
Insights from IgE Immune Surveillance in Allergy and Cancer for Anti-Tumour IgE Treatments


 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Epidemiological Evidence Underlying IgE Immune Surveillance and Links with Cancer
2.1. The Four Hypotheses of IgE and Cancer
2.2. Chronic Inflammation and Cancer Risks Presented by Chronic Allergic Diseases
2.3. The Involvement of Prophylaxis and Th2 Skewing
3. Roles of IgE in Tumour Immunosurveillance
3.1. IgE and Immunosurveillance: Illustrating the Potential for IgE-Based Therapeutics
3.2. Impact of IgE Levels on Malignancy Risk
4. The Combinatorial Hypothesis: Addressing the Gaps
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Coulson, A.; Levy, A.; Gossell-Williams, M. Monoclonal Antibodies in Cancer Therapy: Mechanisms, Successes and Limitations. West Indian Med. J. 2014, 63, 650–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, L.M.; Veeramani, S.; Weiner, G.J. Complement in monoclonal antibody therapy of cancer. Immunol. Res. 2014, 59, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Grilo, A.L.; Mantalaris, A. The Increasingly Human and Profitable Monoclonal Antibody Market. Trends Biotechnol. 2019, 37, 9–16. [Google Scholar] [CrossRef] [Green Version]
- The Antibody Society. Therapeutic Monoclonal Antibodies Approved or in Review in the EU or US. Available online: https://www.antibodysociety.org/resources/approved-antibodies/ (accessed on 11 June 2021).
- Weiner, L.M.; Surana, R.; Wang, S. Monoclonal antibodies: Versatile platforms for cancer immunotherapy. Nat. Rev. Immunol. 2010, 10, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, A.M.; Wolchok, J.D.; Old, L.J. Antibody therapy of cancer. Nat. Rev. Cancer 2012, 12, 278–287. [Google Scholar] [CrossRef]
- Bakema, J.E.; van Egmond, M. Fc receptor-dependent mechanisms of monoclonal antibody therapy of cancer. Curr. Top. Microbiol. Immunol. 2014, 382, 373–392. [Google Scholar] [CrossRef]
- Cruz, E.; Kayser, V. Monoclonal antibody therapy of solid tumors: Clinical limitations and novel strategies to enhance treatment efficacy. Biologics 2019, 13, 33–51. [Google Scholar] [CrossRef] [Green Version]
- DiLillo, D.J.; Ravetch, J.V. Differential Fc-Receptor Engagement Drives an Anti-tumor Vaccinal Effect. Cell 2015, 161, 1035–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasser, M.; Waaga-Gasser, A.M. Therapeutic Antibodies in Cancer Therapy. Adv. Exp. Med. Biol. 2016, 917, 95–120. [Google Scholar] [CrossRef]
- Roghanian, A.; Teige, I.; Mårtensson, L.; Cox, K.L.; Kovacek, M.; Ljungars, A.; Mattson, J.; Sundberg, A.; Vaughan, A.T.; Shah, V.; et al. Antagonistic human FcγRIIB (CD32B) antibodies have anti-tumor activity and overcome resistance to antibody therapy in vivo. Cancer Cell 2015, 27, 473–488. [Google Scholar] [CrossRef] [Green Version]
- Gould, H.J.; Sutton, B.J.; Beavil, A.J.; Beavil, R.L.; McCloskey, N.; Coker, H.A.; Fear, D.; Smurthwaite, L. The biology of IGE and the basis of allergic disease. Annu. Rev. Immunol. 2003, 21, 579–628. [Google Scholar] [CrossRef]
- Engeroff, P.; Caviezel, F.; Mueller, D.; Thoms, F.; Bachmann, M.F.; Vogel, M. CD23 provides a noninflammatory pathway for IgE-allergen complexes. J. Allergy Clin. Immunol. 2020, 145, 301–311.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen-Jarolim, E.; Bax, H.J.; Bianchini, R.; Capron, M.; Corrigan, C.; Castells, M.; Dombrowicz, D.; Daniels-Wells, T.R.; Fazekas, J.; Fiebiger, E.; et al. AllergoOncology—The impact of allergy in oncology: EAACI position paper. Allergy 2017, 72, 866–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, J.; McCraw, A.J.; Nakamura, M.; Osborn, G.; Sow, H.S.; Cox, V.F.; Stavraka, C.; Josephs, D.H.; Spicer, J.F.; Karagiannis, S.N.; et al. IgE Antibodies against Cancer: Efficacy and Safety. Antibodies 2020, 9, 55. [Google Scholar] [CrossRef]
- Sutton, B.J.; Davies, A.M.; Bax, H.J.; Karagiannis, S.N. IgE Antibodies: From Structure to Function and Clinical Translation. Antibodies 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leoh, L.S.; Daniels-Wells, T.R.; Penichet, M.L. IgE immunotherapy against cancer. Curr. Top. Microbiol. Immunol. 2015, 388, 109–149. [Google Scholar] [CrossRef] [Green Version]
- Karagiannis, P.; Singer, J.; Hunt, J.; Gan, S.K.; Rudman, S.M.; Mechtcheriakova, D.; Knittelfelder, R.; Daniels, T.R.; Hobson, P.S.; Beavil, A.J.; et al. Characterisation of an engineered trastuzumab IgE antibody and effector cell mechanisms targeting HER2/neu-positive tumour cells. Cancer Immunol. Immunother. 2009, 58, 915–930. [Google Scholar] [CrossRef] [Green Version]
- Josephs, D.H.; Bax, H.J.; Dodev, T.; Georgouli, M.; Nakamura, M.; Pellizzari, G.; Saul, L.; Karagiannis, P.; Cheung, A.; Herraiz, C.; et al. Anti-Folate Receptor-α IgE but not IgG Recruits Macrophages to Attack Tumors via TNFα/MCP-1 Signaling. Cancer Res. 2017, 77, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Ure, D.M. Negative assoication between allergy and cancer. Scott. Med. J. 1969, 14, 51–54. [Google Scholar] [CrossRef]
- Augustin, R.; Chandradasa, K.D. IgE levels and allergic skin reactions in cancer and non-cancer patients. Int. Arch. Allergy Appl. Immunol. 1971, 41, 141–143. [Google Scholar] [CrossRef]
- McCormick, D.P.; Ammann, A.J.; Ishizaka, K.; Miller, D.G.; Hong, R. A study of allergy in patients with malignant lymphoma and chronic lymphocytic leukemia. Cancer 1971, 27, 93–99. [Google Scholar] [CrossRef]
- Allegra, J.; Lipton, A.; Harvey, H.; Luderer, J.; Brenner, D.; Mortel, R.; Demers, L.; Gillin, M.; White, D.; Trautlein, J. Decreased prevalence of immediate hypersensitivity (atopy) in a cancer population. Cancer Res. 1976, 36, 3225–3226. [Google Scholar]
- Nagy, E.; Berczi, I.; Sehon, A.H. Growth inhibition of murine mammary carcinoma by monoclonal IgE antibodies specific for the mammary tumor virus. Cancer Immunol. Immunother. 1991, 34, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, M.H.; Darcy, P.K.; Trapani, J.A.; MacGregor, D.; Smyth, M.J. Tumor-specific IgE-mediated inhibition of human colorectal carcinoma xenograft growth. Oncol. Res. 1998, 10, 133–142. [Google Scholar] [PubMed]
- Karagiannis, S.N.; Bracher, M.G.; Hunt, J.; McCloskey, N.; Beavil, R.L.; Beavil, A.J.; Fear, D.J.; Thompson, R.G.; East, N.; Burke, F.; et al. IgE-antibody-dependent immunotherapy of solid tumors: Cytotoxic and phagocytic mechanisms of eradication of ovarian cancer cells. J. Immunol. 2007, 179, 2832–2843. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.L.; Pierre, J.; Smith-Norowitz, T.A.; Hagler, M.; Bowne, W.; Pincus, M.R.; Mueller, C.M.; Zenilman, M.E.; Bluth, M.H. Immunoglobulin E antibodies from pancreatic cancer patients mediate antibody-dependent cell-mediated cytotoxicity against pancreatic cancer cells. Clin. Exp. Immunol. 2008, 153, 401–409. [Google Scholar] [CrossRef]
- Karagiannis, S.N.; Bracher, M.G.; Beavil, R.L.; Beavil, A.J.; Hunt, J.; McCloskey, N.; Thompson, R.G.; East, N.; Burke, F.; Sutton, B.J.; et al. Role of IgE receptors in IgE antibody-dependent cytotoxicity and phagocytosis of ovarian tumor cells by human monocytic cells. Cancer Immunol. Immunother. 2008, 57, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Daniels-Wells, T.R.; Helguera, G.; Leuchter, R.K.; Quintero, R.; Kozman, M.; Rodríguez, J.A.; Ortiz-Sánchez, E.; Martínez-Maza, O.; Schultes, B.C.; Nicodemus, C.F.; et al. A novel IgE antibody targeting the prostate-specific antigen as a potential prostate cancer therapy. BMC Cancer 2013, 13, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, E.A.; Brini, A.T.; Yenagi, V.A.; Ferreira, L.M.; Achatz-Straussberger, G.; Ambrosi, A.; Sanvito, F.; Soprana, E.; van Anken, E.; Achatz, G.; et al. Cutting Edge: IgE Plays an Active Role in Tumor Immunosurveillance in Mice. J. Immunol. 2016, 197, 2583–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellizzari, G.; Hoskin, C.; Crescioli, S.; Mele, S.; Gotovina, J.; Chiaruttini, G.; Bianchini, R.; Ilieva, K.; Bax, H.J.; Papa, S.; et al. IgE re-programs alternatively-activated human macrophages towards pro-inflammatory anti-tumoural states. EBioMedicine 2019, 43, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Van Hemelrijck, M.; Garmo, H.; Binda, E.; Hayday, A.; Karagiannis, S.N.; Hammar, N.; Walldius, G.; Lambe, M.; Jungner, I.; Holmberg, L. Immunoglobulin E and cancer: A meta-analysis and a large Swedish cohort study. Cancer Causes Control. 2010, 21, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Wulaningsih, W.; Holmberg, L.; Garmo, H.; Karagiannis, S.N.; Ahlstedt, S.; Malmstrom, H.; Lambe, M.; Hammar, N.; Walldius, G.; Jungner, I.; et al. Investigating the association between allergen-specific immunoglobulin E, cancer risk and survival. Oncoimmunology 2016, 5, e1154250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josephs, D.H.; Spicer, J.F.; Corrigan, C.J.; Gould, H.J.; Karagiannis, S.N. Epidemiological associations of allergy, IgE and cancer. Clin. Exp. Allergy 2013, 43, 1110–1123. [Google Scholar] [CrossRef]
- Liao, H.C.; Wu, S.Y.; Ou, C.Y.; Hsiao, J.R.; Huang, J.S.; Tsai, S.T.; Huang, C.C.; Wong, T.Y.; Lee, W.T.; Chen, K.C.; et al. Allergy symptoms, serum total immunoglobulin E, and risk of head and neck cancer. Cancer Causes Control. 2016, 27, 1105–1115. [Google Scholar] [CrossRef]
- Hayes, M.D.; Ward, S.; Crawford, G.; Seoane, R.C.; Jackson, W.D.; Kipling, D.; Voehringer, D.; Dunn-Walters, D.; Strid, J. Inflammation-induced IgE promotes epithelial hyperplasia and tumour growth. eLife 2020, 9, e51862. [Google Scholar] [CrossRef]
- Karim, A.F.; Westenberg, L.E.H.; Eurelings, L.E.M.; Otten, R.; Gerth van Wijk, R. The association between allergic diseases and cancer: A systematic review of the literature. Neth. J. Med. 2019, 77, 42–66. [Google Scholar]
- Wang, L.; Bierbrier, R.; Drucker, A.M.; Chan, A.W. Noncutaneous and Cutaneous Cancer Risk in Patients with Atopic Dermatitis: A Systematic Review and Meta-analysis. JAMA Dermatol 2020, 156, 158–171. [Google Scholar] [CrossRef]
- Kantor, E.D.; Hsu, M.; Du, M.; Signorello, L.B. Allergies and Asthma in Relation to Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, E.; Kaprio, J.; Vesterinen, E.; Koskenvuo, M. Mortality of adults with asthma: A prospective cohort study. Thorax 1997, 52, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Vesterinen, E.; Pukkala, E.; Timonen, T.; Aromaa, A. Cancer incidence among 78,000 asthmatic patients. Int. J. Epidemiol. 1993, 22, 976–982. [Google Scholar] [CrossRef]
- González-Pérez, A.; Fernández-Vidaurre, C.; Rueda, A.; Rivero, E.; García Rodríguez, L.A. Cancer incidence in a general population of asthma patients. Pharmacoepidemiol. Drug Saf. 2006, 15, 131–138. [Google Scholar] [CrossRef]
- Ji, J.; Shu, X.; Li, X.; Sundquist, K.; Sundquist, J.; Hemminki, K. Cancer risk in hospitalised asthma patients. Br. J. Cancer 2009, 100, 829–833. [Google Scholar] [CrossRef]
- Vandentorren, S.; Baldi, I.; Annesi Maesano, I.; Charpin, D.; Neukirch, F.; Filleul, L.; Cantagrel, A.; Tessier, J.F. Long-term mortality among adults with or without asthma in the PAARC study. Eur. Respir. J. 2003, 21, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Hemminki, K.; Försti, A.; Fallah, M.; Sundquist, J.; Sundquist, K.; Ji, J. Risk of cancer in patients with medically diagnosed hay fever or allergic rhinitis. Int. J. Cancer 2014, 135, 2397–2403. [Google Scholar] [CrossRef] [Green Version]
- Kozłowska, R.; Bożek, A.; Jarząb, J. Association between cancer and allergies. Allergy Asthma Clin. Immunol. 2016, 12, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Zein, M.; Parent, M.E.; Kâ, K.; Siemiatycki, J.; St-Pierre, Y.; Rousseau, M.C. History of asthma or eczema and cancer risk among men: A population-based case-control study in Montreal, Quebec, Canada. Ann. Allergy Asthma Immunol. 2010, 104, 378–384. [Google Scholar] [CrossRef]
- Castaing, M.; Youngson, J.; Zaridze, D.; Szeszenia-Dabrowska, N.; Rudnai, P.; Lissowska, J.; Fabiánová, E.; Mates, D.; Bencko, V.; Foretova, L.; et al. Is the risk of lung cancer reduced among eczema patients? Am. J. Epidemiol. 2005, 162, 542–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberger, A.; Bickeböller, H.; McCormack, V.; Brenner, D.R.; Duell, E.J.; Tjønneland, A.; Friis, S.; Muscat, J.E.; Yang, P.; Wichmann, H.E.; et al. Asthma and lung cancer risk: A systematic investigation by the International Lung Cancer Consortium. Carcinogenesis 2012, 33, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Rittmeyer, D.; Lorentz, A. Relationship between allergy and cancer: An overview. Int. Arch. Allergy Immunol. 2012, 159, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.Y.; Chen, Y.J.; Lin, M.W.; Chen, T.J.; Chu, S.Y.; Chen, C.C.; Lee, D.D.; Chang, Y.T.; Wang, W.J.; Liu, H.N. Cancer risk in patients with allergic rhinitis, asthma and atopic dermatitis: A nationwide cohort study in Taiwan. Int. J. Cancer 2012, 130, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.C.; Chen, Y.; Krewski, D.; Ghadirian, P.; Thun, M.J.; Calle, E.E. Cancer mortality among US men and women with asthma and hay fever. Am. J. Epidemiol. 2005, 162, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Linos, E.; Raine, T.; Alonso, A.; Michaud, D. Atopy and risk of brain tumors: A meta-analysis. J. Natl. Cancer Inst. 2007, 99, 1544–1550. [Google Scholar] [CrossRef] [Green Version]
- Amirian, E.S.; Zhou, R.; Wrensch, M.R.; Olson, S.H.; Scheurer, M.E.; Il’yasova, D.; Lachance, D.; Armstrong, G.N.; McCoy, L.S.; Lau, C.C.; et al. Approaching a Scientific Consensus on the Association between Allergies and Glioma Risk: A Report from the Glioma International Case-Control Study. Cancer Epidemiol. Biomark. Prev. 2016, 25, 282–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana, A.; Wentworth, C.E.; Fernández-Vidaurre, C.; Schlienger, R.G.; Conde, E.; Arellano, F.M. Incidence of cancer in the general population and in patients with or without atopic dermatitis in the U.K. Br. J. Dermatol. 2010, 163, 1036–1043. [Google Scholar] [CrossRef]
- Talbot-Smith, A.; Fritschi, L.; Divitini, M.L.; Mallon, D.F.; Knuiman, M.W. Allergy, atopy, and cancer: A prospective study of the 1981 Busselton cohort. Am. J. Epidemiol. 2003, 157, 606–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandini, S.; Stanganelli, I.; Palli, D.; De Giorgi, V.; Masala, G.; Caini, S. Atopic dermatitis, naevi count and skin cancer risk: A meta-analysis. J. Dermatol. Sci. 2016, 84, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.O.; Svaerke, C.; Körmendiné Farkas, D.; Olesen, A.B.; Kragballe, K.; Sørensen, H.T. Atopic dermatitis and risk of skin cancer: A Danish nationwide cohort study (1977–2006). Am. J. Clin. Dermatol. 2012, 13, 29–36. [Google Scholar] [CrossRef]
- Wiemels, J.L.; Wiencke, J.K.; Li, Z.; Ramos, C.; Nelson, H.H.; Karagas, M.R. Risk of squamous cell carcinoma of the skin in relation to IgE: A nested case-control study. Cancer Epidemiol. Biomark. Prev. 2011, 20, 2377–2383. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Zens, M.S.; Duell, E.; Perry, A.E.; Chapman, M.S.; Karagas, M.R. History of allergy and atopic dermatitis in relation to squamous cell and Basal cell carcinoma of the skin. Cancer Epidemiol. Biomark. Prev. 2015, 24, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Deckert, S.; Kopkow, C.; Schmitt, J. Nonallergic comorbidities of atopic eczema: An overview of systematic reviews. Allergy 2014, 69, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halling-Overgaard, A.S.; Ravnborg, N.; Silverberg, J.I.; Egeberg, A.; Thyssen, J.P. Atopic dermatitis and cancer in solid organs: A systematic review and meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2019, 33, e81–e82. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, C.; Talamini, R.; Franceschi, S.; Negri, E.; Giacosa, A.; La Vecchia, C. Allergy and the risk of selected digestive and laryngeal neoplasms. Eur. J. Cancer Prev. 2004, 13, 173–176. [Google Scholar] [CrossRef]
- Fereidouni, M.; Ferns, G.A.; Bahrami, A. Current status and perspectives regarding the association between allergic disorders and cancer. IUBMB Life 2020, 72, 1322–1339. [Google Scholar] [CrossRef]
- Zhao, H.; Cai, W.; Su, S.; Zhi, D.; Lu, J.; Liu, S. Allergic conditions reduce the risk of glioma: A meta-analysis based on 128,936 subjects. Tumour. Biol. 2014, 35, 3875–3880. [Google Scholar] [CrossRef]
- Amirian, E.S.; Marquez-Do, D.; Bondy, M.L.; Scheurer, M.E. Antihistamine use and immunoglobulin E levels in glioma risk and prognosis. Cancer Epidemiol. 2013, 37, 908–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheurer, M.E.; Amirian, E.S.; Davlin, S.L.; Rice, T.; Wrensch, M.; Bondy, M.L. Effects of antihistamine and anti-inflammatory medication use on risk of specific glioma histologies. Int. J. Cancer 2011, 129, 2290–2296. [Google Scholar] [CrossRef] [Green Version]
- Schlehofer, B.; Blettner, M.; Preston-Martin, S.; Niehoff, D.; Wahrendorf, J.; Arslan, A.; Ahlbom, A.; Choi, W.N.; Giles, G.G.; Howe, G.R.; et al. Role of medical history in brain tumour development. Results from the international adult brain tumour study. Int. J. Cancer 1999, 82, 155–160. [Google Scholar] [CrossRef]
- McCarthy, B.J.; Rankin, K.; Il’yasova, D.; Erdal, S.; Vick, N.; Ali-Osman, F.; Bigner, D.D.; Davis, F. Assessment of type of allergy and antihistamine use in the development of glioma. Cancer Epidemiol. Biomark. Prev. 2011, 20, 370–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, D.S.; Church, M.K. Pharmacology of Antihistamines. World Allergy Organ. J. 2011, 4, S22–S27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tepper, R.I.; Coffman, R.L.; Leder, P. An eosinophil-dependent mechanism for the antitumor effect of interleukin-4. Science 1992, 257, 548–551. [Google Scholar] [CrossRef]
- Saleh, M.; Wiegmans, A.; Malone, Q.; Stylli, S.S.; Kaye, A.H. Effect of in situ retroviral interleukin-4 transfer on established intracranial tumors. J. Natl. Cancer Inst. 1999, 91, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, H.; Villa, L.; Attanucci, J.; Erff, M.; Fellows, W.K.; Lotze, M.T.; Pollack, I.F.; Chambers, W.H. Cytokine gene therapy of gliomas: Effective induction of therapeutic immunity to intracranial tumors by peripheral immunization with interleukin-4 transduced glioma cells. Gene Ther. 2001, 8, 1157–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedetti, S.; Pirola, B.; Pollo, B.; Magrassi, L.; Bruzzone, M.G.; Rigamonti, D.; Galli, R.; Selleri, S.; Di Meco, F.; De Fraja, C.; et al. Gene therapy of experimental brain tumors using neural progenitor cells. Nat. Med. 2000, 6, 447–450. [Google Scholar] [CrossRef]
- Schwartzbaum, J.; Seweryn, M.; Holloman, C.; Harris, R.; Handelman, S.K.; Rempala, G.A.; Huang, R.P.; Burkholder, B.; Brandemihl, A.; Kallberg, H.; et al. Association between Prediagnostic Allergy-Related Serum Cytokines and Glioma. PLoS ONE 2015, 10, e0137503. [Google Scholar] [CrossRef]
- Gohar, M.K.; Ammar, M.G.; Alnagar, A.A.; Abd-ElAziz, H.A. Serum IgE and Allergy Related Genotypes of IL-4R α and IL-13 Genes: Association with Glioma Susceptibility and Glioblastoma Prognosis. Egypt J. Immunol. 2018, 25, 19–33. [Google Scholar]
- Jin, T.B.; Du, S.; Zhu, X.K.; Li, G.; Ouyang, Y.; He, N.; Zhang, Z.; Zhang, Y.; Kang, L.; Yuan, D. Polymorphism in the IL4R gene and clinical features are associated with glioma prognosis: Analyses of case-cohort studies. Medicine 2016, 95, e4231. [Google Scholar] [CrossRef] [PubMed]
- Calboli, F.C.; Cox, D.G.; Buring, J.E.; Gaziano, J.M.; Ma, J.; Stampfer, M.; Willett, W.C.; Tworoger, S.S.; Hunter, D.J.; Camargo, C.A., Jr.; et al. Prediagnostic plasma IgE levels and risk of adult glioma in four prospective cohort studies. J. Natl. Cancer Inst. 2011, 103, 1588–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, G.; Hayes, M.D.; Seoane, R.C.; Ward, S.; Dalessandri, T.; Lai, C.; Healy, E.; Kipling, D.; Proby, C.; Moyes, C.; et al. Epithelial damage and tissue γδ T cells promote a unique tumor-protective IgE response. Nat. Immunol. 2018, 19, 859–870. [Google Scholar] [CrossRef]
- McCoy, K.D.; Harris, N.L.; Diener, P.; Hatak, S.; Odermatt, B.; Hangartner, L.; Senn, B.M.; Marsland, B.J.; Geuking, M.B.; Hengartner, H.; et al. Natural IgE production in the absence of MHC Class II cognate help. Immunity 2006, 24, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Palm, N.W.; Rosenstein, R.K.; Medzhitov, R. Allergic host defences. Nature 2012, 484, 465–472. [Google Scholar] [CrossRef]
- Daniels, T.R.; Leuchter, R.K.; Quintero, R.; Helguera, G.; Rodríguez, J.A.; Martínez-Maza, O.; Schultes, B.C.; Nicodemus, C.F.; Penichet, M.L. Targeting HER2/neu with a fully human IgE to harness the allergic reaction against cancer cells. Cancer Immunol. Immunother. 2012, 61, 991–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagiannis, S.N.; Josephs, D.H.; Karagiannis, P.; Gilbert, A.E.; Saul, L.; Rudman, S.M.; Dodev, T.; Koers, A.; Blower, P.J.; Corrigan, C.; et al. Recombinant IgE antibodies for passive immunotherapy of solid tumours: From concept towards clinical application. Cancer Immunol. Immunother. 2012, 61, 1547–1564. [Google Scholar] [CrossRef]
- Staff, C.; Magnusson, C.G.; Hojjat-Farsangi, M.; Mosolits, S.; Liljefors, M.; Frödin, J.E.; Wahrén, B.; Mellstedt, H.; Ullenhag, G.J. Induction of IgM, IgA and IgE antibodies in colorectal cancer patients vaccinated with a recombinant CEA protein. J. Clin. Immunol. 2012, 32, 855–865. [Google Scholar] [CrossRef]
- Saul, L.; Josephs, D.H.; Cutler, K.; Bradwell, A.; Karagiannis, P.; Selkirk, C.; Gould, H.J.; Jones, P.; Spicer, J.F.; Karagiannis, S.N. Comparative reactivity of human IgE to cynomolgus monkey and human effector cells and effects on IgE effector cell potency. MAbs 2014, 6, 509–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, M.; Souri, E.A.; Osborn, G.; Laddach, R.; Chauhan, J.; Stavraka, C.; Lombardi, S.; Black, A.; Khiabany, A.; Khair, D.O.; et al. IgE Activates Monocytes from Cancer Patients to Acquire a Pro-Inflammatory Phenotype. Cancers 2020, 12, 3376. [Google Scholar] [CrossRef] [PubMed]
- Olteanu, H.; Fenske, T.S.; Harrington, A.M.; Szabo, A.; He, P.; Kroft, S.H. CD23 expression in follicular lymphoma: Clinicopathologic correlations. Am. J. Clin. Pathol. 2011, 135, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linderoth, J.; Jerkeman, M.; Cavallin-Ståhl, E.; Kvaløy, S.; Torlakovic, E. Immunohistochemical expression of CD23 and CD40 may identify prognostically favorable subgroups of diffuse large B-cell lymphoma: A Nordic Lymphoma Group Study. Clin. Cancer Res. 2003, 9, 722–728. [Google Scholar]
- Ye, Z.S.; Fan, L.N.; Wang, L.; Yang, B.; Wang, L.F.; Liu, Y.S.; Ji, S.N.; Xu, H.Z.; Xiao, C.X. Immunoglobulin E induces colon cancer cell apoptosis via enhancing cyp27b1 expression. Am. J. Transl. Res. 2016, 8, 5715–5722. [Google Scholar]
- Spillner, E.; Plum, M.; Blank, S.; Miehe, M.; Singer, J.; Braren, I. Recombinant IgE antibody engineering to target EGFR. Cancer Immunol. Immunother. 2012, 61, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Coney, L.R.; Tomassetti, A.; Carayannopoulos, L.; Frasca, V.; Kamen, B.A.; Colnaghi, M.I.; Zurawski, V.R., Jr. Cloning of a tumor-associated antigen: MOv18 and MOv19 antibodies recognize a folate-binding protein. Cancer Res. 1991, 51, 6125–6132. [Google Scholar]
- Karagiannis, S.N.; Wang, Q.; East, N.; Burke, F.; Riffard, S.; Bracher, M.G.; Thompson, R.G.; Durham, S.R.; Schwartz, L.B.; Balkwill, F.R.; et al. Activity of human monocytes in IgE antibody-dependent surveillance and killing of ovarian tumor cells. Eur. J. Immunol. 2003, 33, 1030–1040. [Google Scholar] [CrossRef]
- Pellizzari, G.; Bax, H.J.; Josephs, D.H.; Gotovina, J.; Jensen-Jarolim, E.; Spicer, J.F.; Karagiannis, S.N. Harnessing Therapeutic IgE Antibodies to Re-educate Macrophages against Cancer. Trends Mol. Med. 2020, 26, 615–626. [Google Scholar] [CrossRef]
- Karagiannis, S.N.; Josephs, D.H.; Bax, H.J.; Spicer, J.F. Therapeutic IgE Antibodies: Harnessing a Macrophage-Mediated Immune Surveillance Mechanism against Cancer. Cancer Res. 2017, 77, 2779–2783. [Google Scholar] [CrossRef] [Green Version]
- Spicer, J.; Basu, B.; Montes, A.; Banerji, U.; Kristeleit, R.; Veal, G.J.; Corrigan, C.; Till, S.; Nintos, G.; Brier, T.; et al. Phase 1 Trial of MOv18, a First-in-Class IgE Antibody Therapy for Cancer. Available online: https://www.abstractsonline.com/pp8/#!/9045/presentation/10640,%20April%2027-29,%202020 (accessed on 2 August 2020).
- Nieters, A.; Łuczyńska, A.; Becker, S.; Becker, N.; Vermeulen, R.; Overvad, K.; Aleksandrova, K.; Boeing, H.; Lagiou, P.; Trichopoulos, D.; et al. Prediagnostic immunoglobulin E levels and risk of chronic lymphocytic leukemia, other lymphomas and multiple myeloma-results of the European Prospective Investigation into Cancer and Nutrition. Carcinogenesis 2014, 35, 2716–2722. [Google Scholar] [CrossRef] [Green Version]
- Wiemels, J.L.; Wilson, D.; Patil, C.; Patoka, J.; McCoy, L.; Rice, T.; Schwartzbaum, J.; Heimberger, A.; Sampson, J.H.; Chang, S.; et al. IgE, allergy, and risk of glioma: Update from the San Francisco Bay Area Adult Glioma Study in the temozolomide era. Int. J. Cancer 2009, 125, 680–687. [Google Scholar] [CrossRef]
- Wiemels, J.L.; Wiencke, J.K.; Patoka, J.; Moghadassi, M.; Chew, T.; McMillan, A.; Miike, R.; Barger, G.; Wrensch, M. Reduced immunoglobulin E and allergy among adults with glioma compared with controls. Cancer Res. 2004, 64, 8468–8473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matta, G.M.; Battaglio, S.; Dibello, C.; Napoli, P.; Baldi, C.; Ciccone, G.; Coscia, M.; Boccadoro, M.; Massaia, M. Polyclonal immunoglobulin E levels are correlated with hemoglobin values and overall survival in patients with multiple myeloma. Clin. Cancer Res. 2007, 13, 5348–5354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrensch, M.; Wiencke, J.K.; Wiemels, J.; Miike, R.; Patoka, J.; Moghadassi, M.; McMillan, A.; Kelsey, K.T.; Aldape, K.; Lamborn, K.R.; et al. Serum IgE, tumor epidermal growth factor receptor expression, and inherited polymorphisms associated with glioma survival. Cancer Res. 2006, 66, 4531–4541. [Google Scholar] [CrossRef] [Green Version]
- Singer, J.; Achatz-Straussberger, G.; Bentley-Lukschal, A.; Fazekas-Singer, J.; Achatz, G.; Karagiannis, S.N.; Jensen-Jarolim, E. AllergoOncology: High innate IgE levels are decisive for the survival of cancer-bearing mice. World Allergy Organ. J. 2019, 12, 100044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlehofer, B.; Siegmund, B.; Linseisen, J.; Schüz, J.; Rohrmann, S.; Becker, S.; Michaud, D.; Melin, B.; Bas Bueno-de-Mesquita, H.; Peeters, P.H.; et al. Primary brain tumours and specific serum immunoglobulin E: A case-control study nested in the European Prospective Investigation into Cancer and Nutrition cohort. Allergy 2011, 66, 1434–1441. [Google Scholar] [CrossRef]
- Ferastraoaru, D.; Rosenstreich, D. IgE deficiency and prior diagnosis of malignancy: Results of the 2005–2006 National Health and Nutrition Examination Survey. Ann. Allergy Asthma Immunol. 2018, 121, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Ferastraoaru, D.; Rosenstreich, D. IgE deficiency is associated with high rates of new malignancies: Results of a longitudinal cohort study. J. Allergy Clin. Immunol. Pract. 2020, 8, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Ferastraoaru, D.; Schwartz, D.; Rosenstreich, D. Increased Malignancy Rate in Children With IgE Deficiency: A Single-center Experience. J. Pediatr. Hematol. Oncol. 2021, 43, e472–e477. [Google Scholar] [CrossRef]
- Magen, E.; Schlesinger, M.; David, M.; Ben-Zion, I.; Vardy, D. Selective IgE deficiency, immune dysregulation, and autoimmunity. Allergy Asthma Proc. 2014, 35, e27–e33. [Google Scholar] [CrossRef]
- Ellison-Loschmann, L.; Benavente, Y.; Douwes, J.; Buendia, E.; Font, R.; Alvaro, T.; Kogevinas, M.; de Sanjosé, S. Immunoglobulin E levels and risk of lymphoma in a case-control study in Spain. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1492–1498. [Google Scholar] [CrossRef] [Green Version]
- Melbye, M.; Smedby, K.E.; Lehtinen, T.; Rostgaard, K.; Glimelius, B.; Munksgaard, L.; Schöllkopf, C.; Sundström, C.; Chang, E.T.; Koskela, P.; et al. Atopy and risk of non-Hodgkin lymphoma. J. Natl. Cancer Inst. 2007, 99, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferastraoaru, D.; Gross, R.; Rosenstreich, D. Increased malignancy incidence in IgE deficient patients not due to concomitant Common Variable Immunodeficiency. Ann. Allergy Asthma Immunol. 2017, 119, 267–273. [Google Scholar] [CrossRef]
- Nakamura, M.; Bax, H.J.; Scotto, D.; Souri, E.A.; Sollie, S.; Harris, R.J.; Hammar, N.; Walldius, G.; Winship, A.; Ghosh, S.; et al. Immune mediator expression signatures are associated with improved outcome in ovarian carcinoma. Oncoimmunology 2019, 8, e1593811. [Google Scholar] [CrossRef]
- Holgate, S.; Casale, T.; Wenzel, S.; Bousquet, J.; Deniz, Y.; Reisner, C. The anti-inflammatory effects of omalizumab confirm the central role of IgE in allergic inflammation. J. Allergy Clin. Immunol. 2005, 115, 459–465. [Google Scholar] [CrossRef]
- Johnston, A.; Smith, C.; Zheng, C.; Aaron, S.D.; Kelly, S.E.; Skidmore, B.; Wells, G.A. Influence of prolonged treatment with omalizumab on the development of solid epithelial cancer in patients with atopic asthma and chronic idiopathic urticaria: A systematic review and meta-analysis. Clin. Exp. Allergy 2019, 49, 1291–1305. [Google Scholar] [CrossRef]
- Li, J.; Goulding, M.; Seymour, S.; Starke, P. EXCELS study results do not rule out potential cancer risk with omalizumab. J. Allergy Clin. Immunol. 2015, 135, 289. [Google Scholar] [CrossRef]
- Ferastraoaru, D.; Bax, H.J.; Bergmann, C.; Capron, M.; Castells, M.; Dombrowicz, D.; Fiebiger, E.; Gould, H.J.; Hartmann, K.; Jappe, U.; et al. AllergoOncology: Ultra-low IgE, a potential novel biomarker in cancer-a Position Paper of the European Academy of Allergy and Clinical Immunology (EAACI). Clin. Transl. Allergy 2020, 10, 32. [Google Scholar] [CrossRef]
- Prizment, A.E.; Folsom, A.R.; Cerhan, J.R.; Flood, A.; Ross, J.A.; Anderson, K.E. History of allergy and reduced incidence of colorectal cancer, Iowa Women’s Health Study. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2357–2362. [Google Scholar] [CrossRef] [Green Version]
- Eppel, A.; Cotterchio, M.; Gallinger, S. Allergies are associated with reduced pancreas cancer risk: A population-based case-control study in Ontario, Canada. Int. J. Cancer 2007, 121, 2241–2245. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.H.; Hsu, M.; Satagopan, J.M.; Maisonneuve, P.; Silverman, D.T.; Lucenteforte, E.; Anderson, K.E.; Borgida, A.; Bracci, P.M.; Bueno-de-Mesquita, H.B.; et al. Allergies and risk of pancreatic cancer: A pooled analysis from the Pancreatic Cancer Case-Control Consortium. Am. J. Epidemiol. 2013, 178, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Muskens, I.S.; Zhou, M.; McCoy, L.; Bracci, P.M.; Hansen, H.M.; Gauderman, W.J.; Wiencke, J.K.; Wrensch, M.R.; Wiemels, J.L. Immune factors preceding diagnosis of glioma: A Prostate Lung Colorectal Ovarian Cancer Screening Trial nested case-control study. Neurooncol. Adv. 2019, 1, vdz031. [Google Scholar] [CrossRef] [Green Version]
- Savage, N. Searching for the roots of brain cancer. Nature 2018, 561, S50–S51. [Google Scholar] [CrossRef]
- Schwartzbaum, J.; Ding, B.; Johannesen, T.B.; Osnes, L.T.; Karavodin, L.; Ahlbom, A.; Feychting, M.; Grimsrud, T.K. Association between prediagnostic IgE levels and risk of glioma. J. Natl. Cancer Inst. 2012, 104, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Whiteside, S.; Markova, M.; Chin, A.; Lam, C.; Dharmani-Khan, P.; Modi, M.; Khan, F.; Storek, J. Influence of Chemotherapy on Allergen-Specific IgE. Int. Arch. Allergy Immunol. 2018, 177, 145–152. [Google Scholar] [CrossRef]



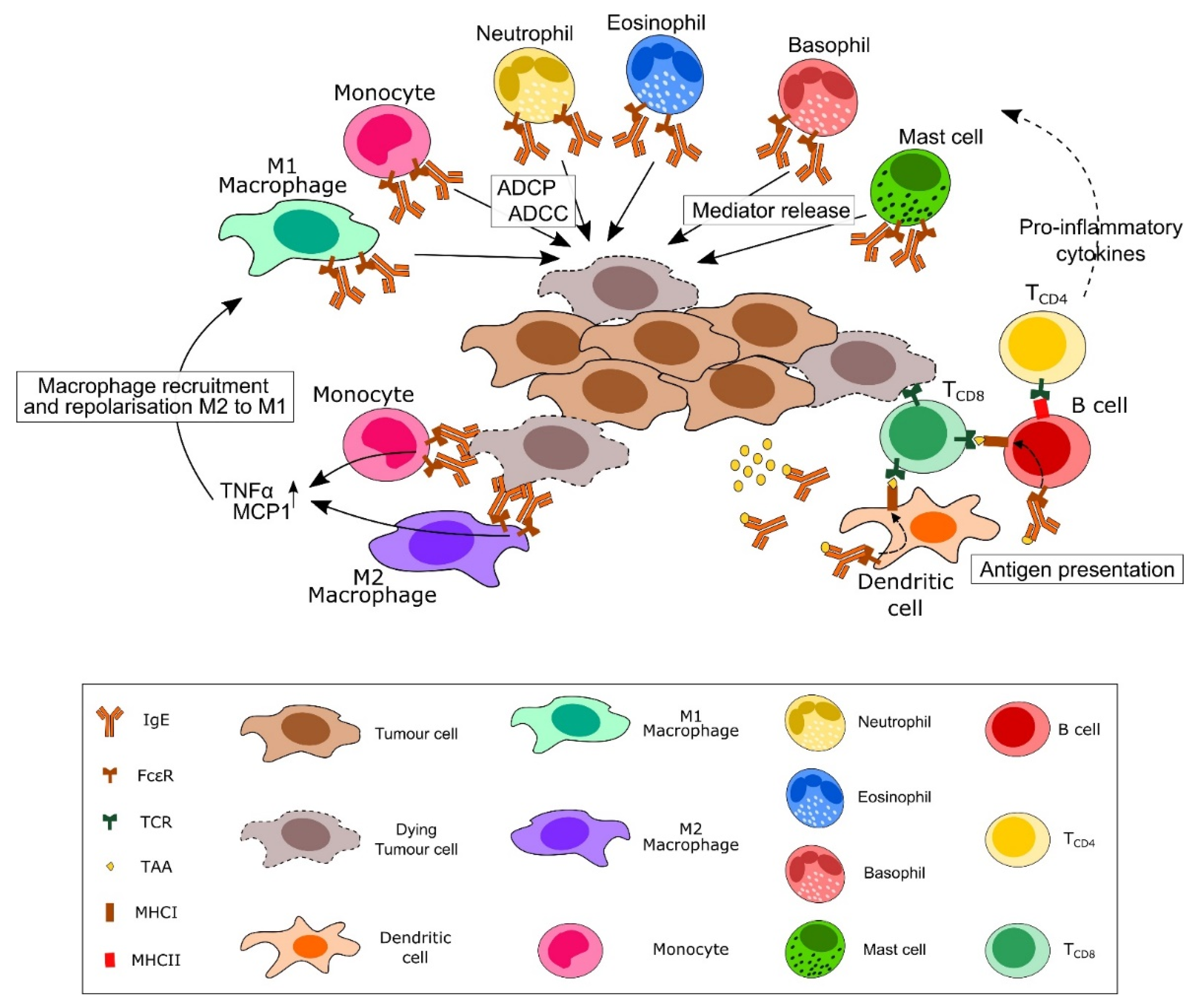{kind=link}
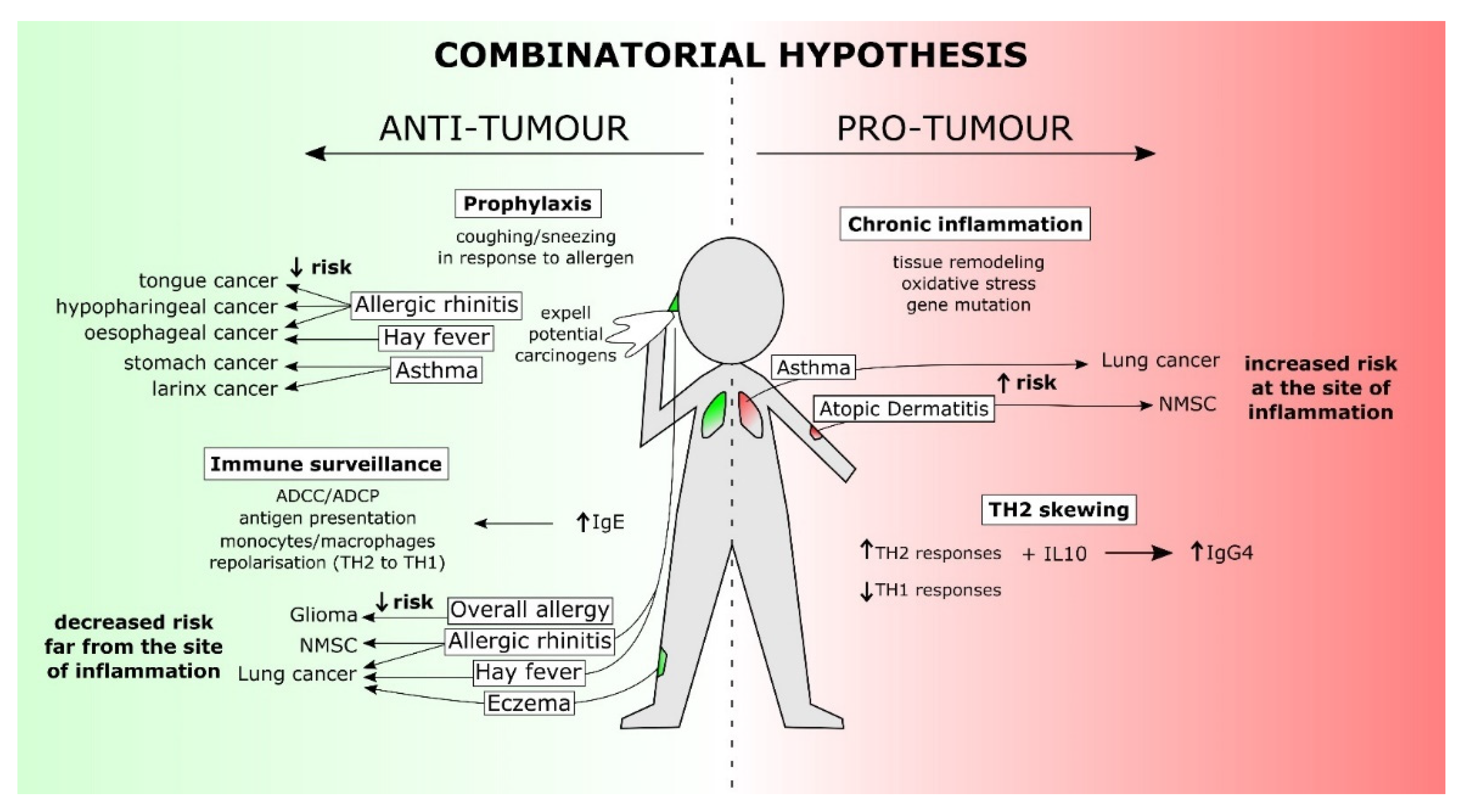{kind=link}
| Hypothesis | Description | Cancer Risk |
|---|---|---|
| Chronic Inflammation [34] | Allergy-induced events including inflammatory cell infiltration, tissue remodelling, and enzyme activation drive mutation of tumour suppressor genes, apoptotic proteins and other factors involved in regulation of cell growth, promoting growth of cancerous cells.Cancer risk will be increased at sites of chronic inflammation. | Increased |
| Immunosurveillance [34] | Allergy reflects general immune hyperresponsiveness; natural immunosurveillance is enhanced. Enhanced immunosurveillance is reflected by high serum IgE, potentially triggered against undiagnosed tumours instead of allergens. IgE/atopy drives activation of powerful effector cells capable of detecting and mounting responses against tumour cells | Decreased |
| Prophylaxis [35] | Physical effects of allergy such as coughing or sneezing act to expel potentially mutagenic or carcinogenic toxins before they can trigger malignancy. Allergy symptoms themselves are beneficial and not necessarily IgE. | Decreased |
| Th2 Immune Skewing [34] | Atopy drives an inappropriate skewing towards T-helper 2 (Th2)-based immune responses, diverting away from potentially tumour-eradicating inflammatory T-helper 1 (Th1) responses. High serum IgE concentrations as a result of Th2 skewing occupy receptors and prevent binding of anti-tumour IgE to Fcε receptor I (FcεRI)-expressing effector cells. Tissues affected by atopy are more sensitive to cancer development due to Th2 skewing. | Increased |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCraw, A.J.; Chauhan, J.; Bax, H.J.; Stavraka, C.; Osborn, G.; Grandits, M.; López-Abente, J.; Josephs, D.H.; Spicer, J.; Wagner, G.K.; et al. Insights from IgE Immune Surveillance in Allergy and Cancer for Anti-Tumour IgE Treatments. Cancers 2021, 13, 4460. https://doi.org/10.3390/cancers13174460
McCraw AJ, Chauhan J, Bax HJ, Stavraka C, Osborn G, Grandits M, López-Abente J, Josephs DH, Spicer J, Wagner GK, et al. Insights from IgE Immune Surveillance in Allergy and Cancer for Anti-Tumour IgE Treatments. Cancers. 2021; 13(17):4460. https://doi.org/10.3390/cancers13174460
Chicago/Turabian StyleMcCraw, Alex J., Jitesh Chauhan, Heather J. Bax, Chara Stavraka, Gabriel Osborn, Melanie Grandits, Jacobo López-Abente, Debra H. Josephs, James Spicer, Gerd K. Wagner, and et al. 2021. "Insights from IgE Immune Surveillance in Allergy and Cancer for Anti-Tumour IgE Treatments" Cancers 13, no. 17: 4460. https://doi.org/10.3390/cancers13174460