
Abdominal Obesity-Related Disturbance of Insulin Sensitivity Is Associated with CD8+ EMRA Cells in the Elderly

 , , , , , and
, , , , , and
Abstract
:1. Introduction
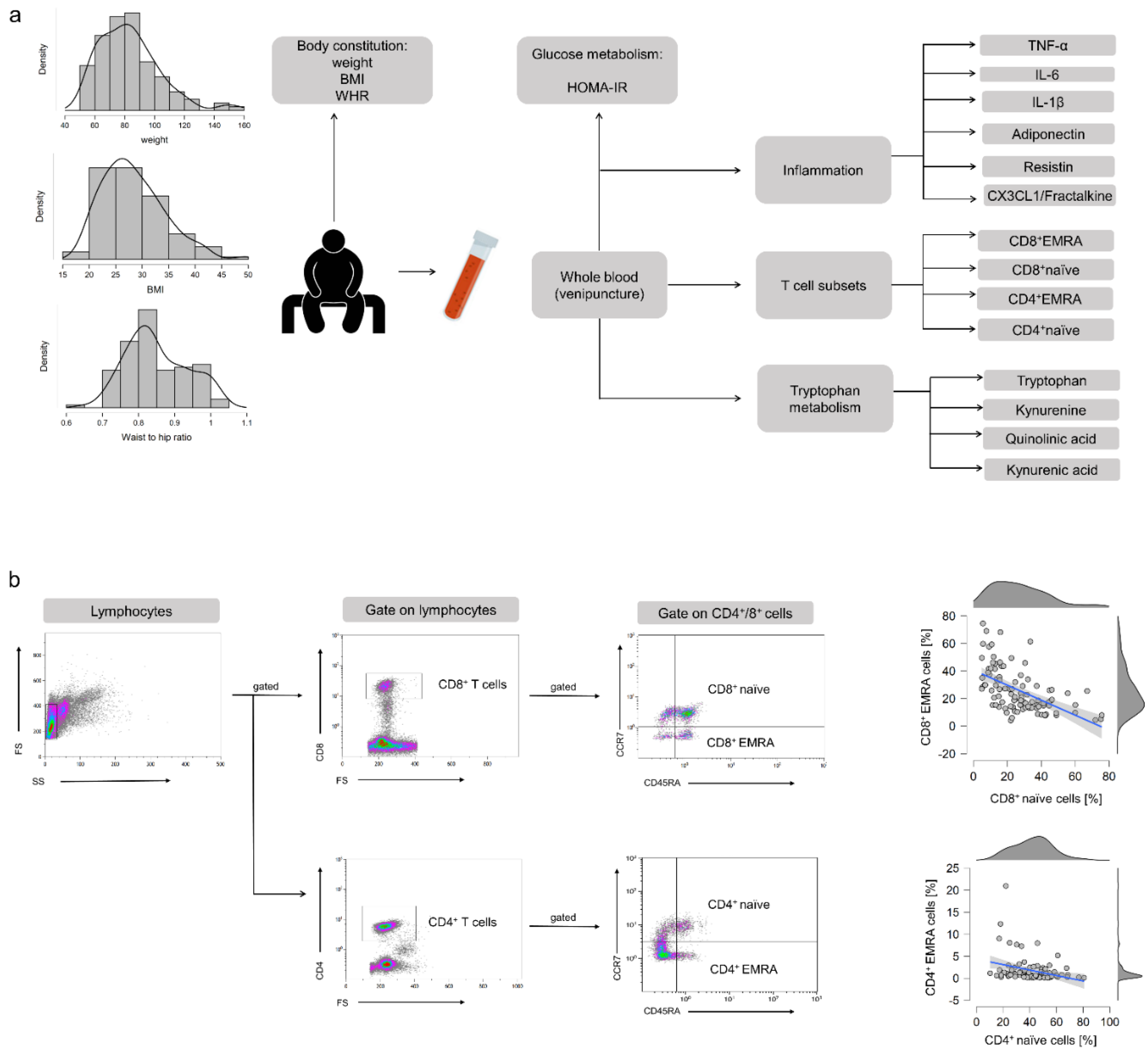
2. Materials and Methods
2.1. Study Participants
2.2. Body Weight and Body Composition
2.3. Blood Sampling
2.4. Analysis of Glucose Metabolism and Insulin Resistance
2.5. Analysis of T Cell Subpopulations
2.6. Analysis of Tryptophan Metabolites
2.7. Analysis of Plasma Cytokines
2.8. Analysis of Cytomegalovirus (CMV) Serostatus
2.9. Statistical Analysis
3. Results
3.1. Baseline Characteristics
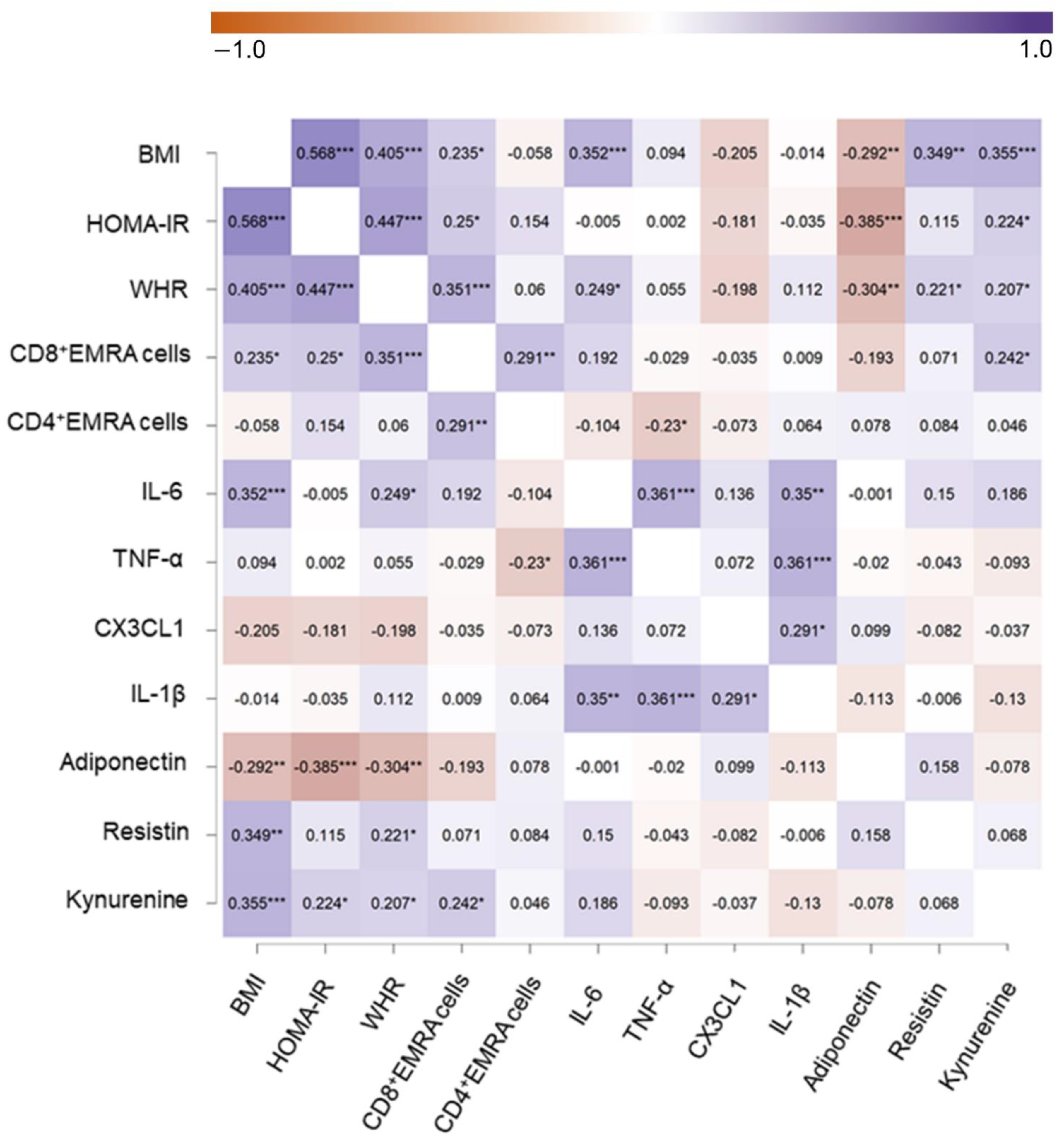
3.2. Associations between Body Composition, Glucose Metabolism, T-EMRA Cells, Trp Metabolites, and Cytokine Status
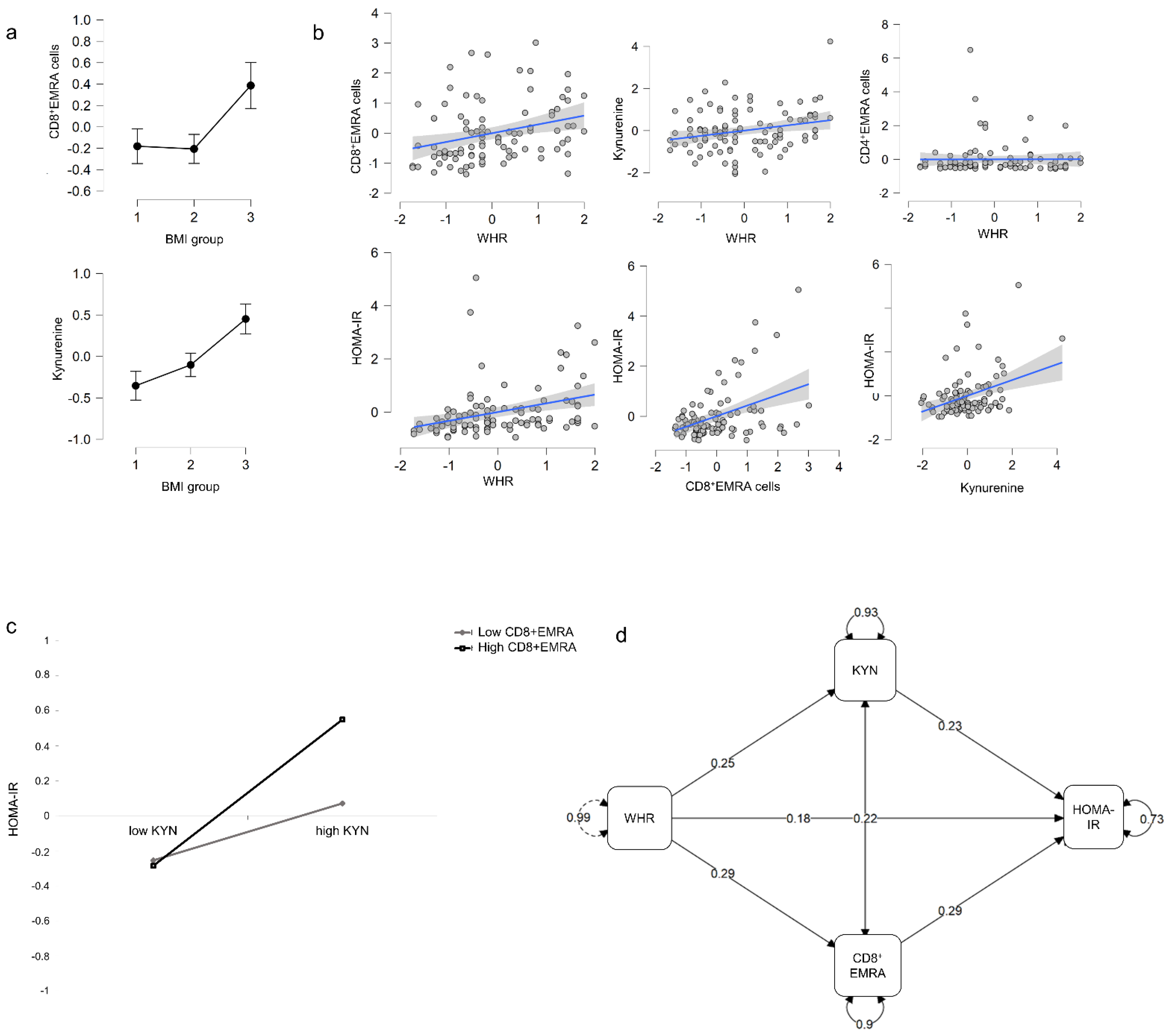
3.3. Effect of BMI on T-EMRA Cells and Trp Metabolism
3.4. Moderation of the Glucose Metabolism by Abdominal Obesity, the Proportion of T-EMRA Cells, and Trp Metabolites
3.5. Abdominal Obesity Affects T-EMRA Cells and Trp Metabolites, Which in Turn Influences HOMA-IR
4. Discussion
4.1. The Role of T-EMRA Cells in Obesity-Related Disturbance of Glucose Homeostasis
4.2. KYN as a Mediator between Abdominal Fat and a Disturbed Glucose Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Makinodan, T. Studies on the influence of age on immune response to understand the biology of immunosenescence. Exp. Gerontol. 1998, 33, 27–38. [Google Scholar] [CrossRef]
- Goronzy, J.J.; Fang, F.; Cavanagh, M.M.; Qi, Q.; Weyand, C.M. Naive T cell maintenance and function in human aging. J. Immunol. 2015, 194, 4073–4080. [Google Scholar] [CrossRef]
- Jacomet, F.; Cayssials, E.; Basbous, S.; Levescot, A.; Piccirilli, N.; Desmier, D.; Robin, A.; Barra, A.; Giraud, C.; Guilhot, F.; et al. Evidence for eomesodermin-expressing innate-like CD8(+) KIR/NKG2A(+) T cells in human adults and cord blood samples. Eur. J. Immunol. 2015, 45, 1926–1933. [Google Scholar] [CrossRef]
- Goronzy, J.J.; Weyand, C.M. Successful and Maladaptive T Cell Aging. Immunity 2017, 46, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wertheimer, A.M.; Bennett, M.S.; Park, B.; Uhrlaub, J.L.; Martinez, C.; Pulko, V.; Currier, N.L.; Nikolich-Žugich, D.; Kaye, J.; Nikolich-Žugich, J. Aging and cytomegalovirus infection differentially and jointly affect distinct circulating T cell subsets in humans. J. Immunol. 2014, 192, 2143–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, K.; Ogonek, J.; Varanasi, P.R.; Luther, S.; Bünting, I.; Thomay, K.; Behrens, Y.L.; Mischak-Weissinger, E.; Hambach, L. Human CD8+ CD57- TEMRA cells: Too young to be called “old”. PLoS ONE 2017, 12, e0177405. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Karandikar, N.J.; Betts, M.R.; Ambrozak, D.R.; Hill, B.J.; Crotty, L.E.; Casazza, J.P.; Kuruppu, J.; Migueles, S.A.; Connors, M.; et al. Expression of CD57 defines replicative senescence and antigen-induced apoptotic death of CD8+ T cells. Blood 2003, 101, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Larbi, A.; Derhovanessian, E.; Ozcelik, D.; Naumova, E.; Pawelec, G. Multiparameter flow cytometric analysis of CD4 and CD8 T cell subsets in young and old people. Immun. Ageing 2008, 25, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, T.; Schulte, S.; Warrington, K.J.; Kopecky, S.L.; Frye, R.L.; Goronzy, J.J.; Weyand, C.M. T-cell-mediated lysis of endothelial cells in acute coronary syndromes. Circulation 2002, 105, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Tilly, G.; Doan-Ngoc, T.M.; Yap, M.; Caristan, A.; Jacquemont, L.; Danger, R.; Cadoux, M.; Bruneau, S.; Giral, M.; Guerif, P.; et al. IL-15 Harnesses Pro-inflammatory Function of TEMRA CD8 in Kidney-Transplant Recipients. Front. Immunol. 2017, 8, 778. [Google Scholar] [CrossRef] [Green Version]
- Spyridopoulos, I.; Martin-Ruiz, C.; Hilkens, C.; Yadegarfar, M.E.; Isaacs, J.; Jagger, C.; Kirkwood, T.; von Zglinicki, T. CMV seropositivity and T-cell senescence predict increased cardiovascular mortality in octogenarians: Results from the Newcastle 85+ study. Aging Cell 2016, 15, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Yap, M.; Boeffard, F.; Clave, E.; Pallier, A.; Danger, R.; Giral, M.; Dantal, J.; Foucher, Y.; Guillot-Gueguen, C.; Toubert, A.; et al. Expansion of highly differentiated cytotoxic terminally differentiated effector memory CD8+ T cells in a subset of clinically stable kidney transplant recipients: A potential marker for late graft dysfunction. J. Am. Soc. Nephrol. 2014, 25, 1856–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boren, E.; Gershwin, M.E. Inflamm-aging: Autoimmunity, and the immune-risk phenotype. Autoimmun. Rev. 2004, 3, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C. Inflammaging as a major characteristic of old people: Can it be prevented or cured? Nutr. Rev. 2007, 65, 173–176. [Google Scholar] [CrossRef]
- Broadley, I.; Pera, A.; Morrow, G.; Davies, K.A.; Kern, F. Expansions of Cytotoxic CD4+CD28- T Cells Drive Excess Cardiovascular Mortality in Rheumatoid Arthritis and Other Chronic Inflammatory Conditions and Are Triggered by CMV Infection. Front. Immunol. 2017, 8, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikby, A.; Månsson, I.A.; Johansson, B.; Strindhall, J.; Nilsson, S.E. The immune risk profile is associated with age and gender: Findings from three Swedish population studies of individuals 20–100 years of age. Biogerontology 2008, 9, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Bardini, G.; Dicembrini, I.; Cresci, B.; Rotella, C.M. Inflammation markers and metabolic characteristics of subjects with 1-h plasma glucose levels. Diabetes Care 2010, 33, 411–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgartner, R.; Forteza, M.J.; Ketelhuth, D.F.J. The interplay between cytokines and the Kynurenine pathway in inflammation and atherosclerosis. Cytokine 2019, 122, 154148. [Google Scholar] [CrossRef]
- Song, P.; Ramprasath, T.; Wang, H.; Zou, M.H. Abnormal kynurenine pathway of tryptophan catabolism in cardiovascular diseases. Cell Mol. Life Sci. 2017, 74, 2899–2916. [Google Scholar] [CrossRef] [PubMed]
- Joisten, N.; Kummerhoff, F.; Koliamitra, C.; Schenk, A.; Walzik, D.; Hardt, L.; Knoop, A.; Thevis, M.; Kiesl, D.; Metcalfe, A.J.; et al. Exercise and the Kynurenine pathway: Current state of knowledge and results from a randomized cross-over study comparing acute effects of endurance and resistance training. Exerc. Immunol. Rev. 2020, 26, 24–42. [Google Scholar]
- Wasserfurth, P.; Nebl, J.; Boßlau, T.K.; Krüger, K.; Hahn, A.; Schuchardt, J.P. Intake of Calanus finmarchicus oil for 12 weeks improves omega-3 index in healthy older subjects engaging in an exercise programme. Br. J. Nutr. 2020, 125, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Noble, R.E. Waist-to-hip ratio versus BMI as predictors of cardiac risk in obese adult women. West. J. Med. 2001, 174, 240–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, A.S. Insulin resistance with aging: Effects of diet and exercise. Sports Med. 2000, 30, 327–346. [Google Scholar] [CrossRef] [PubMed]
- Parel, Y.; Chizzolini, C. CD4+ CD8+ double positive (DP) T cells in health and disease. Autoimmun. Rev. 2004, 3, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.D.; Sivarapatna, A.; Gress, R.E. The aging immune system and its relationship with cancer. Aging Health 2011, 7, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Duggal, N.A. Reversing the immune ageing clock: Lifestyle modifications and pharmacological interventions. Biogerontology 2018, 19, 481–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, E.G.; Murphy, W.J. Obesity induced T cell dysfunction and implications for cancer immunotherapy. Curr. Opin. Immunol. 2018, 51, 181–186. [Google Scholar] [CrossRef]
- Yang, H.; Youm, Y.H.; Vandanmagsar, B.; Rood, J.; Kumar, K.G.; Butler, A.A.; Dixit, V.D. Obesity accelerates thymic aging. Blood 2009, 114, 3803–3812. [Google Scholar] [CrossRef] [Green Version]
- Shirakawa, K.; Yan, X.; Shinmura, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Yamamoto, T.; Anzai, A.; Isobe, S.; Yoshida, N.; et al. Obesity accelerates T cell senescence in murine visceral adipose tissue. J. Clin. Investig. 2016, 126, 4626–4639. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.S.; Kim, S.Y.; Kim, J.T.; Lee, Y.S.; Moon, J.S.; Kim, M.; Kang, Y.E.; Joung, K.H.; Lee, J.H.; Kim, H.J.; et al. T-cell senescence contributes to abnormal glucose homeostasis in humans and mice. Cell Death Dis. 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, A.K.; Xu, M.; Zhu, Y.; Pirtskhalava, T.; Weivoda, M.M.; Hachfeld, C.M.; Prata, L.G.; van Dijk, T.H.; Verkade, E.; Casaclang-Verzosa, G.; et al. Targeting senescent cells alleviates obesity-induced metabolic dysfunction. Aging Cell 2019, 18, e12950. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Vazquez, I.; Fernández-Veledo, S.; Krämer, D.K.; Vila-Bedmar, R.; Garcia-Guerra, L.; Lorenzo, M. Insulin resistance associated to obesity: The link TNF-alpha. Arch. Physiol. Biochem. 2008, 114, 183–194, Erratum in 2009, 115, 117. [Google Scholar] [CrossRef]
- Kim, J.H.; Bachmann, R.A.; Chen, J. Interleukin-6 and insulin resistance. Vitam. Horm. 2009, 80, 613–633. [Google Scholar] [CrossRef]
- Dienz, O.; Rincon, M. The effects of IL-6 on CD4 T cell responses. Clin. Immunol. 2009, 130, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Raynal, S.; Bailbé, D.; Gausseres, B.; Carbonne, C.; Autier, V.; Movassat, J.; Kergoat, M.; Portha, B. Expression of the kynurenine pathway enzymes in the pancreatic islet cells. Activation by cytokines and glucolipotoxicity. Biochim. Biophys. Acta 2015, 1852, 980–991. [Google Scholar] [CrossRef] [Green Version]
- Wolowczuk, I.; Hennart, B.; Leloire, A.; Bessede, A.; Soichot, M.; Taront, S.; Caiazzo, R.; Raverdy, V.; Pigeyre, M.; ABOS Consortium; et al. Tryptophan metabolism activation by indoleamine 2,3-dioxygenase in adipose tissue of obese women: An attempt to maintain immune homeostasis and vascular tone. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batatinha, H.A.P.; Rosa Neto, J.C.; Krüger, K. Inflammatory features of obesity and smoke exposure and the immunologic effects of exercise. Exerc. Immunol. Rev. 2019, 25, 96–111. [Google Scholar] [PubMed]
- Oxenkrug, G. Insulin resistance and dysregulation of tryptophan-kynurenine and kynurenine-nicotinamide adenine dinucleotide metabolic pathways. Mol. Neurobiol. 2013, 48, 294–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, A.; Morley, J.E. Obesity in the Elderly. Available online: https://www.ncbi.nlm.nih.gov/books/NBK532533/ (accessed on 11 December 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | |
|---|---|
| Sex [f/m] | [96/38] |
| Age [years] | 59.2 ± 5.6 |
| Height [m] | 170.9 ± 8.7 |
| Body weight [kg] | 83.0 ± 20.3 |
| BMI [kg/m2] | 28.3 ± 5.8 |
| Waist circumference [cm] | 93.30 ± 14.53 |
| Hip circumference [cm] | 108.32 ± 12.15 |
| WHR | 0.85 ± 0.09 |
| Fasting Glucose [mg/dL] | 93.13 ± 16.78 |
| HbA1c [%] | 5.44 ± 0.45 |
| Insulin [µU/mL] | 11.39 ± 7.96 |
| HOMA-Index | 2.72 ± 2.26 |
| CMV positive [Yes/no] | [72/63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boßlau, T.K.; Wasserfurth, P.; Krüger, B.; Reichel, T.; Palmowski, J.; Nebl, J.; Weyh, C.; Schenk, A.; Joisten, N.; Stahl, F.; et al. Abdominal Obesity-Related Disturbance of Insulin Sensitivity Is Associated with CD8+ EMRA Cells in the Elderly. Cells 2021, 10, 998. https://doi.org/10.3390/cells10050998
Boßlau TK, Wasserfurth P, Krüger B, Reichel T, Palmowski J, Nebl J, Weyh C, Schenk A, Joisten N, Stahl F, et al. Abdominal Obesity-Related Disturbance of Insulin Sensitivity Is Associated with CD8+ EMRA Cells in the Elderly. Cells. 2021; 10(5):998. https://doi.org/10.3390/cells10050998
Chicago/Turabian StyleBoßlau, Tim K., Paulina Wasserfurth, Britta Krüger, Thomas Reichel, Jana Palmowski, Josefine Nebl, Christopher Weyh, Alexander Schenk, Niklas Joisten, Frank Stahl, and et al. 2021. "Abdominal Obesity-Related Disturbance of Insulin Sensitivity Is Associated with CD8+ EMRA Cells in the Elderly" Cells 10, no. 5: 998. https://doi.org/10.3390/cells10050998