
The Soluble Factor from Oral Cancer Cell Lines Inhibits Interferon-γ Production by OK-432 via the CD40/CD40 Ligand Pathway

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Preparation of PBMCs
2.3. Preparation of CM
2.4. In Vitro Simulation Model of the Oral Cancer Patients
2.5. Isolation of Total RNA
2.6. cDNA Microarray
2.7. Quantitative Reverse Transcription–Polymerase Chain Reaction
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Effects of Anti-IL-12 Neutralizing Antibodies (Abs), Recombinant IL-12 (rIL-12), or Anti-CD40 Ab on PBMCs by OK-432 Treatment
2.10. Data Analysis
2.11. Statistical Analysis
3. Results
3.1. Effect of CM on the Cytokine Gene Expressions in PBMCs by OK-432 Treatment (cDNA Microarray)
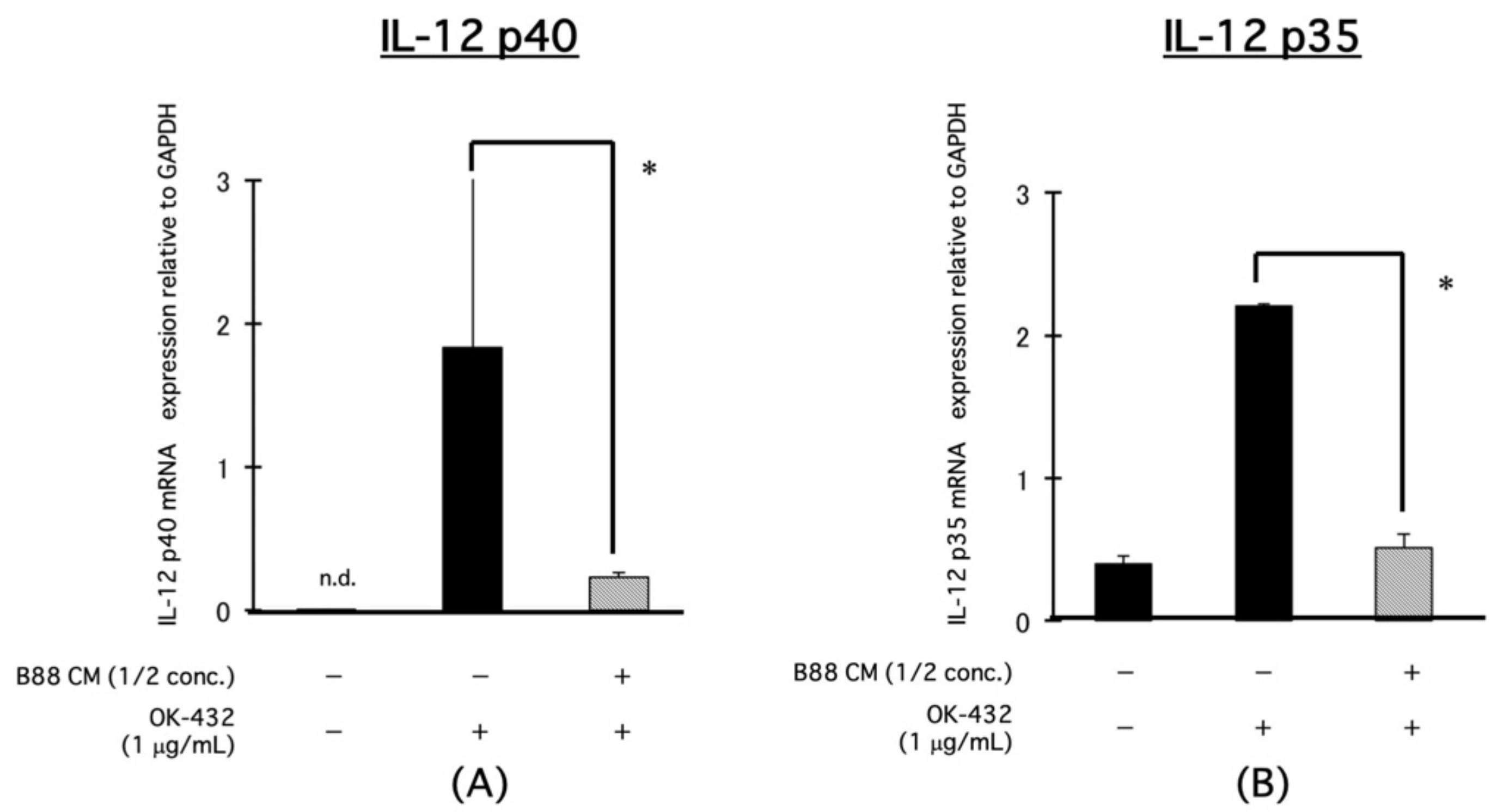
3.2. Blocking the Effect of CM on the mRNA Expression of IL-12-Related Molecules, IL-12p40, and IL-12p35 in PBMCs by OK-432 Treatment
3.3. Blocking the Effect of CM on the mRNA Expression of IL-12-Related Molecules, IL-12p40, and IL-12p35 in PBMCs by OK-432 Treatment
3.4. Effects of Anti-IL-12 Neutralizing Ab or rIL-12 on IFN-γ Production from PBMCs by OK-432 Treatment
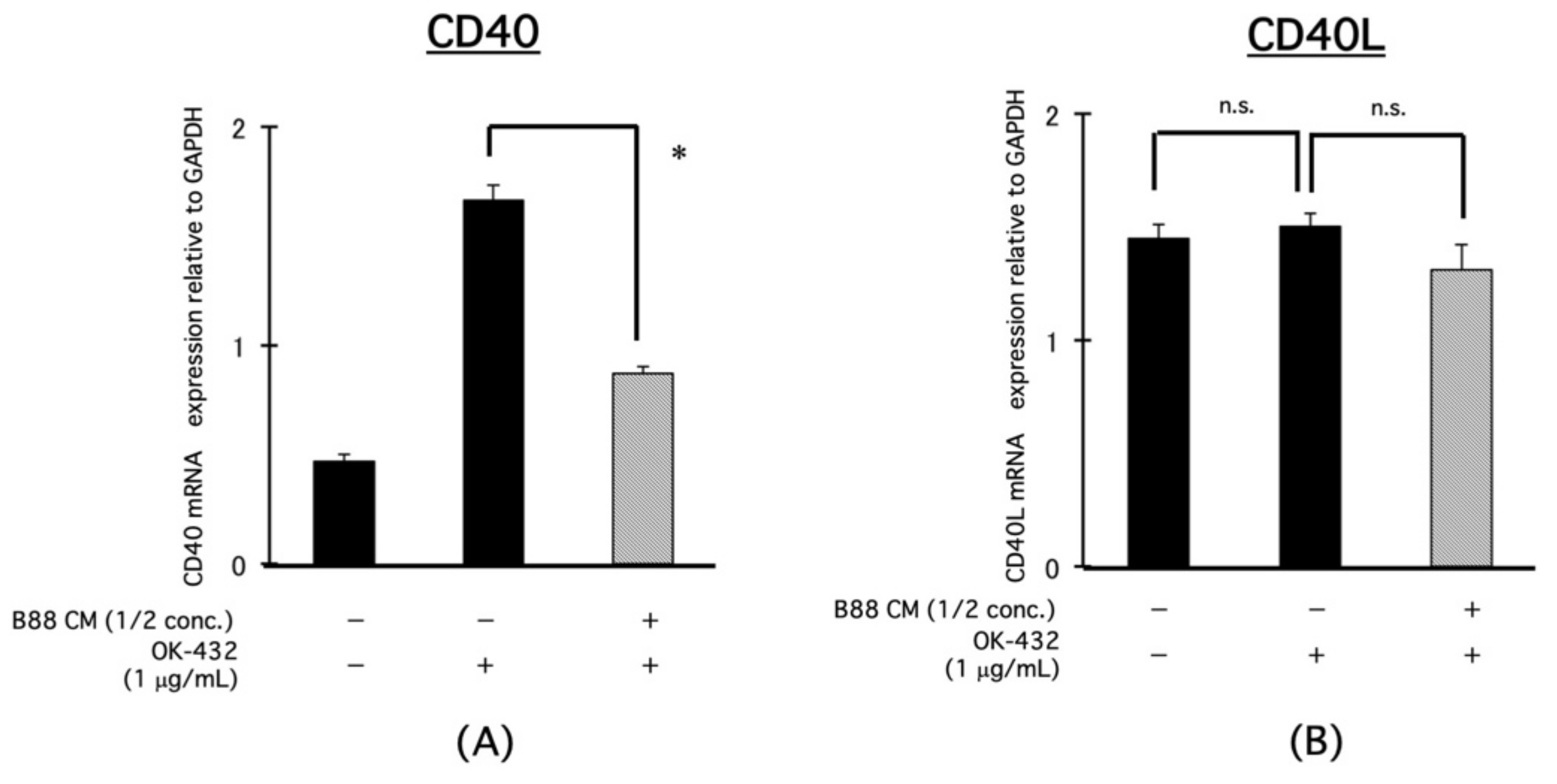
3.5. Blocking Effect of CM on the mRNA Expression of IL-12-Related Molecules, CD40, and CD40L in PBMCs by OK-432 Treatment
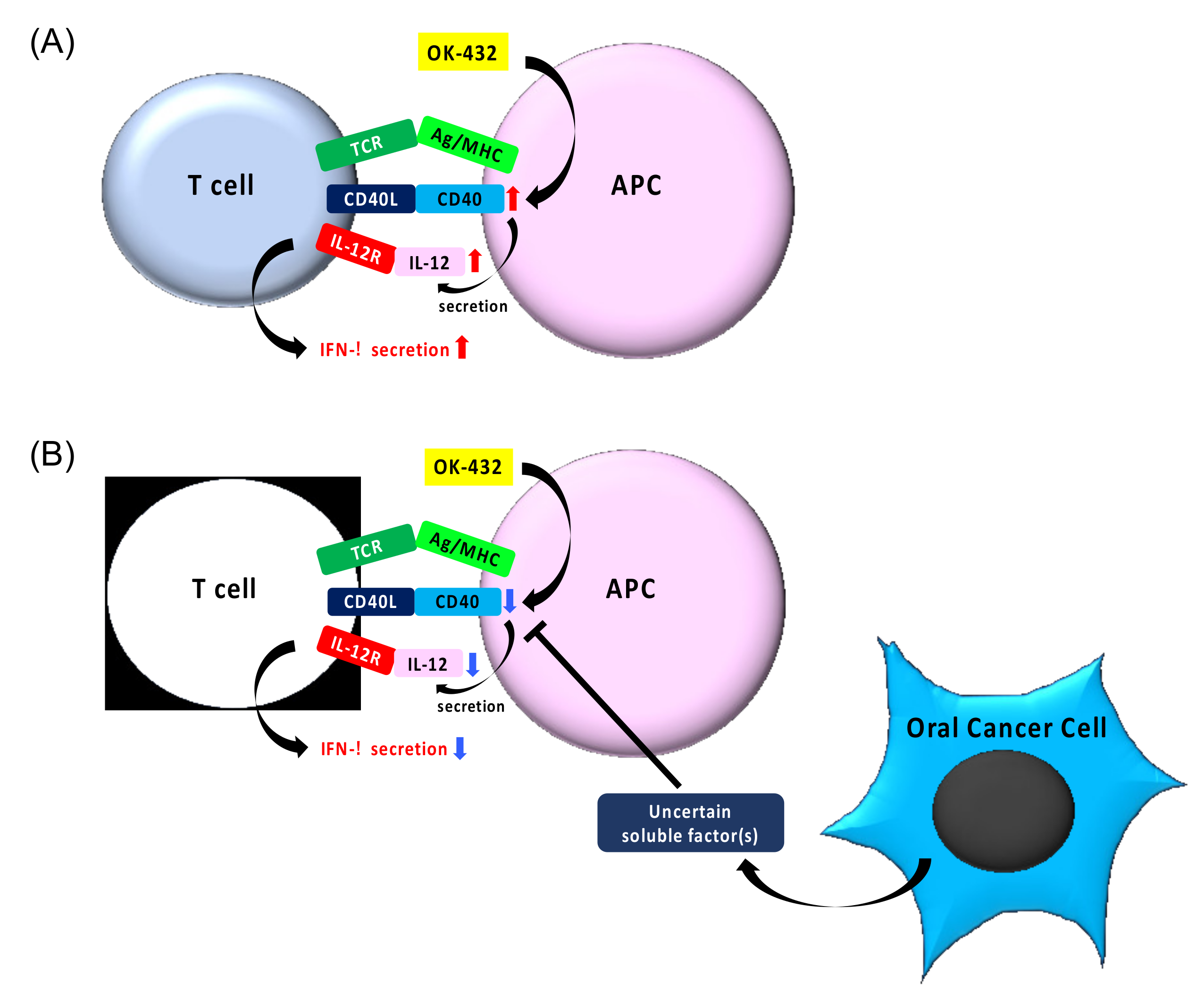
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Okamoto, H.; Shoin, S.; Koshimura, S.; Shimizu, R. Studies on the anticancer and streptolysin S-forming abilities of hemolytic streptococci. Jpn. J. Microbiol. 1967, 11, 323–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Iwa, T. Clinical value of immunotherapy with the streptococcal preparation OK-432 in non-small cell lung cancer. J. Biol. Response Mod. 1987, 6, 169–180. [Google Scholar] [PubMed]
- Katano, M.; Torisu, M. New approach to management of malignant ascites with a streptococcal preparation, OK-432. II. Intraperitoneal inflammatory cell-mediated tumor cell destruction. Surgery 1983, 93, 365–373. [Google Scholar]
- Uchida, A.; Micksche, M. Intrapleural administration of OK432 in cancer patients: Activation of NK cells and reduction of suppressor cells. Int. J. Cancer 1983, 31, 1–5. [Google Scholar] [CrossRef]
- Uchida, A.; Micksche, M.; Hoshino, T. Intrapleural administration of OK432 in cancer patients: Augmentation of autologous tumor killing activity of tumor-associated large granular lymphocytes. Cancer Immunol. Immunother. 1984, 18, 5–12. [Google Scholar] [CrossRef]
- Oshimi, K.; Kano, S.; Takaku, F.; Okumura, K. Augmentation of mouse natural killer cell activity by a streptococcal preparation, OK-432. J. Natl. Cancer Inst. 1980, 65, 1265–1269. [Google Scholar]
- Oshimi, K.; Wakasugi, H.; Seki, H.; Kano, S. Streptococcal preparation OK-432 augments cytotoxic activity against an erythroleukemic cell line in humans. Cancer Immunol. Immunother. 1980, 9, 187–191. [Google Scholar] [CrossRef]
- Matsubara, S.; Suzuki, F.; Ishida, N. Induction of immune interferon in mice treated with a bacterial immunopotentiator, OK-432. Cancer Immunol. Immunother. 1979, 6, 41–45. [Google Scholar] [CrossRef]
- Sato, M.; Hayashi, Y.; Yashida, H.; Yanagawa, T.; Yura, Y.; Urata, M.; Furumoto, M. Effect of immunotherapy with a streptococcal preparation, OK-432, on the peripheral killer lymphocyte population in patients with head and neck cancer. In Immunopharmacological Aspects of OK-432 in Humans; Excerpta Medica: Tokyo, Japan, 1986; pp. 78–88. [Google Scholar]
- Kaji, R.; Yoshida, H.; Yanagawa, T.; Sato, M. Monoclonal antibody to a human salivary gland adenocarcinoma cell line: Augmentation of antibody-dependent cell-mediated cytotoxicity activity by streptococcal preparation OK-432 in human salivary gland adenocarcinoma-bearing nude mice given the antibody. J. Biol. Response Mod. 1989, 8, 488–500. [Google Scholar]
- Nakahara, S.; Tsunoda, T.; Baba, T.; Asabe, S.; Tahara, H. Dendritic cells stimulated with a bacterial product, OK-432, efficiently induce cytotoxic T lymphocytes specific to tumor rejection peptide. Cancer Res. 2003, 63, 4112–4118. [Google Scholar]
- Sato, M.; Yoshida, H.; Yanagawa, T.; Yura, Y.; Urata, M.; Atsumi, M.; Hayashi, Y.; Takegawa, Y. Effects of intradermal administration of streptococcal preparation OK-432 on interferon and natural killer cell activities in patients with oral cancer. Int. J. Oral. Surg. 1984, 13, 7–15. [Google Scholar] [CrossRef]
- Sato, M.; Harada, K.; Yoshida, H.; Yura, Y.; Azuma, M.; Iga, H.; Bando, T.; Kawamata, H.; Takegawa, Y. Therapy for oral squamous cell carcinoma by tegafur and streptococcal agent OK-432 in combination with radiotherapy: Association of the therapeutic effect with differentiation and apoptosis in the cancer cells. Apoptosis 1997, 2, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.A. CD40: A cytokine receptor in search of a ligand. Tissue Antigens. 1990, 36, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Van Kooten, C.; Banchereau, J. CD40-CD40 ligand. J. Leukoc. Biol. 2000, 67, 2–17. [Google Scholar] [CrossRef]
- Kobayashi, M.; Fitz, L.; Ryan, M.; Hewick, R.M.; Clarck, S.C.; Chan, S.; Loudon, R.; Sherman, F.; Perussia, B.; Trinchieri, G. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J. Exp. Med. 1989, 170, 827–845. [Google Scholar] [CrossRef]
- Chan, S.H.; Perussia, B.; Gupta, J.W.; Kobayashi, M.; Pospisili, M.; Young, H.A.; Wolf, S.F.; Young, D.; Clark, S.C.; Trinchieri, G. Induction of interferon-γ production by natural killer cell stimulatory factor: Characterization of the responder cells and synergy with other inducers. J. Exp. Med. 1991, 173, 869–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.H.; Kobayashi, M.; Santoli, D.; Perussia, B.; Trinchieri, G. Mechanisms of IFN-γ induction by natural killer cell stimulatory factor (NKSF/IL-12). Role of transcription and mRNA stability in the synergistic interaction between NKSF and IL-2. J. Immunol. 1992, 148, 92–98. [Google Scholar] [PubMed]
- Wolf, S.F.; Temple, P.A.; Kobayashi, M.; Young, D.; Dicig, M.; Lowe, L.; Dzialo, R.; Fitz, L.; Ferenz, C.; Hewick, R.M. Cloning of cDNA for natural killer cell stimulatory factor, a heterodimeric cytokine with multiple biologic effects on T and natural killer cells. J. Immunol. 1991, 146, 3074–3081. [Google Scholar] [PubMed]
- Gubler, U.; Chua, A.O.; Schoenhaut, D.S.; Dwyer, C.M.; McComas, W.; Motyka, R.; Navabi, N.; Wolitzky, A.G.; Quinn, P.M.; Familletti, P.C. Coexpression of two distinct genes is required to generate secreted bioactive cytotoxic lymphocyte maturation factor. Proc. Natl. Acad. Sci. USA 1991, 88, 4143–4147. [Google Scholar] [CrossRef] [Green Version]
- Trinchieri, G. Interleukin-12: A cytokine at the interface of inflammation and immunity. Adv. Immunol. 1998, 70, 83–243. [Google Scholar]
- Wang, T.; Niu, G.; Kortylewski, M.; Burdelya, L.; Shain, K.; Zhang, S.; Bhattacharya, R.; Gabrilovich, D.; Heller, R.; Coppola, D.; et al. Regulation of the innate and adaptive immune responses by Stat-3 signaling in tumor cells. Nat. Med. 2004, 10, 48–54. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Okamoto, M.; Oshikawa, T.; Tano, T.; Ohe, G.; Furuichi, S.; Nishikawa, H.; Ahmed, S.U.; Akashi, S.; Miyake, K.; Takeuchi, O.; et al. Involvement of Toll-like receptor 4 signaling in interferon-gamma production and antitumor effect by streptococcal agent OK-432. J. Natl. Cancer Inst. 2003, 95, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Ohe, G.; Sasai, A.; Uchida, D.; Tamatani, T.; Nagai, H.; Miyamoto, Y. Effect of soluble factors derived from oral cancer cells on the production of interferon-gamma from peripheral blood mononuclear cells following stimulation with OK-432. Oncol. Rep. 2013, 30, 945–951. [Google Scholar] [CrossRef]
- Uchida, D.; Begum, N.M.; Almofti, A.; Nakashiro, K.; Kawamata, H.; Tateishi, Y.; Hamakawa, H.; Yoshida, H.; Sato, M. Possible role of stromal-cell-derived factor-1/CXCR4 signaling on lymph node metastasis of oral squamous cell carcinoma. Exp. Cell Res. 2003, 290, 289–302. [Google Scholar] [CrossRef]
- Kawamata, H.; Nakashiro, K.; Uchida, D.; Harada, K.; Yoshida, H.; Sato, M. Possible contribution of active MMP2 to lymph-node metastasis and secreted cathepsin L to bone invasion of newly established human oral-squamous-cancer cell lines. Int. J. Cancer 1997, 70, 120–127. [Google Scholar] [CrossRef]
- Yanagawa, T.; Hayashi, Y.; Yoshida, H.; Yura, Y.; Nagamine, S.; Bando, T.; Sato, M. An adenoid squamous carcinoma-forming cell line established from an oral keratinizing squamous cell carcinoma expressing carcinoembryonic antigen. Am. J. Pathol. 1986, 124, 496–509. [Google Scholar]
- Boyüm, A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of monouclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand. J. Clin. Lab. Invest. Suppl. 1968, 97, 77–89. [Google Scholar] [PubMed]
- Ma, X.; D’Andrea, A.; Kubin, M.; Aste-Amezaga, M.; Sartori, A.; Monteiro, J.; Showe, L.; Wysocka, M.; Trinchieri, G. Production of interleukin-12. Res. Immunol. 1995, 146, 432–438. [Google Scholar] [CrossRef]
- Kato, T.; Hakamada, R.; Yamane, H.; Nariuchi, H. Induction of IL-12 p40 messenger RNA expression and IL-12 production of macrophages via CD40-CD40 ligand interaction. J. Immunol. 1996, 156, 3932–3938. [Google Scholar]
- Nemunaitis, J.; Fong, T.; Robbins, J.M.; Edelman, G.; Edwards, W.; Paulson, R.S.; Bruce, J.; Ognoskie, N.; Wynne, D.; Pike, M.; et al. Phase I trial of interferon-gamma (IFN-gamma) retroviral vector administered intratumorally to patients with metastatic melanoma. Cancer Gene Ther. 1999, 6, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasu, Y.; Bangma, C.H.; Hull, G.W.; Lee, H.M.; Hu, J.; Wang, J.; McCurdy, M.A.; Shimura, S.; Yang, G.; Timme, T.L.; et al. Adenovirus-mediated interleukin-12 gene therapy for prostate cancer: Suppression of orthotopic tumor growth and pre-established lung metastases in an orthotopic model. Gene Ther. 1999, 6, 338–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saika, T.; Kusaka, N.; Mouraviev, V.; Satoh, T.; Kumon, H.; Timme, T.L.; Thompson, T.C. Therapeutic effects of adoptive splenocyte transfer following in situ AdIL-12 gene therapy in a mouse prostate cancer model. Cancer Gene Ther. 2006, 13, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Shurin, M.R.; Yurkovetsky, Z.R.; Tourkova, I.L.; Balkir, L.; Shurin, G.V. Inhibition of CD40 expression and CD40-mediated dendritic cell function by tumor-derived IL-10. Int. J. Cancer 2002, 101, 61–68. [Google Scholar] [CrossRef]
- Mendez-Samperio, P.; Garcia, E.; Vazquez, A.; Palma, J. Regulation of interleukin-8 by interleukin-10 and transforming growth factor beta in human monocytes infected with mycobacterium bovis. Clin. Diagn. Lab. Immunol. 2002, 9, 802–807. [Google Scholar] [PubMed] [Green Version]
- Banchereau, J.; Bazan, F.; Blanchard, D.; Briere, F.; Galizzi, J.P.; van Kooten, C.; Liu, Y.J.; Rousset, F.; Saeland, S. The CD40 antigen and its ligand. Annu. Rev. Immunol. 1994, 12, 881–922. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, J.F.; Wang, Y.D.; Qiu, Y.H.; Pan, J.Z.; Xie, W.; Jiang, L.Y.; Klein, B.; Zhang, X.G. An agonist anti-human CD40 monoclonal antibody that induces dendritic cell formation and maturation and inhibits proliferation of a myeloma cell line. Hybridoma 1999, 18, 471–478. [Google Scholar] [PubMed]
- Mauri, C.; Mars, L.T.; Londei, M. Therapeutic activity of agonistic monoclonal antibodies against CD40 in a chronic autoimmune inflammatory process. Nat. Med. 2000, 6, 673–679. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohe, G.; Kudo, Y.; Kamada, K.; Mouri, Y.; Takamaru, N.; Kudoh, K.; Kurio, N.; Miyamoto, Y. The Soluble Factor from Oral Cancer Cell Lines Inhibits Interferon-γ Production by OK-432 via the CD40/CD40 Ligand Pathway. Cancers 2021, 13, 3301. https://doi.org/10.3390/cancers13133301
Ohe G, Kudo Y, Kamada K, Mouri Y, Takamaru N, Kudoh K, Kurio N, Miyamoto Y. The Soluble Factor from Oral Cancer Cell Lines Inhibits Interferon-γ Production by OK-432 via the CD40/CD40 Ligand Pathway. Cancers. 2021; 13(13):3301. https://doi.org/10.3390/cancers13133301
Chicago/Turabian StyleOhe, Go, Yasusei Kudo, Kumiko Kamada, Yasuhiro Mouri, Natsumi Takamaru, Keiko Kudoh, Naito Kurio, and Youji Miyamoto. 2021. "The Soluble Factor from Oral Cancer Cell Lines Inhibits Interferon-γ Production by OK-432 via the CD40/CD40 Ligand Pathway" Cancers 13, no. 13: 3301. https://doi.org/10.3390/cancers13133301