
ΔNp73, TAp73 and Δ133p53 Extracellular Vesicle Cargo as Early Diagnosis Markers in Colorectal Cancer

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Subjects
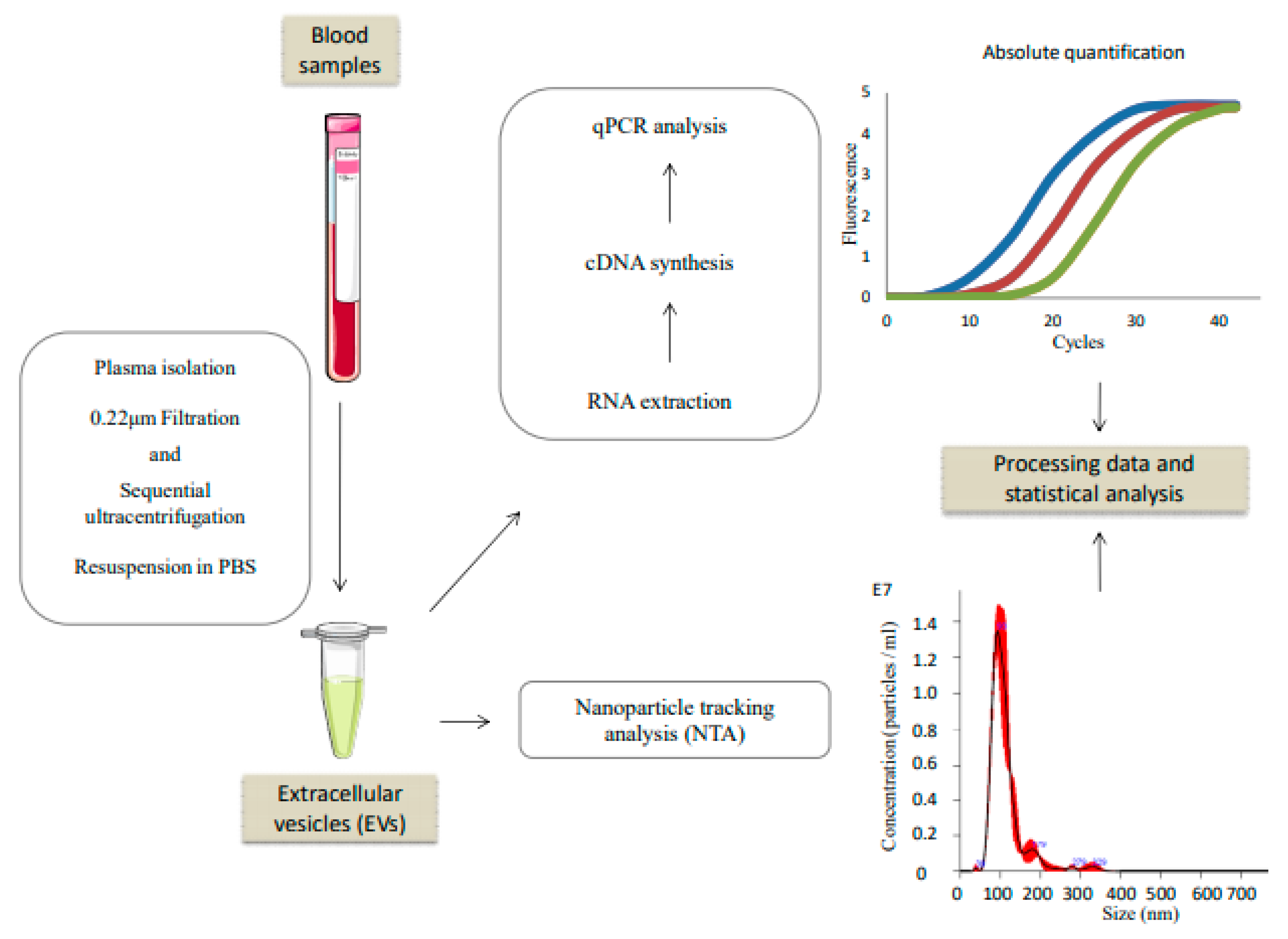
2.2. Extracellular Vesicles Isolation
2.3. Nanoparticle Tracking Analysis (NTA)
2.4. RNA Extraction, Reverse Transcription and qPCR
2.5. Statistical Analysis
3. Results
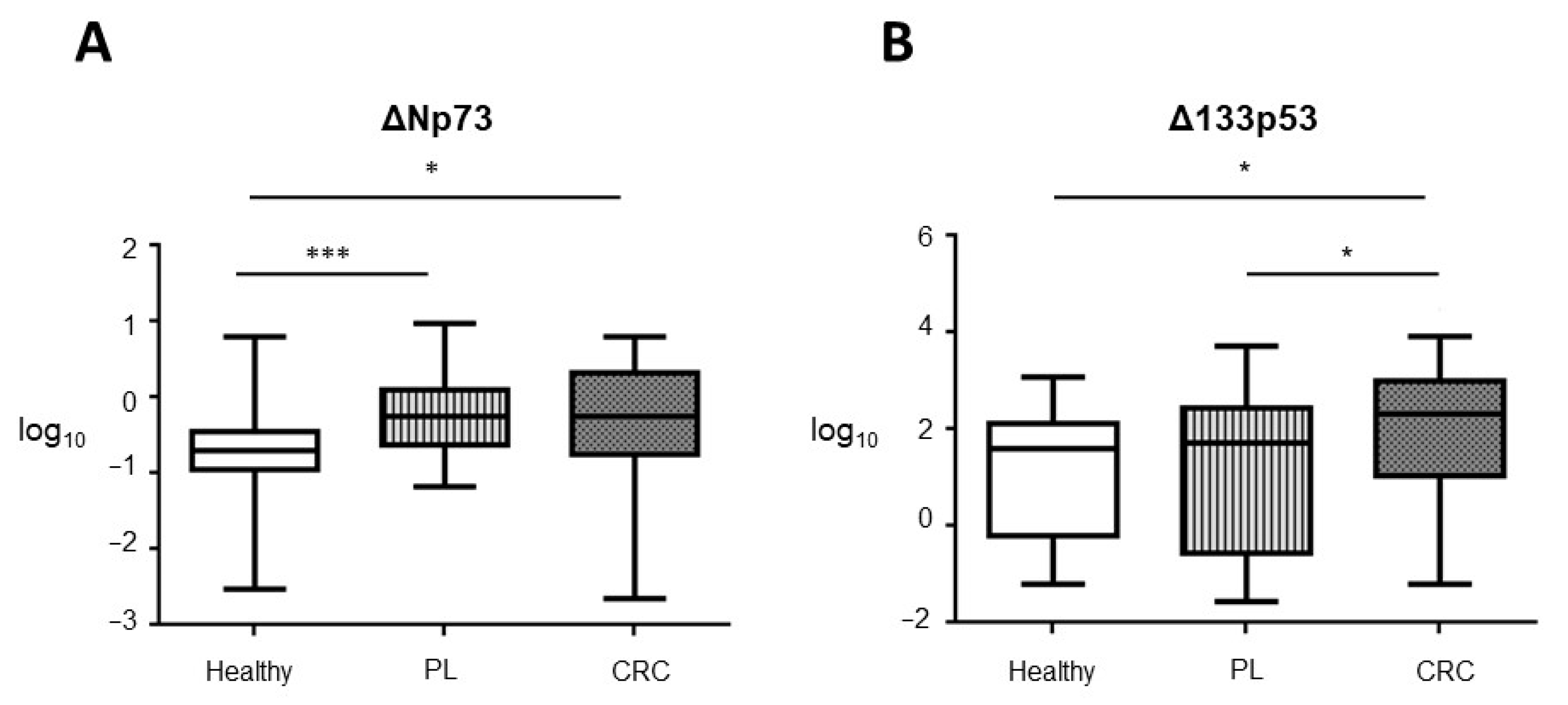
3.1. TAp73, ΔNp73, and Δ133p53 EVs Content in Control, PL and CRC Individuals
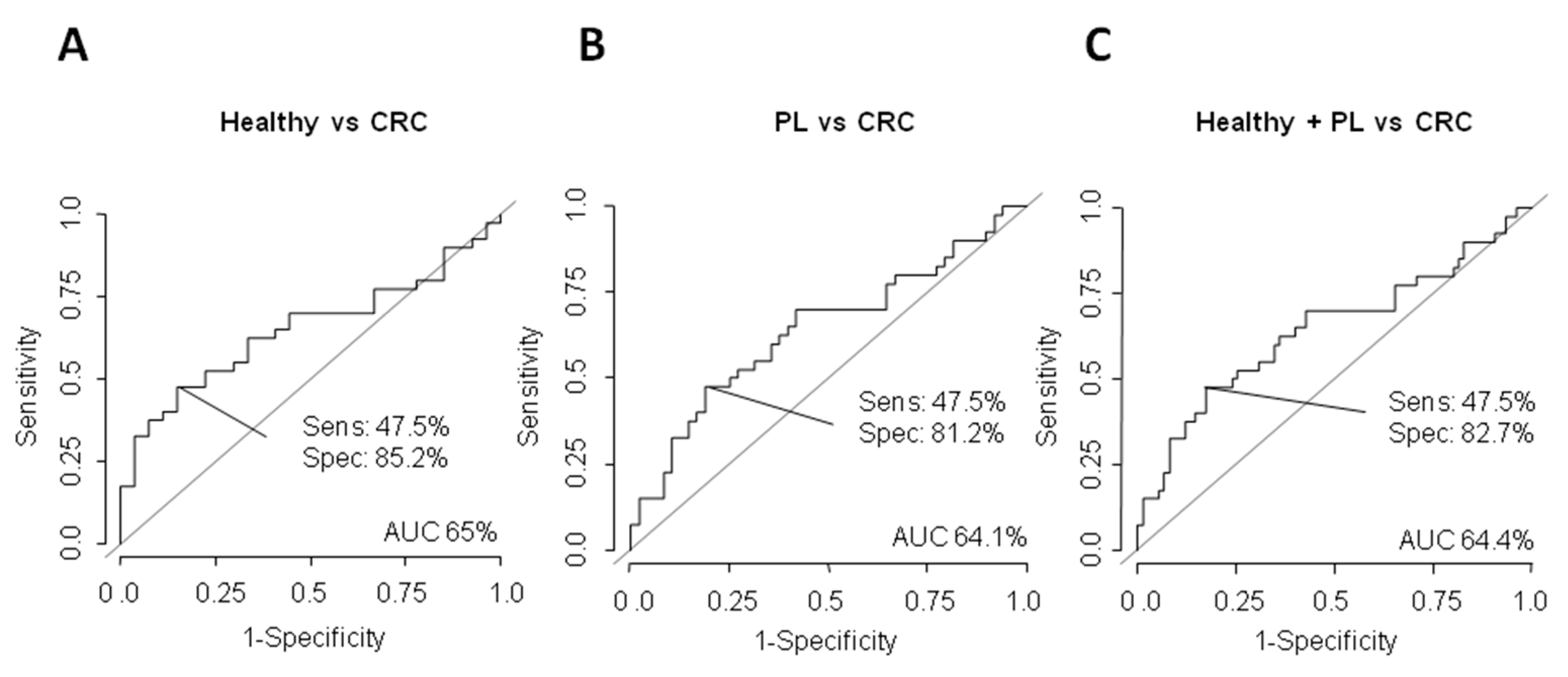
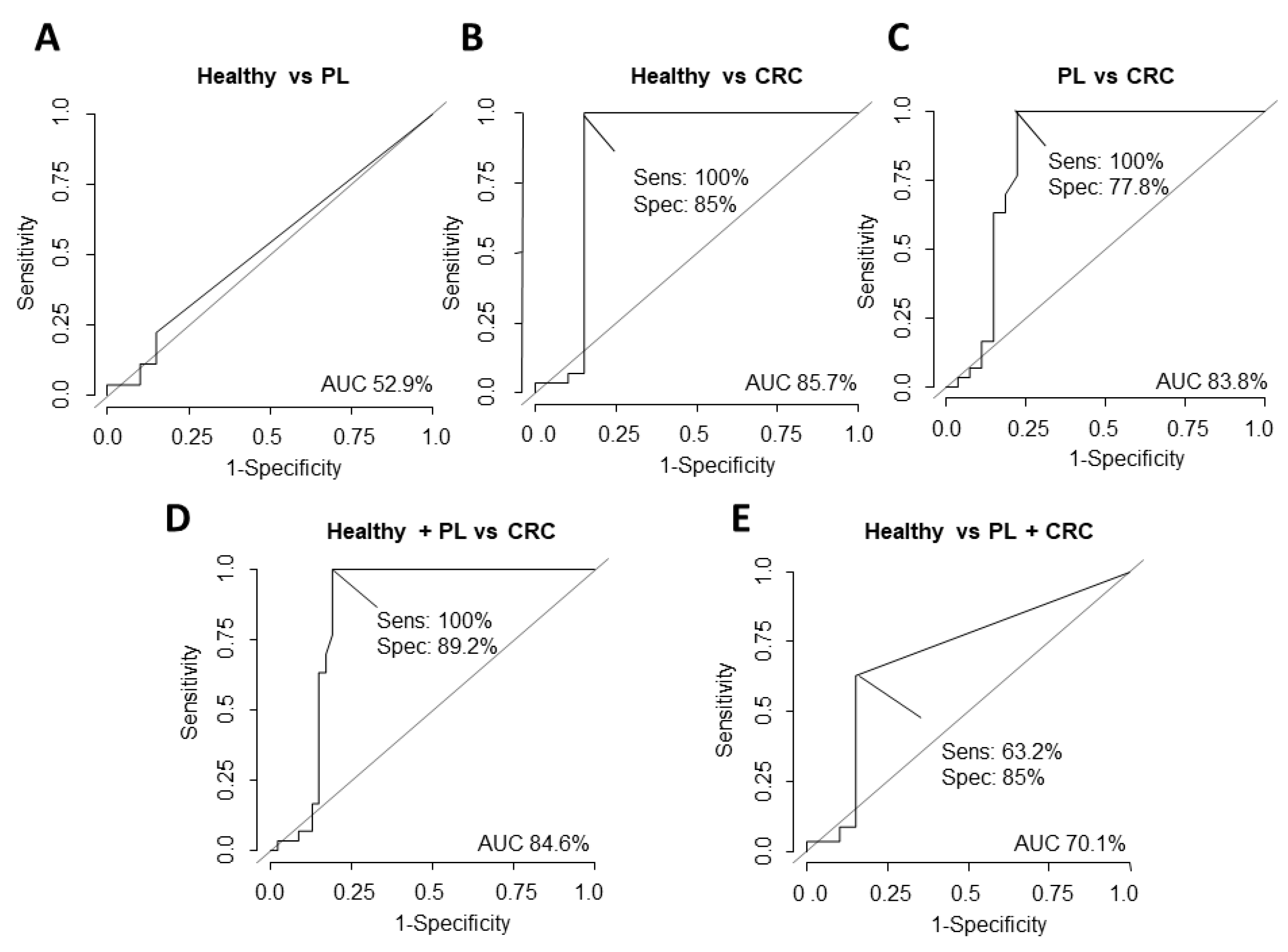
3.2. Potential Value of ΔNp73 and Δ133p53 Versus CEA as Diagnostic Biomarkers
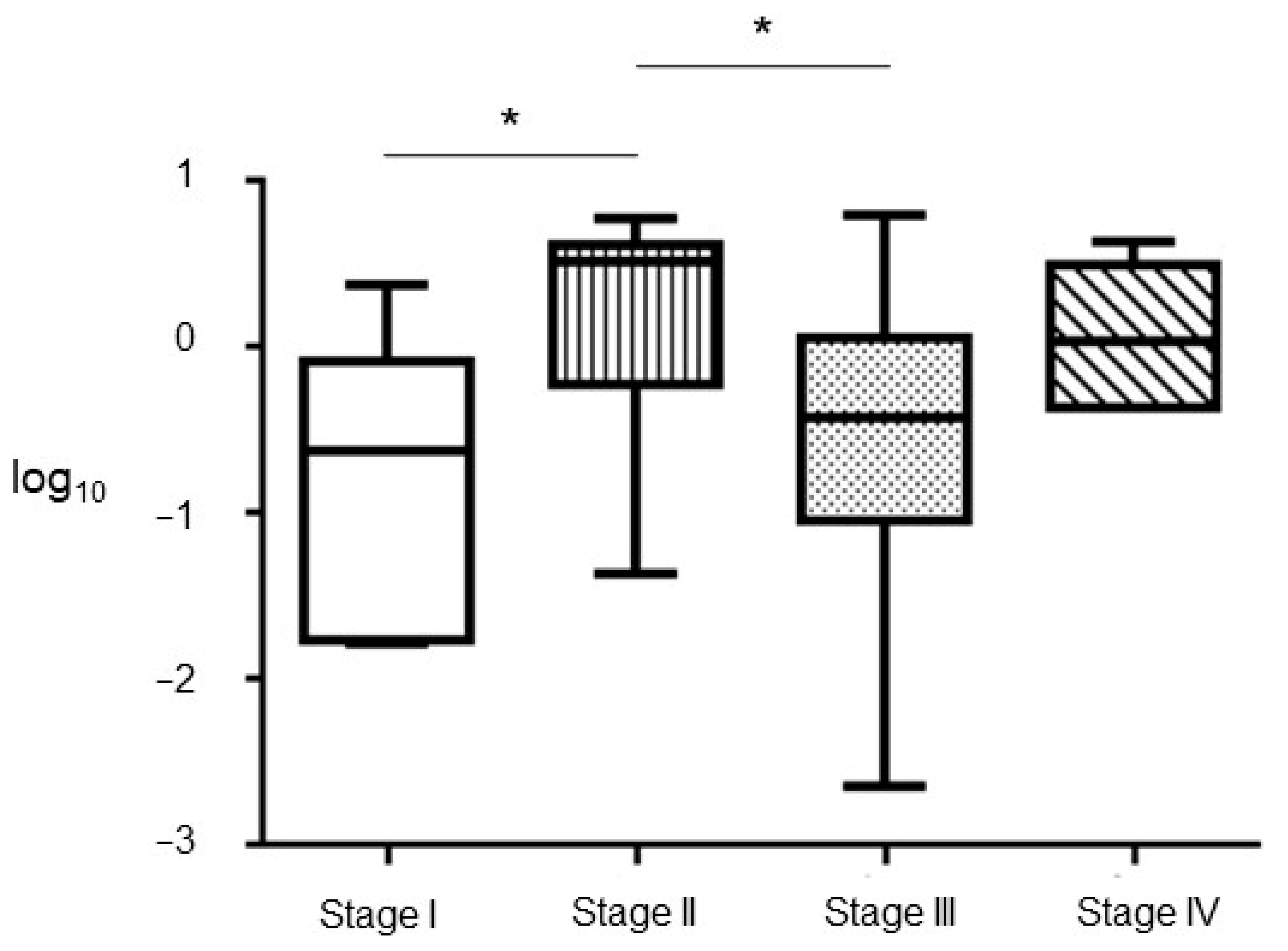
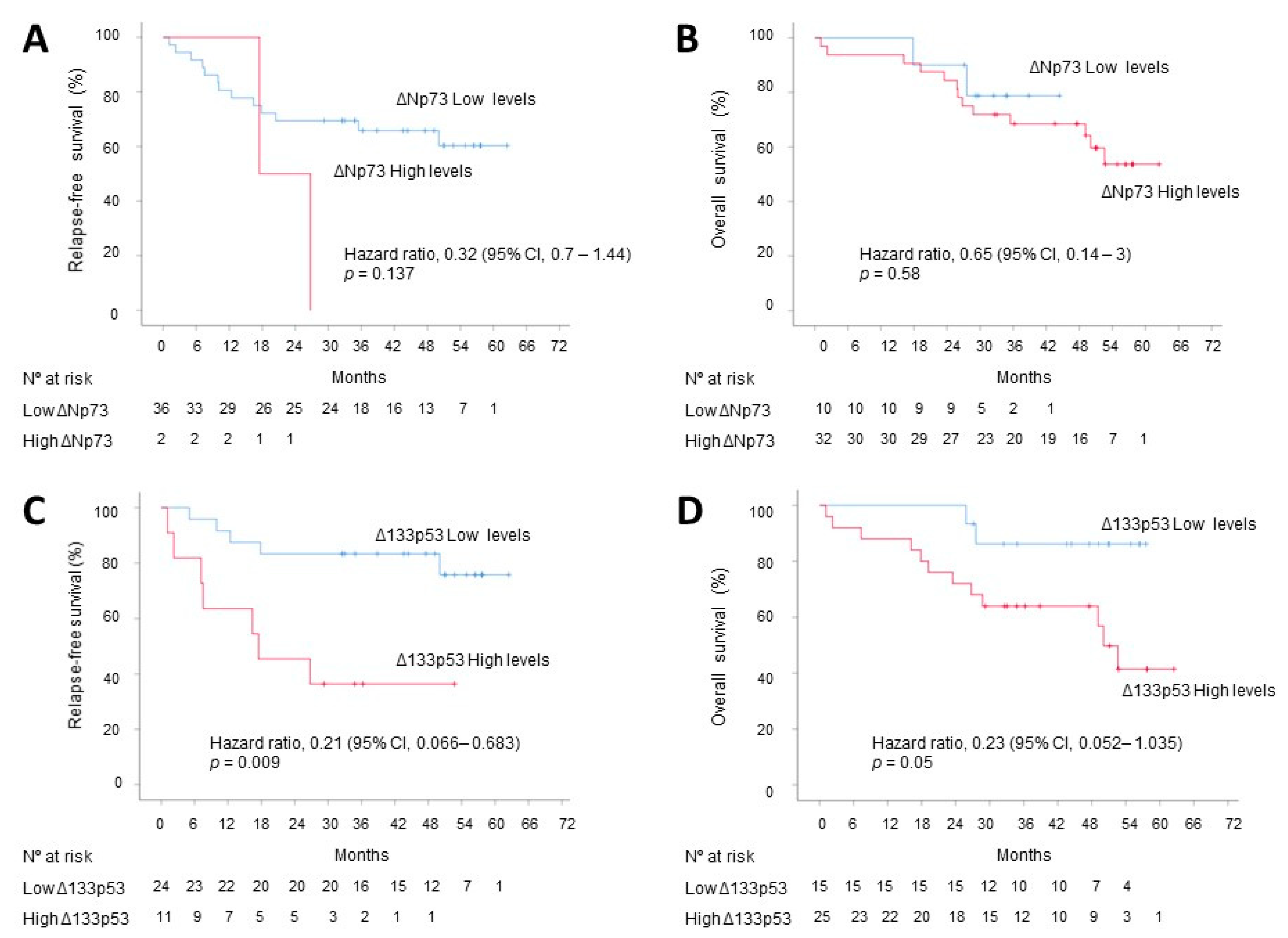
3.3. Correlation with Tumoral Stage, DFS and OS
3.4. Correlation between Seroreactivity of p53 Family Members and EVs Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Argilés, G.; Tabernero, J.; Labianca, R.; Hochhauser, D.; Salazar, R.; Iveson, T.; Laurent-Puig, P.; Quirke, P.; Yoshino, T.; Taieb, J.; et al. Localised colon cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2020, 31, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Lopes, G.; Stern, M.C.; Temin, S.; Sharara, A.I.; Cervantes, A.; Costas-Chavarri, A.; Engineer, R.; Hamashima, C.; Ho, G.F.; Huitzil, F.D.; et al. Early detection for colorectal cancer: ASCO resource-stratified guideline. J. Glob. Oncol. 2019, 5, 1–22. [Google Scholar]
- Lin, J.S.; Piper, M.A.; Perdue, L.A.; Rutter, C.M.; Webber, E.M.; O’Connor, E.; Smith, N.; Whitlock, E.P. Screening for colorectal cancer: Updated evidence report and systematic review for the US preventive services task force. Jama 2016, 315, 2576–2594. [Google Scholar] [CrossRef] [Green Version]
- Force, U.S.P.S.T.; Bibbins-Domingo, K.; Grossman, D.C.; Curry, S.J.; Davidson, K.W.; Epling, J.W., Jr.; García, F.; Gillman, M.W.; Harper, D.M.; Kemper, A.R.; et al. Screening for colorectal cancer: US preventive services task force recommendation statement. Jama 2016, 315, 2564–2575. [Google Scholar]
- Zauber, A.; Knudsen, A.; Rutter, C.M.; Lansdorp-Vogelaar, I.; Kuntz, K.M. Evaluating the benefits and harms of colorectal cancer screening strategies: A collaborative modeling approach. AHRQ Publication No. 14-05203-EF-2. Rockville, MD: Agency for Healthcare Research and Quality. Available online: https://www.uspreventiveservicestaskforce.org (accessed on 1 October 2015).
- Knudsen, A.B.; Zauber, A.G.; Rutter, C.M.; Naber, S.K.; Doria-Rose, V.P.; Pabiniak, C.; Pabiniak, C.; Johanson, C.; Fischer, S.E.; Lansdorp-Vogelaar, I.; et al. Estimation of benefits, burden, and harms of colorectal cancer screening strategies: Modeling study for the US preventive services task force. Jama 2016, 315, 2595–2609. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Niu, Y.; Li, K.; Liu, X.; Chen, H.; Gao, C. A systematic review and meta-analysis of diagnostic and prognostic serum biomarkers of colorectal cancer. PLoS ONE 2014, 9, e103910. [Google Scholar] [CrossRef] [PubMed]
- Siravegna, G.; Marsoni, S.; Siena, S.; Bardelli, A. Integrating liquid biopsies into the management of cancer. Nat. Rev. Clin. Oncol. 2017, 14, 531–548. [Google Scholar] [CrossRef]
- Whiteside, T.L. Validation of plasma-derived small extracellular vesicles as cancer biomarkers. Nat. Rev. Clin. Oncol. 2020, 17, 719–720. [Google Scholar] [CrossRef]
- Dai, J.; Su, Y.; Zhong, S.; Cong, L.; Liu, B.; Yang, J.; Tao, Y.; He, Z.; Chen, C.; Jiang, Y. Exosomes: Key players in cancer and potential therapeutic strategy. Sig. Transduct. Target. Ther. 2020, 5, 145. [Google Scholar] [CrossRef]
- Siskova, A.; Cervena, K.; Kral, J.; Hucl, T.; Vodicka, P.; Vymetalkova, V. Colorectal adenomas-genetics and searching for new molecular screening biomarkers. Int. J. Molec. Sci. 2020, 21, 3260. [Google Scholar] [CrossRef]
- Rodriguez, N.; Pelaez, A.; Barderas, R.; Dominguez, G. Clinical implications of the deregulated TP73 isoforms expression in cancer. Clin. Translat. Oncol. 2018, 20, 827–836. [Google Scholar] [CrossRef]
- Sun, X.F. p73 overexpression is a prognostic factor in patients with colorectal adenocarcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2002, 8, 165–170. [Google Scholar]
- Dominguez, G.; Garcia, J.M.; Pena, C.; Silva, J.; Garcia, V.; Martinez, L.; Maximiano, C.; Gómez, M.E.; Rivera, J.A.; García-Andrade, C.; et al. DeltaTAp73 upregulation correlates with poor prognosis in human tumors: Putative in vivo network involving p73 isoforms, p53, and E2F-1. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 805–815. [Google Scholar] [CrossRef]
- Soldevilla, B.; Diaz, R.; Silva, J.; Campos-Martin, Y.; Munoz, C.; Garcia, V.; García, J.M.; Peña, C.; Herrera, M.; Rodriguez, M.; et al. Prognostic impact of DeltaTAp73 isoform levels and their target genes in colon cancer patients. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 6029–6039. [Google Scholar] [CrossRef] [Green Version]
- Soldevilla, B.; Rodriguez, M.; San Millan, C.; Garcia, V.; Fernandez-Perianez, R.; Gil-Calderon, B.; Martín, P.; García-Grande, A.; Silva, J.; Bonilla, F.; et al. Tumor-derived exosomes are enriched in DeltaNp73, which promotes oncogenic potential in acceptor cells and correlates with patient survival. Hum. Molec. Genet. 2014, 23, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Sui, L.; Du, M.; Wang, Q. Meta-analysis of TP73 polymorphism and cervical cancer. Genet. Molec. Res. 2017, 16. [Google Scholar] [CrossRef]
- Arvanitis, D.A.; Spandidos, D.A. Deregulation of the G1/S phase transition in cancer and squamous intraepithelial lesions of the uterine cervix: A case control study. Oncol. Rep. 2008, 20, 751–760. [Google Scholar]
- Chen, Y.K.; Hsue, S.S.; Lin, L.M. p73 expression for human buccal epithelial dysplasia and squamous cell carcinoma: Does it correlate with nodal status of carcinoma and is there a relationship with malignant change of epithelial dysplasia? Head Neck 2004, 26, 945–952. [Google Scholar] [CrossRef]
- Chen, K.M.; Stephen, J.K.; Raju, U.; Worsham, M.J. Delineating an epigenetic continuum for initiation, transformation and progression to breast cancer. Cancers 2011, 3, 1580–1592. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Zhang, Y.; Jiang, K.; Ye, S.; Chen, S.; Zhang, Q.; Peng, J.; Chen, J. p73 coordinates with Δ133p53 to promote DNA double-strand break repair. Cell Death Diff. 2018, 25, 1063–1079. [Google Scholar] [CrossRef]
- Liu, K.; Lin, J. Δ133p53 decreases the chemosensitivity of carcinoma cell line H1299. Cell Biol. Int. 2019, 43, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Arsic, N.; Ho-Pun-Cheung, A.; Lopez-Crapez, E.; Assenat, E.; Jarlier, M.; Anguille, C.; Colard, M.; Pezet, M.; Roux, P.; Gadea, G. The p53 isoform Δ133p53ss regulates cancer cell apoptosis in a RhoB-dependent manner. PLoS ONE 2017, 12, e0172125. [Google Scholar]
- Horikawa, I.; Park, K.Y.; Isogaya, K.; Hiyoshi, Y.; Li, H.; Anami, K.; Robles, A.I.; Mondal, A.M.; Fujita, K.; Serrano, M.; et al. Δ133p53 represses p53-inducible senescence genes and enhances the generation of human induced pluripotent stem cells. Cell Death Diff. 2017, 24, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, I.; Fujita, K.; Jenkins, L.M.; Hiyoshi, Y.; Mondal, A.M.; Vojtesek, B.; Lane, D.P.; Appella, E.; Harris, C.C. Autophagic degradation of the inhibitory p53 isoform Delta133p53α as a regulatory mechanism for p53-mediated senescence. Nat. Commun. 2014, 5, 4706. [Google Scholar] [CrossRef] [Green Version]
- Joruiz, S.M.; Beck, J.A.; Horikawa, I.; Harris, C.C. The Δ133p53 isoforms, tuners of the p53 pathway. Cancers 2020, 12, 3422. [Google Scholar] [CrossRef]
- Bourdon, J.C.; Fernandes, K.; Murray-Zmijewski, F.; Liu, G.; Diot, A.; Xirodimas, D.P.; Saville, M.K.; Lane, D.P. p53 isoforms can regulate p53 transcriptional activity. Genes Dev. 2005, 19, 2122–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Mondal, A.M.; Horikawa, I.; Nguyen, G.H.; Kumamoto, K.; Sohn, J.J.; Bowman, E.D.; Mathe, E.A.; Schetter, A.J.; Pine, S.R.; et al. p53 isoforms Δ133p53 and p53β are endogenous regulators of replicative cellular senescence. Nat. Cell Biol. 2009, 11, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Campbell, H.; Fleming, N.; Roth, I.; Mehta, S.; Wiles, A.; Williams, G.; Vennin, C.; Arsic, N.; Parkin, A.; Pajic, M.; et al. 133p53 isoform promotes tumour invasion and metastasis via interleukin-6 activation of JAK-STAT and RhoA-ROCK signalling. Nat. Commun. 2018, 9, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, H.; Garmy-Susini, B.; Ainaoui, N.; Van Den Berghe, L.; Peurichard, A.; Javerzat, S.; Bikfalvi, A.; Lane, D.P.; Bourdon, J.C.; Prats, A.C. The p53 isoform, Δ133p53aα, stimulates angiogenesis and tumour progression. Oncogene 2013, 32, 2150–2160. [Google Scholar] [CrossRef] [Green Version]
- Gadea, G.; Arsic, N.; Fernandes, K.; Diot, A.; Joruiz, S.M.; Abdallah, S.; Meuray, V.; Vinot, S.; Anguille, C.; Remenyi, J.; et al. TP53 drives invasion through expression of its Δ133p53β variant. eLife 2016, 5, e14734. [Google Scholar] [CrossRef]
- Kazantseva, M.; Mehta, S.; Eiholzer, R.A.; Gimenez, G.; Bowie, S.; Campbell, H.; Reily-Bell, A.L.; Roth, I.; Ray, S.; Drummond, C.J.; et al. The Δ133p53β isoform promotes an immunosuppressive environment leading to aggressive prostate cancer. Cell Death Dis. 2019, 10, 631. [Google Scholar] [CrossRef]
- Garranzo-Asensio, M.; Guzman-Aranguez, A.; Poves, C.; Fernandez-Acenero, M.J.; Montero-Calle, A.; Ceron, M.A.; Fernandez-Diez, S.; Rodríguez, N.; Gómez de Cedrón, M.; Ramírez de Molina, A.; et al. The specific seroreactivity to Np73 isoforms shows higher diagnostic ability in colorectal cancer patients than the canonical p73 protein. Sci. Rep. 2019, 9, 13547. [Google Scholar] [CrossRef]
- Muller, M.; Schilling, T.; Sayan, A.E.; Kairat, A.; Lorenz, K.; Schulze-Bergkamen, H.; Oren, M.; Koch, A.; Tannapfel, A.; Stremmel, W.; et al. TAp73/Δ Np73 influences apoptotic response, chemosensitivity and prognosis in hepatocellular carcinoma. Cell Death Diff. 2005, 12, 1564–1577. [Google Scholar] [CrossRef]
- Casciano, I.; Mazzocco, K.; Boni, L.; Pagnan, G.; Banelli, B.; Allemanni, G.; Ponzoni, M.; Tonini, G.P.; Romani, M. Expression of ΔNp73 is a molecular marker for adverse outcome in neuroblastoma patients. Cell Death Diff. 2002, 9, 246–251. [Google Scholar] [CrossRef]
- Uramoto, H.; Sugio, K.; Oyama, T.; Nakata, S.; Ono, K.; Morita, M.; Funa, K.; Yasumoto, K. Expression of ΔNp73 predicts poor prognosis in lung cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 6905–6911. [Google Scholar] [CrossRef] [Green Version]
- Vilgelm, A.E.; Hong, S.M.; Washington, M.K.; Wei, J.; Chen, H.; El-Rifai, W.; Zaika, A. Characterization of ΔNp73 expression and regulation in gastric and esophageal tumors. Oncogene 2010, 29, 5861–5868. [Google Scholar] [CrossRef] [Green Version]
- Lucena-Araujo, A.R.; Kim, H.T.; Thome, C.; Jacomo, R.H.; Melo, R.A.; Bittencourt, R.; Pasquini, R.; Pagnano, K.; Glória, A.B.; Chauffaille, M.; et al. High ΔNp73/TAp73 ratio is associated with poor prognosis in acute promyelocytic leukemia. Blood 2015, 126, 2302–2306. [Google Scholar] [CrossRef] [Green Version]
- Toumi, A.A.; El Hadj Oel, A.; Ben Mahmoud, L.K.; Ben Hmida Ael, M.; Chaar, I.; Gharbi, L.; Mzabi, S.; Bouraoui, S. The prognostic value of p73 overexpression in colorectal carcinoma: A clinicopathologic, immunohistochemical, and statistical study of 204 patients. AIMM 2010, 18, 128–136. [Google Scholar] [CrossRef]
- Su, X.L.; Ouyang, X.H.; Yan, M.R.; Liu, G.R. p73 expression and its clinical significance in colorectal cancer. Colorectal Dis. Off. J. Assoc. Coloproctol. G. B. Irel. 2009, 11, 960–963. [Google Scholar] [CrossRef]
- Leupin, N.; Luthi, A.; Novak, U.; Grob, T.J.; Hugli, B.; Graber, H.; Fey, M.F.; Tobler, A. P73 status in B-cell chronic lymphocytic leukaemia. Leukemia Lymphoma 2004, 45, 1205–1207. [Google Scholar] [CrossRef]
- Faridoni-Laurens, L.; Tourpin, S.; Alsafadi, S.; Barrois, M.; Temam, S.; Janot, F.; Koscielny, S.; Bosq, J.; Bénard, J.; Ahomadegbe, J.C. Involvement of N-terminally truncated variants of p73, ΔTAp73, in head and neck squamous cell cancer: A comparison with p53 mutations. Cell Cycle 2008, 7, 1587–1596. [Google Scholar] [CrossRef]
- Peinado, H.; Aleckovic, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; García-Santos, G.; Ghajar, C.; et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Al-Nedawi, K.; Meehan, B.; Micallef, J.; Lhotak, V.; May, L.; Guha, A.; Rak, J. Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nat. Cell Biol. 2008, 10, 619–624. [Google Scholar] [CrossRef]
- Fakih, M.G.; Padmanabhan, A. CEA monitoring in colorectal cancer. What you should know. Oncology 2006, 20, 579–587. [Google Scholar]
- Fletcher, R.H. Carcinoembryonic antigen. Ann. Intern. Med. 1986, 104, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Wanebo, H.J.; Rao, B.; Pinsky, C.M.; Hoffman, R.G.; Stearns, M.; Schwartz, M.K.; Oettgen, H.F. Preoperative carcinoembryonic antigen level as a prognostic indicator in colorectal cancer. N. Engl. J. Med. 1978, 299, 448–451. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Zhou, Y.; Sheng, S.; Qian, S.Y.; Huo, X. Evaluation of serum CEA, CA19-9, CA72-4, CA125 and ferritin as diagnostic markers and factors of clinical parameters for colorectal cancer. Sci. Rep. 2018, 8, 2732. [Google Scholar] [CrossRef]
- Chevinsky, A.H. CEA in tumors of other than colorectal origin. Semin. Surg. Oncol. 1991, 7, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Bormer, O.P. Standardization, specificity, and diagnostic sensitivity of four immunoassays for carcinoembryonic antigen. Clin. Chem. 1991, 37, 231–236. [Google Scholar] [CrossRef]
- Taylor, R.N.; Fulford, K.M.; Huong, A.Y. Results of a nationwide proficiency test for carcinoembryonic antigen. J. Clin. Microbiol. 1977, 5, 433–438. [Google Scholar] [PubMed]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat. Cell Biol. 2015, 17, 816–826. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Healthy Individuals | Subjects with PL | CRC Patients |
|---|---|---|---|
| n | 29 | 49 | 42 |
| Sex | |||
| Male | 10 (34.48%) | 24 (48.98%) | 22 (52.38%) |
| Female | 19 (65.52%) | 25 (51.02%) | 20 (47.62%) |
| Age (years) | |||
| Median, range | 59 (42–79) | 62 (31–81) | 71 (49–89) |
| CEA (ng/mL) | |||
| Median, range | 0.0 (0.0–81.9) | 0.0 (0.0–135.2) | 3.1 (0.5–94.9) |
| Tumor stage | |||
| I | 5 (11.905%) | ||
| II | 9 (21.43%) | ||
| III | 23 (54.76%) | ||
| IV | 5 (11.905%) | ||
| Tumor location | |||
| Right colon | 14 (33.33%) | ||
| Left colon | 17 (40.48%) | ||
| Rectum | 11 (26.19%) | ||
| DFS (n = 38) | |||
| Median, CI | NR | ||
| 3 years DFS (95%, CI) | 75% (61–89) | ||
| OS | |||
| Median, CI | NR | ||
| 3 years OS (95%, CI) | 69% (55–83) | ||
| Seroreactiviy vs. EVs Content | Healthy Subjects | PL | CRC |
|---|---|---|---|
| p53 autoantibodies vs. EVs Δ133p53 | 0.194 | −0.303 | −0.339 |
| p73 autoantibodies vs. EVs ΔNp73 | −0.110 | 0.127 | 0.018 |
| ΔNp73α autoantibodies vs. EVs ΔNp73 | −0.064 | 0.050 | −0.548 |
| ΔNp73β autoantibodies vs. EVs ΔNp73 | −0.064 | 0.018 | −0.305 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Cobos, J.; Viñal, D.; Poves, C.; Fernández-Aceñero, M.J.; Peinado, H.; Pastor-Morate, D.; Prieto, M.I.; Barderas, R.; Rodríguez-Salas, N.; Domínguez, G. ΔNp73, TAp73 and Δ133p53 Extracellular Vesicle Cargo as Early Diagnosis Markers in Colorectal Cancer. Cancers 2021, 13, 2240. https://doi.org/10.3390/cancers13092240
Rodríguez-Cobos J, Viñal D, Poves C, Fernández-Aceñero MJ, Peinado H, Pastor-Morate D, Prieto MI, Barderas R, Rodríguez-Salas N, Domínguez G. ΔNp73, TAp73 and Δ133p53 Extracellular Vesicle Cargo as Early Diagnosis Markers in Colorectal Cancer. Cancers. 2021; 13(9):2240. https://doi.org/10.3390/cancers13092240
Chicago/Turabian StyleRodríguez-Cobos, Javier, David Viñal, Carmen Poves, María J. Fernández-Aceñero, Héctor Peinado, Daniel Pastor-Morate, Mª Isabel Prieto, Rodrigo Barderas, Nuria Rodríguez-Salas, and Gemma Domínguez. 2021. "ΔNp73, TAp73 and Δ133p53 Extracellular Vesicle Cargo as Early Diagnosis Markers in Colorectal Cancer" Cancers 13, no. 9: 2240. https://doi.org/10.3390/cancers13092240