Author Contributions
J.W.T., G.W. and Y.G. designed the project and directed the study; F.L., H.A.K., D.W., J.M., Y.S., X.L., L.P., J.K., S.H.D. and K.W. performed the experiments, data analysis and interpretation; M.H., H.E., J.C., J.W.T., G.W. and Y.G. participated in the data analysis and interpretation; H.E., J.C., M.H., L.P., S.H.D., J.W.T., G.W. and Y.G. drafted and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Figure 1.
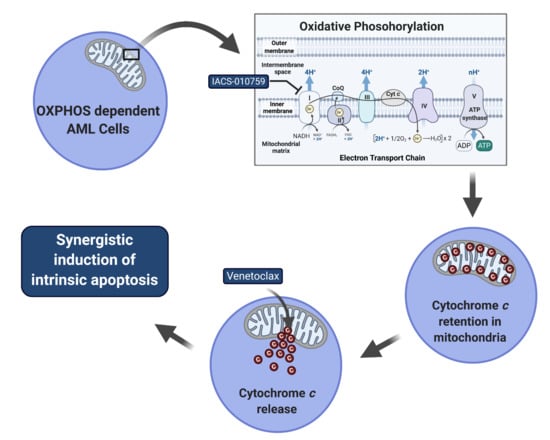
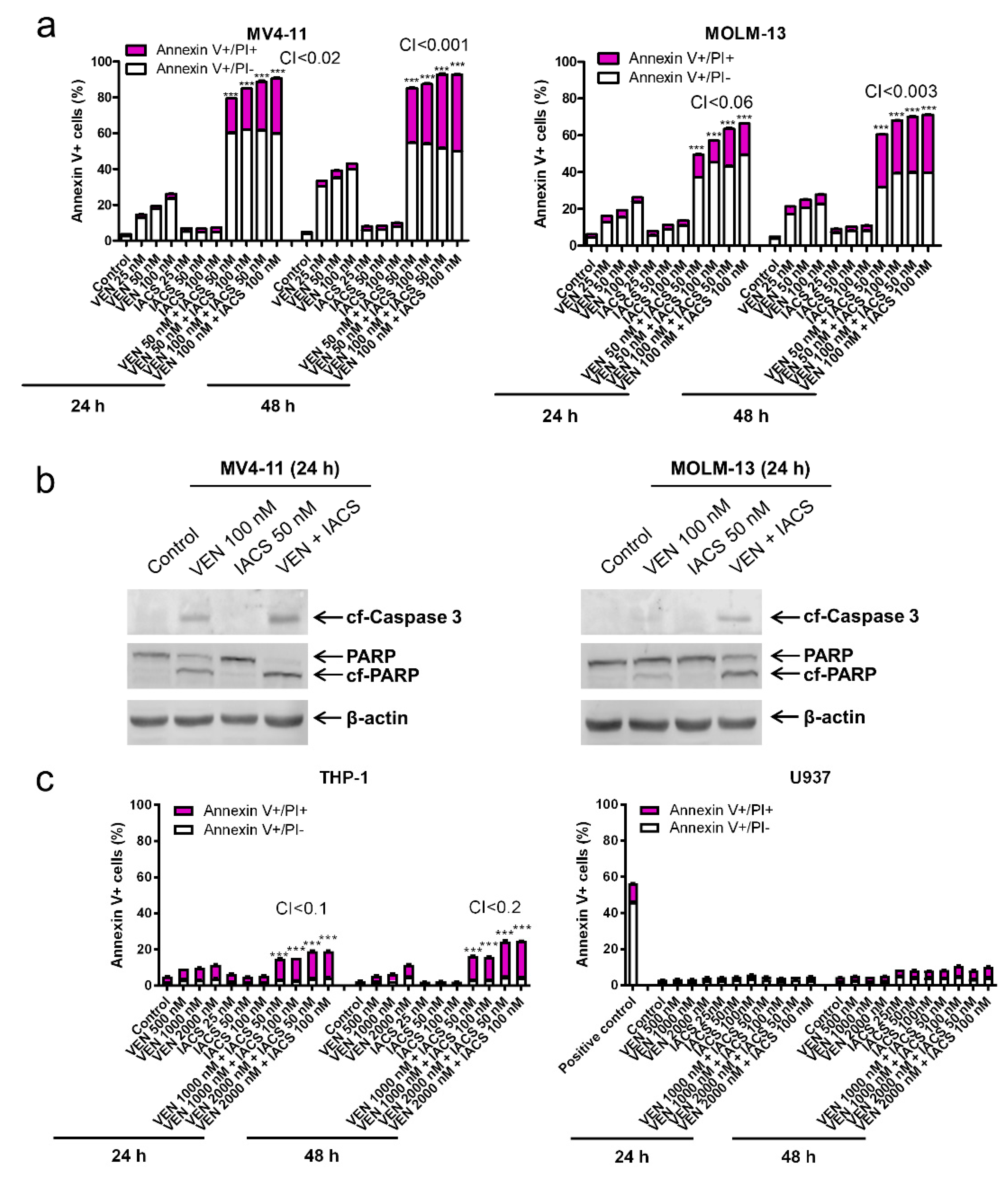
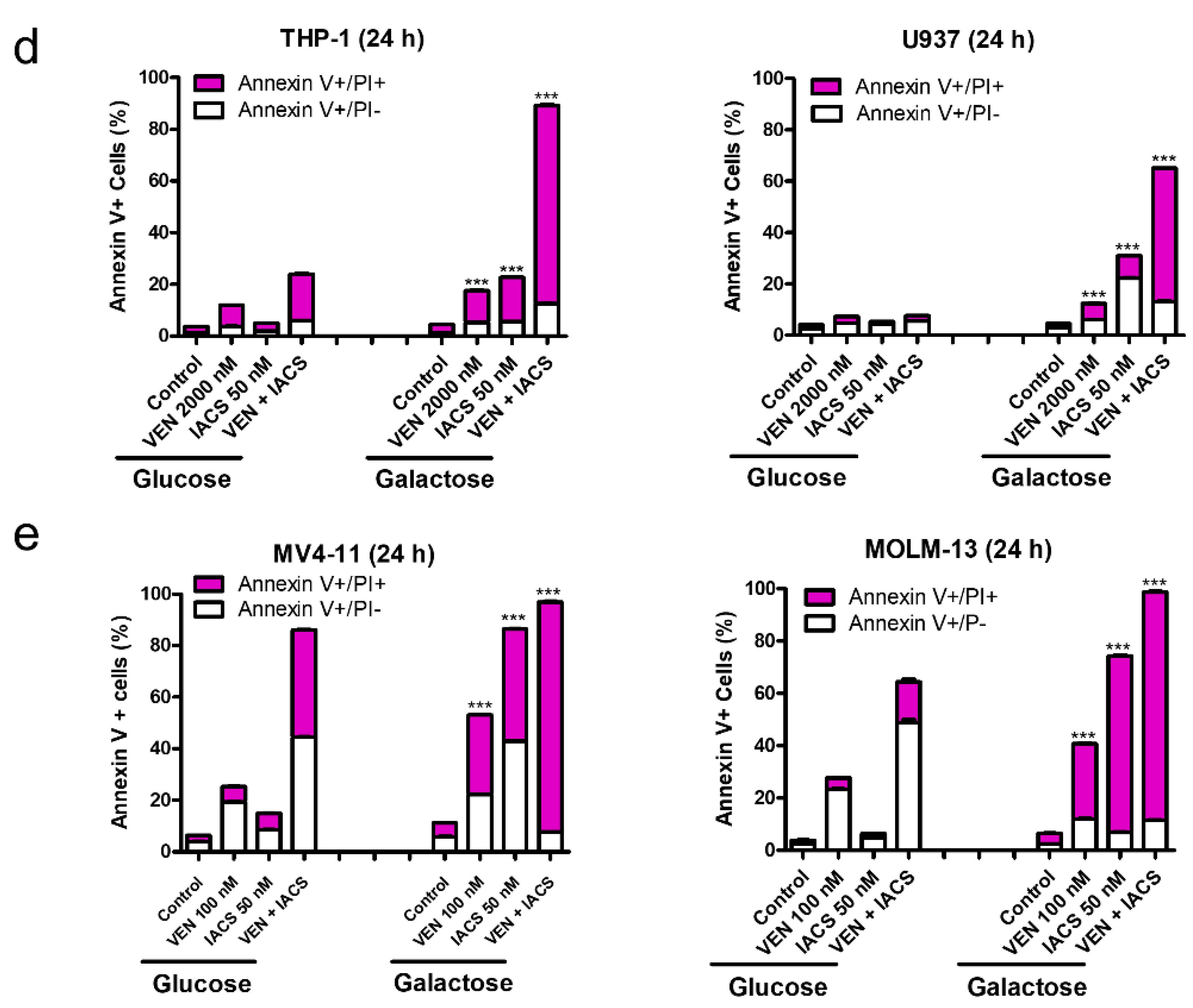
IACS-010759 synergizes with venetoclax in inducing apoptosis in oxidative phosphorylation (OXPHOS)-dependent acute myeloid leukemia (AML) cell lines. (a,c) MV4-11, MOLM-13, THP-1, and U937 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax alone or in combination for 24 or 48 h. The cells were stained with Annexin V-FITC/propidium iodide (PI) and analyzed by flow cytometry. Combination index (CI) values were calculated using CompuSyn software. CI < 1, CI = 1, CI > 1 indicate synergistic, additive, and antagonistic effect, respectively. *** indicates p < 0.001 when compared to vehicle control and individual drug treatments. (b) MV4-11 and MOLM-13 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax alone or in combination for 24 h. Whole cell lysates were subjected to western blotting. (d,e) THP-1, U937, MV4-11, and MOLM-13 cells were cultured in media containing glucose or galactose as the sole sugar source, in the absence or presence of IACS-010759 and venetoclax, for 12 h. Cells were stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 when compared to the corresponding values in cells cultured in media supplemented with glucose.
Figure 1.
IACS-010759 synergizes with venetoclax in inducing apoptosis in oxidative phosphorylation (OXPHOS)-dependent acute myeloid leukemia (AML) cell lines. (a,c) MV4-11, MOLM-13, THP-1, and U937 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax alone or in combination for 24 or 48 h. The cells were stained with Annexin V-FITC/propidium iodide (PI) and analyzed by flow cytometry. Combination index (CI) values were calculated using CompuSyn software. CI < 1, CI = 1, CI > 1 indicate synergistic, additive, and antagonistic effect, respectively. *** indicates p < 0.001 when compared to vehicle control and individual drug treatments. (b) MV4-11 and MOLM-13 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax alone or in combination for 24 h. Whole cell lysates were subjected to western blotting. (d,e) THP-1, U937, MV4-11, and MOLM-13 cells were cultured in media containing glucose or galactose as the sole sugar source, in the absence or presence of IACS-010759 and venetoclax, for 12 h. Cells were stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 when compared to the corresponding values in cells cultured in media supplemented with glucose.
![Cancers 12 02400 g001a]()
![Cancers 12 02400 g001b]()
Figure 2.
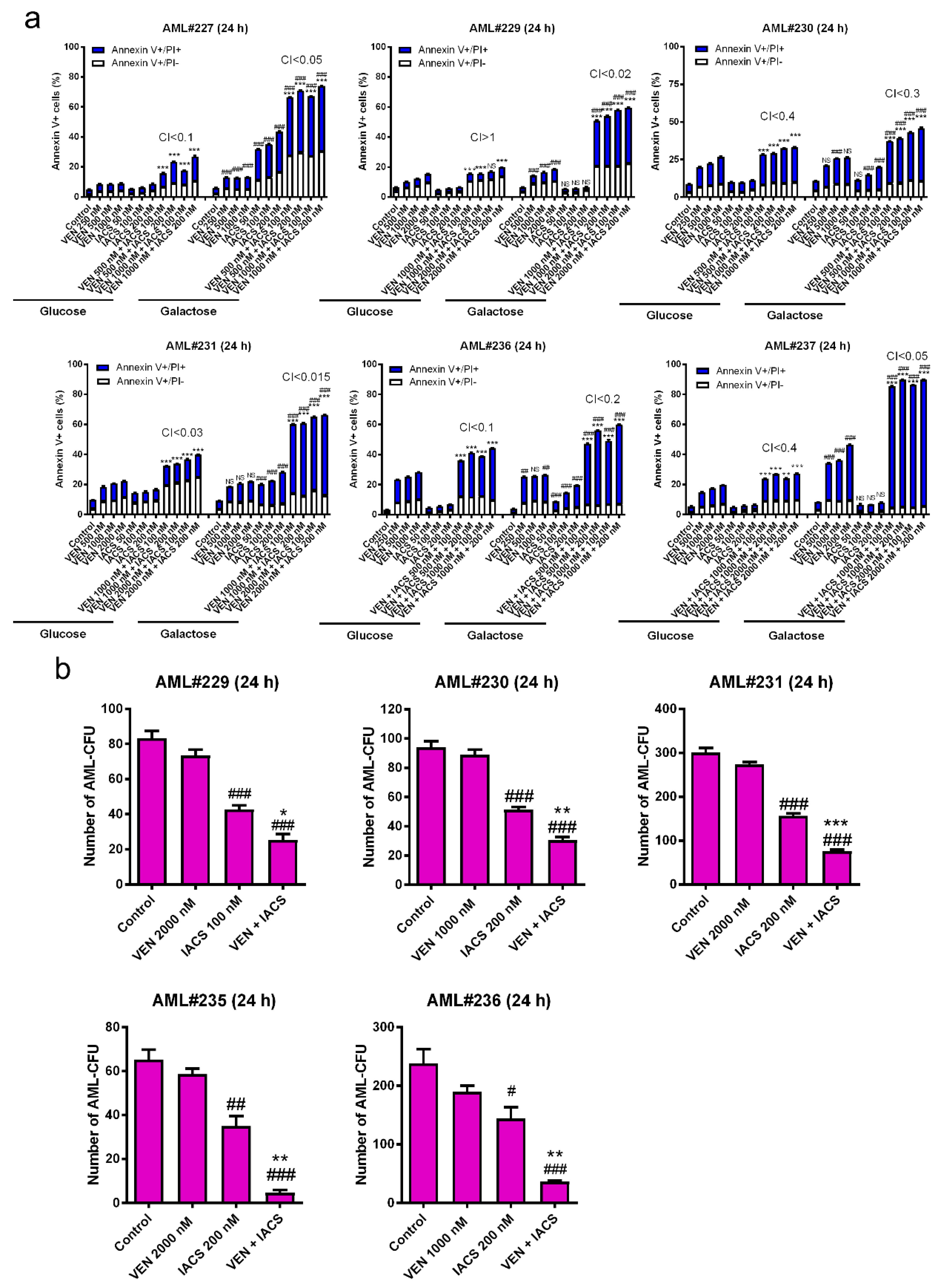
IACS-010759 and venetoclax synergistically induce cell death in primary AML cells and cooperatively reduce the colony forming capacity of AML progenitor cells. (a) AML patient samples (1 × 106 cells/condition) were treated with IACS-010759 and venetoclax alone or in combination for 24 h in the presence of glucose or galactose as the primary sugar source. The cells were split into three tubes and then stained with Annexin V-FITC/propidium iodide (PI) and analyzed by flow cytometry. Combination index (CI) values were calculated using CompuSyn software. CI < 1, CI = 1, CI > 1 indicate synergistic, additive, and antagonistic effect, respectively. *** indicates p < 0.001 when compared to vehicle control and individual drug treatment, ### indicates p < 0.001 when compared to the corresponding values in cells cultured in media supplemented with glucose, and ns indicates not significant. (b) Primary AML patient samples cultured in media supplemented with glucose were treated with vehicle control, venetoclax, IACS-010759, or in combination for 24 h (1.5 × 106 cells/treatment condition) and then plated in methylcellulose in triplicate (5 × 105 cells/plate). After incubation for 14 days, the number of surviving AML cells capable of generating leukemia colonies (AML-CFUs) were enumerated. Data are presented as mean ± SEM. # indicates p < 0.05, ## indicates p < 0.01, and ### indicates p < 0.001 compared to control. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to single drug treatments. Technical triplicates were performed.
Figure 2.
IACS-010759 and venetoclax synergistically induce cell death in primary AML cells and cooperatively reduce the colony forming capacity of AML progenitor cells. (a) AML patient samples (1 × 106 cells/condition) were treated with IACS-010759 and venetoclax alone or in combination for 24 h in the presence of glucose or galactose as the primary sugar source. The cells were split into three tubes and then stained with Annexin V-FITC/propidium iodide (PI) and analyzed by flow cytometry. Combination index (CI) values were calculated using CompuSyn software. CI < 1, CI = 1, CI > 1 indicate synergistic, additive, and antagonistic effect, respectively. *** indicates p < 0.001 when compared to vehicle control and individual drug treatment, ### indicates p < 0.001 when compared to the corresponding values in cells cultured in media supplemented with glucose, and ns indicates not significant. (b) Primary AML patient samples cultured in media supplemented with glucose were treated with vehicle control, venetoclax, IACS-010759, or in combination for 24 h (1.5 × 106 cells/treatment condition) and then plated in methylcellulose in triplicate (5 × 105 cells/plate). After incubation for 14 days, the number of surviving AML cells capable of generating leukemia colonies (AML-CFUs) were enumerated. Data are presented as mean ± SEM. # indicates p < 0.05, ## indicates p < 0.01, and ### indicates p < 0.001 compared to control. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to single drug treatments. Technical triplicates were performed.
![Cancers 12 02400 g002]()
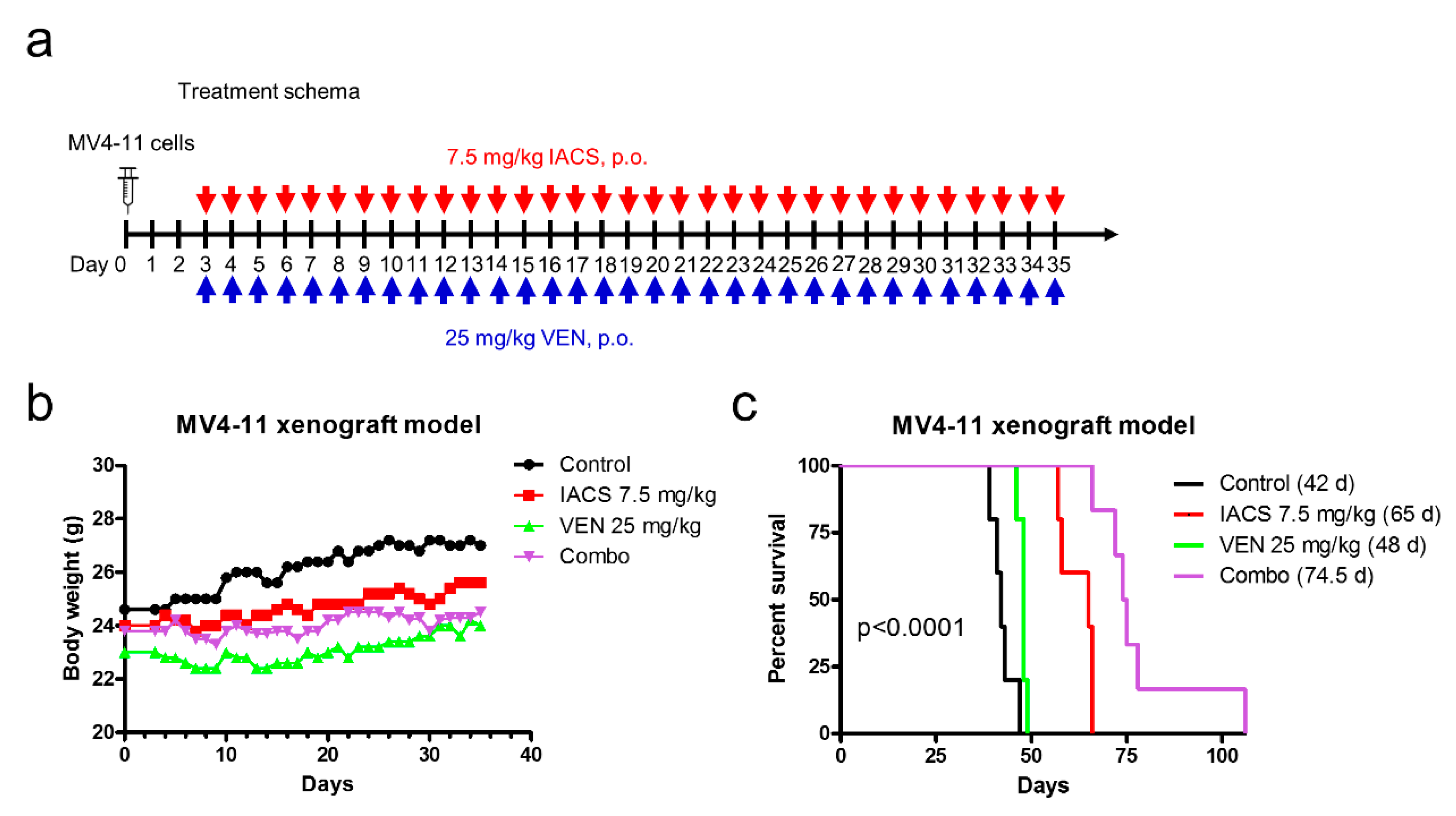
Figure 3.
IACS-010759 and venetoclax combination shows in vivo efficacy in an AML cell line-derived xenograft mouse model. (a) NSGS mice were injected with MV4-11 cells (1 × 106 cells/mouse, via tail vein injection). Three days post cell injection the mice were randomized and treated. Vehicle control, 7.5 mg/kg IACS-010759, 25 mg/kg venetoclax, or 7.5 mg/kg IACS-010759 + 25 mg/kg venetoclax (n = 5) were administered on a daily basis via oral gavage for a total of 33 days. (b,c) Average mouse body weights were measured on a daily basis and graphed (panel b). Overall survival probability, estimated with the Kaplan-Meier method, is shown in panel c.
Figure 3.
IACS-010759 and venetoclax combination shows in vivo efficacy in an AML cell line-derived xenograft mouse model. (a) NSGS mice were injected with MV4-11 cells (1 × 106 cells/mouse, via tail vein injection). Three days post cell injection the mice were randomized and treated. Vehicle control, 7.5 mg/kg IACS-010759, 25 mg/kg venetoclax, or 7.5 mg/kg IACS-010759 + 25 mg/kg venetoclax (n = 5) were administered on a daily basis via oral gavage for a total of 33 days. (b,c) Average mouse body weights were measured on a daily basis and graphed (panel b). Overall survival probability, estimated with the Kaplan-Meier method, is shown in panel c.
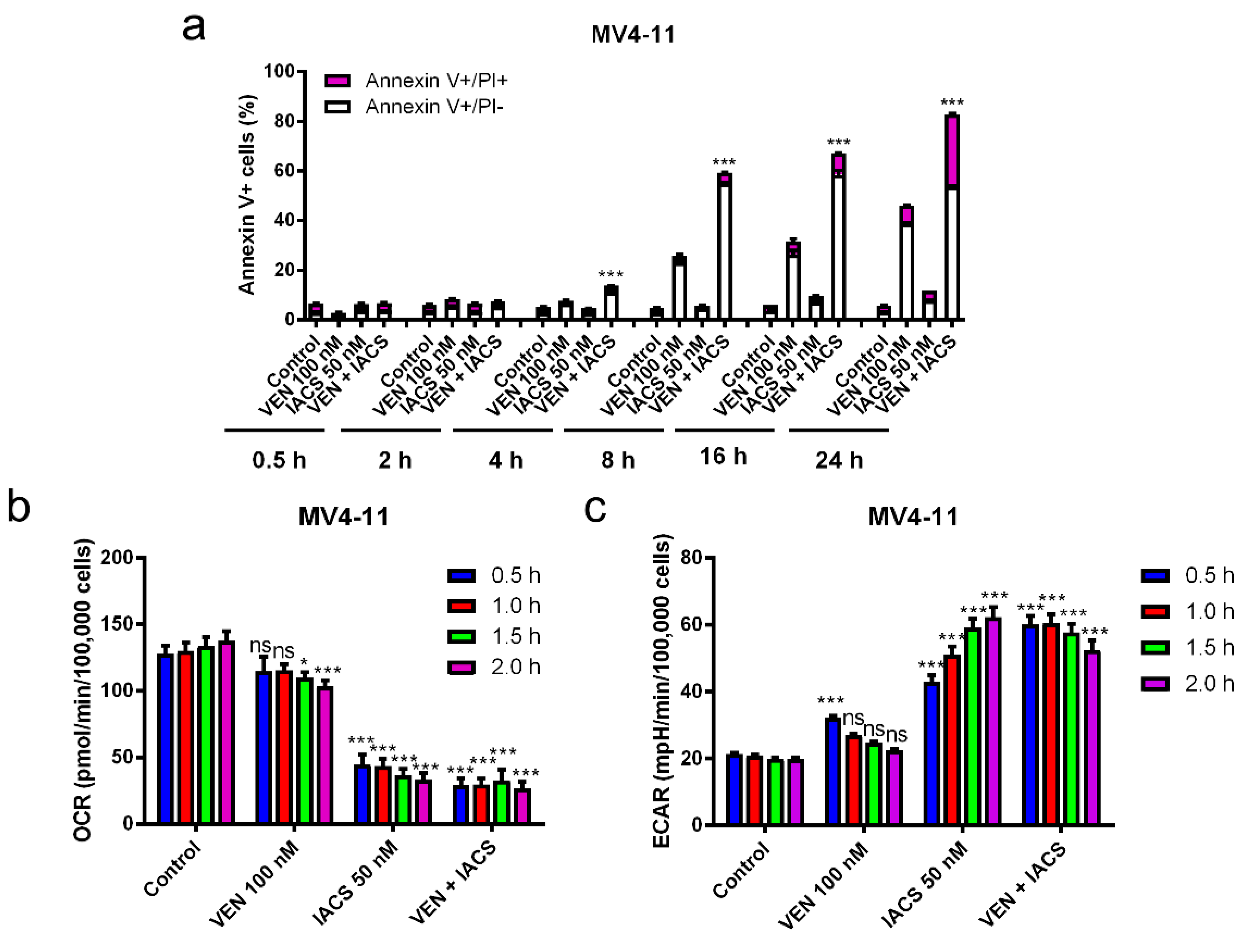
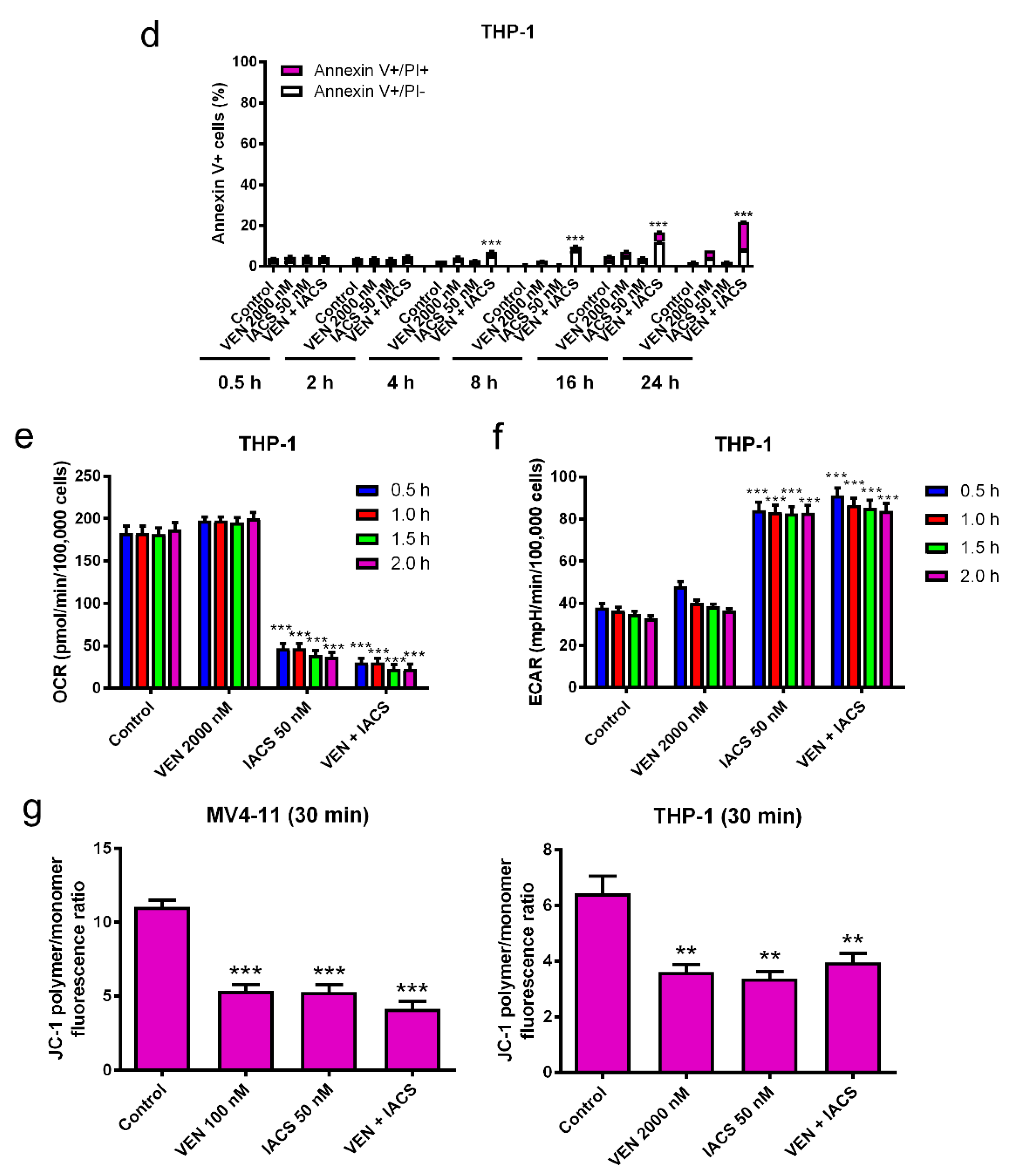
Figure 4.
Inhibition of OXPHOS by IACS-010759 treatment is not enhanced by venetoclax. (a,d) MV4-11 and THP-1 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax alone and in combination for up to 24 h. Cells were then stained with Annexin V/PI and analyzed by flow cytometry. ** indicates p < 0.01 and *** indicates p < 0.001 compared to vehicle control and single drug treatments. (b,c,e,f) MV4-11 and THP-1 cells cultured in media supplemented with glucose were plated in Seahorse media and incubated in a Seahorse analyzer. IACS-010759 and venetoclax were injected into the cell culture while in the Seahorse analyzer. Oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) were measured every 30 min for up to 2 h after injection of the drugs. * indicates p < 0.05, *** indicates p < 0.001, and ns indicates not significant compared to vehicle control. (g) MV4-11 and THP-1 cells cultured in media supplemented with glucose were treated with vehicle control, venetoclax, IACS-010759, or in combination for 30 min and then the cells were subjected to the JC-1 assay. ** indicates p < 0.01 and *** indicates p < 0.001 compared to vehicle control.
Figure 4.
Inhibition of OXPHOS by IACS-010759 treatment is not enhanced by venetoclax. (a,d) MV4-11 and THP-1 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax alone and in combination for up to 24 h. Cells were then stained with Annexin V/PI and analyzed by flow cytometry. ** indicates p < 0.01 and *** indicates p < 0.001 compared to vehicle control and single drug treatments. (b,c,e,f) MV4-11 and THP-1 cells cultured in media supplemented with glucose were plated in Seahorse media and incubated in a Seahorse analyzer. IACS-010759 and venetoclax were injected into the cell culture while in the Seahorse analyzer. Oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) were measured every 30 min for up to 2 h after injection of the drugs. * indicates p < 0.05, *** indicates p < 0.001, and ns indicates not significant compared to vehicle control. (g) MV4-11 and THP-1 cells cultured in media supplemented with glucose were treated with vehicle control, venetoclax, IACS-010759, or in combination for 30 min and then the cells were subjected to the JC-1 assay. ** indicates p < 0.01 and *** indicates p < 0.001 compared to vehicle control.
![Cancers 12 02400 g004a]()
![Cancers 12 02400 g004b]()
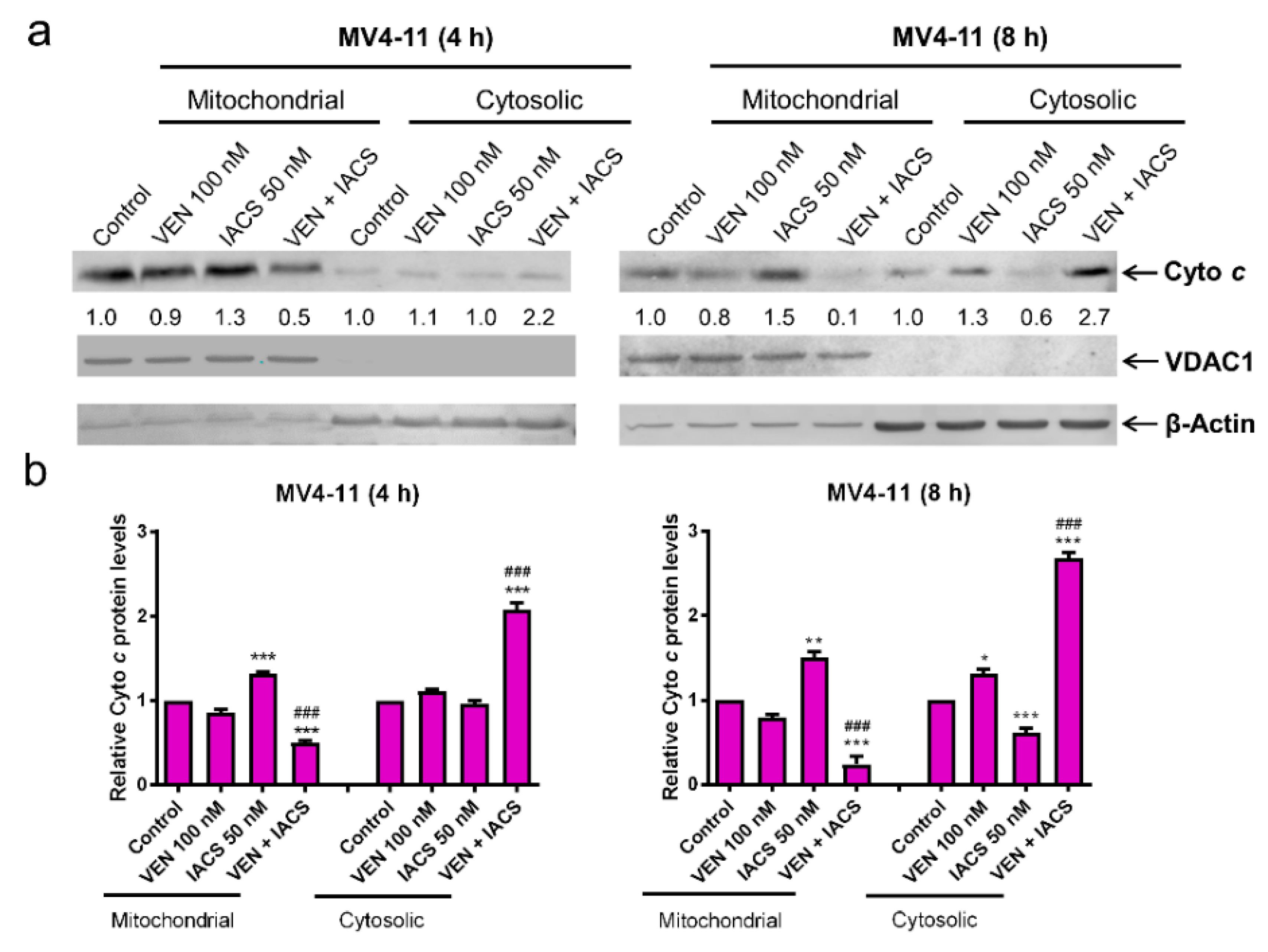
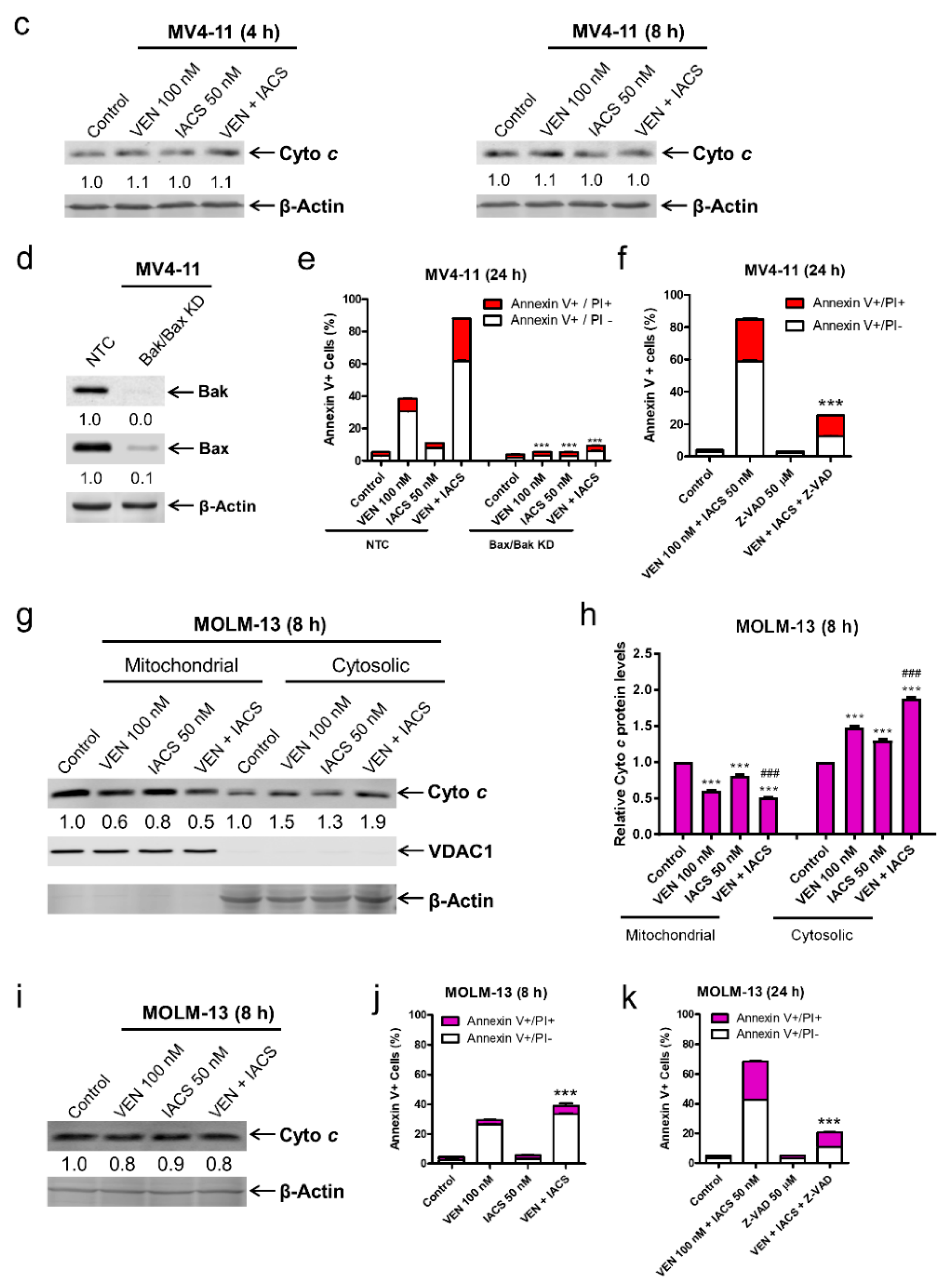
Figure 5.
IACS-010759 and venetoclax cooperatively induce intrinsic apoptosis. (a,b) MV4-11 cells cultured in media supplemented with glucose were treated with vehicle control, IACS-010759, venetoclax, or in combination for 4 or 8 h. Cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel b. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to vehicle control; ### indicates p < 0.001 compared to single drug treatments. (c) MV4-11 cells were cultured and treated as described in panel a. Whole cell lysates were subjected to western blot analysis. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin, and compared to the vehicle control. (d,e) Lentiviral shRNA double knockdown of Bak and Bax (designated Bak/Bax KD) was performed in MV4-11 cells. Non-template control (NTC)-shRNA was used as the control for the Bak/Bax double knockdown. Western blots probed with anti-Bak, -Bax, or -β-actin antibody are shown in panel d. shRNA knockdown cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax, alone or combined, for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the NTC-shRNA cells. (f) MV4-11 cells cultured in media supplemented with glucose were treated with combined IACS-010759 and venetoclax in the absence or presence of the pan-caspase inhibitor Z-VAD-FMK for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the combined venetoclax and IACS-010759 treatment. (g,h) MOLM-13 cells cultured in media supplemented with glucose were treated with vehicle control, IACS-010759, venetoclax, or in combination for 8 h. (i) Cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel h. *** indicates p < 0.001 compared to vehicle control; ### indicates p < 0.001 compared to single drug treatments. (j) MOLM-13 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax, alone or combined, for 8 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the vehicle control and single drug treatments. (k) MOLM-13 cells cultured in media supplemented with glucose were treated with combined IACS-010759 and venetoclax in the absence or presence of the pan-caspase inhibitor Z-VAD-FMK for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the combined venetoclax and IACS-010759 treatment.
Figure 5.
IACS-010759 and venetoclax cooperatively induce intrinsic apoptosis. (a,b) MV4-11 cells cultured in media supplemented with glucose were treated with vehicle control, IACS-010759, venetoclax, or in combination for 4 or 8 h. Cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel b. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to vehicle control; ### indicates p < 0.001 compared to single drug treatments. (c) MV4-11 cells were cultured and treated as described in panel a. Whole cell lysates were subjected to western blot analysis. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin, and compared to the vehicle control. (d,e) Lentiviral shRNA double knockdown of Bak and Bax (designated Bak/Bax KD) was performed in MV4-11 cells. Non-template control (NTC)-shRNA was used as the control for the Bak/Bax double knockdown. Western blots probed with anti-Bak, -Bax, or -β-actin antibody are shown in panel d. shRNA knockdown cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax, alone or combined, for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the NTC-shRNA cells. (f) MV4-11 cells cultured in media supplemented with glucose were treated with combined IACS-010759 and venetoclax in the absence or presence of the pan-caspase inhibitor Z-VAD-FMK for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the combined venetoclax and IACS-010759 treatment. (g,h) MOLM-13 cells cultured in media supplemented with glucose were treated with vehicle control, IACS-010759, venetoclax, or in combination for 8 h. (i) Cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel h. *** indicates p < 0.001 compared to vehicle control; ### indicates p < 0.001 compared to single drug treatments. (j) MOLM-13 cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax, alone or combined, for 8 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the vehicle control and single drug treatments. (k) MOLM-13 cells cultured in media supplemented with glucose were treated with combined IACS-010759 and venetoclax in the absence or presence of the pan-caspase inhibitor Z-VAD-FMK for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the combined venetoclax and IACS-010759 treatment.
![Cancers 12 02400 g005a]()
![Cancers 12 02400 g005b]()
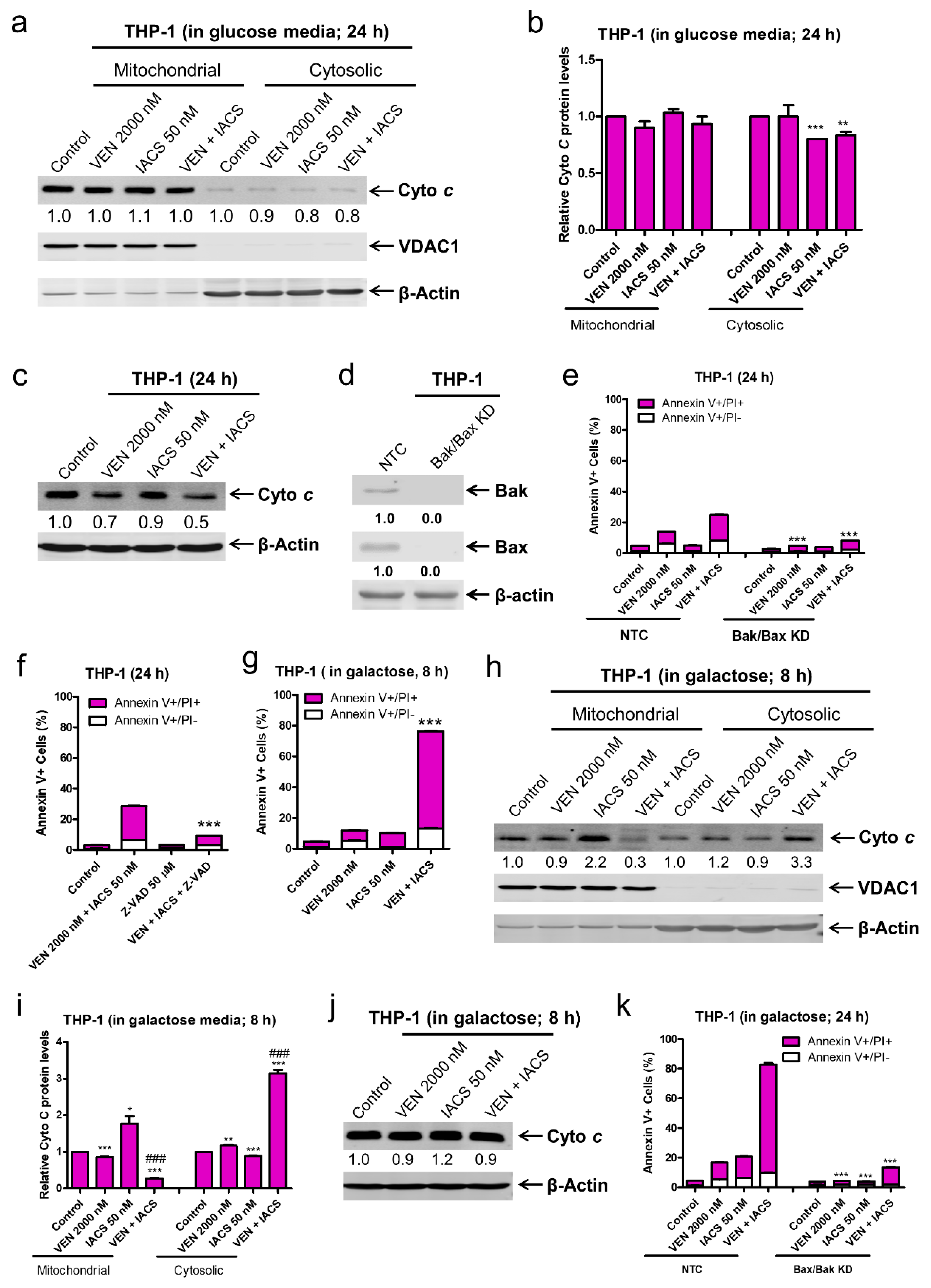
Figure 6.
Forcing glycolysis-reliant AML cells to utilize OXPHOS renders them susceptibility to IACS-010759 and venetoclax induced intrinsic apoptosis mediated by cytochrome c release. (a,b) THP-1 cells, cultured in media supplemented with glucose, were treated with vehicle control, IACS-010759, venetoclax, or in combination for 24 h. Cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to Western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel b. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to vehicle control. (c) THP-1 cells were cultured and treated as described in panel a. Whole cell lysates were subjected to Western blot analysis. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin, and compared to the vehicle control. (d,e) Lentiviral shRNA double knockdown of Bak and Bax (designated Bak/Bax KD) was performed in THP-1 cells. Non-template control (NTC)-shRNA was used as the control for the Bak/Bax double knockdown. Western blots probed with anti-Bak, -Bax, or -β-actin antibody are shown in panel d. shRNA knockdown cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax, alone or combined, for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the NTC-shRNA cells. (f) THP-1 cells cultured in media supplemented with glucose were treated with combined IACS-010759 and venetoclax in the absence or presence of the pan-caspase inhibitor Z-VAD-FMK for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the combined venetoclax and IACS-010759 treatment. (g–i) THP-1 cells cultured in media supplemented with galactose were treated with vehicle control, IACS-010759, venetoclax, or in combination for 8 h. In panel g, cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the vehicle control and single drug treatments. For panels (h,i), cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to Western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel i. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to vehicle control, ### indicates p < 0.001. (j) THP-1 cells were cultured and treated as described in panel (g). Whole cell lysates were subjected to Western blot analysis. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin, and compared to the vehicle control. (k) THP-1 NTC and Bak/Bax double knockdown cells cultured in media supplemented with galactose were treated with IACS-010759 and venetoclax, alone or combined, for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the NTC-shRNA cells.
Figure 6.
Forcing glycolysis-reliant AML cells to utilize OXPHOS renders them susceptibility to IACS-010759 and venetoclax induced intrinsic apoptosis mediated by cytochrome c release. (a,b) THP-1 cells, cultured in media supplemented with glucose, were treated with vehicle control, IACS-010759, venetoclax, or in combination for 24 h. Cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to Western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel b. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to vehicle control. (c) THP-1 cells were cultured and treated as described in panel a. Whole cell lysates were subjected to Western blot analysis. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin, and compared to the vehicle control. (d,e) Lentiviral shRNA double knockdown of Bak and Bax (designated Bak/Bax KD) was performed in THP-1 cells. Non-template control (NTC)-shRNA was used as the control for the Bak/Bax double knockdown. Western blots probed with anti-Bak, -Bax, or -β-actin antibody are shown in panel d. shRNA knockdown cells cultured in media supplemented with glucose were treated with IACS-010759 and venetoclax, alone or combined, for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the NTC-shRNA cells. (f) THP-1 cells cultured in media supplemented with glucose were treated with combined IACS-010759 and venetoclax in the absence or presence of the pan-caspase inhibitor Z-VAD-FMK for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the combined venetoclax and IACS-010759 treatment. (g–i) THP-1 cells cultured in media supplemented with galactose were treated with vehicle control, IACS-010759, venetoclax, or in combination for 8 h. In panel g, cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the vehicle control and single drug treatments. For panels (h,i), cellular fractionation was performed. Mitochondrial and cytosolic fractions were subjected to Western blot analysis. This experiment was performed 2 independent times in triplicate. One representative image is shown. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin or VDAC1, and compared to the vehicle control. Results from one representative experiment are graphed as mean ± SEM in panel i. * indicates p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001 compared to vehicle control, ### indicates p < 0.001. (j) THP-1 cells were cultured and treated as described in panel (g). Whole cell lysates were subjected to Western blot analysis. Relative densitometry measurements were determined using Odyssey Software V3.0, normalized to β-actin, and compared to the vehicle control. (k) THP-1 NTC and Bak/Bax double knockdown cells cultured in media supplemented with galactose were treated with IACS-010759 and venetoclax, alone or combined, for 24 h. Cells were then stained with Annexin V-FITC/PI and analyzed by flow cytometry. *** indicates p < 0.001 compared to the NTC-shRNA cells.
![Cancers 12 02400 g006]()
Table 1.
Patient characteristics of primary AML patient samples.
Table 1.
Patient characteristics of primary AML patient samples.
| Patient | Gender | Age (Year) | Disease Status | CYTOGENETICS | Blast Purity (%) | Gene Mutation |
|---|
| AML#227 | Male | 32 | Newly diagnosed | 47, XY,+14, inv(16)(p13q22)/46, XY, inv(16)(p13q22)/46, XY | 87.5 | PRAME, WT1, JAK2, NRAS |
| AML#229 | Male | 9 | Newly diagnosed | 46, XY, t(15,17)(q24;q21)/46, XY | 92.0 | PML-RARαL, EP300, KRAS |
| AML#230 | Female | 16 | Newly diagnosed | 46, XX, t(8;21)(q22;q22)/46, XX | 59.0 | AML-ETO, WT1, PRAME |
| AML#231 | Male | 17 | Newly diagnosed | 46, XY, der(19)t(1;19)(q23;p13), + 21 | 74.0 | ND |
| AML#235 | Female | 33 | Newly diagnosed | 46, XX | 87.0 | NPM1 |
| AML#236 | Male | 26 | Newly diagnosed | 46, XY, inv(17)(q10), t(15;17)(q24;21) | 88 | WT1, PRAME, PML-RARα(+) |
| AML#237 | Male | 56 | Relapsed | 46, XY | 88.0 | NA |

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}