
PD-1/PD-L1 Checkpoint Inhibitors Are Active in the Chicken Embryo Model and Show Antitumor Efficacy In Ovo


 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tumor Cell Culture
2.2. Immunophenotyping on Chicken Peripheral Blood Mononuclear Cells (PBMCs)
2.3. Immunohistochemistry (IHC) Detection of Tumor-Infiltrated Lymphocytes
2.4. Binding of Pembrolizumab to Chicken Immune Cells
2.5. In Vitro Pembrolizumab Functionality Test
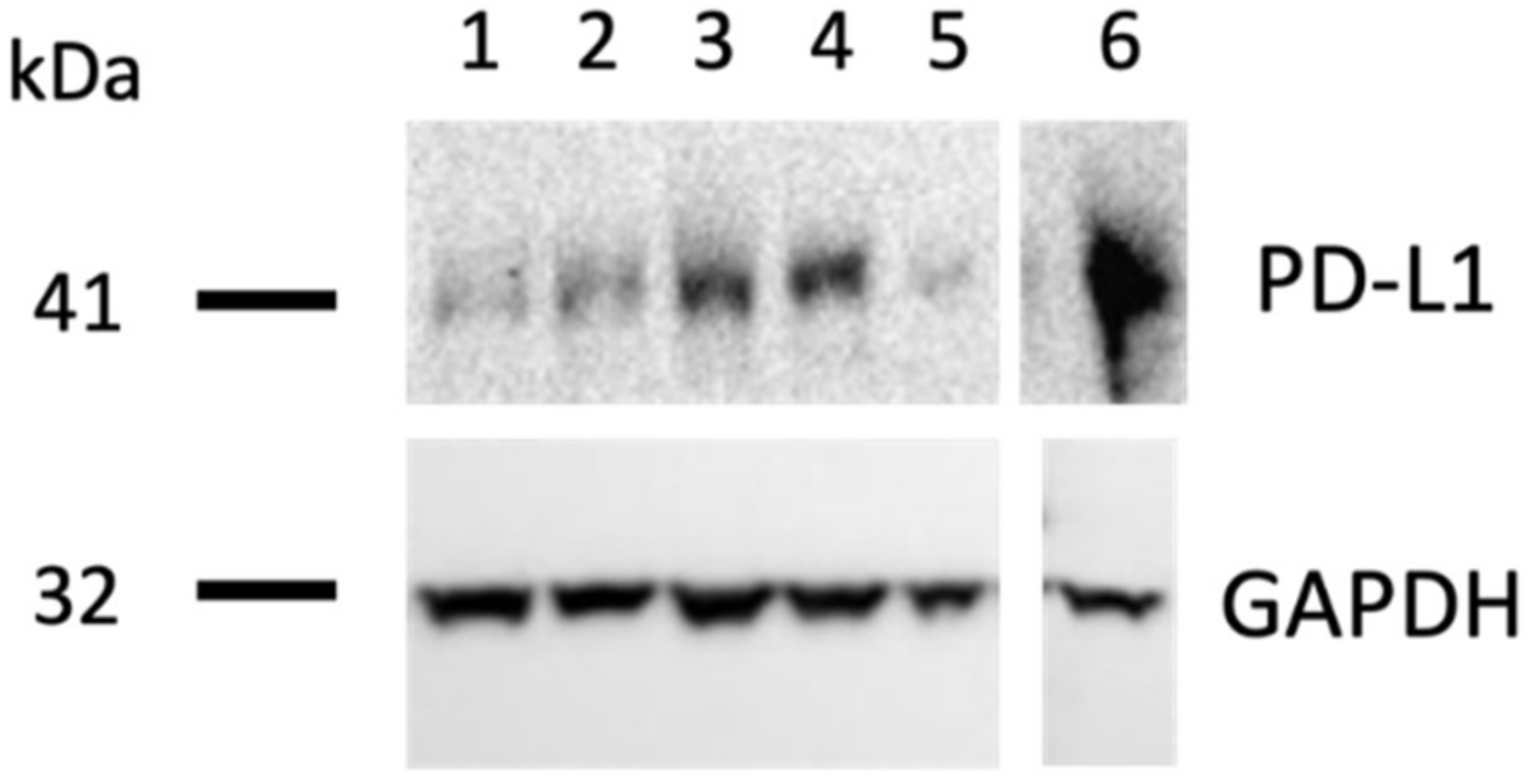
2.6. Western Blot Analysis of PD-L1 Expression In Ovo
2.7. Chicken Chorioallantoic Membrane Assay (CAM) Assay
2.8. Quantitative Evaluation of Immune Cells Infiltration by RT-qPCR
2.9. Statistical Analysis and Significance
3. Results
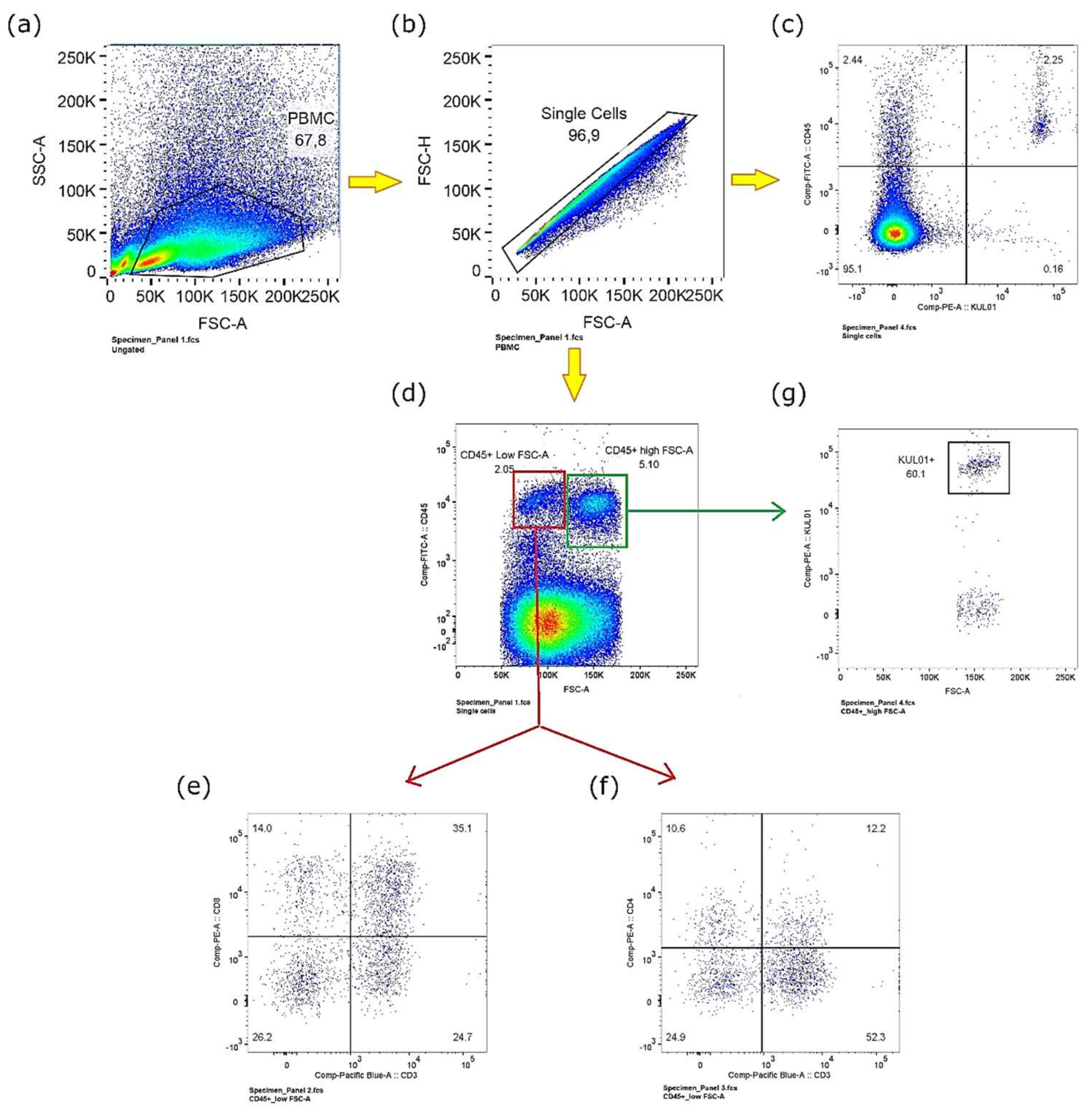
3.1. Immunophenotypic Characterization of Chicken Peripheral Blood Mononuclear Cells
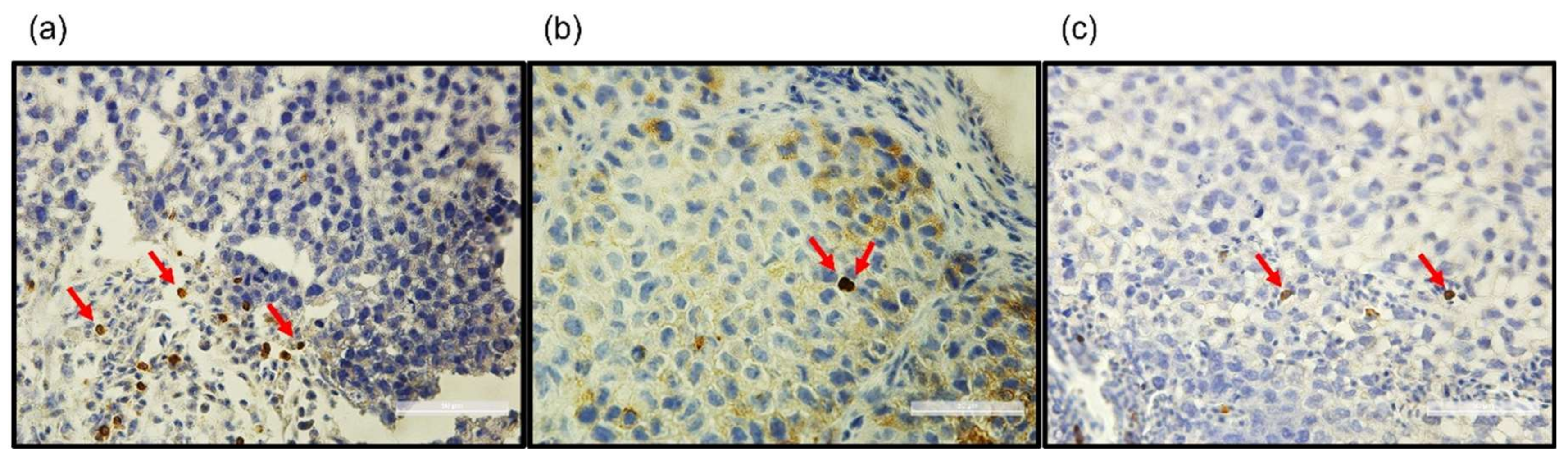
3.2. Tumor-Infiltrating Lymphocytes Are Visualized In Ovo
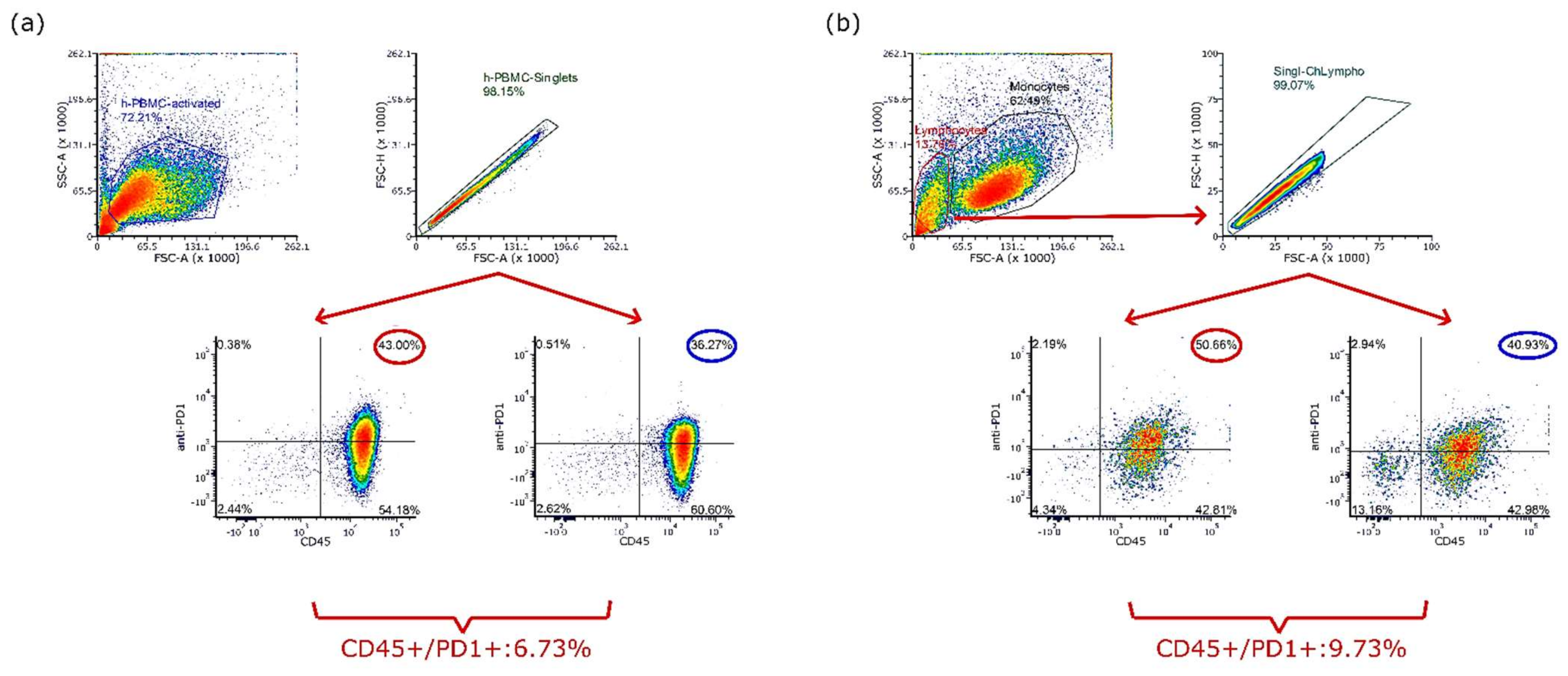
3.3. Anti-Human PD-1 Cross-Reacts with Activated Chicken Splenocytes In Vitro
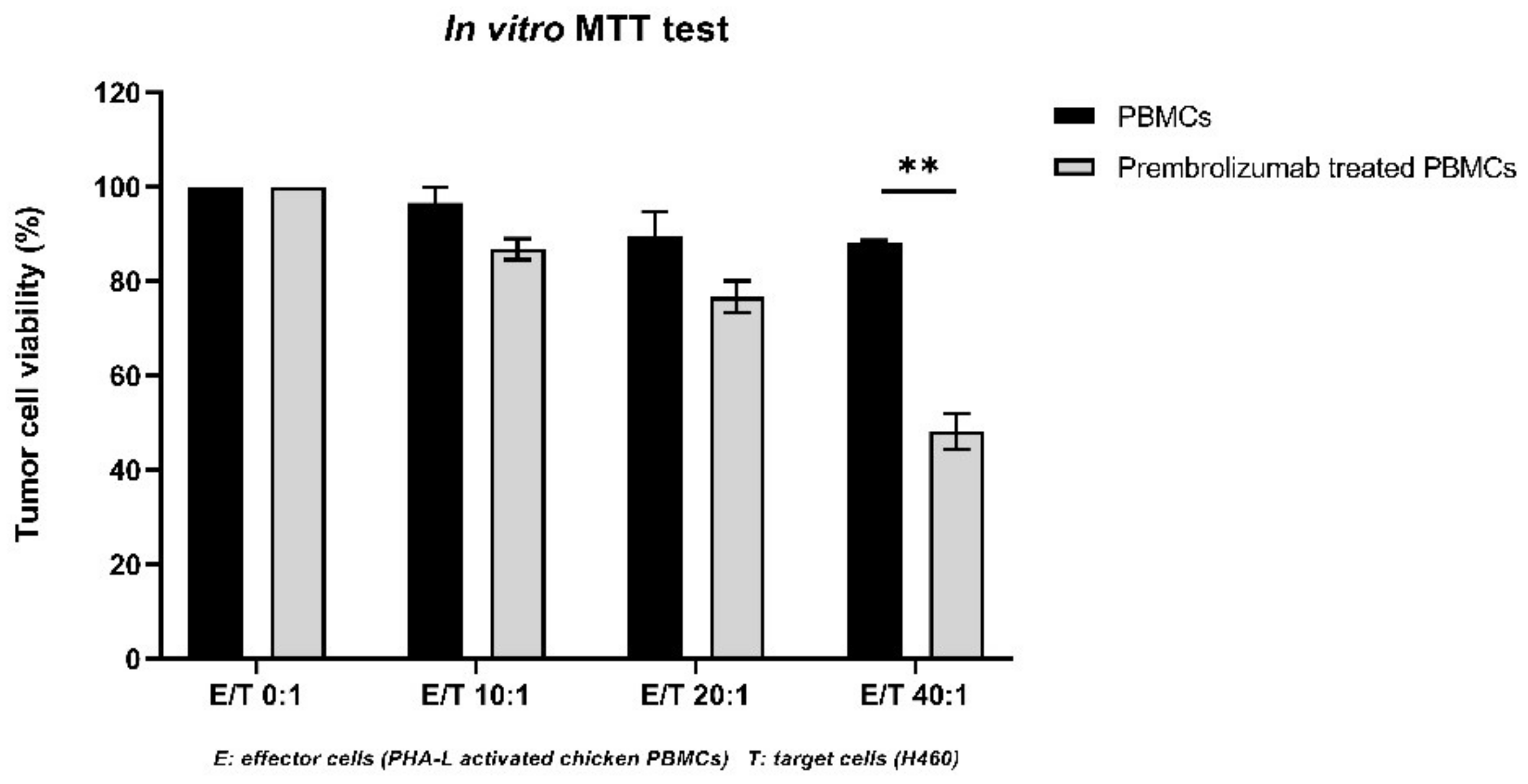
3.4. Pembrolizumab Induces PD-1/PD-L1 Blockade and Restores Chicken T-Cell’s Cytotoxicity against Tumor Cells
3.5. PD-L1 Epitope Is Preserved on Tumors Grown on the CAM
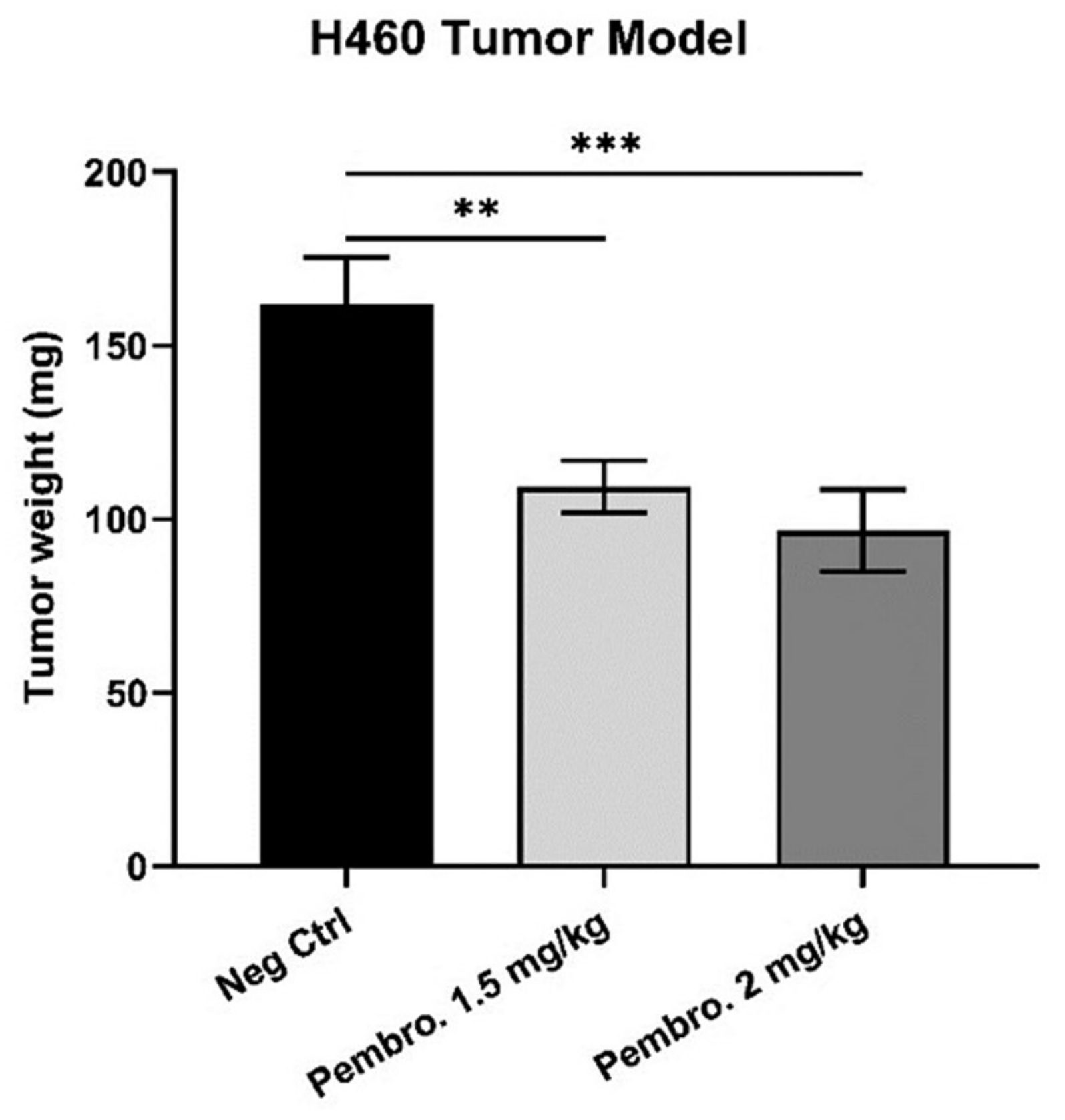
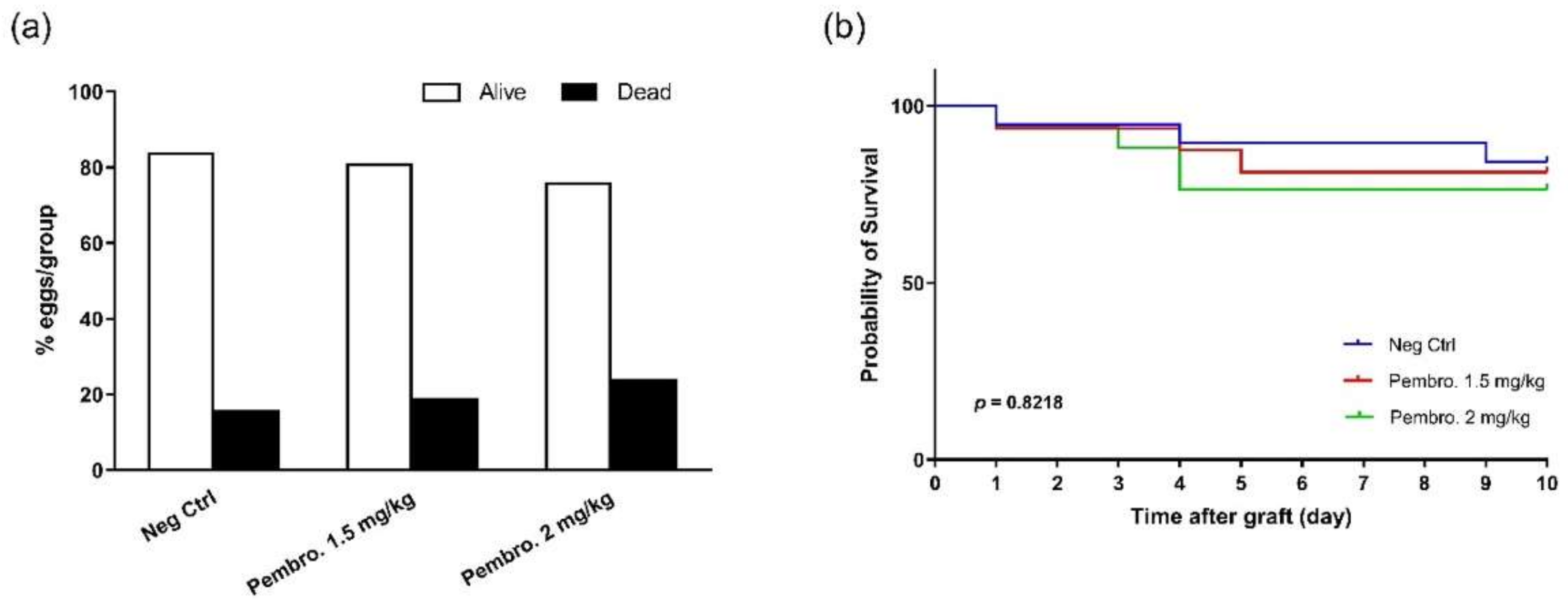
3.6. Pembrolizumab Induces H460 Lung Tumor Growth Regression In Ovo at Safe Doses
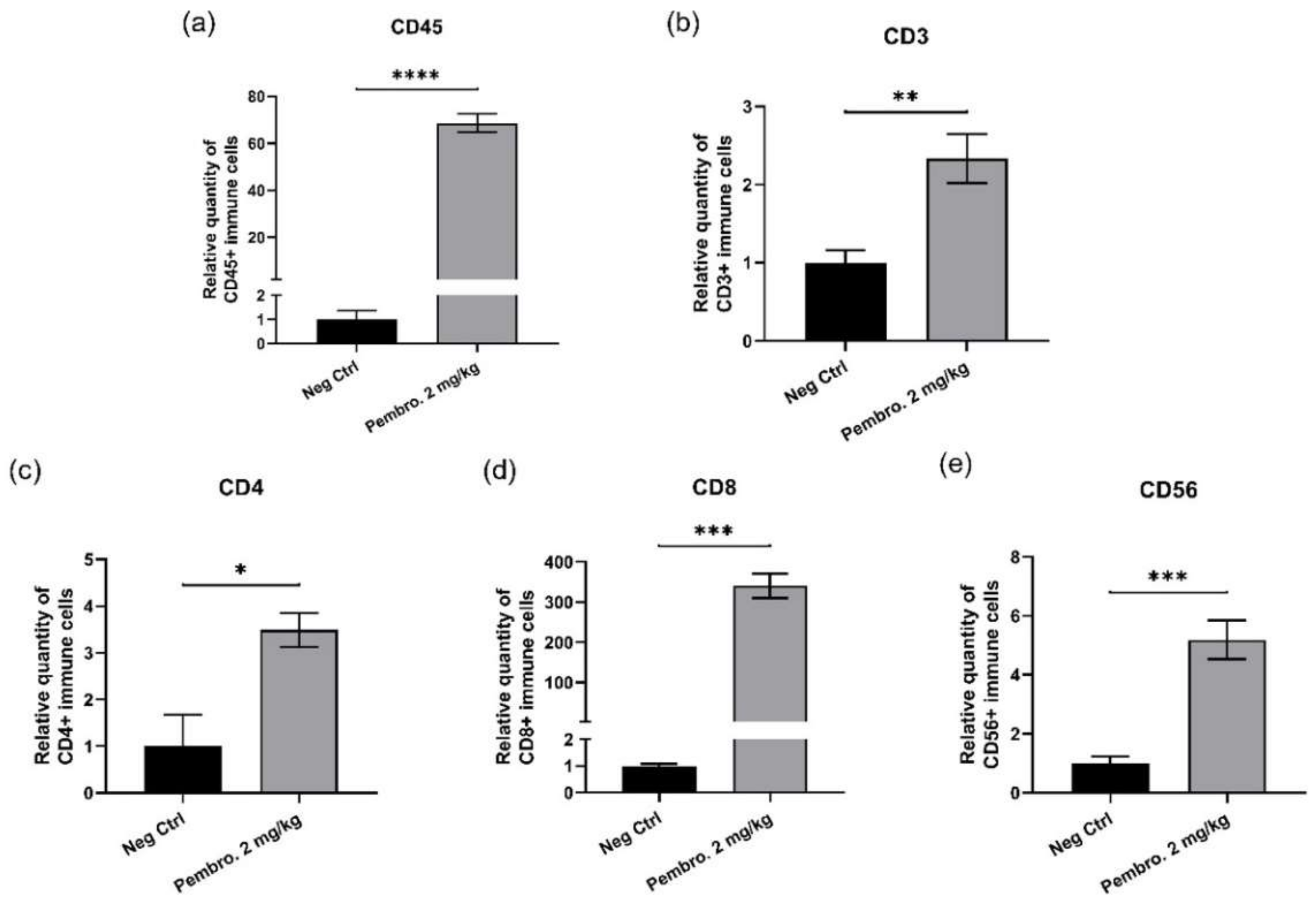
3.7. Pembrolizumab Increases Immune Cell Infiltration in H460 Tumor Xenografted In Ovo
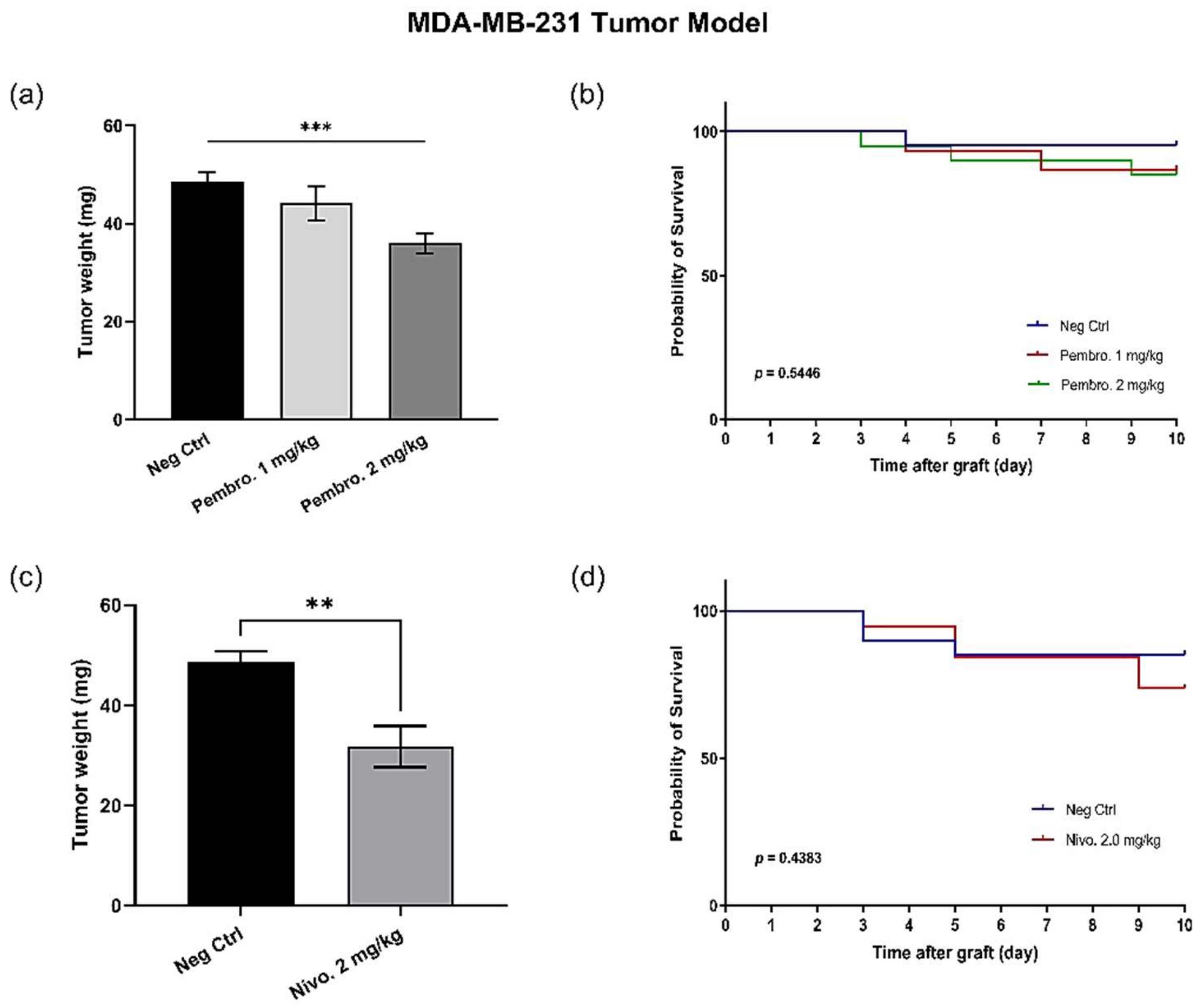
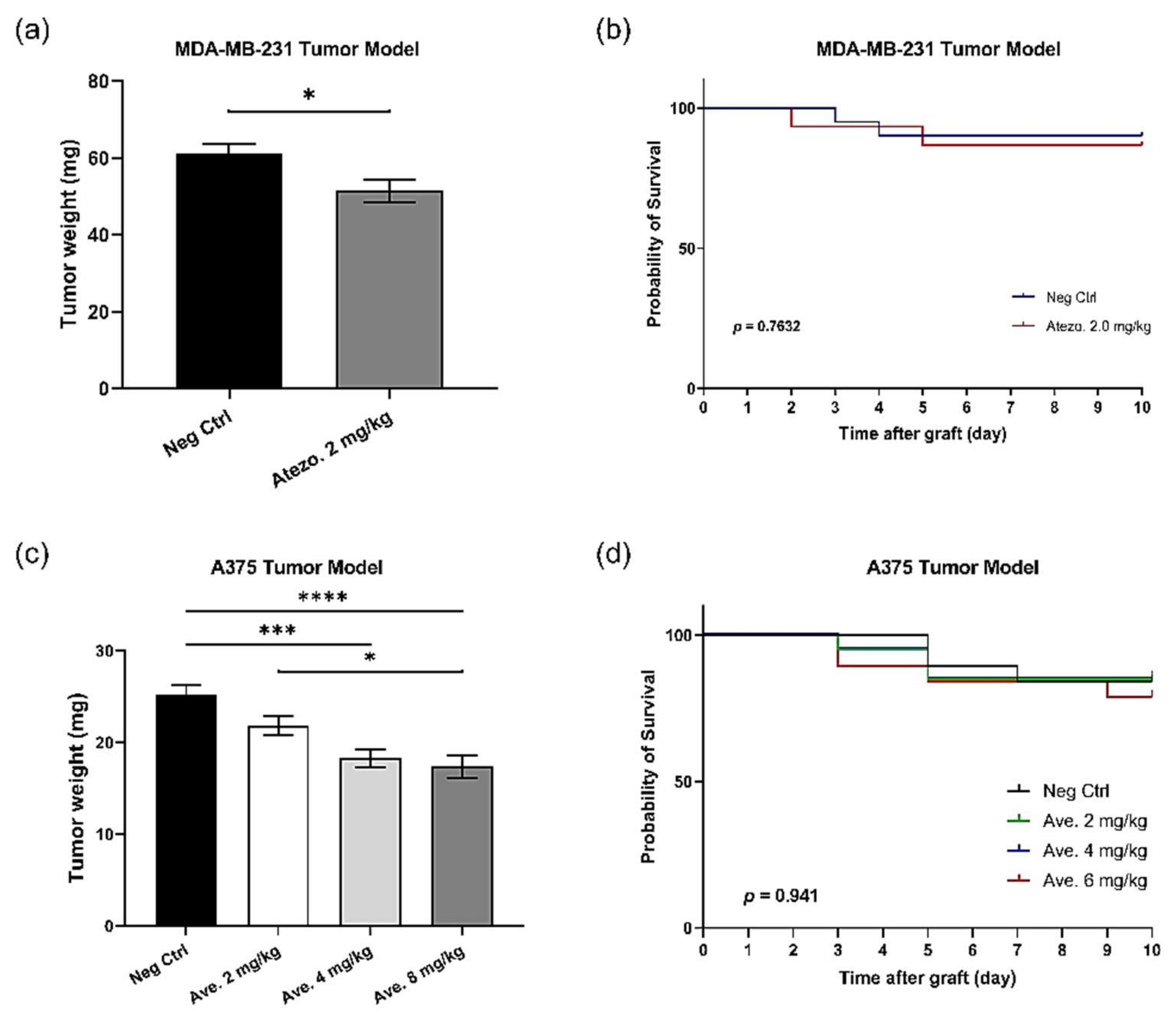
3.8. Efficacy of PD-1 Checkpoint Inhibitors Are also Confirmed with MDA-MB-231 Triple-Negative Breast Cancer In Ovo
3.9. PD-L1 Checkpoint Inhibitors Are Efficient In Ovo
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Zugazagoitia, J.; Guedes, C.; Ponce, S.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Current Challenges in Cancer Treatment. Clin. Ther. 2016, 38, 1551–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattiuzzi, C.; Lippi, G. Current Cancer Epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, D.R.; Krummel, M.F.; Allison, J.P. Enhancement of Antitumor Immunity by CTLA-4 Blockade. Science 1996, 271, 1734–1736. [Google Scholar] [CrossRef] [Green Version]
- Hodi, F.S.; Mihm, M.C.; Soiffer, R.J.; Haluska, F.G.; Butler, M.; Seiden, M.V.; Davis, T.; Henry-Spires, R.; MacRae, S.; Willman, A.; et al. Biologic Activity of Cytotoxic T Lymphocyte-Associated Antigen 4 Antibody Blockade in Previously Vaccinated Metastatic Melanoma and Ovarian Carcinoma Patients. Proc. Natl. Acad. Sci. USA 2003, 100, 4712–4717. [Google Scholar] [CrossRef] [Green Version]
- Ishida, Y.; Agata, Y.; Shibahara, K.; Honjo, T. Induced Expression of PD-1, a Novel Member of the Immunoglobulin Gene Superfamily, upon Programmed Cell Death. EMBO J. 1992, 11, 3887–3895. [Google Scholar] [CrossRef]
- Iwai, Y.; Terawaki, S.; Honjo, T. PD-1 Blockade Inhibits Hematogenous Spread of Poorly Immunogenic Tumor Cells by Enhanced Recruitment of Effector T Cells. Int. Immunol. 2005, 17, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The Clinical Role of the TME in Solid Cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Karpel, M.E.; Boutwell, C.L.; Allen, T.M. BLT Humanized Mice as a Small Animal Model of HIV Infection. Curr. Opin. Virol. 2015, 13, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Shultz, L.D.; Brehm, M.A.; Bavari, S.; Greiner, D.L. Humanized Mice as a Preclinical Tool for Infectious Disease and Biomedical Research: Shultz et Al. Ann. N.Y. Acad. Sci. 2011, 1245, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, X.-J.; Chen, D.-X.; Liu, X.-N.; Wang, X.-J. Humanized Mouse Model: A Review on Preclinical Applications for Cancer Immunotherapy. Am. J. Cancer Res. 2020, 10, 4568–4584. [Google Scholar] [PubMed]
- Rous, P.; Murphy, J.B. Tumor Implantations in the Developing Embryo. J. Am. Med. Assoc. 1911, LVI, 741–742. [Google Scholar] [CrossRef]
- Murphy, J.B. Transplantability of Tissues to the Embryo of Foreign Species: Its Bearing on Questions of Tissue Specificity and Tumor Immunity. J. Exp. Med. 1913, 17, 482–493. [Google Scholar] [CrossRef] [Green Version]
- Achkar, I.W.; Kader, S.; Dib, S.S.; Junejo, K.; Al-Bader, S.B.; Hayat, S.; Bhagwat, A.M.; Rousset, X.; Wang, Y.; Viallet, J.; et al. Metabolic Signatures of Tumor Responses to Doxorubicin Elucidated by Metabolic Profiling in ovo. Metabolites 2020, 10, 268. [Google Scholar] [CrossRef]
- El Hasasna, H.; Saleh, A.; Samri, H.A.; Athamneh, K.; Attoub, S.; Arafat, K.; Benhalilou, N.; Alyan, S.; Viallet, J.; Dhaheri, Y.A.; et al. Rhus Coriaria Suppresses Angiogenesis, Metastasis and Tumor Growth of Breast Cancer through Inhibition of STAT3, NFκB and Nitric Oxide Pathways. Sci. Rep. 2016, 6, 21144. [Google Scholar] [CrossRef]
- Kundeková, B.; Máčajová, M.; Meta, M.; Čavarga, I.; Bilčík, B. Chorioallantoic Membrane Models of Various Avian Species: Differences and Applications. Biology 2021, 10, 301. [Google Scholar] [CrossRef]
- Marcion, G.; Hermetet, F.; Neiers, F.; Uyanik, B.; Dondaine, L.; Dias, A.M.M.; Da Costa, L.; Moreau, M.; Bellaye, P.; Collin, B.; et al. Nanofitins Targeting Heat Shock Protein 110: An Innovative Immunotherapeutic Modality in Cancer. Int. J. Cancer 2021, 148, 3019–3031. [Google Scholar] [CrossRef]
- Nowak-Sliwinska, P.; Segura, T.; Iruela-Arispe, M.L. The Chicken Chorioallantoic Membrane Model in Biology, Medicine and Bioengineering. Angiogenesis 2014, 17, 779–804. [Google Scholar] [CrossRef] [Green Version]
- Rezzola, S.; Loda, A.; Corsini, M.; Semeraro, F.; Annese, T.; Presta, M.; Ribatti, D. Angiogenesis-Inflammation Cross Talk in Diabetic Retinopathy: Novel Insights from the Chick Embryo Chorioallantoic Membrane/Human Vitreous Platform. Front. Immunol. 2020, 11, 581288. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R. The Chick Embryo Chorioallantoic Membrane as an in vivo Experimental Model to Study Multiple Myeloma. In The Enzymes; Elsevier: Amsterdam, The Netherlands, 2019; Volume 46, pp. 23–35. ISBN 978-0-12-817398-5. [Google Scholar]
- Schneider-Stock, R.; Ribatti, D. The CAM Assay as an Alternative In vivo Model for Drug Testing. Organotypic Models in Drug Development. In Handbook of Experimental Pharmacology; Schäfer-Korting, M., Stuchi Maria-Engler, S., Landsiedel, R., Eds.; Springer International Publishing: Cham, Switzerland, 2020; Volume 265, pp. 303–323. ISBN 978-3-030-70062-1. [Google Scholar]
- Garcia, P.; Wang, Y.; Viallet, J.; Macek Jilkova, Z. The Chicken Embryo Model: A Novel and Relevant Model for Immune-Based Studies. Front. Immunol. 2021, 12, 791081. [Google Scholar] [CrossRef] [PubMed]
- Lowenthal, J.W.; Connick, T.; McWaters, P.G.; York, J.J. Development of T Cell Immune Responsiveness in the Chicken. Immunol. Cell Biol. 1994, 72, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, S.; Feng, Y.; Zhang, J.; Du, Y.; Zhang, J.; Ongeval, C.V.; Ni, Y.; Li, Y. Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research. Cells 2021, 10, 463. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane as a Model for Tumor Biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Horita, S.; Nomura, Y.; Sato, Y.; Shimamura, T.; Iwata, S.; Nomura, N. High-Resolution Crystal Structure of the Therapeutic Antibody Pembrolizumab Bound to the Human PD-1. Sci. Rep. 2016, 6, 35297. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.R.; Platero, S.; Saini, K.S.; Curigliano, G.; Anderson, S. Immuno-Oncology Trends: Preclinical Models, Biomarkers, and Clinical Development. J. Immunother. Cancer 2022, 10, e003231. [Google Scholar] [CrossRef]
- Farhood, B.; Najafi, M.; Mortezaee, K. CD8+ Cytotoxic T Lymphocytes in Cancer Immunotherapy: A Review. J. Cell. Physiol. 2019, 234, 8509–8521. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T Cells in Cancer and Cancer Immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Tay, R.E.; Richardson, E.K.; Toh, H.C. Revisiting the Role of CD4+ T Cells in Cancer Immunotherapy—New Insights into Old Paradigms. Cancer Gene Ther. 2021, 28, 5–17. [Google Scholar] [CrossRef]
- Church, S.E.; Jensen, S.M.; Antony, P.A.; Restifo, N.P.; Fox, B.A. Tumor-Specific CD4+ T Cells Maintain Effector and Memory Tumor-Specific CD8+ T Cells. Eur. J. Immunol. 2014, 44, 69–79. [Google Scholar] [CrossRef]
- Lai, Y.-P.; Jeng, C.-J.; Chen, S.-C. The Roles of CD4+ T Cells in Tumor Immunity. ISRN Immunol. 2011, 2011, e497397. [Google Scholar] [CrossRef] [Green Version]
- Ahrends, T.; Borst, J. The Opposing Roles of CD4+ T Cells in Anti-Tumour Immunity. Immunology 2018, 154, 582–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padgett, L.E.; Araujo, D.J.; Hedrick, C.C.; Olingy, C.E. Functional Crosstalk between T Cells and Monocytes in Cancer and Atherosclerosis. J. Leukoc. Biol. 2020, 108, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.B.; Jacob, M.K.; Parajuli, P. Chapter Three-Review of Immune Checkpoint Inhibitors in Immuno-Oncology. In Advances in Pharmacology; Advances in Immunopharmacology; Copple, B.L., Rockwell, C.E., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 91, pp. 111–139. [Google Scholar]
- Liu, J.; Chen, Z.; Li, Y.; Zhao, W.; Wu, J.; Zhang, Z. PD-1/PD-L1 Checkpoint Inhibitors in Tumor Immunotherapy. Front. Pharmacol. 2021, 12, 731798. [Google Scholar] [CrossRef] [PubMed]
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 Pathway in Tolerance and Autoimmunity. Immunol. Rev. 2010, 236, 219–242. [Google Scholar] [CrossRef]
- Elezov, D.S.; Kudryavtsev, I.V. PD-1 Receptor on Immune Cells, Its Expression and Potential Role in Cancer Therapy. Cell. Ther. Transplant. 2019, 8, 8–16. [Google Scholar] [CrossRef]
- Jubel, J.M.; Barbati, Z.R.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. The Role of PD-1 in Acute and Chronic Infection. Front. Immunol. 2020, 11, 487. [Google Scholar] [CrossRef] [Green Version]
- Tovar, Z.; Dauphinée, M.; Talal, N. Synergistic Interaction between Anti-CD3 and IL-2 Demonstrated by Proliferative Response, Interferon Production, and Non-MHC-Restricted Killing. Cell. Immunol. 1988, 117, 12–21. [Google Scholar] [CrossRef]
- Bacot, S.M.; Harper, T.A.; Matthews, R.L.; Fennell, C.J.; Akue, A.; KuKuruga, M.A.; Lee, S.; Wang, T.; Feldman, G.M. Exploring the Potential Use of a PBMC-Based Functional Assay to Identify Predictive Biomarkers for Anti-PD-1 Immunotherapy. Int. J. Mol. Sci. 2020, 21, 9023. [Google Scholar] [CrossRef]
- Wallberg, M.; Recino, A.; Phillips, J.; Howie, D.; Vienne, M.; Paluch, C.; Azuma, M.; Wong, F.S.; Waldmann, H.; Cooke, A. Anti-CD3 Treatment up-Regulates Programmed Cell Death Protein-1 Expression on Activated Effector T Cells and Severely Impairs Their Inflammatory Capacity. Immunology 2017, 151, 248–260. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Usui, Y.; Horie, S.; Futagami, Y.; Yamada, Y.; Ma, J.; Kezuka, T.; Hamada, H.; Usui, T.; Mochizuki, M.; et al. Human Corneal Endothelial Cells Expressing Programmed Death-Ligand 1 (PD-L1) Suppress PD-1+ T Helper 1 Cells by a Contact-Dependent Mechanism. Investig. Ophthalmol. Vis. Sci. 2009, 50, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Chen, P.W.; Li, H.; Alizadeh, H.; Niederkorn, J.Y. PD-L1: PD-1 Interaction Contributes to the Functional Suppression of T-Cell Responses to Human Uveal Melanoma Cells In Vitro. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Champiat, S.; Lambotte, O.; Barreau, E.; Belkhir, R.; Berdelou, A.; Carbonnel, F.; Cauquil, C.; Chanson, P.; Collins, M.; Durrbach, A.; et al. Management of Immune Checkpoint Blockade Dysimmune Toxicities: A Collaborative Position Paper. Ann. Oncol. 2016, 27, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, J.; Page, D.B.; Li, B.T.; Connell, L.C.; Schindler, K.; Lacouture, M.E.; Postow, M.A.; Wolchok, J.D. Toxicities of the Anti-PD-1 and Anti-PD-L1 Immune Checkpoint Antibodies. Ann. Oncol. 2015, 26, 2375–2391. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, M.J.H.; Härtle, S. Chapter 4—B Cells, the Bursa of Fabricius, and the Generation of Antibody Repertoires. In Avian Immunology, 3rd ed.; Kaspers, B., Schat, K.A., Göbel, T.W., Vervelde, L., Eds.; Academic Press: Boston, MA, USA, 2022; pp. 71–99. ISBN 978-0-12-818708-1. [Google Scholar]
- Lee, L.; Samardzic, K.; Wallach, M.; Frumkin, L.R.; Mochly-Rosen, D. Immunoglobulin Y for Potential Diagnostic and Therapeutic Applications in Infectious Diseases. Front. Immunol. 2021, 12, 2257. [Google Scholar] [CrossRef]
- Ching, K.H.; Collarini, E.J.; Abdiche, Y.N.; Bedinger, D.; Pedersen, D.; Izquierdo, S.; Harriman, R.; Zhu, L.; Etches, R.J.; van de Lavoir, M.-C.; et al. Chickens with Humanized Immunoglobulin Genes Generate Antibodies with High Affinity and Broad Epitope Coverage to Conserved Targets. mAbs 2018, 10, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Davison, F. Chapter 1—The Importance of the Avian Immune System and Its Unique Features. In Avian Immunology, 3rd ed.; Kaspers, B., Schat, K.A., Göbel, T.W., Vervelde, L., Eds.; Academic Press: Boston, MA, USA, 2022; pp. 1–9. ISBN 978-0-12-818708-1. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Rousset, X.; Prunier, C.; Garcia, P.; Dosda, E.; Leplus, E.; Viallet, J. PD-1/PD-L1 Checkpoint Inhibitors Are Active in the Chicken Embryo Model and Show Antitumor Efficacy In Ovo. Cancers 2022, 14, 3095. https://doi.org/10.3390/cancers14133095
Wang Y, Rousset X, Prunier C, Garcia P, Dosda E, Leplus E, Viallet J. PD-1/PD-L1 Checkpoint Inhibitors Are Active in the Chicken Embryo Model and Show Antitumor Efficacy In Ovo. Cancers. 2022; 14(13):3095. https://doi.org/10.3390/cancers14133095
Chicago/Turabian StyleWang, Yan, Xavier Rousset, Chloé Prunier, Paul Garcia, Emilien Dosda, Estelle Leplus, and Jean Viallet. 2022. "PD-1/PD-L1 Checkpoint Inhibitors Are Active in the Chicken Embryo Model and Show Antitumor Efficacy In Ovo" Cancers 14, no. 13: 3095. https://doi.org/10.3390/cancers14133095