macrophages from apoptosis and enhancing Nrf2, Ang-1 and Tie-2 signaling. Mol. Immunol. 2018, 103, 322–335. [Google Scholar] [CrossRef]
- Bitto, A.; Minutoli, L.; Galeano, M.R.; Altavilla, D.; Polito, F.; Fiumara, T.; Calò, M.; Lo Cascio, P.; Zentilin, L.; Giacca, M.; et al. Angiopoietin-1 gene transfer improves impaired wound healing in genetically diabetic micewithout increasing VEGF expression. Clin. Sci. 2008, 114, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Tillet, E.; Vittet, D.; Féraud, O.; Moore, R.; Kemler, R.; Huber, P. N-cadherin deficiency impairs pericyte recruitment, and not endothelial differentiation or sprouting, in embryonic stem cell-derived angiogenesis. Exp. Cell Res. 2005, 310, 392–400. [Google Scholar] [CrossRef]
- Lelièvre, E.C.; Plestant, C.; Boscher, C.; Wolff, E.; Mège, R.M.; Birbes, H. N-cadherin mediates neuronal cell survival through Bim down-regulation. PLoS ONE 2012, 7, e33206. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Application | Source | Dilutions |
|---|---|---|---|---|
| Aβ | Rabbit | WB/IF | Abcam | 1:500 |
| GFAP | Rabbit | WB/IF | Abcam | 1:500 |
| NeuN | Rabbit | WB/IF | Abcam | 1:500 |
| CD31 | Rabbit | WB/IF | Abcam | 1:500 |
| VEGF | Rabbit | WB | Abcam | 1:500 |
| GAPDH | Rabbit | WB | Abcam | 1:2500 |
| N-cadherin | Rabbit | WB | Cell signaling | 1:1000 |
| Ang-1 | Rabbit | WB | Cell signaling | 1:1000 |
| caspase3 | Rabbit | WB/IF | Cell signaling | 1:1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, C.; Liang, Y.; Chen, Y.; Xiong, Y.; She, Y.; Zhong, X.; Chen, H.; Huang, M. Berberine Improves Cognitive Impairment by Simultaneously Impacting Cerebral Blood Flow and β-Amyloid Accumulation in an APP/tau/PS1 Mouse Model of Alzheimer’s Disease. Cells 2021, 10, 1161. https://doi.org/10.3390/cells10051161
Ye C, Liang Y, Chen Y, Xiong Y, She Y, Zhong X, Chen H, Huang M. Berberine Improves Cognitive Impairment by Simultaneously Impacting Cerebral Blood Flow and β-Amyloid Accumulation in an APP/tau/PS1 Mouse Model of Alzheimer’s Disease. Cells. 2021; 10(5):1161. https://doi.org/10.3390/cells10051161
Chicago/Turabian StyleYe, Chenghui, Yubin Liang, Ying Chen, Yu Xiong, Yingfang She, Xiaochun Zhong, Hongda Chen, and Min Huang. 2021. "Berberine Improves Cognitive Impairment by Simultaneously Impacting Cerebral Blood Flow and β-Amyloid Accumulation in an APP/tau/PS1 Mouse Model of Alzheimer’s Disease" Cells 10, no. 5: 1161. https://doi.org/10.3390/cells10051161