Macrophage Involvement in Medication-Related Osteonecrosis of the Jaw (MRONJ): A Comprehensive, Short Review

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
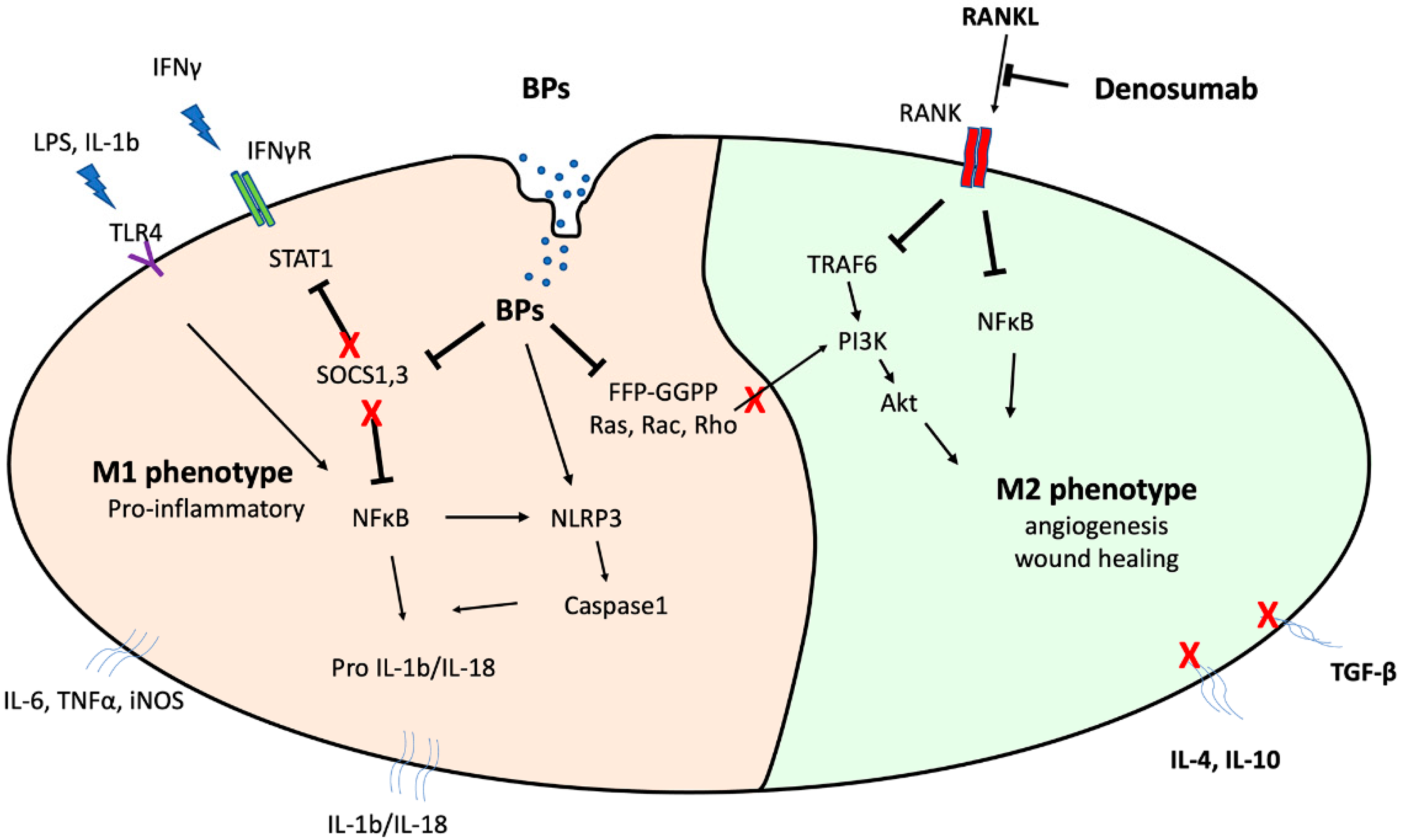
2. Macrophage Involvement in MRONJ Pathophysiology
2.1. Macrophage Biology
2.2. In Vitro Studies
2.3. Alterations in Macrophage Growth and Function
2.4. BP Mechanism of Action against Macrophages
2.5. Alterations in Macrophage Protein Expression
2.6. Alterations in Macrophage Polarization
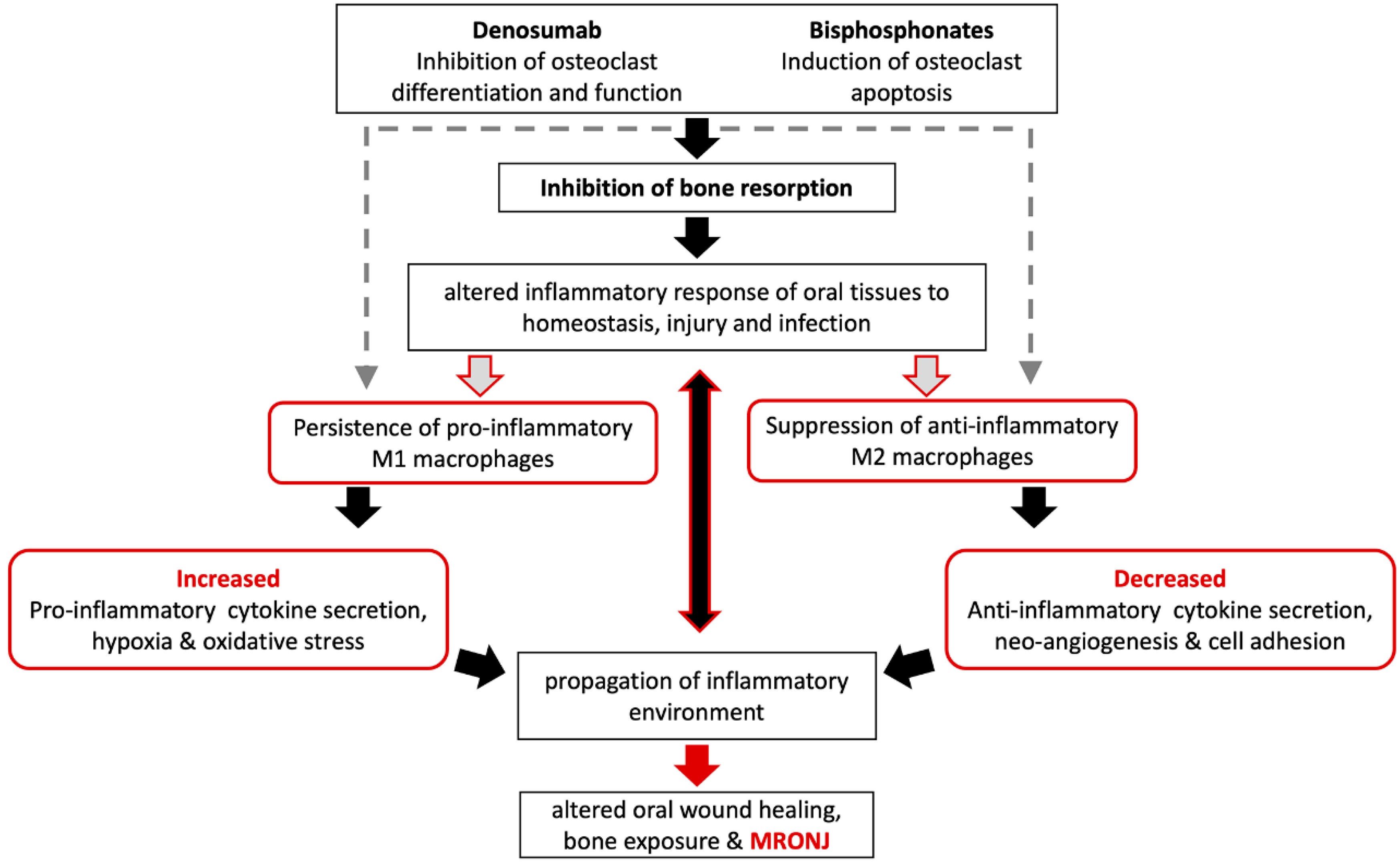
3. Clinical Observations and In Vivo MRONJ Animal Models
3.1. Macrophages May Modulate MRONJ Incidence and Wound Healing
3.2. Macrophage Polarization Changes in MRONJ Tissues
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ruggiero, S.L.; Dodson, T.B.; Fantasia, J.; Goodday, R.; Aghaloo, T.; Mehrotra, B.; O’Ryan, F. American Association of Oral and Maxillofacial Surgeons position paper on medication-related osteonecrosis of the jaw—2014 update. J. Oral Maxillofac. Surg. Off. J. Am. Assoc. Oral Maxillofac. Surg. 2014, 72, 1938–1956. [Google Scholar] [CrossRef] [PubMed]
- Fliefel, R.; Tröltzsch, M.; Kühnisch, J.; Ehrenfeld, M.; Otto, S. Treatment strategies and outcomes of bisphosphonate-related osteonecrosis of the jaw (BRONJ) with characterization of patients: A systematic review. Int. J. Oral Maxillofac. Surg. 2015, 44, 568–585. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, S.; Soutome, S.; Yanamoto, S.; Fujita, S.; Hasegawa, T.; Komori, T.; Kojima, Y.; Miyamoto, H.; Shibuya, Y.; Ueda, N.; et al. Evaluation of the Treatment Strategies for Medication-Related Osteonecrosis of the Jaws (MRONJ) and the Factors Affecting Treatment Outcome: A Multicenter Retrospective Study with Propensity Score Matching Analysis. J. Bone Miner. Res. 2017, 32, 2022–2029. [Google Scholar] [CrossRef] [Green Version]
- Nicolatou-Galitis, O.; Schiødt, M.; Mendes, R.A.; Ripamonti, C.; Hope, S.; Drudge-Coates, L.; Niepel, D.; Van den Wyngaert, T. Medication-related osteonecrosis of the jaw: Definition and best practice for prevention, diagnosis, and treatment. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 127, 117–135. [Google Scholar] [CrossRef] [Green Version]
- Giudice, A.; Antonelli, A.; Muraca, D.; Fortunato, L. Usefulness of advanced-platelet rich fibrin (A-PRF) and injectable-platelet rich fibrin (i-PRF) in the management of a massive medication-related osteonecrosis of the jaw (MRONJ): A 5-years follow-up case report. Indian J. Dent. Res. Off. Publ. Indian Soc. Dent. Res. 2020, 31, 813–818. [Google Scholar] [CrossRef]
- Hadaya, D.; Soundia, A.; Freymiller, E.; Grogan, T.; Elashoff, D.; Tetradis, S.; Aghaloo, T.L. Nonsurgical Management of Medication-Related Osteonecrosis of the Jaws Using Local Wound Care. J. Oral Maxillofac. Surg. Off. J. Am. Assoc. Oral Maxillofac. Surg. 2018, 76, 2332–2339. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, K.; Hayashida, S.; Rokutanda, S.; Kawakita, A.; Soutome, S.; Sawada, S.; Yanamoto, S.; Kojima, Y.; Umeda, M. Surgical strategy for medication-related osteonecrosis of the jaw (MRONJ) on maxilla: A multicenter retrospective study. J. Dent. Sci. 2021, 16, 885–890. [Google Scholar] [CrossRef]
- Beth-Tasdogan, N.H.; Mayer, B.; Hussein, H.; Zolk, O. Interventions for managing medication-related osteonecrosis of the jaw. Cochrane Database Syst. Rev. 2017, 10, Cd012432. [Google Scholar] [CrossRef]
- Owosho, A.A.; Estilo, C.L.; Huryn, J.M.; Yom, S.K. Pentoxifylline and tocopherol in the management of cancer patients with medication-related osteonecrosis of the jaw: An observational retrospective study of initial case series. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 122, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Aghaloo, T.; Hazboun, R.; Tetradis, S. Pathophysiology of Osteonecrosis of the Jaws. Oral Maxillofac. Surg. Clin. North Am. 2015, 27, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.A.; Morrison, A.; Hanley, D.A.; Felsenberg, D.; McCauley, L.K.; O’Ryan, F.; Reid, I.R.; Ruggiero, S.L.; Taguchi, A.; Tetradis, S.; et al. Diagnosis and management of osteonecrosis of the jaw: A systematic review and international consensus. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2015, 30, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Gkouveris, I.; Hadaya, D.; Soundia, A.; Bezouglaia, O.; Chau, Y.; Dry, S.M.; Pirih, F.Q.; Aghaloo, T.L.; Tetradis, S. Vasculature submucosal changes at early stages of osteonecrosis of the jaw (ONJ). Bone 2019, 123, 234–245. [Google Scholar] [CrossRef]
- Soutome, S.; Otsuru, M.; Hayashida, S.; Murata, M.; Yanamoto, S.; Sawada, S.; Kojima, Y.; Funahara, M.; Iwai, H.; Umeda, M.; et al. Relationship between tooth extraction and development of medication-related osteonecrosis of the jaw in cancer patients. Sci. Rep. 2021, 11, 17226. [Google Scholar] [CrossRef] [PubMed]
- Francisconi, C.F.; Vieira, A.E.; Azevedo, M.C.S.; Tabanez, A.P.; Fonseca, A.C.; Trombone, A.P.F.; Letra, A.; Silva, R.M.; Sfeir, C.S.; Little, S.R.; et al. RANKL Triggers Treg-Mediated Immunoregulation in Inflammatory Osteolysis. J. Dent. Res. 2018, 97, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kanayama, K.; Kaur, K.; Tseng, H.C.; Banankhah, S.; Quje, D.T.; Sayre, J.W.; Jewett, A.; Nishimura, I. Osteonecrosis of the Jaw Developed in Mice: Disease variants regulated by γδ t cells in oral mucosal barrier immunity. J. Biol. Chem. 2015, 290, 17349–17366. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Xu, R.; Du, J.; Fu, Y.; Li, S.; Zhang, P.; Liu, L.; Jiang, H. Zoledronic acid promotes TLR-4–mediated M1 macrophage polarization in bisphosphonate-related osteonecrosis of the jaw. FASEB J. 2019, 33, 5208–5219. [Google Scholar] [CrossRef]
- de Barros Silva, P.G.; de Oliveira, C.C.; Brizeno, L.; Wong, D.; Lima Júnior, R.; Gonçalves, R.P.; Sousa, F.B.; Mota, M.; de Albuquerque Ribeiro, R.; Alves, A. Immune cellular profile of bisphosphonate-related osteonecrosis of the jaw. Oral Dis. 2016, 22, 649–657. [Google Scholar] [CrossRef]
- Elsayed, R.; Kurago, Z.; Cutler, C.W.; Arce, R.M.; Gerber, J.; Celis, E.; Sultan, H.; Elashiry, M.; Meghil, M.; Sun, C.; et al. Role of dendritic cell-mediated immune response in oral homeostasis: A new mechanism of osteonecrosis of the jaw. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 2595–2608. [Google Scholar] [CrossRef]
- Hagelauer, N.; Pabst, A.M.; Ziebart, T.; Ulbrich, H.; Walter, C. In vitro effects of bisphosphonates on chemotaxis, phagocytosis, and oxidative burst of neutrophil granulocytes. Clin. Oral Investig. 2015, 19, 139–148. [Google Scholar] [CrossRef]
- Kikuiri, T.; Kim, I.; Yamaza, T.; Akiyama, K.; Zhang, Q.; Li, Y.; Chen, C.; Chen, W.; Wang, S.; Le, A.D.; et al. Cell-based immunotherapy with mesenchymal stem cells cures bisphosphonate-related osteonecrosis of the jaw-like disease in mice. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2010, 25, 1668–1679. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, J.W.P.; Forster, C.; Sun, C.; Peel, S.; Glogauer, M. Zoledronate and pamidronate depress neutrophil functions and survival in mice. Br. J. Pharmacol. 2012, 165, 532–539. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Atsuta, I.; Liu, S.; Chen, C.; Shi, S.; Shi, S.; Le, A.D. IL-17-mediated M1/M2 macrophage alteration contributes to pathogenesis of bisphosphonate-related osteonecrosis of the jaws. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 3176–3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Wang, Z.; Zhou, T.; Shan, L. Determination of the molecular mechanism by which macrophages and γδ-T cells contribute to ZOL-induced ONJ. Aging 2020, 12, 20743–20752. [Google Scholar] [CrossRef] [PubMed]
- Orsini, G.; Failli, A.; Legitimo, A.; Adinolfi, B.; Romanini, A.; Consolini, R. Zoledronic acid modulates maturation of human monocyte-derived dendritic cells. Exp. Biol. Med. 2011, 236, 1420–1426. [Google Scholar] [CrossRef]
- Di Vito, A.; Chiarella, E.; Baudi, F.; Scardamaglia, P.; Antonelli, A.; Giudice, D.; Barni, T.; Fortunato, L.; Giudice, A. Dose-Dependent Effects of Zoledronic Acid on Human Periodontal Ligament Stem Cells: An In Vitro Pilot Study. Cell Transplant. 2020, 29, 963689720948497. [Google Scholar] [CrossRef]
- Kaiser, T.; Geiger, K.; Wallwiener, D.; Klein, G.; Fehm, T.N. Effects of bisphosphonates on human osteoblasts as an important constituent of the bone marrow microenvironment. J. Clin. Oncol. 2010, 28, e13620. [Google Scholar] [CrossRef]
- Liu, D.; Du, J.; Sun, J.; Li, M. Parathyroid hormone-related protein inhibits nitrogen-containing bisphosphonate-induced apoptosis of human periodontal ligament fibroblasts by activating MKP1 phosphatase. Bioengineered 2021, 12, 1997–2006. [Google Scholar] [CrossRef]
- Shi, Y.; Li, M.; Yu, Y.; Zhou, Y.; Zhang, W.; Hua, H.; Wang, S. Stress response in periodontal ligament stem cells may contribute to bisphosphonate-associated osteonecrosis of the jaw: A gene expression array analysis. Mol. Med. Rep. 2020, 22, 2043–2051. [Google Scholar] [CrossRef]
- Walter, C.; Klein, M.O.; Pabst, A.; Al-Nawas, B.; Duschner, H.; Ziebart, T. Influence of bisphosphonates on endothelial cells, fibroblasts, and osteogenic cells. Clin. Oral Investig. 2010, 14, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets. Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-wound Healing Phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef]
- Lavin, Y.; Mortha, A.; Rahman, A.; Merad, M. Regulation of macrophage development and function in peripheral tissues. Nat. Rev. Immunol. 2015, 15, 731–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoefert, S.; Hoefert, C.S.; Albert, M.; Munz, A.; Grimm, M.; Northoff, H.; Reinert, S.; Alexander, D. Zoledronate but not denosumab suppresses macrophagic differentiation of THP-1 cells. An aetiologic model of bisphosphonate-related osteonecrosis of the jaw (BRONJ). Clin. Oral Investig. 2015, 19, 1307–1318. [Google Scholar] [CrossRef]
- Hoefert, S.; Schmitz, I.; Weichert, F.; Gaspar, M.; Eufinger, H. Macrophages and bisphosphonate-related osteonecrosis of the jaw (BRONJ): Evidence of local immunosuppression of macrophages in contrast to other infectious jaw diseases. Clin. Oral Investig. 2015, 19, 497–508. [Google Scholar] [CrossRef]
- Paschalidi, P.; Gkouveris, I.; Soundia, A.; Kalfarentzos, E.; Vardas, E.; Georgaki, M.; Kostakis, G.; Erovic, B.M.; Tetradis, S.; Perisanidis, C.; et al. The role of M1 and M2 macrophage polarization in progression of medication-related osteonecrosis of the jaw. Clin. Oral Investig. 2020, 25, 2845–2857. [Google Scholar] [CrossRef]
- Huang, R.; Wang, X.; Zhou, Y.; Xiao, Y. RANKL-induced M1 macrophages are involved in bone formation. Bone Res. 2017, 5, 17019. [Google Scholar] [CrossRef]
- Canton, J. Phagosome maturation in polarized macrophages. J. Leukoc. Biol. 2014, 96, 729–738. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, X.H.; Jin, L. Macrophage Polarization in Physiological and Pathological Pregnancy. Front. Immunol. 2019, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Riboldi, E.; Ippolito, A.; Sica, A. Molecular and epigenetic basis of macrophage polarized activation. Semin. Immunol. 2015, 27, 237–248. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, J.J.; Paul, W.E. Mechanisms underlying lineage commitment and plasticity of helper CD4+ T cells. Science 2010, 327, 1098–1102. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Plüddemann, A. Tissue macrophages: Heterogeneity and functions. BMC Biol. 2017, 15, 53. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS−) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Ginhoux, F.; Guilliams, M. Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity 2016, 44, 439–449. [Google Scholar] [CrossRef]
- Perdiguero, E.G.; Geissmann, F. The development and maintenance of resident macrophages. Nat. Immunol. 2016, 17, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Pittet, M.J.; Nahrendorf, M.; Swirski, F.K. The journey from stem cell to macrophage. Ann. N. Y. Acad. Sci. 2014, 1319, 1–18. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, H.; Shi, Q. Macrophages and bone inflammation. J. Orthop. Transl. 2017, 10, 86–93. [Google Scholar] [CrossRef]
- Batoon, L.; Millard, S.M.; Raggatt, L.J.; Pettit, A.R. Osteomacs and Bone Regeneration. Curr. Osteoporos. Rep. 2017, 15, 385–395. [Google Scholar] [CrossRef]
- Patntirapong, S.; Poolgesorn, M. Alteration of macrophage viability, differentiation, and function by bisphosphonates. Oral Dis. 2018, 24, 1294–1302. [Google Scholar] [CrossRef]
- Hoefert, S.; Sade Hoefert, C.; Munz, A.; Schmitz, I.; Grimm, M.; Yuan, A.; Northoff, H.; Reinert, S.; Alexander, D. Effect of bisphosphonates on macrophagic THP-1 cell survival in bisphosphonate-related osteonecrosis of the jaw (BRONJ). Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 121, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Hoefert, S.; Sade Hoefert, C.; Munz, A.; Northoff, H.; Yuan, A.; Reichenmiller, K.; Reinert, S.; Grimm, M. Altered macrophagic THP-1 cell phagocytosis and migration in bisphosphonate-related osteonecrosis of the jaw (BRONJ). Clin. Oral Investig. 2016, 20, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Okinaga, T.; Hikiji, H.; Ariyoshi, W.; Yoshiga, D.; Habu, M.; Tominaga, K.; Nishihara, T. Zoledronic acid exacerbates inflammation through M1 macrophage polarization. Inflamm. Regen. 2018, 38, 16. [Google Scholar] [CrossRef] [PubMed]
- Patntirapong, S.; Phupunporn, P.; Vanichtantiphong, D.; Thanetchaloempong, W. Inhibition of macrophage viability by bound and free bisphosphonates. Acta Histochem. 2019, 121, 400–406. [Google Scholar] [CrossRef]
- Muratsu, D.; Yoshiga, D.; Taketomi, T.; Onimura, T.; Seki, Y.; Matsumoto, A.; Nakamura, S. Zoledronic acid enhances lipopolysaccharide-stimulated proinflammatory reactions through controlled expression of SOCS1 in macrophages. PLoS ONE 2013, 8, e67906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimachi, K.; Kajiya, H.; Nakayama, S.; Ikebe, T.; Okabe, K. Zoledronic acid inhibits RANK expression and migration of osteoclast precursors during osteoclastogenesis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2011, 383, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Shikama, Y.; Nagai, Y.; Okada, S.; Oizumi, T.; Shimauchi, H.; Sugawara, S.; Endo, Y. Pro-IL-1β accumulation in macrophages by alendronate and its prevention by clodronate. Toxicol. Lett. 2010, 199, 123–128. [Google Scholar] [CrossRef]
- Yang, X.; Xu, X.; Chen, J.; Wang, Q.; Wang, G.; Ai, X.; Wang, X.; Pan, J. Zoledronic acid regulates the synthesis and secretion of IL-1β through Histone methylation in macrophages. Cell Death Discov. 2020, 6, 47. [Google Scholar] [CrossRef]
- Morita, M.; Iwasaki, R.; Sato, Y.; Kobayashi, T.; Watanabe, R.; Oike, T.; Nakamura, S.; Keneko, Y.; Miyamoto, K.; Ishihara, K.; et al. Elevation of pro-inflammatory cytokine levels following anti-resorptive drug treatment is required for osteonecrosis development in infectious osteomyelitis. Sci. Rep. 2017, 7, 46322. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, W.; Lee, S.; Xu, Q.; Naji, A.; Le, A.D. Bisphosphonate Induces Osteonecrosis of the Jaw in Diabetic Mice via NLRP3/Caspase-1-Dependent IL-1β Mechanism. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2015, 30, 2300–2312. [Google Scholar] [CrossRef] [Green Version]
- Scheller, E.L.; Hankenson, K.D.; Reuben, J.S.; Krebsbach, P.H. Zoledronic acid inhibits macrophage SOCS3 expression and enhances cytokine production. J. Cell. Biochem. 2011, 112, 3364–3372. [Google Scholar] [CrossRef] [Green Version]
- Hayano, H.; Kuroshima, S.; Sasaki, M.; Tamaki, S.; Inoue, M.; Ishisaki, A.; Sawase, T. Distinct immunopathology in the early stages between different antiresorptives-related osteonecrosis of the jaw-like lesions in mice. Bone 2020, 135, 115308. [Google Scholar] [CrossRef]
- Wehrhan, F.; Moebius, P.; Amann, K.; Ries, J.; Preidl, R.; Neukam, F.W.; Weber, M. Macrophage and osteoclast polarization in bisphosphonate associated necrosis and osteoradionecrosis. J. Cranio-Maxillo-Facial Surg. Off. Publ. Eur. Assoc. Cranio-Maxillo-Facial Surg. 2017, 45, 944–953. [Google Scholar] [CrossRef]
- Kuroshima, S.; Al-Salihi, Z.; Yamashita, J. Mouse anti-RANKL antibody delays oral wound healing and increases TRAP-positive mononuclear cells in bone marrow. Clin. Oral Investig. 2016, 20, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, S.; Kuroshima, S.; Hayano, H.; Nakajima, K.; Kakehashi, H.; Ishisaki, A.; Sawase, T. Dynamic polarization shifting from M1 to M2 macrophages in reduced osteonecrosis of the jaw-like lesions by cessation of anti-RANKL antibody in mice. Bone 2020, 141, 115560. [Google Scholar] [CrossRef]
- Dougall, W.C.; Glaccum, M.; Charrier, K.; Rohrbach, K.; Brasel, K.; De Smedt, T.; Daro, E.; Smith, J.; Tometsko, M.E.; Maliszewski, C.R.; et al. RANK is essential for osteoclast and lymph node development. Genes Dev. 1999, 13, 2412–2424. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.Y.; Yoshida, H.; Sarosi, I.; Tan, H.L.; Timms, E.; Capparelli, C.; Morony, S.; Oliveira-dos-Santos, A.J.; Van, G.; Itie, A.; et al. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Seshasayee, D.; Wang, H.; Lee, W.P.; Gribling, P.; Ross, J.; Van Bruggen, N.; Carano, R.; Grewal, I.S. A novel in vivo role for osteoprotegerin ligand in activation of monocyte effector function and inflammatory response. J. Biol. Chem. 2004, 279, 30202–30209. [Google Scholar] [CrossRef] [Green Version]
- Coxon, F.P.; Thompson, K.; Roelofs, A.J.; Ebetino, F.H.; Rogers, M.J. Visualizing mineral binding and uptake of bisphosphonate by osteoclasts and non-resorbing cells. Bone 2008, 42, 848–860. [Google Scholar] [CrossRef]
- Artz, J.D.; Wernimont, A.K.; Dunford, J.E.; Schapira, M.; Dong, A.; Zhao, Y.; Lew, J.; Russell, R.G.; Ebetino, F.H.; Oppermann, U.; et al. Molecular characterization of a novel geranylgeranyl pyrophosphate synthase from Plasmodium parasites. J. Biol. Chem. 2011, 286, 3315–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, R.G. Bisphosphonates: The first 40 years. Bone 2011, 49, 2–19. [Google Scholar] [CrossRef]
- Licata, A.A. Discovery, clinical development, and therapeutic uses of bisphosphonates. Ann. Pharmacother. 2005, 39, 668–677. [Google Scholar] [CrossRef]
- Herre, J.; Marshall, A.S.; Caron, E.; Edwards, A.D.; Williams, D.L.; Schweighoffer, E.; Tybulewicz, V.; Reis e Sousa, C.; Gordon, S.; Brown, G.D. Dectin-1 uses novel mechanisms for yeast phagocytosis in macrophages. Blood 2004, 104, 4038–4045. [Google Scholar] [CrossRef] [Green Version]
- Rougerie, P.; Miskolci, V.; Cox, D. Generation of membrane structures during phagocytosis and chemotaxis of macrophages: Role and regulation of the actin cytoskeleton. Immunol. Rev. 2013, 256, 222–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, J.L.; Hartwig, J.H.; Tardy, Y.; Dewey, C.F., Jr. Measuring actin dynamics in endothelial cells. Microsc. Res. Tech. 1998, 43, 385–394. [Google Scholar] [CrossRef]
- Wong, B.; Lumma, W.C.; Smith, A.M.; Sisko, J.T.; Wright, S.D.; Cai, T.Q. Statins suppress THP-1 cell migration and secretion of matrix metalloproteinase 9 by inhibiting geranylgeranylation. J. Leukoc. Biol. 2001, 69, 959–962. [Google Scholar]
- Akula, M.K.; Shi, M.; Jiang, Z.; Foster, C.E.; Miao, D.; Li, A.S.; Zhang, X.; Gavin, R.M.; Forde, S.D.; Germain, G.; et al. Control of the innate immune response by the mevalonate pathway. Nat. Immunol. 2016, 17, 922–929. [Google Scholar] [CrossRef]
- Muller, A.L.; Freed, D.H. Basic and Clinical Observations of Mevalonate Depletion on the Mevalonate Signaling Pathway. Curr. Mol. Pharmacol. 2017, 10, 6–12. [Google Scholar] [CrossRef]
- Tseng, H.C.; Kanayama, K.; Kaur, K.; Park, S.H.; Park, S.; Kozlowska, A.; Sun, S.; McKenna, C.E.; Nishimura, I.; Jewett, A. Bisphosphonate-induced differential modulation of immune cell function in gingiva and bone marrow in vivo: Role in osteoclast-mediated NK cell activation. Oncotarget 2015, 6, 20002–20025. [Google Scholar] [CrossRef] [PubMed]
- Awad, F.; Assrawi, E.; Jumeau, C.; Georgin-Lavialle, S.; Cobret, L.; Duquesnoy, P.; Piterboth, W.; Thomas, L.; Stankovic-Stojanovic, K.; Louvrier, C.; et al. Impact of human monocyte and macrophage polarization on NLR expression and NLRP3 inflammasome activation. PLoS ONE 2017, 12, e0175336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.C.; Li, Z.; Xu, W.; Xiang, C.H.; Ma, Y.F. Luteolin alleviates NLRP3 inflammasome activation and directs macrophage polarization in lipopolysaccharide-stimulated RAW264.7 cells. Am. J. Transl. Res. 2018, 10, 265–273. [Google Scholar]
- Ferrari, S.; Lewiecki, E.M.; Butler, P.W.; Kendler, D.L.; Napoli, N.; Huang, S.; Crittenden, D.B.; Pannacciulli, N.; Siris, E.; Binkley, N. Favorable skeletal benefit/risk of long-term denosumab therapy: A virtual-twin analysis of fractures prevented relative to skeletal safety events observed. Bone 2020, 134, 115287. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Bisphosphonates | Cellular Activity | Polarization | Protein Expression |
|---|---|---|---|
| THP1 cells | (↓) viability [33,52,53] (↓) migration [54] (↓) phagocytosis [54] | (↑) M1 (–) M2 [55] (↑) M1 (↓) M2 [16] | (↑) MMP9 (–) MMP2 [52] |
| RAW264.7 cells | (↓) viability [56] (↑) apoptosis [57] (↓) migration [58] | (↑) IL1 [59,60] (↑) (IL1b, IL6, TNFα,NO) [40] | |
| BMDMs | (↑) M1 (↓) M2 [16] | (↑) (IL1b, IL6, TNFα) [61] (↑) IL1b [62] (↑) IL6 [63] | |
| Mouse MRONJ models | (↑) M1 [16] (↑) M1\(↓) M2 [22] (↓) M2 [64] | (↑) (IL6, IL1b, caspase1) [60,62] | |
| Human MRONJ tissue | (↓) differentiation [34] | (↑) M1 (↓) M2 [35] (↑) M1 [65] | (↑) IL6 (↓) IL10 [35] |
| Denosumab/ anti-RANKL | |||
| THP1 cells | (–) viability [33] | ||
| BMDMs | (–) viability [66] (–) number [67] | (–) IL10 (–)TGFb [67] | |
| Mouse MRONJ models | (↓) wound healing [66,67] (–) differentiation [68,69] (↓) functionality [70] | (–) M1 (–) M2 [64] (↑) M1 (↑↑) * M2 [67] | |
| Human MRONJ tissue | (↑) M1 (↓) M2 [35] | (↑) IL6 (↓) IL10 [35] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gkouveris, I.; Soundia, A.; Gouveris, P.; Zouki, D.; Hadaya, D.; Tetradis, S. Macrophage Involvement in Medication-Related Osteonecrosis of the Jaw (MRONJ): A Comprehensive, Short Review. Cancers 2022, 14, 330. https://doi.org/10.3390/cancers14020330
Gkouveris I, Soundia A, Gouveris P, Zouki D, Hadaya D, Tetradis S. Macrophage Involvement in Medication-Related Osteonecrosis of the Jaw (MRONJ): A Comprehensive, Short Review. Cancers. 2022; 14(2):330. https://doi.org/10.3390/cancers14020330
Chicago/Turabian StyleGkouveris, Ioannis, Akrivoula Soundia, Panagiotis Gouveris, Dionysia Zouki, Danny Hadaya, and Sotirios Tetradis. 2022. "Macrophage Involvement in Medication-Related Osteonecrosis of the Jaw (MRONJ): A Comprehensive, Short Review" Cancers 14, no. 2: 330. https://doi.org/10.3390/cancers14020330