
Syndecan-1 Overexpressing Mesothelioma Cells Inhibit Proliferation, Wound Healing, and Tube Formation of Endothelial Cells


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
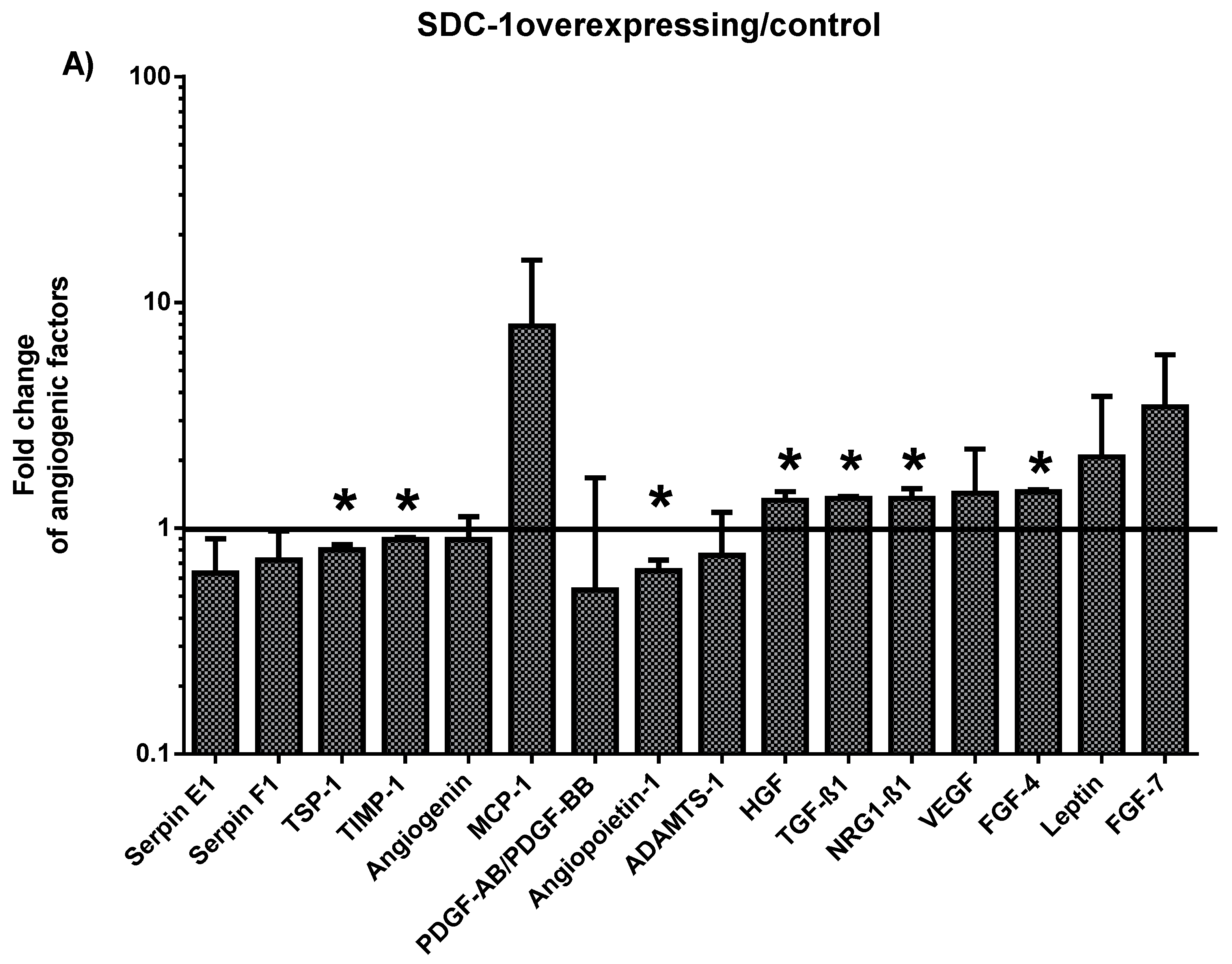
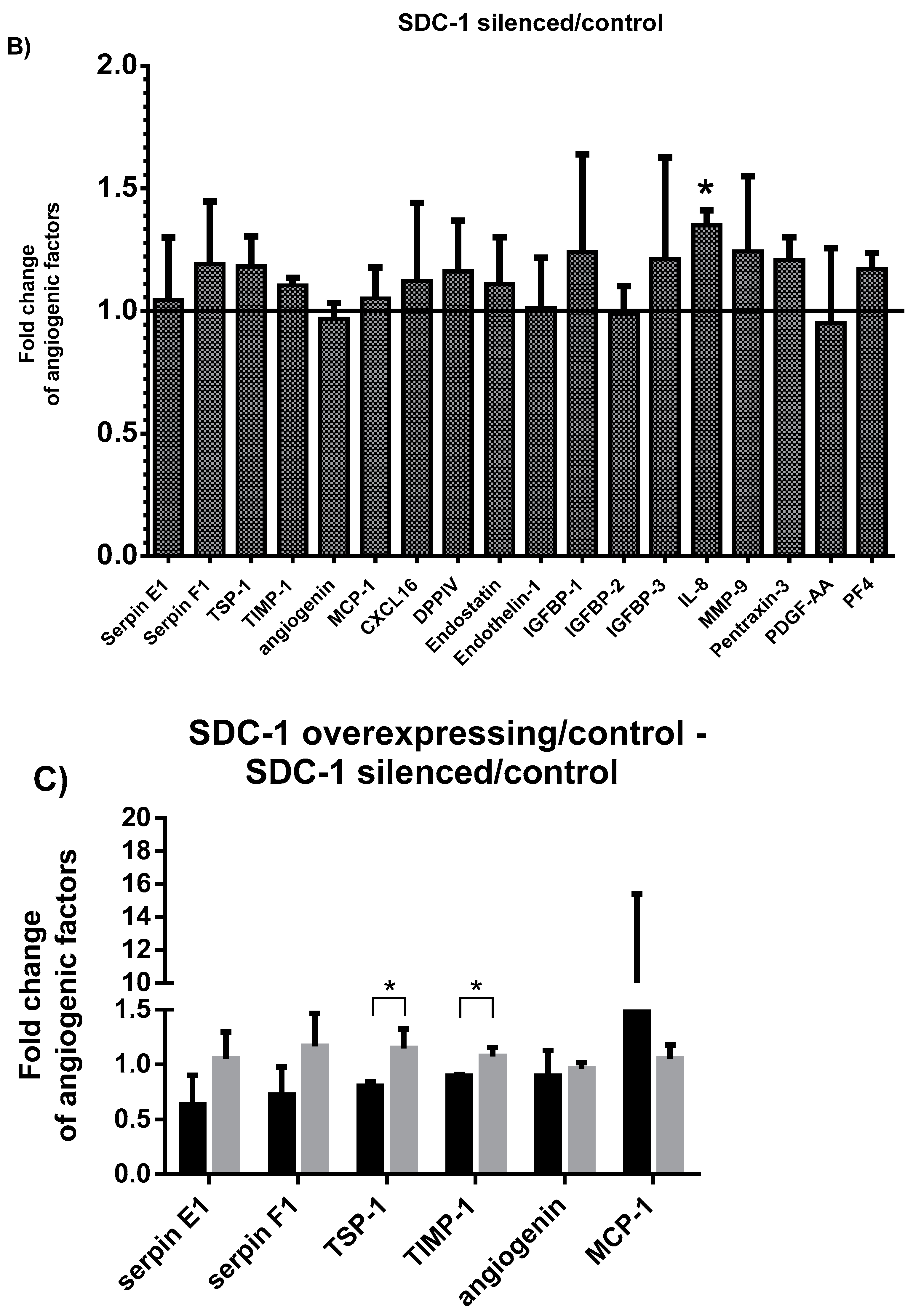
2.1. Characterization of the Angiogenesis-Related Proteins Secreted by SDC-1 Overexpressing and SDC-1 Silenced Mesothelioma Cells
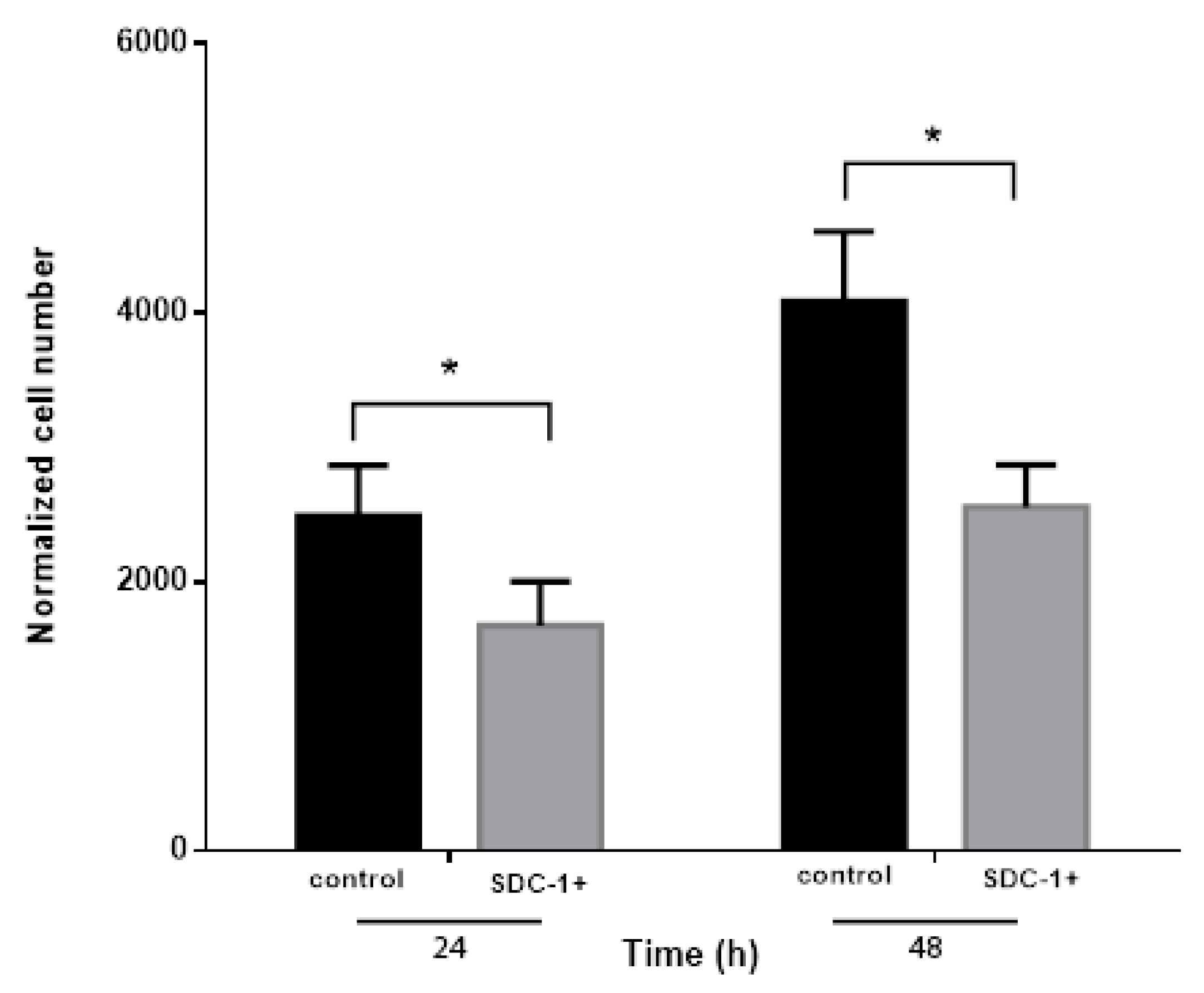
2.2. Conditioned Medium from SDC-1 Over-Expressing Mesothelioma Cells Inhibits Endothelial Cell Proliferation
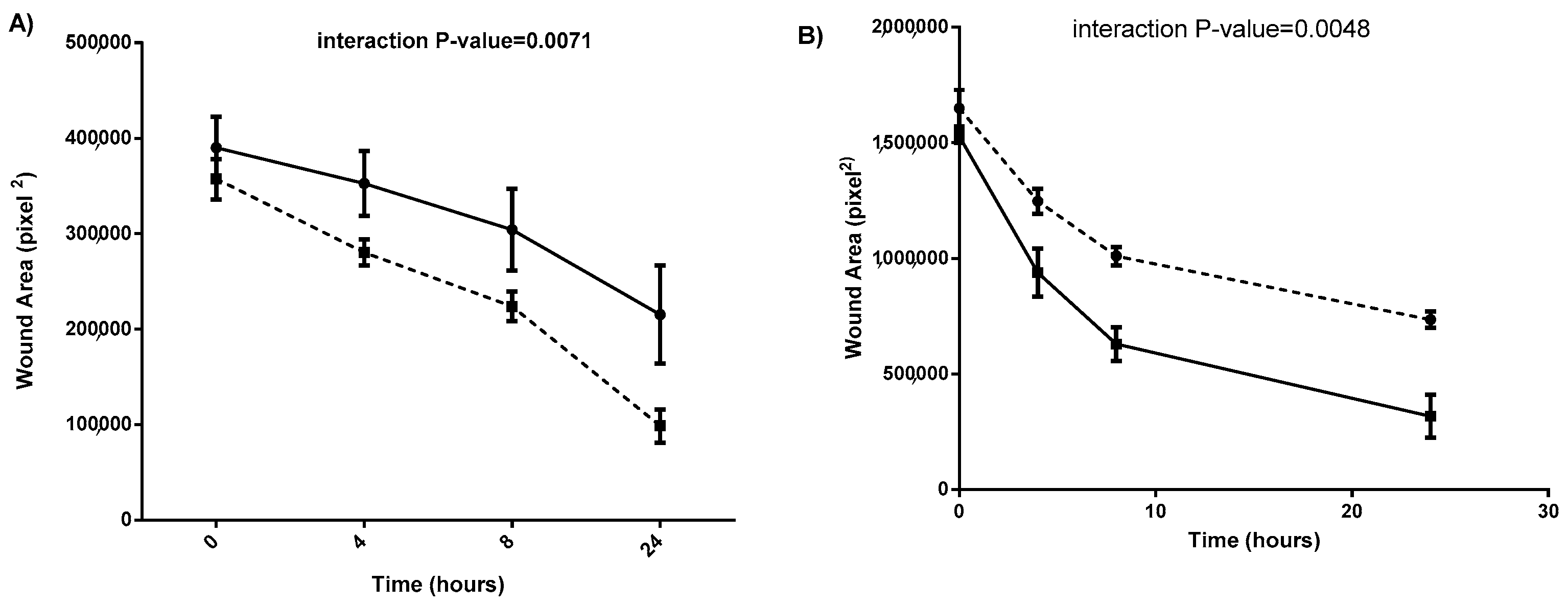
2.3. Conditioned Medium from SDC-1 Overexpressing Mesothelioma Cells Inhibits Endothelial Cell Wound Healing Assay
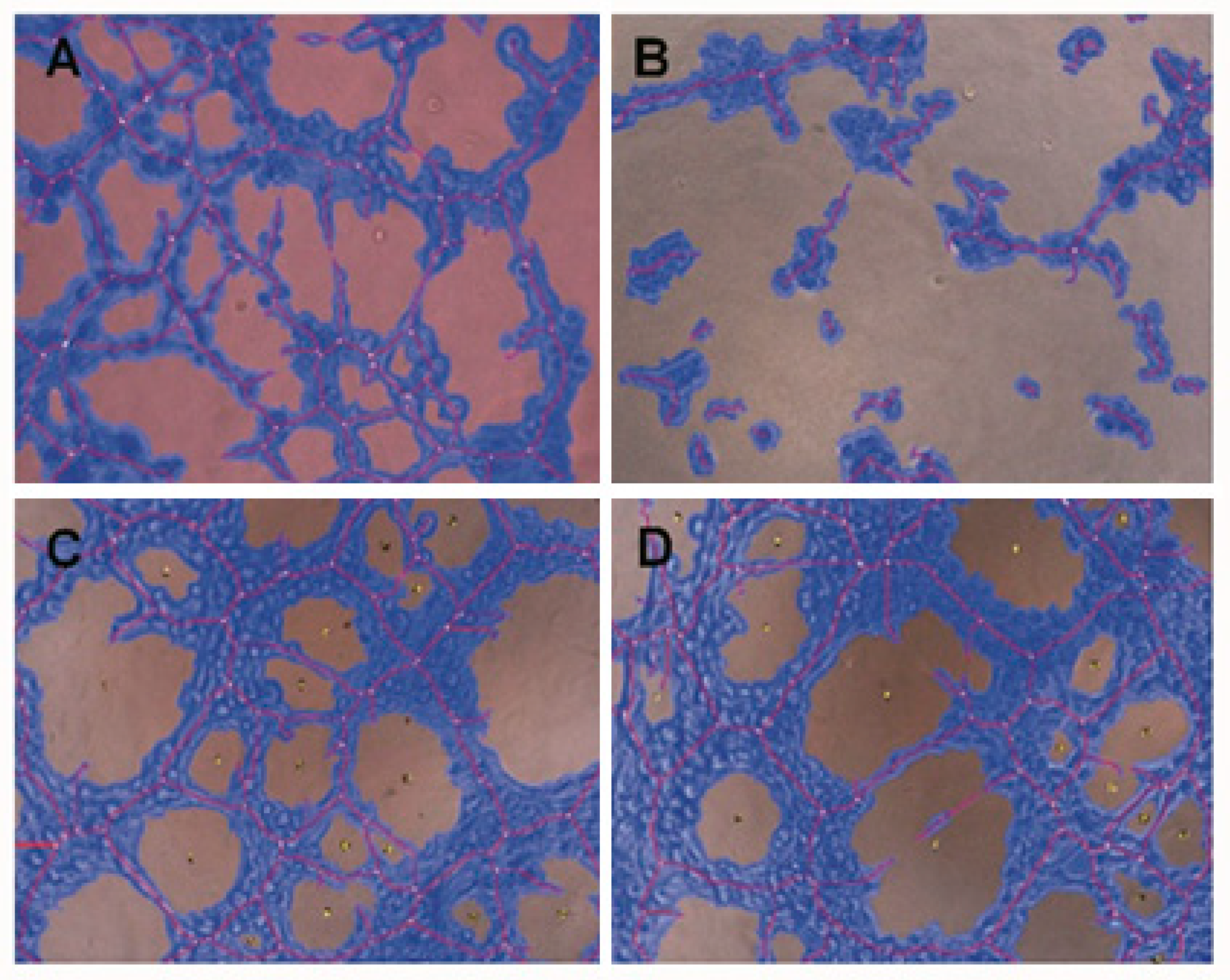
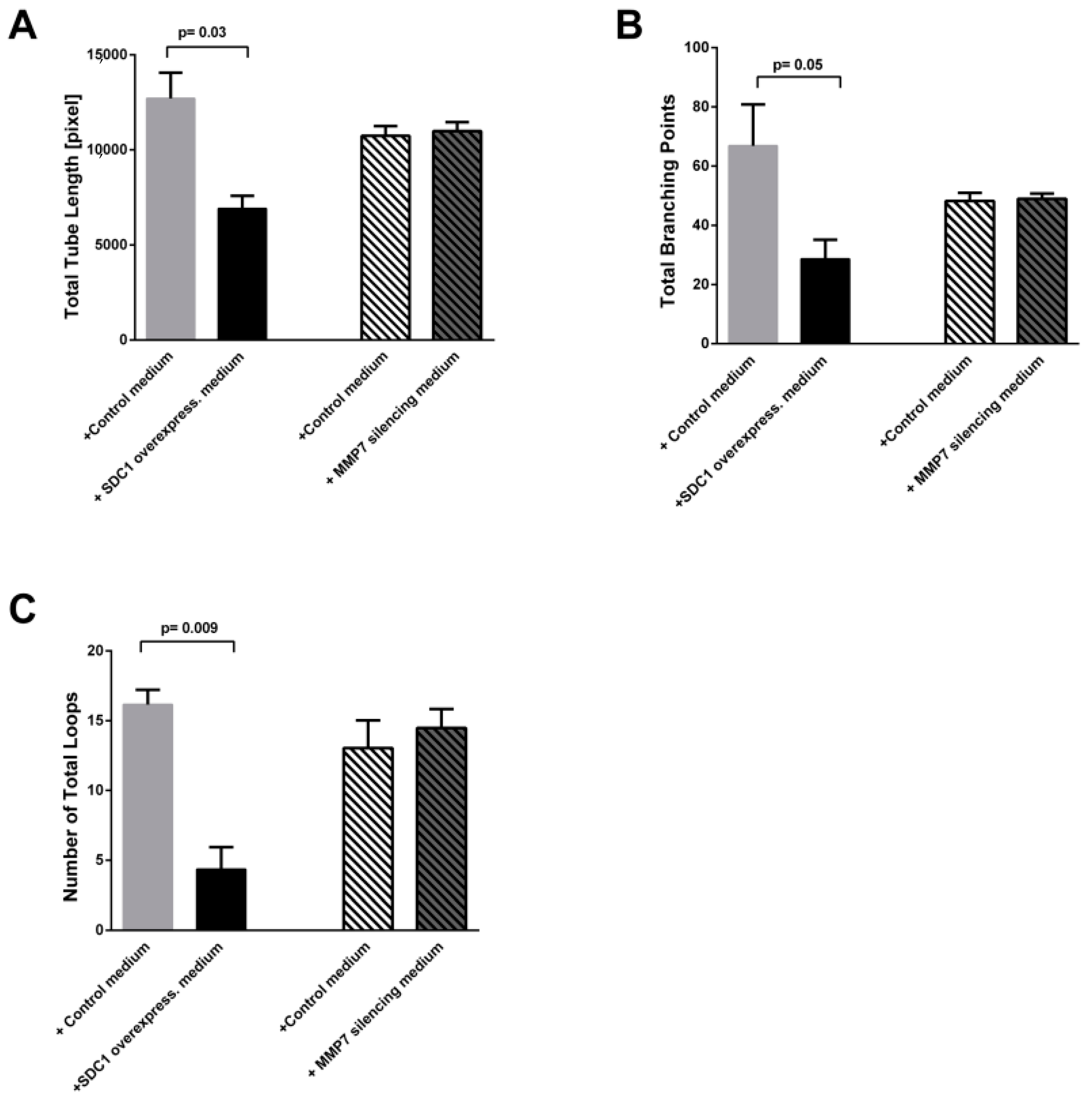
2.4. Conditioned Medium from SDC-1 Over-Expressing Cells Hampers HUVEC Cell Tubulogenesis
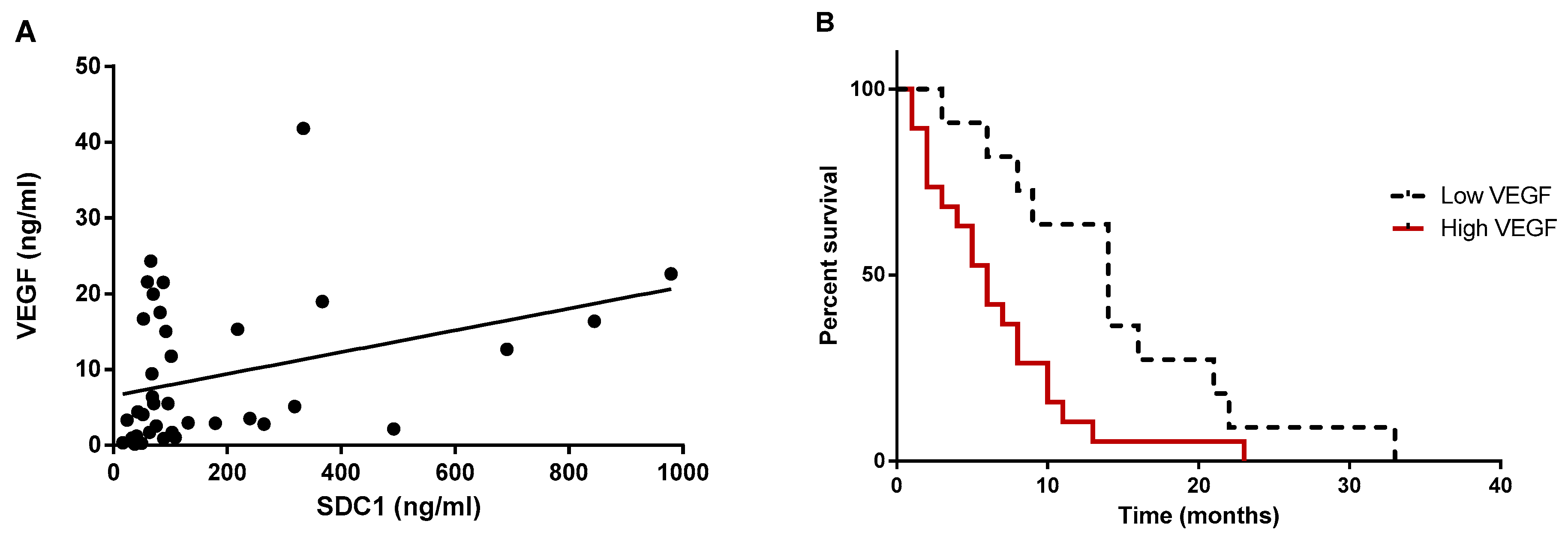
2.5. Shed SDC-1 Is Positively Correlated with VEGF in Pleural Effusions from Mesothelioma Patients
2.6. Mesothelioma Patients with Higher Levels of VEGF in Pleural Effusions Displayed Shorter Survival
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture Conditions
4.2. SDC-1 Silencing by Transfection with Small Interfering RNAs (siRNAs)
4.3. Conditioned Medium Preparation
4.4. Pleural Effusions
4.5. Proteome Profiler Human Angiogenesis Antibody Array
4.6. Endothelial Cell Proliferation Assay
4.7. Wound Healing Assay
4.8. Endothelial Cell Chemotaxis Assay
4.9. Endothelial Cell Tube Formation Assay
4.10. Interference with SDC-1 Shedding by MMP-7 Gene Silencing
4.11. Soluble SDC-1 and VEGF Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. SDC-1 Measurement by Luminex Assay with Human Premixed Multi-Analyte Kit
4.13. Fluorescence Activated Cell Sorting (FACS)
4.14. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carbone, M.; Ly, B.H.; Dodson, R.F.; Pagano, I.; Morris, P.T.; Dogan, U.A.; Gazdar, A.F.; Pass, H.I.; Yang, H. Malignant mesothelioma: Facts, Myths, and Hypotheses. J. Cell. Physiol. 2011, 227, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Bononi, A.; Napolitano, A.; Pass, H.I.; Yang, H.; Carbone, M. Latest developments in our understanding of the pathogenesis of mesothelioma and the design of targeted therapies. Expert Rev. Respir. Med. 2015, 9, 633–654. [Google Scholar] [CrossRef] [Green Version]
- Mineo, T.C.; Ambrogi, V. Malignant pleural mesothelioma: Factors influencing the prognosis. Oncology 2012, 26, 1164–1175. [Google Scholar] [PubMed]
- Van Moorst, M.; Dass, C.R. Methods for co-culturing tumour and endothelial cells: Systems and their applications. J. Pharm. Pharmacol. 2011, 63, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, A.F. Mechanisms of angiogenesis. Biochemistry 2008, 73, 751–762. [Google Scholar] [CrossRef] [PubMed]
- König, J.-E.; Tolnay, E.; Wiethege, T.; Müller, K.-M. Expression of vascular endothelial growth factor in diffuse malignant pleural mesothelioma. Virchows. Archiv. 1999, 435, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Shridhar, V.; Bright, R.K.; Kalemkerian, G.P.; Du, W.; Carbone, M.; Watanabe, Y.; Pass, H.I. VEGF and VEGF type C play an important role in angiogenesis and lymphangiogenesis in human malignant mesothelioma tumours. Br. J. Cancer 1999, 81, 54–61. [Google Scholar] [CrossRef]
- Kumar-Singh, S.; Vermeulen, P.B.; Weyler, J.; Segers, K.; Weyn, B.; van Daele, A.; Dirix, L.V.; van Oosterom, A.T.; van Marck, E. Evaluation of tumour angiogenesis as a prognostic marker in malignant mesothelioma. J. Pathol. 1997, 182, 211–216. [Google Scholar] [CrossRef]
- Kumar-Singh, S.; Weyler, J.; Martin, M.J.H.; Vermeulen, P.; Van Marck, E. Angiogenic cytokines in mesothelioma: A study of VEGF, FGF-1 and -2, and TGF? expression. J. Pathol. 1999, 189, 72–78. [Google Scholar] [CrossRef]
- Zalcman, G.; Mazieres, J.; Margery, J.; Greillier, L.; Audigier-Valette, C.; Moro-Sibilot, D.; Molinier, O.; Corre, R.; Monnet, I.; Gounant, V.; et al. Bevacizumab for newly diagnosed pleural mesothelioma in the Mesothelioma Avastin Cisplatin Pemetrexed Study (MAPS): A randomised, controlled, open-label, phase 3 trial. Lancet 2016, 387, 1405–1414. [Google Scholar] [CrossRef]
- Grosso, F.; Steele, N.; Novello, S.; Nowak, A.K.; Popat, S.; Greillier, L.; John, T.; Leighl, N.B.; Reck, M.; Taylor, P.; et al. Nintedanib Plus Pemetrexed/Cisplatin in Patients With Malignant Pleural Mesothelioma: Phase II Results From the Randomized, Placebo-Controlled LUME-Meso Trial. J. Clin. Oncol. 2017, 35, 3591–3600. [Google Scholar] [CrossRef]
- Zong, F.; Fthenou, E.; Castro, J.; Pã©Terfia, B.; Kovalszky, I.; Szilã¡k, L.; Tzanakakis, G.; Dobra, K. Effect of syndecan-1 overexpression on mesenchymal tumour cell proliferation with focus on different functional domains. Cell Prolif. 2009, 43, 29–40. [Google Scholar] [CrossRef]
- Szatmári, T.; Mundt, F.; Heidari-Hamedani, G.; Zong, F.; Ferolla, E.; Alexeyenko, A.; Hjerpe, A.; Dobra, K. Novel Genes and Pathways Modulated by Syndecan-1: Implications for the Proliferation and Cell-Cycle Regulation of Malignant Mesothelioma Cells. PLoS ONE 2012, 7, e48091. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.H.-F.; Aquino, R.S.; Park, P.W. Molecular functions of syndecan-1 in disease. Matrix Biol. 2012, 31, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Rapraeger, A.C. Synstatin: A selective inhibitor of the syndecan-1-coupled IGF1R-αvβ3 integrin complex in tumorigenesis and angiogenesis. FEBS J. 2013, 280, 2207–2215. [Google Scholar] [CrossRef] [PubMed]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Pajoohesh-Ganji, A. Syndecan-1 and Its Expanding List of Contacts. Adv. Wound Care 2015, 4, 235–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mythreye, K.; Blobe, G.C. Proteoglycan signaling co-receptors: Roles in cell adhesion, migration and invasion. Cell Sign. 2009, 21, 1548–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar-Singh, S.; Jacobs, W.; Dhaene, K.; Weyn, B.; Bogers, J.; Weyler, J.; Marck, E.V. Syndecan-1 expression in malignant mesothelioma: Correlation with cell differentiation, WT1 expression, and clinical outcome. J. Pathol. 1998, 186, 300–305. [Google Scholar] [CrossRef]
- Mundt, F.; Heidari-Hamedani, G.; Nilsonne, G.; Metintas, M.; Hjerpe, A.; Dobra, K. Diagnostic and Prognostic Value of Soluble Syndecan-1 in Pleural Malignancies. BioMed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zong, F.; Fthenou, E.; Mundt, F.; Szatmári, T.; Kovalszky, I.; Szilák, L.; Brodin, D.; Tzanakakis, G.; Hjerpe, A.; Dobra, K. Specific Syndecan-1 Domains Regulate Mesenchymal Tumor Cell Adhesion, Motility and Migration. PLoS ONE 2011, 6, e14816. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J. Endothelial-Cells and Angiogenic Growth-Factors in Cancer Growth and Metastasis—Introduction. Cancer Metastasis Rev. 1990, 9, 171–174. [Google Scholar] [CrossRef]
- Robinson, B.W.S.; Lake, R.A. Medical progress—Advances in Malignant Mesothelioma. N. Engl. J. Med. 2005, 353, 1591–1603. [Google Scholar] [CrossRef] [Green Version]
- Masood, R.; Kundra, A.; Zhu, S.; Xia, G.; Scalia, P.; Smith, D.; Gill, P.S. Malignant mesothelioma growth inhibition by agents that target the VEGF and VEGF-C autocrine loops. Int. J. Cancer 2003, 104, 603–610. [Google Scholar] [CrossRef]
- Mussap, M.; Cibecchini, F.; Noto, A.; Fanos, V. In search of biomarkers for diagnosing and managing neonatal sepsis: The role of angiopoietins. J. Matern. Fetal Neonatal Med. 2013, 26, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.N.; Tozawa, Y.; Deutsch, U.; Wolburg-Buchholz, K.; Fujiwara, Y.; Gendron-Maguire, M.; Gridley, T.; Wolburg, H.; Risau, W.; Qin, Y. Distinct roles of the receptor tyrosine kinases Tie-1 and Tie-2 in blood vessel formation. Nat. Cell Biol. 1995, 376, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Tabata, C.; Hirayama, N.; Tabata, R.; Yasumitsu, A.; Yamada, S.; Murakami, A.; Iida, S.; Tamura, K.; Fukuoka, K.; Kuribayashi, K.; et al. A novel clinical role for angiopoietin-1 in malignant pleural mesothelioma. Eur. Respir. J. 2010, 36, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Mikami, K.; Tabata, C.; Tabata, R.; Nogi, Y.; Terada, T.; Honda, M.; Kamiya, H.; Nishizaki, T.; Nakano, T. Clinical Significance of Serum Angiopoietin-1 in Malignant Peritoneal Mesothelioma. Cancer Investig. 2013, 31, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Shridhar, V.; Kalemkerian, G.P.; Bright, R.K.; Watanabe, Y.; Pass, H.I. Thrombospondin-1 expression and clinical implications in malignant pleural mesothelioma. Cancer 1999, 85, 2570–2576. [Google Scholar] [CrossRef]
- Russell, K.S.; Stern, D.F.; Polverini, P.J.; Bender, J.R. Neuregulin activation of ErbB receptors in vascular endothelium leads to angiogenesis. Am. J. Physiol. Content 1999, 277, H2205–H2211. [Google Scholar] [CrossRef]
- Mundt, F.; Johansson, H.J.; Forshed, J.; Arslan, S.; Metintas, M.; Dobra, K.; Lehtiö, J.; Hjerpe, A. Proteome Screening of Pleural Effusions Identifies Galectin 1 as a Diagnostic Biomarker and Highlights Several Prognostic Biomarkers for Malignant Mesothelioma. Mol. Cell. Proteom. 2014, 13, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Ries, C. Cytokine functions of TIMP-1. Cell. Mol. Life Sci. 2014, 71, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Bussolino, F.; Di Renzo, M.F.; Ziche, M.; Bocchietto, E.; Olivero, M.; Naldini, L.; Gaudino, G.; Tamagnone, L.; Coffer, A.; Comoglio, P.M. Hepatocyte growth factor is a potent angiogenic factor which stimulates endothelial cell motility and growth. J. Cell Biol. 1992, 119, 629–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thayaparan, T.; Spicer, J.F.; Maher, J. The role of the HGF/Met axis in mesothelioma. Biochem. Soc. Trans. 2016, 44, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, T.T.; Markkanen, J.E.; Arve, K.; Rutanen, J.; Kettunen, M.I.; Vajanto, I.; Jauhiainen, S.; Cashion, L.; Gruchala, M.; Närvänen, O.; et al. Fibroblast growth factor-4 induces vascular permeability, angiogenesis, and arteriogenesis in a rabbit hind limb ischemia model. FASEB J. 2002, 17, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Therapeutics targeting angiogenesis: Genetics and epigenetics, extracellular miRNAs and signaling networks (Review). Int. J. Mol. Med. 2013, 32, 763–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepper, M.S.; Vassalli, J.-D.; Orci, L.; Montesano, R. Biphasic Effect of Transforming Growth Factor-β1 on in Vitro Angiogenesis. Exp. Cell Res. 1993, 204, 356–363. [Google Scholar] [CrossRef]
- Chen, P.; Abacherli, L.E.; Nadler, S.T.; Wang, Y.; Li, Q.; Parks, W.C. MMP7 Shedding of Syndecan-1 Facilitates Re-Epithelialization by Affecting α2β1 Integrin Activation. PLOS ONE 2009, 4, e6565. [Google Scholar] [CrossRef] [Green Version]
- Comar, M.; Zanotta, N.; Zanconati, F.; Cortale, M.; Bonotti, A.; Cristaudo, A.; Bovenzi, M. Chemokines involved in the early inflammatory response and in pro-tumoral activity in asbestos-exposed workers from an Italian coastal area with territorial clusters of pleural malignant mesothelioma. Lung Cancer 2016, 94, 61–67. [Google Scholar] [CrossRef]
- Yasumitsu, A.; Tabata, C.; Tabata, R.; Hirayama, N.; Murakami, A.; Yamada, S.; Terada, T.; Iida, S.; Tamura, K.; Fukuoka, K.; et al. Clinical Significance of Serum Vascular Endothelial Growth Factor in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2010, 5, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, N.; Tabata, C.; Tabata, R.; Maeda, R.; Yasumitsu, A.; Yamada, S.; Kuribayashi, K.; Fukuoka, K.; Nakano, T. Pleural Effusion VEGF Levels As A Prognostic Factor Of Malignant Pleural Mesothelioma. Am. J. Respir. Crit. Care Med. 2011, 105, 137–142. [Google Scholar]
- Wang, X.; Zuo, D.; Chen, Y.; Li, W.; Liu, R.; He, Y.; Ren, L.; Zhou, L.; Deng, T.; Ying, G.; et al. Shed Syndecan-1 is involved in chemotherapy resistance via the EGFR pathway in colorectal cancer. Br. J. Cancer 2014, 111, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, A.; Uyama, T.; Kobayashi, F.; Yamada, S.; Sugahara, K.; Rapraeger, A.C.; Sanderson, R.D. Heparanase-enhanced shedding of syndecan-1 by myeloma cells promotes endothelial invasion and angiogenesis. Blood 2010, 115, 2449–2457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharbaran, R. Advances in the molecular functions of syndecan-1 (SDC1/CD138) in the pathogenesis of malignancies. Crit. Rev. Oncol. 2015, 94, 1–17. [Google Scholar] [CrossRef]
- Ramani, V.C.; Sanderson, R.D. Chemotherapy stimulates syndecan-1 shedding: A potentially negative effect of treatment that may promote tumor relapse. Matrix Biol. 2014, 35, 215–222. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javadi, J.; Heidari-Hamedani, G.; Schmalzl, A.; Szatmári, T.; Metintas, M.; Aspenström, P.; Hjerpe, A.; Dobra, K. Syndecan-1 Overexpressing Mesothelioma Cells Inhibit Proliferation, Wound Healing, and Tube Formation of Endothelial Cells. Cancers 2021, 13, 655. https://doi.org/10.3390/cancers13040655
Javadi J, Heidari-Hamedani G, Schmalzl A, Szatmári T, Metintas M, Aspenström P, Hjerpe A, Dobra K. Syndecan-1 Overexpressing Mesothelioma Cells Inhibit Proliferation, Wound Healing, and Tube Formation of Endothelial Cells. Cancers. 2021; 13(4):655. https://doi.org/10.3390/cancers13040655
Chicago/Turabian StyleJavadi, Joman, Ghazal Heidari-Hamedani, Angelika Schmalzl, Tünde Szatmári, Muzaffer Metintas, Pontus Aspenström, Anders Hjerpe, and Katalin Dobra. 2021. "Syndecan-1 Overexpressing Mesothelioma Cells Inhibit Proliferation, Wound Healing, and Tube Formation of Endothelial Cells" Cancers 13, no. 4: 655. https://doi.org/10.3390/cancers13040655