MicroRNA Regulation of the Small Rho GTPase Regulators—Complexities and Opportunities in Targeting Cancer Metastasis


1
Center for Molecular Imaging, Department of Radiology, University of Michigan, 109 Zina Pitcher Place, Ann Arbor, MI 48109, USA
2
Department of Toxicology and Cancer Biology, College of Medicine, University of Kentucky, 1095 V A Drive, Lexington, KY 40536, USA
*
Authors to whom correspondence should be addressed.
Cancers 2020, 12(5), 1092; https://doi.org/10.3390/cancers12051092
Submission received: 25 March 2020
/
Revised: 24 April 2020
/
Accepted: 25 April 2020
/
Published: 28 April 2020
(This article belongs to the Special Issue Rho Family of GTPases in Cancer)
Abstract
:The small Rho GTPases regulate important cellular processes that affect cancer metastasis, such as cell survival and proliferation, actin dynamics, adhesion, migration, invasion and transcriptional activation. The Rho GTPases function as molecular switches cycling between an active GTP-bound and inactive guanosine diphosphate (GDP)-bound conformation. It is known that Rho GTPase activities are mainly regulated by guanine nucleotide exchange factors (RhoGEFs), GTPase-activating proteins (RhoGAPs), GDP dissociation inhibitors (RhoGDIs) and guanine nucleotide exchange modifiers (GEMs). These Rho GTPase regulators are often dysregulated in cancer; however, the underlying mechanisms are not well understood. MicroRNAs (miRNAs), a large family of small non-coding RNAs that negatively regulate protein-coding gene expression, have been shown to play important roles in cancer metastasis. Recent studies showed that miRNAs are capable of directly targeting RhoGAPs, RhoGEFs, and RhoGDIs, and regulate the activities of Rho GTPases. This not only provides new evidence for the critical role of miRNA dysregulation in cancer metastasis, it also reveals novel mechanisms for Rho GTPase regulation. This review summarizes recent exciting findings showing that miRNAs play important roles in regulating Rho GTPase regulators (RhoGEFs, RhoGAPs, RhoGDIs), thus affecting Rho GTPase activities and cancer metastasis. The potential opportunities and challenges for targeting miRNAs and Rho GTPase regulators in treating cancer metastasis are also discussed. A comprehensive list of the currently validated miRNA-targeting of small Rho GTPase regulators is presented as a reference resource.
Keywords:
Rho GTPases; Rho GTPase regulators; RhoGEFs; RhoGAPs; RhoGDIs; microRNAs; cancer; metastasis1. Introduction
Cancer progression is highlighted by changes in cancer cells that promote aggressiveness allowing cells to acquire a greater metastatic potential. Once cancer cells in the primary tumor gain the ability to invade the surrounding tissue, motile cells pass through the basement membrane and the extracellular matrix (ECM) penetrating into the lymphatic or vascular circulation. These motile cells travel through the circulatory system until they arrest at a different locations, extravasate through the vascular basement membrane and the ECM into the new environment where they gain epithelial characteristics and form a secondary or metastatic lesion. Because metastasis is the leading cause of mortality in cancer patients, recent research has focused on identifying and understanding the underlying mechanisms that contribute to metastasis. Numerous studies demonstrated that small Rho GTPases are key regulators of cell adhesion, migration and invasion, thus playing crucial roles in cancer metastasis (for reviews see [1,2,3]). It is well established that the activities of small Rho GTPases are tightly regulated mainly by the following four groups of regulators: guanine nucleotide exchange factors (GEFs), GTPase-activating proteins (GAPs), guanosine diphosphate (GDP) dissociation inhibitors (GDIs) and guanine nucleotide exchange modifiers (GEMs) [4,5,6,7]. However, much less is known about how the activities of small Rho GTPase regulators are regulated.
Although elucidating the underlying mechanisms of cancer metastasis has been the focus for many years, the connection between microRNAs (miRNAs), a family of small non-coding RNAs, and Rho GTPase regulators has only recently become a focused topic in cancer metastasis studies. There is a growing body of evidence revealing the critical involvement of miRNAs in the tight spatiotemporal regulation of actin-based physiology. Moreover, depending on the specific context, miRNAs can have a tumor suppressive or oncogenic role in cancer. We understand that miRNAs can directly regulate the expression of Rho GTPases and this was reviewed elsewhere [8]. In this review, we focused on recent exciting findings showing that miRNAs play important roles in regulating Rho GTPase regulators (RhoGEFs, RhoGAPs, RhoGDIs), eventually affecting small Rho GTPase activities and cancer metastasis. A comprehensive list of the currently validated miRNA-targeting of small Rho GTPase regulators is presented.
2. MicroRNA Biogenesis and Function
Although the basic features of microRNA biogenesis and its mechanism of action were established over a decade ago [9,10,11], subsequent years have shown a vast accumulation of new information that has not only deciphered the mechanistic details, but has also demonstrated that miRNAs are key regulatory hubs for cancer. Here, we provide only a brief introduction to miRNA biogenesis and function for context as we discuss their direct role in modulating mechanisms that contribute to cancer progression (we have previously reviewed miRNA biogenesis in more detail [12,13,14]).
MicroRNAs (miRNAs or miRs), are a subclass of small (~21–23 nucleotides) non-coding RNA molecules that negatively regulate protein-coding gene expression. In terms of biogenesis (Figure 1), a functionally mature miRNA is derived from the cleavage of a double-stranded ~70 nt RNA hairpin precursor in the cytosol. These miRNA precursors are typically located either within the introns of a host protein-coding gene or in intergenic regions, and are transcribed in the nucleus by either RNA polymerase II or III. However, the cases in which miRNA precursors were found within the exons of transcripts and in antisense transcripts have been reported [15,16]. Once excised from the precursor RNA hairpin, a mature miRNA is then loaded into the RNA-induced silencing complex (RISC), where miRNAs are then able to negatively regulate the expression of target genes. Functionally, miRNAs elicit this negative regulation typically by imperfect base pairing with the 3′ untranslated region (3′UTR) of the target messenger RNA (mRNA) through the miRNA seed sequence. The seed sequence is the second to eighth nucleotide at the 5′ end of a mature miRNA. The binding of a miRNA to its target mRNAs can induce mRNA degradation, translational inhibition or direct cleavage, depending on the sequence complementarity [11,17,18,19]. The expression of miRNAs can be regulated through interactions with and the modifications of their promoters. In addition to the promoter-mediated control of expression, miRNA function can be controlled through the binding and sequestration of the mature miRNA in the cytosol by long non-coding RNAs (lncRNAs) as well as circular ncRNAs (circRNAs), termed competing endogenous RNAs (ceRNAs) [20]. Due to their direct role in regulating the gene expression of most of the human genome [16,21,22], miRNAs are directly involved in almost all aspects of cellular functions. Specifically, the cellular pathways that underlie cancer progression are regulated by either oncogenic or tumor suppressive miRNAs [12,23]. This suggests that miRNAs are central regulatory elements and represent a promising avenue for therapeutic intervention.
3. The Small Rho GTPases
Ras small GTPases are a superfamily of monomeric hydrolases that are found in all eukaryotic cells and function similarly to the α-subunits of heterotrimeric G proteins. Small GTPases act as molecular switches to facilitate cell activities including proliferation, morphology change, adhesion, migration, invasion and nuclear or vesicular transport, among others. This molecular switching is driven by binding and hydrolyzing GTP, leading to the transition of small GTPases between three conformational states; 1) the GDP-bound, 2) the GTP-bound and 3) the empty state that transiently exists between the replacement of GDP with GTP in the guanine nucleotide binding site [24]. The three main areas of the GTPase that change between GTP- or GDP-bound states are referred to as the phosphate binding loop (P-loop), switch 1 (residues 30–40, also known as the effector loop) and switch 2 (residues 60–76) [25,26], all of which reside within the GTP-binding site of the GTPase. The GDP-bound state is generally considered inactive (“off”), while the GTP-bound form is the active (“on”) form which allows GTPases to move to the cell membrane region and interact with downstream effectors.
The small Rho GTPase family is one of the five originally classified major subfamilies of the Ras small GTPase superfamily [27]. It consists of 20 small (190–250 residues) molecules which control the cytoskeleton and cell morphology specifically by regulating actin dynamics (Table 1). They share ~30% sequence identity with the other Ras superfamily proteins and between 40–95% sequence identity within the subfamily. In addition to containing sequence motifs common to all Ras small GTPases, what structurally separates Rho small GTPases from other proteins in the Ras superfamily is the insertion of 9–12 residues, after residue 122 located between the fifth β-strand and fourth α-helix within the GTPase domain [28,29]. The majority of Rho GTPases undergo C-terminal post-translational modifications by isoprenoid lipids or palmitate fatty acids [30,31], which help localize their subcellular localization and association with membranes or organelles. In addition to modifications to their C-terminal, Rho GTPases are also directly regulated by GTPase-activating proteins (GAPs), guanine nucleotide exchange factors (GEFs), GDP dissociation inhibitors (GDIs) and guanine nucleotide exchange modifiers (GEMs) discussed later in this review. Of the 20 members of the Rho GTPase subfamily (Table 1), the best characterized Rho molecules are RhoA, Rac1, and Cdc42. RhoA promotes actin–myosin contractility and thus controls stress fiber and focal adhesion formation and turnover. Rac1 drives actin polymerization for the formation of membrane ruffling and lamellipodia, or the large projection at the leading edge of the migrating cell. Cdc42 regulates the formation of filopodia, which are actin-rich, finger-like projections that exude from the lamellipodia at the leading edge of the migrating cell.
4. Regulators of the Small Rho GTPases
The “on” and “off” states of the Rho GTPases can be accelerated by the interaction with certain regulators of G-protein signaling (Figure 2) [4,5,6]. GTPase-activating proteins (GAPs) accelerate the Rho GTPases intrinsic phosphatase capability, putting the GTPase into the “off” state. Conversely, guanine nucleotide exchange factors (GEFs) activate Rho GTPases by rapidly exchanging GDP with the GTP. GDP dissociation factors (GDIs) also act to put the Rho GTPases into the “off” state by binding and sequestering Rho GTPases. Since the number of GAPs and GEFs outnumbers the number of Rho GTPases by over 3 to 1, many of these GAPs, GEFs and GDIs target the same Rho GTPase. However, some of these GAPs, GEFs and GDIs have been shown to be specific for a single Rho GTPase over the others. Moreover, these regulators of small G-protein signaling have been shown to be regulated by the small Rho GTPases themselves.
4.1. GTPase-Activating Proteins (GAPs)
The Rho GTPase-activating proteins (RhoGAPs) are one of the major regulators of Rho GTPases found in all eukaryotes. They are defined by the presence of a conserved 150 residue RhoGAP domain, which is distinct from GAPs for other classes of GTPases. This domain consists of nine α helices and a conserved arginine residue in a loop structure [47]. The RhoGAP domain gives RhoGAPs their function because it is sufficient for the binding to GTP-bound Rho proteins as well as accelerating their GTPase activity. Currently, there are over 60 RhoGAPs reported in humans (Table 2), far outnumbering their Rho GTPase substrates. This suggests that each RhoGAP may play a specialized role in regulating the multiple GTPase activity, location, or effector association and mandates the tight control of RhoGAP activity so that Rho GTPases are not always in the “off” state. Traditionally, GAPs were thought of as tumor suppressors. However, as some recent work has demonstrated, the overexpression of RhoGAPs in some cancers [48,49,50] and the interaction between the Rho GTPases and RhoGAPs may be more complex and context-dependent than originally thought. Expanding this to miRNAs, this would also suggest that miRNAs that directly target RhoGAPs also play a context-dependent role in cancer progression.
Insight into how RhoGAPs facilitate the intrinsic GTPase activity of Rho GTPases stems from structural work on Ras GTPase-activating proteins (RasGAPs). This is because even though the GAP domains differ, the tertiary structure and the fundamental GTPase-activating mechanism are similar to that of the RasGAPs [260,261]. Firstly, the RhoGAP interacts with the P-loop, switch 1 and the switch 2 regions of the GTP-binding site. This causes a slight rotation of the GTPase, placing the arginine residue of the RhoGAP directly into the active site of the Rho GTPase [262,263]. This not only begins to stimulate the catalysis of GTP, but also stabilizes any charges developed during the transitional state [264]. The inserted arginine interacts with Gln61 in the switch 2 region of the GTPase. The Gln61 residue is important because it is responsible for the interaction with a water molecule used to stimulate the intrinsic GTPase catalytic activity. This water-mediated nucleophilic attack is performed on the terminal ɣ phosphate of a GTP molecule, producing GDP and an inorganic phosphate. Although it is not well known how RhoGAPs choose their specific Rho GTPase target, it has been suggested that this is achieved by recognizing sequences in the α3 helix of the Rho GTPase [265,266].
4.2. Guanine Nucleotide Exchange Factors (GEFs)
Rho guanine nucleotide exchange factors (RhoGEFs) are directly responsible for the activation of Rho GTPases by catalyzing the exchange of GDP for GTP. Some RhoGEFs display specificity toward a single Rho GTPase, while others exhibit more promiscuity. As in the case of RhoGAPs, the highly controlled regulation of RhoGEF activity is paramount to ensure that Rho GTPases are not always in the “on” state. This regulation and specificity of RhoGEFs is determined by regulatory mechanisms such as posttranslational modifications, having differing sensitivities to lipids, direct binding to surface receptors, and an association with specialized complexes [267,268,269]. Based upon their functions, this would suggest that aberrant RhoGEF regulation, such as increased gene expression or mutations causing constitutive activation, is a main driver behind cancer progression. Indeed, RhoGEFs are positive regulators of cancer progression, where their increased expression drives cancer cell migration, invasion, adhesion and metastasis. Therefore, this suggests that miRNAs that directly target RhoGEFs function as tumor suppressors. However, this idea has been challenged recently where a RhoGEF (ARHGEF10) was shown to function as a tumor suppressor in pancreatic ductal adenocarcinoma [270].
There are two subfamilies of RhoGEFs: the diffuse B-cell lymphoma (Dbl) and dedicator of cytokinesis (DOCK) families. The 73 members of the Dbl family (Table 2) share a ~200 residue catalytic Dbl homology (DH) domain immediately preceding an adjacent regulatory ~100 residue pleckstrin homology (PH) domain [271]. However, some family members possess tandem DH and PH domains or completely lack a PH domain [272]. They also differ significantly in the N- and C-terminal sequences, which is used to regulate the intrinsic RhoGEF catalytic activity, localization, or complex association as described above. Additionally, it should be noted that unlike humans, plants do not possess Dbl RhoGEFs [273]. Functionally, the DH domain of Dbl RhoGEFs is responsible for facilitating the exchange of GDP for GTP by stabilizing switch 1, the remodeling of the alanine near the Mg2+ binding site in switch 2, and stabilizing the P-loop of the Rho GTPase [6]. Dbl RhoGEFs have been shown to act on RhoA, Rac1, and Cdc42. Conversely, the DOCK RhoGEFS (Table 2) are characterized by a conserved Dock-homology region-2 (DHR2) domain that serves as the catalytic domain and the DHR1 domain that locates them to specific membranes. What makes these RhoGEFs unique is that the DHR2 domain stabilizes the switch 1 of the Rho GTPase using interactions not seen in other typical RhoGEFs. DOCK proteins also utilize a valine residue to help dissociate the bound GDP, but does not distort switch 2 of the GTPase like Dbl family members [6]. DOCK proteins are shown to act primarily as RhoGEFs for Rac1 and Cdc42, but not RhoA [274,275].
4.3. GDP Dissociation Inhibitors (GDIs)
In addition to RhoGAPs and RhoGEFs, Rho GTPase GDP dissociation inhibitors (RhoGDIs) perform a unique function in the regulation of Rho GTPases. Characterized by a conserved ~60 residue N-terminal domain, RhoGDIs prevent Rho GTPases from activation and the subsequent interactions with downstream effectors through three different mechanisms: (1) binding and sequestering Rho GTPases in the inactive, GDP-bound form preventing activation by RhoGEFs. (2) binding and sequestering Rho GTPases in the active, GTP-bound form preventing the hydrolysis of GTP by either the intrinsic or the RhoGAP-stimulated GTPase activity, and (3) modulating the cycling of Rho GTPases between the cytosolic and membrane localization [276,277]. These biochemical functions mean that RhoGDIs have dual roles in the cell; they form soluble complexes with GDP-bound Rho GTPases in the cytosol, but also monitor Rho GTPases at the site of action on membranes.
To elicit their effects, RhoGDIs recognize the isoprenoid geranylgeranyl lipids at the C-terminus of the Rho GTPase [276,277]. Once bound to the Rho GTPase, the N-terminal domain of the RhoGDI interacts with the switch 1 and switch 2 of the Rho GTPase, restricting the spatial flexibility needed to exchange GDP or hydrolyze GTP. In contrast to the large number of RhoGAPs and RhoGEFs, only three RhoGDIs have been identified in mammals (Table 2). RhoGDIα (also known as RhoGDI1) is the most commonly found and ubiquitously expressed RhoGDI and it is able to form complexes with most members of the Rho family [250,253,278]. RhoGDIβ (also known as RhoGDI2) is predominantly expressed in hematopoietic cells [279,280], but its dysregulation is also found in certain cancer types [281,282,283,284]. It can interact with several Rho GTPases, but the affinity for complexing is 10–20 fold lower than that of RhoGDIα [256,285]. Lastly, RhoGDIɣ (also known as RhoGDI3) is preferentially expressed in the brain, lung, kidney, testis and pancreas [258,259], and is targeted to the Golgi complex through its N-terminal domain where it predominantly interacts with RhoB and RhoG [257,258]. The dysregulation of the RhoGDIs is linked to cancer cell migration, invasion, and metastasis [286,287], but the downstream effects of the altered RhoGDI expression seems to be context and cancer-type dependent. Additionally, it has been shown that RhoGDI mRNA can interact with regulators critical to the miRNA biogenesis, stability and activity [288], suggesting a more diverse role for RhoGDIs in cancer cells.
4.4. Guanine Nucleotide Exchange Modulators (GEMs)
In addition to GAPs, GEFs, and GDIs, another family of GTPase regulators has recently been identified. The guanine nucleotide exchange modulators (GEMs) are unique because they function as both a GEF and a GDI depending on context [7,289]. The members of this family are characterized by a ~30 residue domain that directly binds to G proteins. GEMs share little sequence homology between family members and act as central regulators to diverse G protein signaling cascades. The prototypical GEM family member is Girdin (for Girder of actin [290], also known as GIV [291], HkRP1 [292], or APE [293]), which is a multi-domain cytosolic protein that was identified to regulate the actin cytoskeleton during cell migration. Although it has been shown to drive actin cytoskeletal remodeling and bind to α-subunits of heterotrimeric G proteins like other GTPase regulators, these studies were not performed using human Rho GTPases. Therefore, it is not yet known if GEMs can directly modulate Rho GTPases activity.
5. MiRNAs Target RhoGAPs, RhoGEFs and RhoGDIs to Regulate Rho GTPase Activities and Cancer Progression
In addition to directly targeting Rho GTPases themselves [8], miRNAs are also directly involved upstream by targeting the modulators of Rho GTPase activity. This section will focus on miRNAs that have been identified to directly bind to the 3′UTR of RhoGAPs, RhoGEFs and RhoGDIs in cancer. The decreased expression of these Rho GTPase modulators has context-dependent effects on processes of cancer progression which we review here. A full list of the miRNAs that have been shown to reduce the expressions of RhoGAP, RhoGEF and RhoGDI mRNA and/or protein expression is found in Table 3.
5.1. MiRNA Targeting of RhoGAPs
Many studies including ours have shown that active Rho GTPases promote cancer cell survival, proliferation, migration and tumor metastasis [426]. Therefore, RhoGAPs, which are negative regulators of Rho GTPases, are usually considered to have tumor-suppressive functions [427]. However, data elucidating the physiologic function of the miRNA targeting of RhoGAPs in cancer currently show the most context-dependent effects of the three groups of Rho GTPase regulators. For example, it was found that the enforced expression of miR-34a in lung cancer cells [296] or miR-509 in primary osteosarcoma cells [297] reduced the growth and migration in vitro, and tumor invasion and metastasis in mice through targeting ARHGAP1. Furthermore, decreased ARHGAP1 expression by miR-509 sensitized cells to cisplatin, a commonly used chemotherapeutic [297]. However, Satterfield et al. found that the reduction of ARHGAP1 by miR-130b in Ewing sarcoma induced the cancer cell growth, migration and invasion in vitro, and promoted lung colonization when injected into the tail vein of Rag2−/− mice [298]. In a different study, MDA-MB-231 breast cancer cells stably expressing miR-940 injected into the calvarial or tibial bones of BALB/cAJcl-nu/nu mice resulted in tumors with enhanced osteoblastic lesions [299]. This study determined that ARHGAP1 was a direct target of miR-940 and the reduction of ARHGAP1 led to an increase in osteoblastic lesions.
The context-dependent effects of the miRNA targeting of ARHGAP5 have also been found. Since, Wang et al. previously found that miR-486-5p was significantly downregulated in non-small cell lung cancer (NSCLC), and they next aimed to identify the effects of miR-486 on NSCLC cancer progression [300]. The ectopic addition of miR-486-5p into A549 and H157 NSCLC cell lines resulted in the significant inhibition of cell growth as well as cell migration and invasion as determined by a Transwell migration assay. A tail vein injection of the cells transfected with miR-486-5p demonstrated that miR-486-5p could reduce colonization and growth in the lungs. Mechanistically, they determined that the oncogenic ARHGAP5 was a direct target of miR-486-5p, and the silencing of ARHGAP5 recapitulated the phenotypes of increased miR-486-5p expression [300]. Conversely, in two separate studies the induction of miR-494 by ionizing radiation or miR-744 by lactic acid enhanced the cell motility and invasion through the targeting of ARHGAP5 [301,302]. Interestingly, instead of decreasing the expression of ARHGAP5, it was shown that miR-744 bound directly to the promoter and increased the expression of ARHGAP5 to drive these processes of cancer progression [302,303]. This suggests a more complex regulatory mechanism of miRNAs with ARHGAP5 in cancer.
The miR-200 family is well known to regulate many aspects of cancer progression [12]. Therefore, it is not surprising that works have identified the effects of the miR-200 family on RhoGAPs. Deleted in liver cancer 1 (DLC1, also known as ARHGAP7) is typically thought of as a tumor suppressor, however, studying the miR-200 family discovered the inconsistent effects of targeting on cancer progression. In colorectal cancer, Wu and colleagues found that miR-141 drove cancer progression [306]. In addition to miR-141 inversely correlating with DLC1, the transient or stable expression of miR-141 in Lovo colorectal cancer cells promoted cell growth, migration and invasion, as well as tumor growth. They found that the miR-141-induced cell growth was accompanied by an increase of cells in the G2/M phase and a decrease in the G0/G1 phase of the cell cycle, suggesting that miR-141 could promote cell cycle progression. Xiao et al. found that the expression of another miR-200 family member, miR-429, similarly increased cell growth by directly targeting DLC1 in non-small cell lung cancer (NSCLC) [308]. In both studies, the expression of DLC1 lacking the 3′UTR in miRNA-expressing cells was able to overcome the oncogenic effects, confirming the oncogenic mechanism of miR-141 and -429 through targeting DLC1. Conversely, miR-200a, -200b and -200c were shown to suppress cancer progression [310]. In direct contrast to the study with miR-429 [308], the re-expression of another miR-200 family members not only resulted in an increase in E-cadherin, characteristic of a non-migratory and invasive epithelial cell, but also reduced the gene expression profile of a metastatic lung cell line in response to the direct targeting of DLC1 in NSCLC [310]. Moreover, Ibrahim et al. showed that the miR-200c-mediated DLC1 mRNA inhibition led to enhanced growth and colony formation, but reduced migration and invasion in serous ovarian cancer [311]. Since members of the miR-200 family share the same targeting seed sequence [12] and conflicting results are even obtained within the same cancer type [308,310], defining contexts in which environmental factors contribute to the miRNA regulating effects of cancer progression warrants further investigation.
Although the targeting of the aforementioned RhoGAPs generated both controversial and context-dependent results, some RhoGAPs maintain a more consistent role in cancer. Examples of this include; (1) the reduction of ARHGAP18 expression by the stable expression of either miR-153 or miR-200b inhibited the cancer cell migration and metastasis in liver and breast cancer cell lines [50,318], (2) the targeting of RACGAP1 by miR-192, -204, or -4324 induced the cell cycle arrest and reduced migration and invasion in osteosarcoma [356], pancreatic [357] and bladder cancer [358], respectively, and (3) the targeting of ARHGAP37 (STARD13) by miR-9 or miR-125b drove cancer cell growth, migration, invasion and metastasis [323,324,325,326]. Together these data suggest that based upon the currently available literature, some RhoGAPs act specifically as oncogenes or tumor suppressors.
IQGAP1 is also targeted by miRNAs, and although it is a scaffolding RhoGAP that does not possess intrinsic GTPase-activating capabilities, it has been shown to function as an oncogene in cancer. The study by Furuta and colleagues found that miR-124 and miR-203 were frequently methylated in hepatocellular carcinoma (HCC), and inversely correlated with patient prognosis [330]. The ectopic expression of miR-124 or miR-203 both not only suppressed hepatocellular carcinoma cell growth, but also induced cell cycle arrest and apoptosis in part by the direct targeting of IQGAP1. The regulation of IQGAP1 expression by miR-124 has also been identified in glioblastoma [331] and in endometrial cancer [332]. In both of these studies, the suppression of IQGAP1 either by direct repression or the ectopic expression of miR-124 blunted cell migration and invasion. These data suggest miR-124 as a tumor suppressor in multiple cancer types. Furthermore, Sun et al. found that miR-506 was inversely correlated with patient prognosis and the tumor stage in breast cancer [333]. In addition to the reduction of cell growth, invasion and adhesion, the expression of miR-506 also reduced MAPK/ERK signaling. This group found that miR-506 suppressed these functions and pathways by targeting IQGAP1, and the expression of IQGAP1 lacking the 3′UTR rescued the effects of miR-506 on breast cancer progression [333]. Collectively, these data suggest that although IQGAP1 lacks the typical GTPase-activating function, the dysregulation of IQGAP1 regulation is a critical step in cancer progression for different cancer types.
5.2. MiRNA Targeting of RhoGEFs
Since RhoGEFs are positive regulators of Rho GTPases, they are generally thought to have oncogenic functions in cancer. Therefore, miRNAs that directly target RhoGEFs function as tumor suppressors. Current literature supported this as the reduction of RhoGEF expression by miRNAs generally blunts cancer progression. One of the most well known Dbl family RhoGEFs is the son of sevenless 1 (SOS1), which not only activates Rho GTPases, but is also known to scaffold and activate Ras [428]. Due to its central function in cancer progression, many studies have found that the downregulation of SOS1 expression blocks many aspects that define cancer. In support of this, studies have found that both miR-20b and miR-4728 expression levels were significantly reduced in papillary thyroid cancer (PTC) compared to adjacent normal thyroid tissue, but that low levels of these miRNAs correlated with worse patient prognosis [383,389]. The ectopic expression of either of these miRNAs inhibited PTC cell growth, migration and invasion. In support of the critical function of SOS1 in Ras-mediated signaling, the expression of miR-20b and miR-4728 significantly blunted MAPK/ERK signaling by directly targeting SOS1. It was also found that the expression of SOS1 was able to overcome miRNA-mediated inhibition on growth, migration, invasion and signaling [383]. SOS1 can also promote bone metastasis through the direct modulation of breast cancer stem cell (CSC) migration and invasion [388]. Lin et al. found that miR-628 was significantly downregulated in bone metastatic breast cancer cells compared to the cells of the primary tumor. As CSCs are the major driving force behind metastasis, they next determined the effects of miR-628 expression on the functions of CSCs. The ectopic expression of a miR-628 mimic in CD44+/CD24− breast CSCs significantly reduced cell migration and invasion as assayed by a Transwell assay. Additionally, the miR-628 mimic also reduced the expression of mesenchymal markers, vimentin and Snail, and increased the E-cadherin epithelial marker, suggesting a non-migratory phenotype. This study determined that miR-628 attenuates CSC migration and invasion through the direct targeting of the SOS1 3′UTR, and that the SOS1 expression was able to overcome the inhibitory effects of miR-628 on migration and invasion [388]. These data collectively demonstrate that SOS1 is an oncogene and miRNAs that directly target SOS1 act as tumor suppressors.
Another the well studied RhoGEFs is T-lymphoma invasion and metastasis-inducing protein 1, or TIAM1. Although originally identified as a Rac-specific GEF, studies have also identified GEF effects on RhoA and CDC42, albeit to a less efficient extent [226], where TIAM1 operates as a potent oncogene. Two members of the miR-10 family (miR-10a and miR-10b) have been shown to function as tumor suppressors by directly targeting TIAM1 [391,392,393,394]. The expression of these two miRNAs were found to be significantly reduced in tumor tissue compared to the matched normal tissue [391,393,394]. The stable expression of these two miRNAs inhibited Rac activation, cell growth, migration and invasion, as well as induced apoptosis by regulating TIAM1 expression. Furthermore, these results translated to mice as the expressions of miR-10a and miR-10b were found to reduce tumor growth and metastasis in esophageal squamous cell carcinoma [391] and cervical cancer [393], respectively. It was found that miR-10b was epigenetically silenced by DNA methylation [393,394], suggesting that demethylating agents could reactivate tumor suppressors to combat cancer progression. It should also be noted that some studies have identified an oncogenic role for miRNAs targeting TIAM1 [404,405], thus more work is needed to uncover the context-dependent effects of TIAM1 on cancer progression.
Outside of the hematopoietic system, VAV2 is critical for regulating tumorigenesis and cancer progression. As with the other central Dbl-like RhoGEFs described above, miRNAs target and regulate the expression of VAV2, leading to the suppression of cancer cell growth, migration and invasion. The study by Bischoff and colleagues found that miR-149 functioned as a key metastatic suppressor through VAV2-Rac downregulation in basal-like breast cancer [407]. Compared to luminal and HER2-positive breast cancer, the basal-like cells displayed significantly lower levels of miR-149. This group found that although the stable expression of miR-149 did not affect cell growth, it had detrimental effects on cell invasion in a Transwell assay towards multiple stimuli, including collagen and a combination of serum and EGF. A further analysis of single cells found that the distance and velocity were unchanged, but that the net displacement was reduced by miR-149, suggesting a defect in the directional migration of these cells. Mechanistically, the downregulation of VAV2 by miR-149 resulted in the impaired activation of Src and paxillin, key signaling molecules downstream of integrin engagement, drastically reducing cell spreading and the adhesion needed for migration [407]. In addition to regulating migration and invasion [406,407], the reduction of VAV2 has been linked to the inhibition of epithelial-mesenchymal transition (EMT), a process where a normally polar, epithelial cell undergoes a change to a mesenchymal-like cell to take on the characteristics of a mesenchymal cell and become more motile and invasive, as well as angiogenesis [408,409].
Similar to the Dbl family, current work suggested that DOCK family RhoGEFs typically act as oncogenic drivers of cancer. Many studies on the DOCK family have centered around their role in EMT. For example, it was shown that DOCK1 was able to promote an EMT phenotype in glioma and breast cancer [412,413]. This was shown to be accomplished through a DOCK1-mediated interleukin-8- or -22-driven NFĸB/Snail signaling mechanism. The expression of miR-31 or -486 was able to blunt this signaling pathway, promoting a non-migratory epithelial-like phenotype in cells and inhibited metastasis, through the downregulation of DOCK1 [412,413]. Similarly, the cooperation between Twist1 and BMI1 to suppress let-7i expression promoted the acquisition of properties of the EMT, namely cell migration and stem-like properties in head and neck squamous cell carcinoma (HNSCC) [414]. The loss of let-7i leads to an increase in DOCK3 expression, which promotes RAC1 activation and EMT properties. Interestingly, the re-expression of let-7i did not change the expression of EMT markers in 2D or 3D cultures, but did blunt the mesenchymal movement, invasion and metastasis [414], suggesting that let-7i can target characteristics of the EMT without directly affecting the expression of typical EMT markers.
5.3. MiRNA Targeting of RhoGDIs
The expression of RhoGDIs is dysregulated in many cancers and they have been shown to mediate processes directly linked to tumorigenesis and cancer progression. Much of the current work has focused on the most prominent member of the RhoGDI family, RhoGDIα. In patients with glioma, Lin et al. found that the expression of RhoGDIα protein, but not the mRNA, was frequently downregulated in the high grade glioma compared to the lower grade and matched normal brain tissue, suggesting miRNA regulation [288]. Indeed, miR-151, as well as miR-16, were found to directly target RhoGDIα mRNA to positively regulate cell migration and invasion. Interestingly, this group found that the binding of PCBP2 (multifunctional nucleic acid binding protein 2) facilitated miR-151 and -16 binding to the 3′UTR of RhoGDIα by changing the mRNA secondary structure [288], suggesting a more complex mechanism for the RhoGDIα regulation. Similar results were found in hepatocellular carcinoma [421], prostate [422] and ovarian [423] cancer, where miR-151 promoted tumor progression through processes involving synergistic cooperation with focal adhesion kinase (FAK) [421] and the reduction of Akt/mTOR signaling [423] by the direct regulation of RhoGDIα. Like the other RhoGTPase regulators, the effects of the suppression of RhoGDIα on cancer progression seem to be context dependent as the miRNA suppression of RhoGDIα in colorectal cancer resulted in suppressed cancer cell migration, invasion and metastasis [418,419].
6. Preclinically and Clinically Targeting miRNAs and RhoGAPs, RhoGEFs, and RhoGDIs
One question that remains to be answered is how to successfully resolve the context-dependent pro- and anti-metastatic functions of individual Rho GTPases in order to develop proper therapies. While the majority of RhoA, Rac1 and Cdc42 in vitro studies supported a pro-metastatic function, it was within in vivo studies that opposing functions were found. This may be explained by the differences in the model systems used for these experiments. Firstly, cell lines rely on key signaling pathways, including the Rho GTPases, to facilitate their adaption to rapid growth on plastic. Therefore, cell lines could be hypersensitive to any change in Rho GTPase signaling pathways not seen in in vivo systems. Additionally, it has been shown that the RhoGEF P-Rex1 was positively correlated with estrogen receptor expression and inversely correlated with PI3K levels in breast cancer [429], suggesting that the hormone receptor status and PI3K status may relate to, and be important for interpreting, the contradicting results in vitro. Secondly, Rho GTPases may also be involved in positive or negative feedback loops with the tumor microenvironment that regulate the metastatic potential of a tumor, which is not completely mimicked in the cell culture systems. Lastly, manipulating the expression levels of single Rho GTPase in cell lines may disrupt the level or activity of other Rho GTPases within the cell, affect homeostasis and lead to a poor drug efficacy.
Rho GTPases themselves are not considered viable clinical targets. This is due to their (1) structure, which provides limited small-molecule binding pockets, (2) high affinity for GTP and GDP, and (3) high concentration of GTP available in the cells [430,431]. Therefore, a more reasonable approach to limit Rho GTPases in cancer is to target the activators of Rho GTPases. Since the enhanced activation or overexpression of Rho GTPases is common in cancer [432,433], these treatment strategies primarily revolve around blocking the interaction of Rho GTPases with GEFs or GTP nucleotide binding. By targeting GEFs or GTP binding, Rho GTPases lose the ability to quickly exchange GDP for GTP leading to the inhibition of the interaction with downstream effectors, which thus inhibits signaling cascades. Both targeting strategies have been shown to be effective for cancer and have been reviewed more extensively, along with other Rho GTPase targeting strategies, which can be found elsewhere [430,431,434,435].
However, a more straightforward approach to regulating Rho GTPases is to manipulate miRNA expression. As discussed above, a growing body of evidence has demonstrated that miRNAs are effective at regulating Rho GTPase activity in cancer by targeting RhoGAPs, RhoGEFs and RhoGDIs. This provides a rationale for manipulating the miRNAs as a treatment strategy. The main advantage of using miRNAs as therapeutics is that a single miRNA can affect multiple pathways, avoiding the “one-drug, one-target” approach that often leads to resistance [436]. The use of the so-called miRNA mimics is the main and most effective approach for restoring or replacing tumor-suppressing miRNA expression. These are often chemically modified RNA duplexes that can be loaded into the RISC and act similarly to a mature miRNA on downstream targets. Many studies have shown the efficacy of this approach to regulate miRNA targets such as the Rho GTPases and Rho GTPase regulators [361,437,438,439]. These studies have highlighted the potential utility of miRNAs in the clinic, and provided supporting evidence for ongoing miRNA mimics in clinical trials (miR-16 (Mesomir 1), miR-34 (MRX34), and miR-29b (MRG-201)). However, miRNA mimics have hit many roadblocks that currently limit its use in clinics.
On the other hand, the development of miRNA inhibitors to target oncogenic miRNAs is a burgeoning area of research for clinical use. This includes the development of small molecule inhibitors of miRNAs (SMIRs), locked nucleic acid (LNA) antimiRs, antagomiRs and miRNA sponges. Gumireddy and colleagues reported the first SMIR in 2008, when they found a small molecule that effectively inhibited the transcription of miR-21 [440]. Since then, other SMIRs have been identified [441,442,443], however none of these SMIRs have yet been shown to result in an altered expression of the Rho GTPase regulators. LNAs are bicyclic RNA analogs that contains a 2′-O, 4′-C methylene bridge that restricts the flexibility of the ring [444]. LNAs possess an extremely high affinity and specificity for complementary DNA and RNA sequences, making them effective for antagonizing miRNA function [445]. LNAs have been successfully employed resulting in increased levels of RhoGAPs in culture [301]. AntagomiRs are single-stranded RNA molecules designed to be complementary to the target miRNA [446] and have been shown to effectively regulate the activity of Rho GTPases in vivo [438,447]. The last method for miRNA inhibition involved miRNA sponges, or competitive inhibitors that contained multiple, tandem seed sequence binding sites to a miRNA of interest and could inhibit all miRNAs that shared the seed sequence that it expressed [448]. MiRNA sponges have been used to enforce the expression of Rho GTPase regulators in the brain and liver [449,450]. Each of these strategies effectively sequestered the target miRNA and prevented it from being loaded into the RISC. This resulted in the enhanced expression of the miRNA target. However, as with the miRNA mimics, inhibitors of miRNAs are still early in terms of clinical trial effectiveness, but show promise.
Although the targeting of Rho GTPase regulators for cancer with miRNA replacement or miRNA inhibitors is a straightforward idea, the two biggest barriers that miRNA interventions face are the stability issue and the development of effective delivery systems. RNA molecules are inherently unstable due to their 2′-OH chemical group [451,452]. However, recent advances in our understanding of non-coding RNA (ncRNA) biology has led to the production of natural or chemical modifications that increase RNA stability, seen in many of the miRNA inhibitors discussed above. In contrast, the effective delivery of miRNA remains the biggest challenge for miRNA therapies. Delivery systems are currently either viral- or non-viral-based. The toxicity and immunogenicity related to viral vectors limit their use in the clinic, therefore non-viral-based delivery systems have continued to evolve as a promising approach. In addition to the lack of toxicity and immunogenicity, the tolerance of the cargo size and the ease of control over the composition, modification and manufacturing add to the attraction of non-viral-based delivery systems for ncRNA therapies [453,454,455]. In particular, cationic materials that condense negatively charged nucleic acids through electrostatic interactions have shown the efficiency and specificity needed for clinical miRNA therapies. Recent work has tried to modify particle size and surface composition to increase the lower efficiency of delivery when compared to viral vectors and avoid unexpected biological outcomes [453,456,457,458].
7. Conclusions
This review clearly demonstrated that miRNAs are involved in regulating multiple facets of cancer cell biology through the regulation of RhoGAPs, RhoGEFs and RhoGDIs. A comprehensive list of the currently validated RhoGAP, RhoGEF, and RhoGDI miRNA targets is shown in Table 3. Although the targeting of RhoGAPs results in much more context-dependent effects on cancer progression, the current data generally suggest that miRNAs that target RhoGEFs and RhoGDIs act as tumor suppressors and oncogenes, respectively (Figure 3).
The data summarized here demonstrate a strong rationale for targeting Rho GTPase regulators as a potential therapeutic approach for aggressive and highly metastatic cancers. However, no clinically effective drugs targeting these regulators have been approved for cancer therapy. Current studies revolve around inhibiting activators of Rho GTPases (RhoGEFs), however compounds that mimic RhoGAPs by inserting an arginine residue or stimulate GTP hydrolysis may provide another promising avenue for treatment.
MiRNAs are proving to be advantageous in the clinic, particularly in diseases such as cancer, that may not have a single underlying cause. Moreover, some small molecule drugs are currently being developed to target multiple pathways [459], lending credence to the use of miRNAs clinically. In order to better utilize miRNAs that therapeutically target the Rho GTPase regulators, future work will need to better identify the context-dependent effects and mechanisms of the miRNA targeting of RhoGAPs, RhoGEFs and RhoGDIs. Moreover, identifying what leads to the context-dependent tumor suppressor or oncogenic effects of a single miRNA in different cancers will be critical not only to our basic knowledge of cancer progression, but also to enhance the clinical utility for that miRNA (for example, miR-141 targets ARHGAP7 (DLC1) to promote colorectal cancer cell growth, migration and invasion [306], but inhibits liver cancer progression by targeting TIAM1 [400]). In terms of the regulators themselves, future work will need to address whether GEMs directly modulate Rho GTPase activity and signaling as well as the specificity of GEMs for certain members of the Rho GTPase family. Accomplishing both will not only advance our understanding of the functions of these regulators in cancer, but it will also identify other potential miRNA and Rho GTPase regulator targets, increasing their appeal as therapeutics. Additionally, although significant strides have been taken to increase the stability and delivery systems for miRNAs, more work needs to be done to improve the targeting ability of these systems and to enhance therapeutic effects without triggering an immune system response. Even though the use of miRNAs in a clinical setting is currently limited, the recent FDA approval of siRNA for the treatment of the peripheral nerve disease caused by hereditary transthyretin-mediated amyloidosis is promising for ncRNA therapy [460]. Overall, the wide range of interactions between miRNAs and RhoGAPs, RhoGEFs and RhoGDIs in various cancers provides new challenges and opportunities for the development of new general and personalized therapeutic strategies.
Funding
This work was supported by a Research Scholar Grant (RGS-15-026-01-CSM) from the American Cancer Society to C.Y. Brock Humphries, PhD, was supported by an American Cancer Society—Michigan Cancer Research Fund Postdoctoral Fellowship, PF-18-236-01-CCG.
Conflicts of Interest
The authors declare that they have no conflict of interests.
Abbreviations
| 3′ untranslated region | 3′UTR |
| cancer stem cell | CSC |
| diffuse B-cell lymphoma | Dbl |
| Dbl homology | DH |
| DOCK-homology region-1 | DHR1 |
| DOCK-homology region-2 | DHR2 |
| dedicator of cytokinesis | DOCK |
| extracellular matrix | ECM |
| epithelial-to-mesenchymal transition | EMT |
| GTPase-activating protein | GAP |
| guanine nucleotide exchange factor | GEF |
| guanine nucleotide exchange modulator | GEM |
| GDP dissociation inhibitor | GDI |
| guanosine diphosphate | GDP |
| guanosine triphosphate | GTP |
| locked nucleic acid | LNA |
| messenger RNA | mRNA |
| microRNA | miRNA/miR |
| non-coding RNA | ncRNA |
| pleckstrin homology | PH |
| phosphate binding loop | P-loop |
| RNA-induced silencing complex | RISC |
| small molecule inhibitor of miRNA | SMIR |
References
- Ridley, A.J. Rho GTPases and cell migration. J. Cell Sci. 2001, 114, 2713–2722. [Google Scholar]
- Lawson, C.D.; Ridley, A.J. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 2018, 217, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.A.; Govek, E.E.; Böttner, B.; Van Aelst, L. Rho GTPases: Signaling, migration, and invasion. Exp. Cell Res. 2000, 261, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kaibuchi, K.; Kuroda, S.; Amano, M. Regulation of the Cytoskeleton and Cell Adhesion by the Rho Family GTPases in Mammalian Cells. Annu. Rev. Biochem. 1999, 68, 459–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-Binding Proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef]
- Cherfils, J.; Zeghouf, M. Regulation of small GTPases by GEFs, GAPs, and GDIs. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.; Rangamani, P.; Kufareva, I. The GAPs, GEFs, GDIs and…now, GEMs: New kids on the heterotrimeric G protein signaling block. Cell Cycle 2017, 16, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Bi, F.; Zhou, X.; Zheng, Y. Rho GTPase regulation by miRNAs and covalent modifications. Trends Cell Biol. 2012, 22, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of microRNA-target recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphries, B.; Yang, C. The microRNA-200 family: Small molecules with novel roles in cancer development, progression, and therapy. Oncotarget 2015, 6, 6472–6498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphries, B.; Wang, Z.; Yang, C. MicroRNA Regulation of Epigenetic Modifiers in Breast Cancer. Cancers 2019, 11, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Humphries, B.; Yang, C.; Wang, Z. MiR-205 Dysregulations in Breast Cancer: The Complexity and Opportunities. Noncoding RNA 2019, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Lagos-Quintana, M.; Rauhut, R.; Meyer, J.; Borkhardt, A.; Tuschl, T. New microRNAs from mouse and human. RNA 2003, 9, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Pillai, R.S.; Bhattacharyya, S.N.; Filipowicz, W. Repression of protein synthesis by miRNAs: How many mechanisms? Trends Cell Biol. 2007, 17, 118–126. [Google Scholar] [CrossRef]
- Karginov, F.V.; Cheloufi, S.; Chong, M.M.; Stark, A.; Smith, A.D.; Hannon, G.J. Diverse endonucleolytic cleavage sites in the mammalian transcriptome depend upon microRNAs, Drosha, and additional nucleases. Mol. Cell 2010, 38, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Bracken, C.P.; Szubert, J.M.; Mercer, T.R.; Dinger, M.E.; Thomson, D.W.; Mattick, J.S.; Michael, M.Z.; Goodall, G.J. Global analysis of the mammalian RNA degradome reveals widespread miRNA-dependent and miRNA-independent endonucleolytic cleavage. Nucleic Acids Res. 2011, 39, 5658–5668. [Google Scholar] [CrossRef] [Green Version]
- Kartha, R.V.; Subramanian, S. Competing endogenous RNAs (ceRNAs): New entrants to the intricacies of gene regulation. Front. Genet. 2014, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. miRBase: Tools for microRNA genomics. Nucleic Acids Res. 2008, 36, D154–D158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase superfamily: A conserved switch for diverse cell functions. Nature 1990, 348, 125–132. [Google Scholar] [CrossRef]
- Pai, E.F.; Kabsch, W.; Krengel, U.; Holmes, K.C.; John, J.; Wittinghofer, A. Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21 in the triphosphate conformation. Nature 1989, 341, 209–214. [Google Scholar] [CrossRef]
- Milburn, M.V.; Tong, L.; DeVos, A.M.; Brünger, A.; Yamaizumi, Z.; Nishimura, S.; Kim, S.H. Molecular Switch for Signal Transduction: Structural Differences Between Active and Inactive Forms of Protooncogenic ras Proteins. Science 1990, 247, 939–945. [Google Scholar] [CrossRef] [Green Version]
- Kahn, R.A.; Der, C.J.; Bokoch, G.M. The ras superfamily of GTP-binding proteins: Guidelines on nomenclature. FASEB J. 1992, 6, 2512–2513. [Google Scholar] [CrossRef] [Green Version]
- Wennerberg, K.; Der, C.J. Rho-family GTPases: It’s not only Rac and Rho (and I like it). J. Cell Sci. 2004, 117, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Valencia, A.; Chardin, P.; Wittinghofer, A.; Sander, C. The ras Protein Family: Evolutionary Tree and Role of Conserved Amino Acids. Biochemistry 1991, 30, 4637–4648. [Google Scholar] [CrossRef]
- Roberts, P.J.; Mitin, N.; Keller, P.J.; Chenette, E.J.; Madigan, J.P.; Currin, R.O.; Cox, A.D.; Wilson, O.; Kirschmeier, P.; Der, C.J. Rho Family GTPase modification and dependence on CAAX motif-signaled posttranslational modification. J. Biol. Chem. 2008, 283, 25150–25163. [Google Scholar] [CrossRef] [Green Version]
- Adamson, P.; Marshall, C.J.; Hall, A.; Tilbrook, P.A. Post-translational Modifications of p21rho Proteins. J. Biol. Chem. 1992, 267, 20033–20038. [Google Scholar] [PubMed]
- Johnson, D.I. Cdc42: An Essential Rho-Type GTPase Controlling Eukaryotic Cell Polarity. Microbiol. Mol. Biol. Rev. 1999, 63, 54–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etienne-Manneville, S. Cdc42-the centre of polarity. J. Cell Sci. 2004, 117, 1291–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosco, E.E.; Mulloy, J.C.; Zheng, Y. Rac1 GTPase: A “Rac” of all trades. Cell. Mol. Life Sci. 2009, 66, 370–374. [Google Scholar] [CrossRef]
- Gu, Y.; Jia, B.; Yang, F.C.; D’Souza, M.; Harris, C.E.; Derrow, C.W.; Zheng, Y.; Williams, D.A. Biochemical and biological characterization of a human Rac2 GTPase mutant associated with phagocytic immunodeficiency. J. Biol. Chem. 2001, 276, 15929–15938. [Google Scholar] [CrossRef] [Green Version]
- Troeger, A.; Williams, D.A. Hematopoietic-specific Rho GTPases Rac2 and RhoH and human blood disorders. Exp. Cell Res. 2013, 319, 2375–2383. [Google Scholar] [CrossRef] [Green Version]
- De Curtis, I. The Rac3 GTPase in Neuronal Development, Neurodevelopmental Disorders, and Cancer. Cells 2019, 8, 1063. [Google Scholar] [CrossRef] [Green Version]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. Bioessays 2007, 29, 356–370. [Google Scholar] [CrossRef] [Green Version]
- Hanna, S.; El-Sibai, M. Signaling networks of Rho GTPases in cell motility. Cell. Signal. 2013, 25, 1955–1961. [Google Scholar] [CrossRef]
- Sit, S.T.; Manser, E. Rho GTPases and their role in organizing the actin cytoskeleton. J. Cell Sci. 2011, 124, 679–683. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.M.; Ridley, A.J. The RhoB small GTPase in physiology and disease. Small GTPases 2018, 9, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, W.; Rivero, F. Atypical Rho GTPases of the RhoBTB Subfamily: Roles in Vesicle Trafficking and Tumorigenesis. Cells 2016, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthold, J.; Schenkova, K.; Rivero, F. Rho GTPases of the RhoBTB subfamily and tumorigenesis. Acta Pharmacol. Sin. 2008, 29, 285–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.; Pranatharthi, A.; Ross, C.; Srivastava, S. RhoC: A fascinating journey from a cytoskeletal organizer to a Cancer stem cell therapeutic target. J. Exp. Clin. Cancer Res. 2019, 38, 328. [Google Scholar] [CrossRef] [PubMed]
- Phuyal, S.; Farhan, H. Multifaceted Rho GTPase Signaling at the Endomembranes. Front. Cell Dev. Biol. 2019, 7, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, F.M.; Ridley, A.J. Rho GTPases in cancer cell biology. FEBS Lett. 2008, 582, 2093–2101. [Google Scholar] [CrossRef] [Green Version]
- Gamblin, S.J.; Smerdon, S.J. GTPase-activating proteins and their complexes. Curr. Opin. Struct. Biol. 1998, 8, 195–201. [Google Scholar] [CrossRef]
- Lazarini, M.; Traina, F.; Machado-Neto, J.A.; Barcellos, K.S.; Moreira, Y.B.; Brandao, M.M.; Verjovski-Almeida, S.; Ridley, A.J.; Saad, S.T. ARHGAP21 is a RhoGAP for RhoA and RhoC with a role in proliferation and migration of prostate adenocarcinoma cells. Biochim. Biophys. Acta 2013, 1832, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, C.N.; Castellvi-Bel, S.; Chang, L.M.; Bessa, X.; Nakagawa, H.; Harada, H.; Sung, R.K.; Pique, J.M.; Castells, A.; Rustgi, A.K. ARHGAP8 is a novel member of the RHOGAP family related to ARHGAP1/CDC42GAP/p50RHOGAP: Mutation and expression analyses in colorectal and breast cancers. Gene 2004, 336, 59–71. [Google Scholar] [CrossRef]
- Humphries, B.; Wang, Z.; Li, Y.; Jhan, J.R.; Jiang, Y.; Yang, C. ARHGAP18 Downregulation by miR-200b Suppresses Metastasis of Triple-Negative Breast Cancer by Enhancing Activation of RhoA. Cancer Res. 2017, 77, 4051–4064. [Google Scholar] [CrossRef] [Green Version]
- Amin, E.; Jaiswal, M.; Derewenda, U.; Reis, K.; Nouri, K.; Koessmeier, K.T.; Aspenstrom, P.; Somlyo, A.V.; Dvorsky, R.; Ahmadian, M.R. Deciphering the Molecular and Functional Basis of RHOGAP Family Proteins: A systematic approach toward selective inactivation of rho family proteins. J. Biol. Chem. 2016, 291, 20353–20371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, T.H.; Xu, X.; Kaartinen, V.; Heisterkamp, N.; Groffen, J.; Bokoch, G.M. Abr and Bcr are multifunctional regulators of the Rho GTP-binding protein family. Proc. Natl. Acad. Sci. USA 1995, 92, 10282–10286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, K.; Jacques, K.M.; Stauffer, S.; Kubosaki, A.; Zhu, K.; Hirsch, D.S.; Resau, J.; Zheng, Y.; Randazzo, P.A. ARAP1: A Point of Convergence for Arf and Rho Signaling. Mol. Cell 2002, 9, 109–119. [Google Scholar] [CrossRef]
- Yoon, H.Y.; Miura, K.; Cuthbert, E.J.; Davis, K.K.; Ahvazi, B.; Casanova, J.E.; Randazzo, P.A. ARAP2 effects on the actin cytoskeleton are dependent on Arf6-specific GTPase-activating-protein activity and binding to RhoA-GTP. J. Cell Sci. 2006, 119, 4650–4666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krugmann, S.; Williams, R.; Stephens, L.; Hawkins, P.T. ARAP3 is a PI3K- and rap-regulated GAP for RhoA. Curr. Biol. 2004, 14, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Krugmann, S.; Anderson, K.E.; Ridley, S.H.; Risso, N.; McGregor, A.; Coadwell, J.; Davidson, K.; Eguinoa, A.; Ellson, C.D.; Lipp, P.; et al. Identification of ARAP3, a Novel PI3K Effector Regulating Both Arf and Rho GTPases, by Selective Capture on Phosphoinositide Affinity Matrices. Mol. Cell 2002, 9, 95–108. [Google Scholar] [CrossRef]
- Kozma, R.; Ahmed, S.; Best, A.; Lim, L. The GTPase-Activating Protein n-Chimaerin Cooperates with Rac1 and Cdc42Hs To Induce the Formation of Lamellipodia and Filopodia. Mol. Cell. Biol. 1996, 16, 5069–5080. [Google Scholar] [CrossRef] [Green Version]
- Foletta, V.C.; Brown, F.D.; Young, W.S.R. Cloning of rat ARHGAP4/C1, a RhoGAP family member expressed in the nervous system that colocalizes with the Golgi complex and microtubules. Mol. Brain Res. 2002, 107, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Christerson, L.B.; Gallagher, E.; Vanderbilt, C.A.; Whitehurst, A.W.; Wells, C.; Kazempour, R.; Sternweis, P.C.; Cobb, M.H. p115 Rho GTPase activating protein interacts with MEKK1. J. Cell. Physiol. 2002, 192, 200–208. [Google Scholar] [CrossRef]
- Tribioli, C.; Droetto, S.; Bione, S.; Cesareni, G.; Torrisi, M.R.; Lotti, L.V.; Lanfrancone, L.; Toniolo, D.; Pelicci, P. An X chromosome-linked gene encoding a protein with characteristics of a rhoGAP predominantly expressed in hematopoietic cells. Proc. Natl. Acad. Sci. USA 1996, 93, 695–699. [Google Scholar] [CrossRef] [Green Version]
- Burbelo, P.D.; Miyamoto, S.; Utani, A.; Brill, S.; Yamada, K.M.; Hall, A.; Yamada, Y. p190-B, a New Member of the Rho GAP Family, and Rho Are Induced to Cluster after Integrin Cross-linking. J. Biol. Chem. 1995, 270, 30919–30926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, S.K.; Paylor, R.; Jenna, S.; Lamarche-Vane, N.; Armstrong, D.L.; Xu, B.; Mancini, M.A.; Zoghbi, H.Y. Functional analysis of ARHGAP6, a novel GTPaseactivating protein for RhoA. Hum. Mol. Genet. 2000, 9, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Yin, Y. Inhibitory effects of Arhgap6 on cervical carcinoma cells. Tumour Biol. 2016, 37, 1411–1425. [Google Scholar] [CrossRef]
- Wong, C.M.; Lee, J.M.; Ching, Y.P.; Jin, D.Y.; Ng, I.O. Genetic and Epigenetic Alterations of DLC-1 Gene in Hepatocellular Carcinoma. Cancer Res. 2003, 63, 7646–7651. [Google Scholar] [PubMed]
- Shang, X.; Zhou, Y.T.; Low, B.C. Concerted Regulation of Cell Dynamics by BNIP-2 and Cdc42GAP Homology/Sec14p-like, Proline-rich, and GTPase-activating Protein Domains of a Novel Rho GTPase-activating Protein, BPGAP1. J. Biol. Chem. 2003, 278, 45903–45914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, Y.; Kawasoe, T.; Daigo, Y.; Nishiwaki, T.; Ishiguro, H.; Takahashi, M.; Kitayama, J.; Nakamura, Y. Isolation of a novel human gene, ARHGAP9, encoding a rho-GTPase activating protein. Biochem. Biophys. Res. Commun. 2001, 284, 643–649. [Google Scholar] [CrossRef]
- Ren, X.R.; Du, Q.S.; Huang, Y.Z.; Ao, S.Z.; Mei, L.; Xiong, W.C. Regulation of CDC42 GTPase by Proline-rich Tyrosine Kinase 2 Interacting with PSGAP, a Novel Pleckstrin Homology and Src Homology 3 Domain Containing rhoGAP Protein. J. Cell Biol. 2001, 152, 971–984. [Google Scholar] [CrossRef] [Green Version]
- Zanin, E.; Desai, A.; Poser, I.; Toyoda, Y.; Andree, C.; Moebius, C.; Bickle, M.; Conradt, B.; Piekny, A.; Oegema, K. A Conserved RhoGAP Limits M Phase Contractility and Coordinates with Microtubule Asters to Confine RhoA during Cytokinesis. Dev. Cell 2013, 26, 496–510. [Google Scholar] [CrossRef] [Green Version]
- Florio, M.; Albert, M.; Taverna, E.; Namba, T.; Brandl, H.; Lewitus, E.; Haffner, C.; Sykes, A.; Wong, F.K.; Peters, J.; et al. Human-specific gene ARHGAP11B promotes basal progenitor amplification and neocortex expansion. Science 2015, 347, 1465–1470. [Google Scholar] [CrossRef]
- Gentile, A.; D’Alessandro, L.; Lazzari, L.; Martinoglio, B.; Bertotti, A.; Mira, A.; Lanzetti, L.; Comoglio, P.M.; Medico, E. Met-driven invasive growth involves transcriptional regulation of Arhgap12. Oncogene 2008, 27, 5590–5598. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.; Ren, X.R.; Huang, Y.Z.; Xie, Y.; Liu, G.; Saito, H.; Tang, H.; Wen, L.; Brady-Kalnay, S.M.; Mei, L.; et al. Signal Transduction in Neuronal Migration: Roles of GTPase Activating Proteins and the Small GTPase Cdc42 in the Slit-Robo Pathway. Cell 2001, 107, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Endris, V.; Wogatzky, B.; Leimer, U.; Bartsch, D.; Zatyka, M.; Latif, F.; Maher, E.R.; Tariverdian, G.; Kirsch, S.; Karch, D.; et al. The novel Rho-GTPase activating gene MEGAP/ srGAP3 has a putative role in severe mental retardation. Proc. Natl. Acad. Sci. USA 2002, 99, 11754–11759. [Google Scholar] [CrossRef] [Green Version]
- Waltereit, R.; Leimer, U.; von Bohlen Und Halbach, O.; Panke, J.; Holter, S.M.; Garrett, L.; Wittig, K.; Schneider, M.; Schmitt, C.; Calzada-Wack, J.; et al. Srgap3−/− mice present a neurodevelopmental disorder with schizophrenia-related intermediate phenotypes. FASEB J. 2012, 26, 4418–4428. [Google Scholar] [CrossRef] [PubMed]
- Harada, A.; Furuta, B.; Takeuchi, K.; Itakura, M.; Takahashi, M.; Umeda, M. Nadrin, a novel neuron-specific GTPase-activating protein involved in regulated exocytosis. J. Biol. Chem. 2000, 275, 36885–36891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, M.; Hasegawa, H.; Hyodo, T.; Ito, S.; Asano, E.; Yuang, H.; Funasaka, K.; Shimokata, K.; Hasegawa, Y.; Hamaguchi, M.; et al. ARHGAP18, a GTPase-activating protein for RhoA, controls cell shape, spreading, and motility. Mol. Biol. Cell 2011, 22, 3840–3852. [Google Scholar] [CrossRef]
- Chang, G.H.; Lay, A.J.; Ting, K.K.; Zhao, Y.; Coleman, P.R.; Powter, E.E.; Formaz-Preston, A.; Jolly, C.J.; Bower, N.I.; Hogan, B.M.; et al. ARHGAP18: An endogenous inhibitor of angiogenesis, limiting tip formation and stabilizing junctions. Small GTPases 2014, 5, e975002. [Google Scholar] [CrossRef] [Green Version]
- David, M.D.; Petit, D.; Bertoglio, J. The RhoGAP ARHGAP19 controls cytokinesis and chromosome segregation in T lymphocytes. J. Cell Sci. 2014, 127, 400–410. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Sakisaka, T.; Hisata, S.; Baba, T.; Takai, Y. RA-RhoGAP, Rap-activated Rho GTPase-activating protein implicated in neurite outgrowth through Rho. J. Biol. Chem. 2005, 280, 33026–33034. [Google Scholar] [CrossRef] [Green Version]
- Barcellos, K.S.; Bigarella, C.L.; Wagner, M.V.; Vieira, K.P.; Lazarini, M.; Langford, P.R.; Machado-Neto, J.A.; Call, S.G.; Staley, D.M.; Chung, J.Y.; et al. ARHGAP21 protein, a new partner of alpha-tubulin involved in cell-cell adhesion formation and essential for epithelial-mesenchymal transition. J. Biol. Chem. 2013, 288, 2179–2189. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Moreno, V.; Gadea, G.; Ahn, J.; Paterson, H.; Marra, P.; Pinner, S.; Sahai, E.; Marshall, C.J. Rac activation and inactivation control plasticity of tumor cell movement. Cell 2008, 135, 510–523. [Google Scholar] [CrossRef] [Green Version]
- Lavelin, I.; Geiger, B. Characterization of a novel GTPase-activating protein associated with focal adhesions and the actin cytoskeleton. J. Biol. Chem. 2005, 280, 7178–7185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, Y.; Hartwig, J.H.; Stossel, T.P. FilGAP, a Rho- and ROCK-regulated GAP for Rac binds filamin A to control actin remodelling. Nat. Cell Biol. 2006, 8, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Csepanyi-Komi, R.; Sirokmany, G.; Geiszt, M.; Ligeti, E. ARHGAP25, a novel Rac GTPase-activating protein, regulates phagocytosis in human neutrophilic granulocytes. Blood 2012, 119, 573–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, T.; Nemoto, Y.; Nukiwa, T.; Takeshima, H. Identification and characterization of a novel Rho GTPase activating protein implicated in receptor-mediated endocytosis. FEBS Lett. 2004, 566, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.Y.; Taylor, S.H.; Garva, R.; Holmes, D.F.; Zeef, L.A.; Soininen, R.; Boot-Handford, R.P.; Kadler, K.E. Arhgap28 is a RhoGAP that inactivates RhoA and downregulates stress fibers. PLoS ONE 2014, 9, e107036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saras, J.; Franzén, P.; Aspenstrom, P.; Hellman, U.; Gonez, L.J.; Heldin, C.H. A Novel GTPase-activating Protein for Rho Interacts with a PDZ Domain of the Protein-tyrosine Phosphatase PTPL1. J. Biol. Chem. 1997, 272, 24333–24338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naji, L.; Pacholsky, D.; Aspenstrom, P. ARHGAP30 is a Wrch-1-interacting protein involved in actin dynamics and cell adhesion. Biochem. Biophys. Res. Commun. 2011, 409, 96–102. [Google Scholar] [CrossRef]
- Lamarche-Vane, N.; Hall, A. CdGAP, a Novel Proline-rich GTPase-activating Protein for Cdc42 and Rac. J. Biol. Chem. 1998, 273, 29172–29177. [Google Scholar] [CrossRef] [Green Version]
- Okabe, T.; Nakamura, T.; Nishimura, Y.N.; Kohu, K.; Ohwada, S.; Morishita, Y.; Akiyama, T. RICS, a novel GTPase-activating protein for Cdc42 and Rac1, is involved in the beta-catenin-N-cadherin and N-methyl-D-aspartate receptor signaling. J. Biol. Chem. 2003, 278, 9920–9927. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.H.; Hwang, J.; Legendre, M.; Zhang, M.; Kimura, A.; Saltiel, A.R. TCGAP, a multidomain Rho GTPase-activating protein involved in insulin-stimulated glucose transport. EMBO J. 2003, 22, 2679–2691. [Google Scholar] [CrossRef] [Green Version]
- Guerrier, S.; Coutinho-Budd, J.; Sassa, T.; Gresset, A.; Jordan, N.V.; Chen, K.; Jin, W.L.; Frost, A.; Polleux, F. The F-BAR domain of srGAP2 induces membrane protrusions required for neuronal migration and morphogenesis. Cell 2009, 138, 990–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Mi, Y.J.; Dai, Y.K.; Fu, H.L.; Cui, D.X.; Jin, W.L. The inverse F-BAR domain protein srGAP2 acts through srGAP3 to modulate neuronal differentiation and neurite outgrowth of mouse neuroblastoma cells. PLoS ONE 2013, 8, e57865. [Google Scholar] [CrossRef] [PubMed]
- Mason, F.M.; Heimsath, E.G.; Higgs, H.N.; Soderling, S.H. Bi-modal regulation of a formin by srGAP2. J. Biol. Chem. 2011, 286, 6577–6586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croise, P.; Houy, S.; Gand, M.; Lanoix, J.; Calco, V.; Toth, P.; Brunaud, L.; Lomazzi, S.; Paramithiotis, E.; Chelsky, D.; et al. Cdc42 and Rac1 activity is reduced in human pheochromocytoma and correlates with FARP1 and ARHGEF1 expression. Endocr. Relat. Cancer 2016, 23, 281–293. [Google Scholar] [CrossRef] [Green Version]
- Leung, T.H.; Ching, Y.P.; Yam, J.W.; Wong, C.M.; Yau, T.O.; Jin, D.Y.; Ng, I.O. Deleted in liver cancer 2 (DLC2) suppresses cell transformation by means of inhibition of RhoA activity. Proc. Natl. Acad. Sci. USA 2005, 102, 15207–15212. [Google Scholar] [CrossRef] [Green Version]
- Ching, Y.P.; Wong, C.M.; Chan, S.F.; Leung, T.H.; Ng, D.C.; Jin, D.Y.; Ng, I.O. Deleted in liver cancer (DLC) 2 encodes a RhoGAP protein with growth suppressor function and is underexpressed in hepatocellular carcinoma. J. Biol. Chem. 2003, 278, 10824–10830. [Google Scholar] [CrossRef] [Green Version]
- Kawai, K.; Kiyota, M.; Seike, J.; Deki, Y.; Yagisawa, H. START-GAP3/DLC3 is a GAP for RhoA and Cdc42 and is localized in focal adhesions regulating cell morphology. Biochem. Biophys. Res. Commun. 2007, 364, 783–789. [Google Scholar] [CrossRef]
- Lundstrom, A.; Gallio, M.; Englund, C.; Steneberg, P.; Hemphala, J.; Aspenstrom, P.; Keleman, K.; Falileeva, L.; Dickson, B.J.; Samakovlis, C. Vilse, a conserved Rac/Cdc42 GAP mediating Robo repulsion in tracheal cells and axons. Genes Dev. 2004, 18, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Lenhart, K.C.; Bird, K.E.; Suen, A.A.; Rojas, M.; Kakoki, M.; Li, F.; Smithies, O.; Mack, C.P.; Taylor, J.M. The smooth muscle-selective RhoGAP GRAF3 is a critical regulator of vascular tone and hypertension. Nat. Commun. 2013, 4, 2910. [Google Scholar] [CrossRef] [Green Version]
- Cicchetti, P.; Ridley, A.J.; Zheng, Y.; Cerione, R.A.; Baltimore, D. 3BP-1, an SH3 domain binding protein, has GAP activity for Rac and inhibits growth factor-induced membrane ruffling in fibroblasts. EMBO J. 1995, 14, 3127–3135. [Google Scholar] [CrossRef]
- Parrini, M.C.; Sadou-Dubourgnoux, A.; Aoki, K.; Kunida, K.; Biondini, M.; Hatzoglou, A.; Poullet, P.; Formstecher, E.; Yeaman, C.; Matsuda, M.; et al. SH3BP1, an exocyst-associated RhoGAP, inactivates Rac1 at the front to drive cell motility. Mol. Cell 2011, 42, 650–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raynaud, F.; Moutin, E.; Schmidt, S.; Dahl, J.; Bertaso, F.; Boeckers, T.M.; Homburger, V.; Fagni, L. Rho-GTPase-activating protein interacting with Cdc-42-interacting protein 4 homolog 2 (Rich2): A new Ras-related C3 botulinum toxin substrate 1 (Rac1) GTPase-activating protein that controls dendritic spine morphogenesis. J. Biol. Chem. 2014, 289, 2600–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richnau, N.; Aspenstrom, P. Rich, a rho GTPase-activating protein domain-containing protein involved in signaling by Cdc42 and Rac1. J. Biol. Chem. 2001, 276, 35060–35070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kreuk, B.J.; Schaefer, A.; Anthony, E.C.; Tol, S.; Fernandez-Borja, M.; Geerts, D.; Pool, J.; Hambach, L.; Goulmy, E.; Hordijk, P.L. The human minor histocompatibility antigen 1 is a RhoGAP. PLoS ONE 2013, 8, e73962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aresta, S.; de Tand-Heim, M.F.; Béranger, F.; de Gunzburg, J. A novel Rho GTPase-activating-protein interacts with Gem, a member of the Ras superfamily of GTPases. Biochem. J. 2002, 367, 57–65. [Google Scholar] [CrossRef]
- Bauer, H.; Willert, J.; Koschorz, B.; Herrmann, B.G. The t complex-encoded GTPase-activating protein Tagap1 acts as a transmission ratio distorter in mice. Nat. Genet. 2005, 37, 969–973. [Google Scholar] [CrossRef]
- Tamehiro, N.; Nishida, K.; Yanobu-Takanashi, R.; Goto, M.; Okamura, T.; Suzuki, H. T-cell activation RhoGTPase-activating protein plays an important role in TH17-cell differentiation. Immunol. Cell Biol. 2017, 95, 729–735. [Google Scholar] [CrossRef]
- Corvol, H.; Rousselet, N.; Thompson, K.E.; Berdah, L.; Cottin, G.; Foussigniere, T.; Longchampt, E.; Fiette, L.; Sage, E.; Prunier, C.; et al. FAM13A is a modifier gene of cystic fibrosis lung phenotype regulating rhoa activity, actin cytoskeleton dynamics and epithelial-mesenchymal transition. J. Cyst. Fibros. 2018, 17, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Dyson, J.M.; Fedele, C.G.; Davies, E.M.; Becanovic, J.; Mitchell, C.A. Phosphoinositide Phosphatases: Just as Important as the Kinases. In Phosphoinositides I: Enzymes of Synthesis and Degradation; Balla, T., Wymann, M., York, J.D., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2012; Volume 58, pp. 215–280. [Google Scholar]
- Faucherre, A. Lowe syndrome protein OCRL1 interacts with Rac GTPase in the trans-Golgi network. Hum. Mol. Genet. 2003, 12, 2449–2456. [Google Scholar] [CrossRef]
- Van Rahden, V.A.; Brand, K.; Najm, J.; Heeren, J.; Pfeffer, S.R.; Braulke, T.; Kutsche, K. The 5-phosphatase OCRL mediates retrograde transport of the mannose 6-phosphate receptor by regulating a Rac1-cofilin signalling module. Hum. Mol. Genet. 2012, 21, 5019–5038. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.J.; Callow, M.G.; Souza, B.; Polakis, P. IQGAP1, a calmodulin-binding protein with a rasGAP-related domain, is a potential effector for cdc42Hs. EMBO J. 1996, 15, 2997–3005. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Sundaram, A.; Kudo, M.; Farmer, J.; Ganesan, P.; Khalifeh-Soltani, A.; Arjomandi, M.; Atabai, K.; Huang, X.; Sheppard, D. IQGAP1-dependent scaffold suppresses RhoA and inhibits airway smooth muscle contraction. J. Clin. Investig. 2014, 124, 4895–4898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteel, D.E.; Turner, S.; Schwappacher, R.; Rangaswami, H.; Su-Yuo, J.; Zhuang, S.; Boss, G.R.; Pilz, R.B. Rho isoform-specific interaction with IQGAP1 promotes breast cancer cell proliferation and migration. J. Biol. Chem. 2012, 287, 38367–38378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, S.; Fukata, M.; Kobayashi, K.; Nakafuku, M.; Nomura, N.; Iwamatsu, A.; Kaibuchi, K. Identification of IQGAP as a Putative Target for the Small GTPases, Cdc42 and Rac1. J. Biol. Chem. 1996, 271, 23363–23367. [Google Scholar] [CrossRef] [Green Version]
- Brill, S.; Li, S.; Lyman, C.W.; Church, D.M.; Wasmuth, J.J.; Weissbach, L.; Bernards, A.; Snijders, A.J. The Ras GTPase-Activating-Protein-Related Human Protein IQGAP2 Harbors a Potential Actin Binding Domain and Interacts with Calmodulin and Rho Family GTPases. Mol. Cell. Biol. 1996, 16, 4869–4878. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Tao, Y.; Chen, Y.; Xu, W. RhoC regulates the proliferation of gastric cancer cells through interaction with IQGAP1. PLoS ONE 2012, 7, e48917. [Google Scholar] [CrossRef]
- Wang, S.; Watanabe, T.; Noritake, J.; Fukata, M.; Yoshimura, T.; Itoh, N.; Harada, T.; Nakagawa, M.; Matsuura, Y.; Arimura, N.; et al. IQGAP3, a novel effector of Rac1 and Cdc42, regulates neurite outgrowth. J. Cell Sci. 2007, 120, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, J.; Scheel, A.A.; Diekmann, D.; Hall, A.; Ruppert, C.; Bähler, M. A novel type of myosin implicated in signalling by rho family GTPases. EMBO J. 1995, 14, 697–704. [Google Scholar] [CrossRef]
- Zheng, Y.; Bagrodia, S.; Cerione, R.A. Activation of Phosphoinositide 3-Kinase Activity by Cdc42Hs Binding to p85. J. Biol. Chem. 1994, 269, 18727–18730. [Google Scholar]
- Chamberlain, M.D.; Berry, T.R.; Pastor, M.C.; Anderson, D.H. The p85alpha subunit of phosphatidylinositol 3’-kinase binds to and stimulates the GTPase activity of Rab proteins. J. Biol. Chem. 2004, 279, 48607–48614. [Google Scholar] [CrossRef] [Green Version]
- Cariaga-Martinez, A.E.; Cortes, I.; Garcia, E.; Perez-Garcia, V.; Pajares, M.J.; Idoate, M.A.; Redondo-Munoz, J.; Anton, I.M.; Carrera, A.C. Phosphoinositide 3-kinase p85beta regulates invadopodium formation. Biol. Open 2014, 3, 924–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touré, A.; Dorseuil, O.; Morin, L.; Timmons, P.; Jégou, B.; Reibel, L.; Gacon, G. MgcRacGAP, A New Human GTPase-activating Protein for Rac and Cdc42 Similar to Drosophila rotundRacGAP Gene Product, Is Expressed in Male Germ Cells. J. Biol. Chem. 1998, 273, 6019–6023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, C.D.; Fan, C.; Mitin, N.; Baker, N.M.; George, S.D.; Graham, D.M.; Perou, C.M.; Burridge, K.; Der, C.J.; Rossman, K.L. Rho GTPase Transcriptome Analysis Reveals Oncogenic Roles for Rho GTPase-Activating Proteins in Basal-like Breast Cancers. Cancer Res. 2016, 76, 3826–3837. [Google Scholar] [CrossRef] [Green Version]
- Jullien-Flores, V.; Dorseuil, O.; Romero, F.; Letourneur, F.; Saragosti, S.; Berger, R.; Tavitian, A.; Gacon, G.; Camonis, J.H. Bridging Ral GTPase to Rho pathways. RLIP76, a Ral effector with CDC42/Rac GTPase-activating protein activity. J. Biol. Chem. 1995, 270, 22473–22477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topp, J.D.; Gray, N.W.; Gerard, R.D.; Horazdovsky, B.F. Alsin is a Rab5 and Rac1 guanine nucleotide exchange factor. J. Biol. Chem. 2004, 279, 24612–24623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadano, S.; Otomo, A.; Suzuki-Utsunomiya, K.; Kunita, R.; Yanagisawa, Y.; Showguchi-Miyata, J.; Mizumura, H.; Ikeda, J.E. ALS2CL, the novel protein highly homologous to the carboxy-terminal half of ALS2, binds to Rab5 and modulates endosome dynamics. FEBS Lett. 2004, 575, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.J.; Sharma, S.; elMasry, N.; Qiu, R.G.; McCabe, P.; Polakis, P.; Bollag, G. Identification of a Novel Guanine Nucleotide Exchange Factor for the Rho GTPase. J. Biol. Chem. 1996, 271, 25452–25458. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, M.; Gremer, L.; Dvorsky, R.; Haeusler, L.C.; Cirstea, I.C.; Uhlenbrock, K.; Ahmadian, M.R. Mechanistic insights into specificity, activity, and regulatory elements of the regulator of G-protein signaling (RGS)-containing Rho-specific guanine nucleotide exchange factors (GEFs) p115, PDZ-RhoGEF (PRG), and leukemia-associated RhoGEF (LARG). J. Biol. Chem. 2011, 286, 18202–18212. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, R.; Zheng, Y.; Busch, H. Cloning and Characterization of GEF-H1, a Microtubule-associated Guanine Nucleotide Exchange Factor for Rac and Rho GTPases. J. Biol. Chem. 1998, 273, 34954–34960. [Google Scholar] [CrossRef] [Green Version]
- Dubois, F.; Keller, M.; Calvayrac, O.; Soncin, F.; Hoa, L.; Hergovich, A.; Parrini, M.C.; Mazieres, J.; Vaisse-Lesteven, M.; Camonis, J.; et al. RASSF1A Suppresses the Invasion and Metastatic Potential of Human Non-Small Cell Lung Cancer Cells by Inhibiting YAP Activation through the GEF-H1/RhoB Pathway. Cancer Res. 2016, 76, 1627–1640. [Google Scholar] [CrossRef] [Green Version]
- Kamon, H.; Kawabe, T.; Kitamura, H.; Lee, J.; Kamimura, D.; Kaisho, T.; Akira, S.; Iwamatsu, A.; Koga, H.; Murakami, M.; et al. TRIF-GEFH1-RhoB pathway is involved in MHCII expression on dendritic cells that is critical for CD4 T-cell activation. EMBO J. 2006, 25, 4108–4119. [Google Scholar] [CrossRef] [Green Version]
- Arthur, W.T.; Ellerbroek, S.M.; Der, C.J.; Burridge, K.; Wennerberg, K. XPLN, a guanine nucleotide exchange factor for RhoA and RhoB, but not RhoC. J. Biol. Chem. 2002, 277, 42964–42972. [Google Scholar] [CrossRef] [Green Version]
- Gotthardt, K.; Ahmadian, M.R. Asef is a Cdc42-specific guanine nucleotide exchange factor. Biol. Chem. 2007, 388. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Senda, T.; Ishidate, T.; Koyama, R.; Morishita, T.; Iwayama, Y.; Higuchi, O.; Akiyama, T. Asef, a Link Between the Tumor Suppressor APC and G-Protein Signaling. Science 2000, 289, 1194–1197. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Kumamoto, Y.; Wang, P.; Gan, X.; Lehmann, D.; Smrcka, A.V.; Cohn, L.; Iwasaki, A.; Li, L.; Wu, D. Regulation of immature dendritic cell migration by RhoA guanine nucleotide exchange factor Arhgef5. J. Biol. Chem. 2009, 284, 28599–28606. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Chang, S.W.; Tatsumoto, T.; Chan, A.M.; Miki, T. TIM, a Dbl-related protein, regulates cell shape and cytoskeletal organization in a Rho-dependent manner. Cell. Signal. 2005, 17, 461–471. [Google Scholar] [CrossRef]
- Debily, M.A. Expression and molecular characterization of alternative transcripts of the ARHGEF5/TIM oncogene specific for human breast cancer. Hum. Mol. Genet. 2003, 13, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manser, E.; Loo, T.H.; Koh, C.G.; Zhao, Z.S.; Chen, X.Q.; Tan, L.; Tan, I.; Leung, T.; Lim, L. PAK Kinases Are Directly Coupled to the PIX Family of Nucleotide Exchange Factors. Mol. Cell 1998, 1, 183–192. [Google Scholar] [CrossRef]
- Alberts, A.S.; Treisman, R. Activation of RhoA and SAPK/JNK signalling pathways by the RhoA-specific exchange factor mNET1. EMBO J. 1998, 17, 4075–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srougi, M.C.; Burridge, K. The nuclear guanine nucleotide exchange factors Ect2 and Net1 regulate RhoB-mediated cell death after DNA damage. PLoS ONE 2011, 6, e17108. [Google Scholar] [CrossRef]
- Reid, T.; Bathoorn, A.; Ahmadian, M.R.; Collard, J.G. Identification and Characterization of hPEM-2, a Guanine Nucleotide Exchange Factor Specific for Cdc42. J. Biol. Chem. 1999, 274, 33587–33593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohl, M.; Winkler, S.; Wieland, T.; Lutz, S. Gef10—the third member of a Rho-specific guanine nucleotide exchange factor subfamily with unusual protein architecture. Naunyn Schmiedebergs Arch. Pharmacol. 2006, 373, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Winkler, S.; Mohl, M.; Wieland, T.; Lutz, S. GrinchGEF—a novel Rho-specific guanine nucleotide exchange factor. Biochem. Biophys. Res. Commun. 2005, 335, 1280–1286. [Google Scholar] [CrossRef] [PubMed]
- Oleksy, A.; Opalinski, L.; Derewenda, U.; Derewenda, Z.S.; Otlewski, J. The molecular basis of RhoA specificity in the guanine nucleotide exchange factor PDZ-RhoGEF. J. Biol. Chem. 2006, 281, 32891–32897. [Google Scholar] [CrossRef] [Green Version]
- Kristelly, R.; Gao, G.; Tesmer, J.J. Structural determinants of RhoA binding and nucleotide exchange in leukemia-associated Rho guanine-nucleotide exchange factor. J. Biol. Chem. 2004, 279, 47352–47362. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Olson, M.F.; Hall, A.; Cerione, R.A.; Toksoz, D. Direct involvement of the small GTP-binding protein Rho in lbc oncogene function. J. Biol. Chem. 1995, 270, 9031–9034. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, I.P.; Lambert, Q.T.; Glaven, J.A.; Abe, K.; Rossman, K.L.; Mahon, G.M.; Trzaskos, J.M.; Kay, R.; Campbell, S.L.; Der, C.J. Dependence of Dbl and Dbs Transformation on MEK and NF-κB Activation. Mol. Cell. Biol. 1999, 19, 7759–7770. [Google Scholar] [CrossRef] [Green Version]
- Ogita, H.; Kunimoto, S.; Kamioka, Y.; Sawa, H.; Masuda, M.; Mochizuki, N. EphA4-mediated Rho activation via Vsm-RhoGEF expressed specifically in vascular smooth muscle cells. Circ. Res. 2003, 93, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Kusuhara, S.; Fukushima, Y.; Fukuhara, S.; Jakt, L.M.; Okada, M.; Shimizu, Y.; Hata, M.; Nishida, K.; Negi, A.; Hirashima, M.; et al. Arhgef15 promotes retinal angiogenesis by mediating VEGF-induced Cdc42 activation and potentiating RhoJ inactivation in endothelial cells. PLoS ONE 2012, 7, e45858. [Google Scholar] [CrossRef] [Green Version]
- Margolis, S.S.; Salogiannis, J.; Lipton, D.M.; Mandel-Brehm, C.; Wills, Z.P.; Mardinly, A.R.; Hu, L.; Greer, P.L.; Bikoff, J.B.; Ho, H.-Y.H.; et al. EphB-Mediated Degradation of the RhoA GEF Ephexin5 Relieves a Developmental Brake on Excitatory Synapse Formation. Cell 2010, 143, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Hiramoto-Yamaki, N.; Takeuchi, S.; Ueda, S.; Harada, K.; Fujimoto, S.; Negishi, M.; Katoh, H. Ephexin4 and EphA2 mediate cell migration through a RhoG-dependent mechanism. J. Cell Biol. 2010, 190, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.W.; He, X.; Borthwick, K.; Donne, A.J.; Hampson, L.; Hampson, I.N. The HPV16 E6 binding protein Tip-1 interacts with ARHGEF16, which activates Cdc42. Br. J. Cancer 2011, 104, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Park, B.; Kim, G.; Kim, K.; Pak, J.; Kim, K.; Ye, M.B.; Park, S.G.; Park, D. Arhgef16, a novel Elmo1 binding partner, promotes clearance of apoptotic cells via RhoG-dependent Rac1 activation. Biochim. Biophys. Acta 2014, 1843, 2438–2447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rümenapp, U.; Freichel-Blomquist, A.; Wittinghofer, B.; Jakobs, K.H.; Wieland, T. A mammalian Rho-specific guanine-nucleotide exchange factor (p164-RhoGEF) without a pleckstrin homology domain. Biochem. J. 2002, 366, 721–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitin, N.; Rossman, K.L.; Der, C.J. Identification of a novel actin-binding domain within the Rho guanine nucleotide exchange factor TEM4. PLoS ONE 2012, 7, e41876. [Google Scholar] [CrossRef] [PubMed]
- Lutz, S.; Mohl, M.; Rauch, J.; Weber, P.; Wieland, T. RhoGEF17, a Rho-specific guanine nucleotide exchange factor activated by phosphorylation via cyclic GMP-dependent kinase Ialpha. Cell. Signal. 2013, 25, 630–638. [Google Scholar] [CrossRef]
- Niu, J.; Profirovic, J.; Pan, H.; Vaiskunaite, R.; Voyno-Yasenetskaya, T. G Protein betagamma subunits stimulate p114RhoGEF, a guanine nucleotide exchange factor for RhoA and Rac1: Regulation of cell shape and reactive oxygen species production. Circ. Res. 2003, 93, 848–856. [Google Scholar] [CrossRef] [Green Version]
- Blomquist, A.; Schwörer, G.; Schablowski, H.; Psoma, A.; Lehnen, M.; Jakobs, K.H.; Rümenapp, U. Identification and characterization of a novel Rho-specific guanine nucleotide exchange factor. Biochem. J. 2000, 352, 319–325. [Google Scholar] [CrossRef]
- Wang, Y.; Suzuki, H.; Yokoo, T.; Tada-Iida, K.; Kihara, R.; Miura, M.; Watanabe, K.; Sone, H.; Shimano, H.; Toyoshima, H.; et al. WGEF is a novel RhoGEF expressed in intestine, liver, heart, and kidney. Biochem. Biophys. Res. Commun. 2004, 324, 1053–1058. [Google Scholar] [CrossRef]
- Zheng, Y.; Hart, M.J.; Cerione, R.A. Guanine Nucleotide Exchange Catalyzed by dbl Oncogene Product. Methods Enzymol. 1995, 256, 77–84. [Google Scholar]
- Hart, M.J.; Eva, A.; Evans, T.; Aaronson, S.A.; Cerione, R.A. Catalysis of guanine nucleotide exchange on the CDC42Hs protein by the dbl oncogene product. Nature 1991, 354, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Cao, J. Studies on the Regulatory Mechanism of the ULK1 Complex in the Induction of Autophagy. Ph.D. Thesis, University of Minnesota Digital Conservancy, Minneapolis, MN, USA, 2012. [Google Scholar]
- Raimondi, F.; Felline, A.; Fanelli, F. Catching Functional Modes and Structural Communication in Dbl Family Rho Guanine Nucleotide Exchange Factors. J. Chem. Inf. Model. 2015, 55, 1878–1893. [Google Scholar] [CrossRef] [PubMed]
- Debant, A.; Serra-Pagès, C.; Seipel, K.; O’Brien, S.; Tang, M.; Park, S.H.; Streuli, M. The multidomain protein Trio binds the LAR transmembrane tyrosine phosphatase, contains a protein kinase domain, and has separate rac-specific and rho-specific guanine nucleotide exchange factor domains. Proc. Natl. Acad. Sci. USA 1996, 93, 5466–5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, J.; Van Vactor, D. The Trio family of guanine-nucleotide-exchange factors: Regulators of axon guidance. J. Cell Sci. 2001, 114, 1973–1980. [Google Scholar]
- May, V.; Schiller, M.R.; Eipper, B.A.; Mains, R.E. Kalirin Dbl-Homology Guanine Nucleotide Exchange Factor 1 Domain Initiates New Axon Outgrowths via RhoG-Mediated Mechanisms. J. Neurosci. 2002, 22, 6980–6990. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.M.; Huang, J.; Wang, Y.; Eipper, B.A.; Mains, R.E. Kalirin, a Multifunctional Rho Guanine Nucleotide Exchange Factor, Is Necessary for Maintenance ofHippocampal Pyramidal Neuron Dendrites and Dendritic Spines. J. Neurosci. 2003, 23, 10593–10603. [Google Scholar] [CrossRef]
- Colomer, V.; Engelender, S.; Sharp, A.H.; Duan, K.; Cooper, J.K.; Lanahan, A.; Lyford, G.; Worley, P.; Ross, C.A. Huntingtin-associated protein 1 (HAP1) binds to a Trio-like polypeptide, with a rac1 guanine nucleotide exchange factor domain. Hum. Mol. Genet. 1997, 6, 1519–1525. [Google Scholar] [CrossRef] [Green Version]
- Souchet, M.; Portales-Casamar, E.; Mazurais, D.; Schmidt, S.; Léger, I.; Javré, J.L.; Robert, P.; Berrebi-Bertrand, I.; Bril, A.; Gout, B.; et al. Human p63RhoGEF, a novel RhoA-specific guanine nucleotide exchange factor, is localized in cardiac sarcomere. J. Cell Sci. 2002, 115, 629–640. [Google Scholar]
- Guo, X.; Stafford, L.J.; Bryan, B.; Xia, C.; Ma, W.; Wu, X.; Liu, D.; Songyang, Z.; Liu, M. A Rac/Cdc42-specific exchange factor, GEFT, induces cell proliferation, transformation, and migration. J. Biol. Chem. 2003, 278, 13207–13215. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Jin, R.; Qu, G.; Wang, X.; Li, Z.; Yuan, Z.; Zhao, C.; Siwko, S.; Shi, T.; Wang, P.; et al. GPR116, an Adhesion G-Protein-Coupled Receptor, Promotes Breast Cancer Metastasis via the G q-p63RhoGEF-Rho GTPase Pathway. Cancer Res. 2013, 73, 6206–6218. [Google Scholar] [CrossRef] [Green Version]
- Ellerbroek, S.M.; Wennerberg, K.; Arthur, W.T.; Dunty, J.M.; Bowman, D.R.; DeMali, K.A.; Der, C.; Burridge, K. SGEF, a RhoG guanine nucleotide exchange factor that stimulates macropinocytosis. Mol. Biol. Cell 2004, 15, 3309–3319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamah, S.M.; Lin, M.Z.; Goldberg, J.L.; Estrach, S.; Sahin, M.; Hu, L.; Bazalakova, M.; Neve, R.L.; Corfas, G.; Debant, A.; et al. EphA Receptors Regulate Growth Cone Dynamics through the Novel Guanine Nucleotide Exchange Factor Ephexin. Cell 2001, 105, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Dada, O.; Gutowski, S.; Brautigam, C.A.; Chen, Z.; Sternweis, P.C. Direct regulation of p190RhoGEF by activated Rho and Rac GTPases. J. Struct. Biol. 2018, 202, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Sagara, M.; Shibata, Y.; Shirouzu, M.; Yokoyama, S.; Akiyama, T. Identification and characterization of Asef2, a guanine-nucleotide exchange factor specific for Rac1 and Cdc42. Oncogene 2007, 26, 7620–7627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bristow, J.M.; Sellers, M.H.; Majumdar, D.; Anderson, B.; Hu, L.; Webb, D.J. The Rho-family GEF Asef2 activates Rac to modulate adhesion and actin dynamics and thereby regulate cell migration. J. Cell Sci. 2009, 122, 4535–4546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford-Speelman, D.L.; Roche, J.A.; Bowman, A.L.; Bloch, R.J. The rho-guanine nucleotide exchange factor domain of obscurin activates rhoA signaling in skeletal muscle. Mol. Biol. Cell 2009, 20, 3905–3917. [Google Scholar] [CrossRef] [Green Version]
- Coisy-Quivy, M.; Touzet, O.; Bourret, A.; Hipskind, R.A.; Mercier, J.; Fort, P.; Philips, A. TC10 controls human myofibril organization and is activated by the sarcomeric RhoGEF obscurin. J. Cell Sci. 2009, 122, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Solski, P.A.; Wilder, R.S.; Rossman, K.L.; Sondek, J.; Cox, A.D.; Campbell, S.L.; Der, C.J. Requirement for C-terminal sequences in regulation of Ect2 guanine nucleotide exchange specificity and transformation. J. Biol. Chem. 2004, 279, 25226–25233. [Google Scholar] [CrossRef] [Green Version]
- Tatsumoto, T.; Xie, X.; Blumenthal, R.; Okamoto, I.; Miki, T. Human ECT2 Is an Exchange Factor for Rho GTPases, Phosphorylated in G2/M Phases, and Involved in Cytokinesis. J. Cell Biol. 1999, 147, 921–928. [Google Scholar] [CrossRef]
- Wennerberg, K.; Ellerbroek, S.M.; Liu, R.Y.; Karnoub, A.E.; Burridge, K.; Der, C.J. RhoG signals in parallel with Rac1 and Cdc42. J. Biol. Chem. 2002, 277, 47810–47817. [Google Scholar] [CrossRef] [Green Version]
- Miki, T.; Smith, C.L.; Long, J.E.; Eva, A.; Fleming, T.P. Oncogene ect2 is related to regulators of small GTP-binding proteins. Nature 1993, 362, 462–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, M.A.; Kwiatkowski, A.V.; Pellegrini, L.; Cestra, G.; Butler, M.H.; Rossman, K.L.; Serna, D.M.; Sondek, J.; Gertler, F.B.; De Camilli, P. Tuba, a novel protein containing bin/amphiphysin/Rvs and Dbl homology domains, links dynamin to regulation of the actin cytoskeleton. J. Biol. Chem. 2003, 278, 49031–49043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Cai, L.; Zhang, X.; Li, A.; Miao, Y.; Li, Q.; Qiu, X.; Wang, E. ARHGEF39 promotes tumor progression via activation of Rac1/P38 MAPK/ATF2 signaling and predicts poor prognosis in non-small cell lung cancer patients. Lab. Investig. 2018, 98, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Curtis, C.; Hemmeryckx, B.; Haataja, L.; Senadheera, D.; Groffen, J.; Heisterkamp, N. Scambio, a novel guanine nucleotide exchange factor for Rho. Mol. Cancer 2004, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.J.; Nishikawa, K.; Yuda, H.; Wang, Y.L.; Osaka, H.; Fukazawa, N.; Naito, A.; Kudo, Y.; Wada, K.; Aoki, S. Solo/Trio8, a membrane-associated short isoform of Trio, modulates endosome dynamics and neurite elongation. Mol. Cell. Biol. 2006, 26, 6923–6935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abiko, H.; Fujiwara, S.; Ohashi, K.; Hiatari, R.; Mashiko, T.; Sakamoto, N.; Sato, M.; Mizuno, K. Rho guanine nucleotide exchange factors involved in cyclic-stretch-induced reorientation of vascular endothelial cells. J. Cell Sci. 2015, 128, 1683–1695. [Google Scholar] [CrossRef] [Green Version]
- Ueda, H.; Nagae, R.; Kozawa, M.; Morishita, R.; Kimura, S.; Nagase, T.; Ohara, O.; Yoshida, S.; Asano, T. Heterotrimeric G protein betagamma subunits stimulate FLJ00018, a guanine nucleotide exchange factor for Rac1 and Cdc42. J. Biol. Chem. 2008, 283, 1946–1953. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Park, W.S.; Park, B.O.; Kim, C.Y.; Oh, Y.; Kim, J.M.; Choi, H.; Kyung, T.; Kim, C.H.; Lee, G.; et al. PLEKHG3 enhances polarized cell migration by activating actin filaments at the cell front. Proc. Natl. Acad. Sci. USA 2016, 113, 10091–10096. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.; Kamynina, E.; Morley, S.; Chung, S.; Muakkassa, N.; Wang, H.; Brathwaite, S.; Sharma, G.; Manor, D. Plekhg4 is a novel Dbl family guanine nucleotide exchange factor protein for rho family GTPases. J. Biol. Chem. 2013, 288, 14522–14530. [Google Scholar] [CrossRef] [Green Version]
- Mavrakis, K.J.; McKinlay, K.J.; Jones, P.; Sablitzky, F. DEF6, a novel PH-DH-like domain protein, is an upstream activator of the Rho GTPases Rac1, Cdc42, and RhoA. Exp. Cell Res. 2004, 294, 335–344. [Google Scholar] [CrossRef]
- Gupta, S.; Fanzo, J.C.; Hu, C.; Cox, D.; Jang, S.Y.; Lee, A.E.; Greenberg, S.; Pernis, A.B. T cell receptor engagement leads to the recruitment of IBP, a novel guanine nucleotide exchange factor, to the immunological synapse. J. Biol. Chem. 2003, 278, 43541–43549. [Google Scholar] [CrossRef] [Green Version]
- Koyano, Y.; Kawamoto, T.; Shen, M.; Yan, W.; Noshiro, M.; Fujii, K.; Kato, Y. Molecular Cloning and Characterization of CDEP, a Novel Human Protein Containing the Ezrin-like Domain of the Band 4.1 Superfamily and the Dbl Homology Domain of Rho Guanine Nucleotide Exchange Factors. Biochem. Biophys. Res. Commun. 1997, 241, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Cheadle, L.; Biederer, T. The novel synaptogenic protein Farp1 links postsynaptic cytoskeletal dynamics and transsynaptic organization. J. Cell Biol. 2012, 199, 985–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, Y.; Yamauchi, J.; Itoh, H. Src kinase regulates the activation of a novel FGD-1-related Cdc42 guanine nucleotide exchange factor in the signaling pathway from the endothelin A receptor to JNK. J. Biol. Chem. 2003, 278, 29890–29900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, T.; Yamashita, T.; Yamaguchi, A.; Sumimoto, H.; Hosokawa, K.; Tohyama, M. A Novel FERM Domain Including Guanine Nucleotide Exchange Factor Is Involved in Rac Signaling and Regulates Neurite Remodeling. J. Neurosci. 2002, 22, 8504–8513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Fischer, D.J.; Santos, M.F.; Tigyi, G.; Pasteris, N.G.; Gorski, J.L.; Xu, Y. The Faciogenital Dysplasia Gene Product FGD1 Functions as a Cdc42Hs-specific GuanineNucleotide Exchange Factor. J. Biol. Chem. 1996, 271, 33169–33172. [Google Scholar] [CrossRef] [Green Version]
- Huber, C.; Martensson, A.; Bokoch, G.M.; Nemazee, D.; Gavin, A.L. FGD2, a CDC42-specific exchange factor expressed by antigen-presenting cells, localizes to early endosomes and active membrane ruffles. J. Biol. Chem. 2008, 283, 34002–34012. [Google Scholar] [CrossRef] [Green Version]
- Pasteris, N.G.; Nagata, K.; Hall, A.; Gorski, J.L. Isolation, characterization, and mapping of the mouse Fgd3 gene, a new Faciogenital Dysplasia (FGD1; Aarskog Syndrome) gene homologue. Gene 2000, 242, 237–247. [Google Scholar] [CrossRef]
- Ono, Y.; Nakanishi, H.; Nishimura, M.; Kakizaki, M.; Takahashi, K.; Miyahara, M.; Satoh-Horikawa, K.; Mandai, K.; Takai, Y. Two actions of frabin: Direct activation of Cdc42 and indirect activation of Rac. Oncogene 2000, 19, 3050–3058. [Google Scholar] [CrossRef] [Green Version]
- Kurogane, Y.; Miyata, M.; Kubo, Y.; Nagamatsu, Y.; Kundu, R.K.; Uemura, A.; Ishida, T.; Quertermous, T.; Hirata, K.; Rikitake, Y. FGD5 mediates proangiogenic action of vascular endothelial growth factor in human vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 988–996. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Guo, Y.; Negre, J.; Preto, J.; Smithers, C.C.; Azad, A.K.; Overduin, M.; Murray, A.G.; Eitzen, G. Fgd5 is a Rac1-specific Rho GEF that is selectively inhibited by aurintricarboxylic acid. Small GTPases 2019. [Google Scholar] [CrossRef] [PubMed]
- Steenblock, C.; Heckel, T.; Czupalla, C.; Espirito Santo, A.I.; Niehage, C.; Sztacho, M.; Hoflack, B. The Cdc42 guanine nucleotide exchange factor FGD6 coordinates cell polarity and endosomal membrane recycling in osteoclasts. J. Biol. Chem. 2014, 289, 18347–18359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, N.K.; Jenna, S.; Glogauer, M.; Quinn, C.C.; Wasiak, S.; Guipponi, M.; Antonarakis, S.E.; Kay, B.K.; Strossel, T.P.; Lamarche-Vane, N.; et al. Endocytic protein intersectin-l regulates actin assembly via Cdc42 and N-WASP. Nat. Cell Biol. 2001, 3, 927–932. [Google Scholar] [CrossRef]
- McGavin, M.K.; Badour, K.; Hardy, L.A.; Kubiseski, T.J.; Zhang, J.; Siminovitch, K.A. The Intersectin 2 Adaptor Links Wiskott Aldrich Syndrome Protein (WASp)-mediated Actin Polymerization to T Cell Antigen Receptor Endocytosis. J. Exp. Med. 2001, 194, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Kwak, D.; Noh, J.; Lee, M.N.; Lee, C.S.; Suh, P.G.; Ryu, S.H. Phospholipase D2 induces stress fiber formation through mediating nucleotide exchange for RhoA. Cell. Signal. 2011, 23, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Mahankali, M.; Peng, H.J.; Henkels, K.M.; Dinauer, M.C.; Gomez-Cambronero, J. Phospholipase D2 (PLD2) is a guanine nucleotide exchange factor (GEF) for the GTPase Rac2. Proc. Natl. Acad. Sci. USA 2011, 108, 19617–19622. [Google Scholar] [CrossRef] [Green Version]
- De Toledo, M.; Coulon, V.; Schmidt, S.; Fort, P.; Blangy, A. The gene for a new brain specific RhoA exchange factor maps to the highly unstable chromosomal region 1p36.2–1p36.3. Oncogene 2001, 20, 7307–7317. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Asiedu, M.; Adelstein, R.S.; Wei, Q. A Novel Guanine Nucleotide Exchange Factor MyoGEF is Required for Cytokinesis. Cell Cycle 2006, 5, 1234–1239. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, R.; Aresta, S.; Blangy, A.; Del Maestro, L.; Louvard, D.; Arpin, M. Interaction of ezrin with the novel guanine nucleotide exchange factor PLEKHG6 promotes RhoG-dependent apical cytoskeleton rearrangements in epithelial cells. Mol. Biol. Cell 2007, 18, 4780–4793. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Asiedu, M.; Wei, Q. Myosin-interacting guanine exchange factor (MyoGEF) regulates the invasion activity of MDA-MB-231 breast cancer cells through activation of RhoA and RhoC. Oncogene 2009, 28, 2219–2230. [Google Scholar] [CrossRef] [Green Version]
- Welch, H.C.; Coadwell, W.J.; Ellson, C.D.; Ferguson, G.J.; Andrews, S.R.; Erdjument-Bromage, H.; Tempst, P.; Hawkins, P.T.; Stephens, L.R. P-Rex1, a PtdIns(3,4,5)P3- and Gbetagamma-regulated guanine-nucleotide exchange factor for Rac. Cell 2002, 108, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Donald, S.; Hill, K.; Lecureuil, C.; Barnouin, R.; Krugmann, S.; John Coadwell, W.; Andrews, S.R.; Walker, S.A.; Hawkins, P.T.; Stephens, L.R.; et al. P-Rex2, a new guanine-nucleotide exchange factor for Rac. FEBS Lett. 2004, 572, 172–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeldt, H.; Vázquez-Prado, J.; Gutkind, J.S. P-REX2, a novel PI-3-kinase sensitive Rac exchange factor. FEBS Lett. 2004, 572, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Fine, B.; Hodakoski, C.; Koujak, S.; Su, T.; Saal, L.H.; Maurer, M.; Hopkins, B.; Keniry, M.; Sulis, M.L.; Mense, S.; et al. Activation of the PI3K pathway in cancer through inhibition of PTEN by exchange factor P-REX2a. Science 2009, 325, 1261–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyono, M.; Satoh, T.; Kaziro, Y. G protein beta gamma subunit-dependent Rac-guanine nucleotide exchange activity of Ras-GRF1/CDC25(Mm). Proc. Natl. Acad. Sci. USA 1999, 96, 4826–4831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, F.; Sanz-Moreno, V.; Agudo-Ibanez, L.; Wallberg, F.; Sahai, E.; Marshall, C.J.; Crespo, P. RasGRF suppresses Cdc42-mediated tumour cell movement, cytoskeletal dynamics and transformation. Nat. Cell Biol. 2011, 13, 819–826. [Google Scholar] [CrossRef]
- Fan, W.T.; Koch, C.A.; de Hoog, C.L.; Fam, N.P.; Moran, M.F. The exchange factor Ras-GRF2 activates Ras-dependent and Rac-dependent mitogen-activated protein kinase pathways. Curr. Biol. 1998, 8, 935–938. [Google Scholar] [CrossRef] [Green Version]
- Hamel, B.; Monaghan-Benson, E.; Rojas, R.J.; Temple, B.R.; Marston, D.J.; Burridge, K.; Sondek, J. SmgGDS is a guanine nucleotide exchange factor that specifically activates RhoA and RhoC. J. Biol. Chem. 2011, 286, 12141–12148. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Yin, Z.; Zhao, B.; Qi, Y.; Liu, J.; Rahimi, S.A.; Lee, L.Y.; Li, S. Microgravity simulation activates Cdc42 via Rap1GDS1 to promote vascular branch morphogenesis during vasculogenesis. Stem Cell Res. 2017, 25, 157–165. [Google Scholar] [CrossRef]
- Nimnual, A.S.; Yatsula, B.A.; Bar-Sagi, D. Coupling of Ras and Rac Guanosine Triphosphatases Through the Ras Exchanger Sos. Science 1998, 279, 560–563. [Google Scholar] [CrossRef]
- Toffali, L.; Montresor, A.; Mirenda, M.; Scita, G.; Laudanna, C. SOS1, ARHGEF1, and DOCK2 rho-GEFs Mediate JAK-Dependent LFA-1 Activation by Chemokines. J. Immunol. 2017, 198, 708–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, M.; Terada, Y.; Iwamatsu, A.; Shinohara, A.; Mochizuki, N.; Higuchi, M.; Gotoh, Y.; Ihara, S.; Nagata, S.; Itoh, H.; et al. SWAP-70 is a guanine-nucleotide exchange factor that mediates signalling of membrane ruffling. Nature 2002, 416, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Sivalenka, R.R.; Jessberger, R. SWAP-70 regulates c-kit-induced mast cell activation, cell-cell adhesion, and migration. Mol. Cell. Biol. 2004, 24, 10277–10288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, F.; Habets, G.G.; Stam, J.C.; van der Kammen, R.A.; Collard, J.G. A role for Rac in Tiam1-induced membrane ruffling and invasion. Nature 1995, 375, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, M.; Sone, M.; Fukata, M.; Kuroda, S.; Kaibuchi, K.; Nabeshima, Y.; Hama, C. Identification of the stef Gene That Encodes a Novel Guanine Nucleotide Exchange Factor Specific for Rac1. J. Biol. Chem. 1999, 274, 17837–17844. [Google Scholar] [CrossRef] [Green Version]
- Woroniuk, A.; Porter, A.; White, G.; Newman, D.T.; Diamantopoulou, Z.; Waring, T.; Rooney, C.; Strathdee, D.; Marston, D.J.; Hahn, K.M.; et al. STEF/TIAM2-mediated Rac1 activity at the nuclear envelope regulates the perinuclear actin cap. Nat. Commun. 2018, 9, 2124. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Goto, T.M.; Sugimoto, M.; Nishimura, T.; Shinagawa, T.; Ohno, S.; Amano, M.; Kaibuchi, K. Rho-kinase phosphorylates PAR-3 and disrupts PAR complex formation. Dev. Cell 2008, 14, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Snyder, J.T.; Worthylake, D.K.; Rossman, K.L.; Betts, L.; Pruitt, W.M.; Siderovski, D.P.; Der, C.J.; Sondek, J. Structural basis for the selective activation of Rho GTPases by Dbl exchange factors. Nat. Struct. Biol. 2002, 9, 468–475. [Google Scholar] [CrossRef]
- Crespo, P.; Schuebel, K.E.; Ostrom, A.A.; Gutkind, J.S.; Bustelo, X.R. Phosphotyrosine-dependent activation of Rac-l GDP/GTP exchange by the vav proto-oncogene product. Nature 1997, 385, 169–172. [Google Scholar] [CrossRef]
- Schuebel, K.E.; Movilla, N.; Rosa, J.L.; Bustelo, X.R. Phosphorylation-dependent and constitutive activation of Rho proteins by wild-type and oncogenic Vav-2. EMBO J. 1998, 17, 6608–6621. [Google Scholar] [CrossRef] [Green Version]
- Movilla, N.; Bustelo, X.R. Biological and Regulatory Properties of Vav-3, a New Member of the Vav Family of Oncoproteins. Mol. Cell. Biol. 1999, 19, 7870–7885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, L.; Sachdev, P.; Yan, L.; Chan, J.L.; Trenkle, T.; McClelland, M.; Welsh, J.; Wang, L.H. Vav3 Mediates Receptor Protein Tyrosine Kinase Signaling, Regulates GTPase Activity, Modulates Cell Morphology, and Induces Cell Transformation. Mol. Cell. Biol. 2000, 20, 9212–9224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyokawa, E.; Hashimoto, Y.; Kobayashi, S.; Sugimura, H.; Kurata, T.; Matsuda, M. Activation of Rac1 by a Crk SH3-binding protein, DOCK180. Genes Dev. 1998, 12, 3331–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, L.; Fu, H.; Yan, J.; Wang, Y.; Guo, H.; Hao, X.; Xu, X.; Jin, T.; Zhang, N. Association between Gαi2 and ELMO1/Dock180 connects chemokine signalling with Rac activation and metastasis. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Côté, J.F.; Vuori, K. In Vitro Guanine Nucleotide Exchange Activity of DHR-2/DOCKER/CZH2 Domains. Methods Enzymol. 2006, 406, 41–57. [Google Scholar] [PubMed]
- Nishihara, H.; Maeda, M.; Oda, A.; Tsuda, M.; Sawa, H.; Nagashima, K.; Tanaka, S. DOCK2 associates with CrkL and regulates Rac1 in human leukemia cell lines. Blood 2002, 100, 3968–3974. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, H.; Maeda, M.; Tsuda, M.; Makino, Y.; Sawa, H.; Nagashima, K.; Tanaka, S. DOCK2 mediates T cell receptor-induced activation of Rac2 and IL-2 transcription. Biochem. Biophys. Res. Commun. 2002, 296, 716–720. [Google Scholar] [CrossRef]
- Namekata, K.; Enokido, Y.; Iwasawa, K.; Kimura, H. MOCA induces membrane spreading by activating Rac1. J. Biol. Chem. 2004, 279, 14331–14337. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Kinchen, J.M.; Rossman, K.L.; Grimsley, C.; Hall, M.; Sondek, J.; Hengartner, M.O.; Yajnik, V.; Ravichandran, K.S. A Steric-Inhibition Model for Regulation of Nucleotide Exchange via the Dock180 Family of GEFs. Curr. Biol. 2005, 15, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Omi, N.; Kiyokawa, E.; Matsuda, M.; Kinoshita, K.; Yamada, S.; Yamada, K.; Matsushima, Y.; Wang, Y.; Kawai, J.; Suzuki, M.; et al. Mutation of Dock5, a member of the guanine exchange factor Dock180 superfamily, in the rupture of lens cataract mouse. Exp. Eye Res. 2008, 86, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Yamauchi, J.; Sanbe, A.; Tanoue, A. Dock6, a Dock-C subfamily guanine nucleotide exchanger, has the dual specificity for Rac1 and Cdc42 and regulates neurite outgrowth. Exp. Cell Res. 2007, 313, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Watabe-Uchida, M.; John, K.A.; Janas, J.A.; Newey, S.E.; Van Aelst, L. The Rac activator DOCK7 regulates neuronal polarity through local phosphorylation of stathmin/Op18. Neuron 2006, 51, 727–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Johnson, J.L.; Cerione, R.A.; Erickson, J.W. Prenylation and membrane localization of Cdc42 are essential for activation by DOCK7. Biochemistry 2013, 52, 4354–4363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, Y.; Tanaka, Y.; Terasawa, M.; Pieczyk, M.; Habiro, K.; Katakai, T.; Hanawa-Suetsugu, K.; Kukimoto-Niino, M.; Nishizaki, T.; Shirouzu, M.; et al. DOCK8 is a Cdc42 activator critical for interstitial dendritic cell migration during immune responses. Blood 2012, 119, 4451–4461. [Google Scholar] [CrossRef] [Green Version]
- Meller, N.; Irani-Tehrani, M.; Kiosses, W.B.; Del Pozo, M.A.; Schwartz, M.A. Zizimin1, a novel Cdc42 activator, reveals a new GEF domain for Rho proteins. Nat. Cell Biol. 2002, 4, 639–647. [Google Scholar] [CrossRef]
- Gadea, G.; Sanz-Moreno, V.; Self, A.; Godi, A.; Marshall, C.J. DOCK10-mediated Cdc42 activation is necessary for amoeboid invasion of melanoma cells. Curr. Biol. 2008, 18, 1456–1465. [Google Scholar] [CrossRef] [Green Version]
- Nishikimi, A.; Meller, N.; Uekawa, N.; Isobe, K.; Schwartz, M.A.; Maruyama, M. Zizimin2: A novel, DOCK180-related Cdc42 guanine nucleotide exchange factor expressed predominantly in lymphocytes. FEBS Lett. 2005, 579, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Leonard, D.; Hart, M.J.; Platko, J.V.; Eva, A.; Henzel, W.; Evans, T.; Cerione, R.A. The Identification and Characterization of a GDP-dissociation Inhibitor (GDI) for the CDC42Hs Protein. J. Biol. Chem. 1992, 267, 22860–22868. [Google Scholar]
- Gorvel, J.P.; Chang, T.C.; Boretto, J.; Azuma, T.; Chavrier, P. Differential properties of D4/LyGDI versus RhoGDI: Phosphorylation and rho GTPase selectivity. FEBS Lett. 1998, 422, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Fauré, J.; Dagher, M.C. Interactions between Rho GTPases and Rho GDP dissociation inhibitor (Rho-GDI). Biochimie 2001, 83, 409–414. [Google Scholar] [CrossRef]
- Fukumoto, Y.; Kaibuchi, K.; Hori, Y.; Fujioka, H.; Araki, S.; Ueda, T.; Kikuchi, A.; Takai, Y. Molecular cloning and characterization of a novel type of regulatory protein (GDI) for the rho proteins, ras p21-like small GTP-binding proteins. Oncogene 1990, 5, 1321–1328. [Google Scholar] [PubMed]
- Elfenbein, A.; Rhodes, J.M.; Meller, J.; Schwartz, M.A.; Matsuda, M.; Simons, M. Suppression of RhoG activity is mediated by a syndecan 4-synectin-RhoGDI1 complex and is reversed by PKCalpha in a Rac1 activation pathway. J. Cell Biol. 2009, 186, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Urano, E.; Miyauchi, K.; Ichikawa, R.; Hamatake, M.; Misawa, N.; Sato, K.; Ebina, H.; Koyanagi, Y.; Komano, J. The hematopoietic cell-specific Rho GTPase inhibitor ARHGDIB/D4GDI limits HIV type 1 replication. AIDS Res. Hum. Retrovir. 2012, 28, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platko, J.V.; Leonard, D.A.; Adra, C.N.; Shaw, R.J.; Cerione, R.A.; Lim, B. A single residue can modify target-binding affinity and activity of the functional domain of the Rho-subfamily GDP dissociation inhibitors. Proc. Natl. Acad. Sci. USA 1995, 92, 2974–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, N.; Morin, A.; Olofsson, B. RhoGDI-3 regulates RhoG and targets this protein to the Golgi complex through its unique N-terminal domain. Traffic 2002, 3, 341–357. [Google Scholar] [CrossRef]
- Zalcman, G.; Closson, V.; Camonis, J.; Honoré, N.; Rousseau-Merck, M.F.; Tavitian, A.; Olofsson, B. RhoGDI-3 is a new GDP dissociation inhibitor (GDI). Identification of a non-cytosolic GDI protein interacting with the small GTP-binding proteins RhoB and RhoG. J. Biol. Chem. 1996, 271, 30366–30374. [Google Scholar] [CrossRef] [Green Version]
- Adra, C.N.; Manor, D.; Ko, J.L.; Zhu, S.; Horiuchi, T.; Van Aelst, L.; Cerione, R.A.; Lim, B. RhoGDIg: A GDP-dissociation inhibitor for Rho proteins with preferential expression in brain and pancreas. Proc. Natl. Acad. Sci. USA 1997, 94, 4279–4284. [Google Scholar] [CrossRef] [Green Version]
- Rittinger, K.; Taylor, W.R.; Smerdon, S.J.; Gamblin, S.J. Support for shared ancestry of GAPs. Nature 1998, 392. [Google Scholar] [CrossRef]
- Bax, B. Domains of rasGAP and rhoGAP are related. Nature 1998, 392, 447–448. [Google Scholar] [CrossRef]
- Ahmadian, M.R.; Stege, P.; Scheffzek, K.; Wittinghofer, A. Confirmation of the arginine-finger hypothesis for the GAP-stimulated GTP-hydrolysis reaction of Ras. Nat. Struct. Biol. 1997, 4, 686–689. [Google Scholar] [CrossRef]
- Mittal, R.; Ahmadian, M.R.; Goody, R.S.; Wittinghofer, A. Formation of a transition-state analog of the Ras GTPase reaction by Ras-GDP, tetrafluoroaluminate, and GTPase-activating proteins. Science 1996, 273, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.Y.; Zheng, Y. Rho GTPase-activating proteins in cell regulation. Trends Cell Biol. 2003, 13, 13–22. [Google Scholar] [CrossRef]
- Canagarajah, B.; Leskow, F.C.; Ho, J.Y.; Mischak, H.; Saidi, L.F.; Kazanietz, M.G.; Hurley, J.H. Structural mechanism for lipid activation of the Rac-specific GAP, beta2-chimaerin. Cell 2004, 119, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Bao, H.; Li, F.; Wang, C.; Wang, N.; Jiang, Y.; Tang, Y.; Wu, J.; Shi, Y. Structural Basis for the Specific Recognition of RhoA by the Dual GTPase-activating Protein ARAP3. J. Biol. Chem. 2016, 291, 16709–16719. [Google Scholar] [CrossRef] [Green Version]
- Barrio-Real, L.; Kazanietz, M.G. Rho GEFs and Cancer: Linking Gene Expression and Metastatic Dissemination. Sci. Signal. 2012, 5, pe43. [Google Scholar] [CrossRef]
- Garcia-Mata, R.; Burridge, K. Catching a GEF by its tail. Trends Cell Biol. 2007, 17, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Schiller, M.R. Coupling receptor tyrosine kinases to Rho GTPases—GEFs what’s the link. Cell. Signal. 2006, 18, 1834–1843. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Radulovich, N.; Wang, T.; Raghavan, V.; Zhu, C.Q.; Tsao, M.S. Rho guanine nucleotide exchange factor ARHGEF10 is a putative tumor suppressor in pancreatic ductal adenocarcinoma. Oncogene 2020, 39, 308–321. [Google Scholar] [CrossRef]
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: Turning on RHO GTPases with guanine nucleotide-exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef]
- Cook, D.R.; Rossman, K.L.; Der, C.J. Rho guanine nucleotide exchange factors: Regulators of Rho GTPase activity in development and disease. Oncogene 2014, 33, 4021–4035. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Li, H.; Yang, Z. Arabidopsis RopGAPs are a novel family of rho GTPase-activating proteins that require the Cdc42/Rac-interactive binding motif for rop-specific GTPase stimulation. Plant Physiol. 2000, 124, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Cote, J.F.; Vuori, K. Identification of an evolutionarily conserved superfamily of DOCK180-related proteins with guanine nucleotide exchange activity. J. Cell Sci. 2002, 115, 4901–4913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meller, N.; Merlot, S.; Guda, C. CZH proteins: A new family of Rho-GEFs. J. Cell Sci. 2005, 118, 4937–4946. [Google Scholar] [CrossRef] [Green Version]
- DerMardirossian, C.; Bokoch, G.M. GDIs: Central regulatory molecules in Rho GTPase activation. Trends Cell Biol. 2005, 15, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Dransart, E.; Olofsson, B.; Cherfils, J. RhoGDIs revisited: Novel roles in Rho regulation. Traffic 2005, 6, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Kikuchi, A.; Ohga, N.; Yamamoto, J.; Takai, Y. Purification and Characterization from Bovine Brain Cytosol of a Novel Regulatory Protein Inhibiting the Dissociation of GDP from and the Subsequent Binding of GTP to rhoB ~20, a ras p214ike GTPbinding Protein. J. Biol. Chem. 1990, 265, 9373–9380. [Google Scholar] [PubMed]
- Lelias, J.M.; Adra, C.N.; Wulf, G.M.; Guillemot, J.C.; Khagad, M.; Caput, D.; Lim, B. cDNA cloning of a human mRNA preferentially expressed in hematopoietic cells and with homology to a GDP-dissociation inhibitor for the rho GTP-binding proteins. Proc. Natl. Acad. Sci. USA 1993, 90, 1479–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherle, P.; Behrens, T.; Staudt, L.M. Ly-GDI, a GDP-dissociation inhibitor of the RhoA GTP-binding protein, is expressed preferentially in lymphocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 7568–7572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, M.A.; Theodorescu, D. RhoGDI2: A new metastasis suppressor gene: Discovery and clinical translation. Urol. Oncol. 2007, 25, 401–406. [Google Scholar] [CrossRef]
- Abiatari, I.; DeOliveira, T.; Kerkadze, V.; Schwager, C.; Esposito, I.; Giese, N.A.; Huber, P.; Bergman, F.; Abdollahi, A.; Friess, H.; et al. Consensus transcriptome signature of perineural invasion in pancreatic carcinoma. Mol. Cancer Ther. 2009, 8, 1494–1504. [Google Scholar] [CrossRef] [Green Version]
- Seraj, M.J.; Harding, M.A.; Gildea, J.J.; Welch, D.R.; Theodorescu, D. The relationship of BRMS1 and RhoGDI2 gene expression to metastatic potential in lineage related human bladder cancer cell lines. Clin. Exp. Metastasis 2001, 18, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xu, G.; Sotnikova, A.; Szczepanowski, M.; Giefing, M.; Krause, K.; Krams, M.; Siebert, R.; Jin, J.; Klapper, W. Loss of expression of LyGDI (ARHGDIB), a rho GDP-dissociation inhibitor, in Hodgkin lymphoma. Br. J. Haematol. 2007, 139, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Adra, C.N.; Ko, J.; Leonard, D.; Wirth, L.J.; Cerione, R.A.; Lim, B. Identification of a Novel Protein With GDP Dissociation Inhibitor Activity for the Ras-Like Proteins CDC42Hs and Rac I. Genes Chromosomes Cancer 1993, 8, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Harding, M.A.; Theodorescu, D. RhoGDI signaling provides targets for cancer therapy. Eur. J. Cancer 2010, 46, 1252–1259. [Google Scholar] [CrossRef]
- Xie, F.; Shao, S.; Aziz, A.U.R.; Zhang, B.; Wang, H.; Liu, B. Role of Rho-specific guanine nucleotide dissociation inhibitor alpha regulation in cell migration. Acta Histochem 2017, 119, 183–189. [Google Scholar] [CrossRef]
- Lin, X.; Yang, B.; Liu, W.; Tan, X.; Wu, F.; Hu, P.; Jiang, T.; Bao, Z.; Yuan, J.; Qiang, B.; et al. Interplay between PCBP2 and miRNA modulates ARHGDIA expression and function in glioma migration and invasion. Oncotarget 2016, 7, 19483–19498. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Bhandari, D.; Leyme, A.; Aznar, N.; Midde, K.K.; Lo, I.C.; Ear, J.; Niesman, I.; Lopez-Sanchez, I.; Blanco-Canosa, J.B.; et al. GIV/Girdin activates Galphai and inhibits Galphas via the same motif. Proc. Natl. Acad. Sci. USA 2016, 113, E5721–E5730. [Google Scholar] [CrossRef] [Green Version]
- Enomoto, A.; Murakami, H.; Asai, N.; Morone, N.; Watanabe, T.; Kawai, K.; Murakumo, Y.; Usukura, J.; Kaibuchi, K.; Takahashi, M. Akt/PKB regulates actin organization and cell motility via Girdin/APE. Dev. Cell 2005, 9, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Le-Niculescu, H.; Niesman, I.; Fischer, T.; DeVries, L.; Farquhar, M.G. Identification and characterization of GIV, a novel Galpha i/s-interacting protein found on COPI, endoplasmic reticulum-Golgi transport vesicles. J. Biol. Chem. 2005, 280, 22012–22020. [Google Scholar] [CrossRef] [Green Version]
- Simpson, F.; Martin, S.; Evans, T.M.; Kerr, M.; James, D.E.; Parton, R.G.; Teasdale, R.D.; Wicking, C. A novel hook-related protein family and the characterization of hook-related protein 1. Traffic 2005, 6, 442–458. [Google Scholar] [CrossRef]
- Anai, M.; Shojima, N.; Katagiri, H.; Ogihara, T.; Sakoda, H.; Onishi, Y.; Ono, H.; Fujishiro, M.; Fukushima, Y.; Horike, N.; et al. A novel protein kinase B (PKB)/AKT-binding protein enhances PKB kinase activity and regulates DNA synthesis. J. Biol. Chem. 2005, 280, 18525–18535. [Google Scholar] [CrossRef] [Green Version]
- Ge, P.; Cao, L.; Chen, X.; Jing, R.; Yue, W. miR-762 activation confers acquired resistance to gefitinib in non-small cell lung cancer. BMC Cancer 2019, 19, 1203. [Google Scholar] [CrossRef] [PubMed]
- Ouchida, M.; Kanzaki, H.; Ito, S.; Hanafusa, H.; Jitsumori, Y.; Tamaru, S.; Shimizu, K. Novel direct targets of miR-19a identified in breast cancer cells by a quantitative proteomic approach. PLoS ONE 2012, 7, e44095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, Y.H.; Gibbons, D.L.; Chakravarti, D.; Creighton, C.J.; Rizvi, Z.H.; Adams, H.P.; Pertsemlidis, A.; Gregory, P.A.; Wright, J.A.; Goodall, G.J.; et al. ZEB1 drives prometastatic actin cytoskeletal remodeling by downregulating miR-34a expression. J. Clin. Investig. 2012, 122, 3170–3183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, S.L.; Palat, A.; Pan, Y.; Rajapakshe, K.; Mirchandani, R.; Bondesson, M.; Yustein, J.T.; Coarfa, C.; Gunaratne, P.H. MicroRNA-509-3p inhibits cellular migration, invasion, and proliferation, and sensitizes osteosarcoma to cisplatin. Sci. Rep. 2019, 9, 19089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satterfield, L.; Shuck, R.; Kurenbekova, L.; Allen-Rhoades, W.; Edwards, D.; Huang, S.; Rajapakshe, K.; Coarfa, C.; Donehower, L.A.; Yustein, J.T. miR-130b directly targets ARHGAP1 to drive activation of a metastatic CDC42-PAK1-AP1 positive feedback loop in Ewing sarcoma. Int. J. Cancer 2017, 141, 2062–2075. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Ochi, H.; Sunamura, S.; Kosaka, N.; Mabuchi, Y.; Fukuda, T.; Yao, K.; Kanda, H.; Ae, K.; Okawa, A.; et al. Cancer-secreted hsa-miR-940 induces an osteoblastic phenotype in the bone metastatic microenvironment via targeting ARHGAP1 and FAM134A. Proc. Natl. Acad. Sci. USA 2018, 115, 2204–2209. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tian, X.; Han, R.; Zhang, X.; Wang, X.; Shen, H.; Xue, L.; Liu, Y.; Yan, X.; Shen, J.; et al. Downregulation of miR-486-5p contributes to tumor progression and metastasis by targeting protumorigenic ARHGAP5 in lung cancer. Oncogene 2013, 33, 1181–1189. [Google Scholar] [CrossRef] [Green Version]
- Kwak, S.Y.; Yang, J.S.; Kim, B.Y.; Bae, I.H.; Han, Y.H. Ionizing radiation-inducible miR-494 promotes glioma cell invasion through EGFR stabilization by targeting p190B rhoGAP. Biochim. Biophys. Acta 2014, 1843, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Jia, L.; Yu, Y.; Jin, L. Lactic acid induced microRNA-744 enhances motility of SiHa cervical cancer cells through targeting ARHGAP5. Chem. Biol. Interact. 2019, 298, 86–95. [Google Scholar] [CrossRef]
- Fang, Y.; Zhu, X.; Wang, J.; Li, N.; Li, D.; Sakib, N.; Sha, Z.; Song, W. MiR-744 functions as a proto-oncogene in nasopharyngeal carcinoma progression and metastasis via transcriptional control of ARHGAP5. Oncotarget 2015, 6, 13164–13175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, S.; Xiao, R.; Shi, L.; Zhou, X.; Yang, T.; Zhang, M.; Weng, N.; Zhao, X.; Wang, R.; Liu, J.; et al. MicroRNA-18a promotes cancer progression through SMG1 suppression and mTOR pathway activation in nasopharyngeal carcinoma. Cell Death Dis. 2019, 10, 819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.J.; Li, J.S.; Zhou, H.; Xiao, H.X.; Li, Y.; Zhou, T. MicroRNA-106b promotes colorectal cancer cell migration and invasion by directly targeting DLC1. J. Exp. Clin. Cancer Res. 2015, 34, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.P.; Zhu, H.Y.; Sun, X.F.; Chen, L.X.; Zhou, Q.; Chen, J. MicroRNA-141 regulates the tumour suppressor DLC1 in colorectal cancer. Neoplasma 2015, 62, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Li, Y.; Zhang, G. Downregulation of microRNA-301a inhibited proliferation, migration and invasion of non-small cell lung cancer by directly targeting DLC1. Oncol. Lett. 2017, 14, 6017–6023. [Google Scholar] [CrossRef] [Green Version]
- Xiao, P.; Liu, W.; Zhou, H. miR-429 promotes the proliferation of non-small cell lung cancer cells via targeting DLC-1. Oncol. Lett. 2016, 12, 2163–2168. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Liu, Y.; Jiang, J.; Liu, Y.; Yang, Z.; Wu, S.; Cao, W.; Cui, I.H.; Yu, C. IGF2-derived miR-483 mediated oncofunction by suppressing DLC-1 and associated with colorectal cancer. Oncotarget 2016, 7, 48456–48466. [Google Scholar] [CrossRef] [Green Version]
- Pacurari, M.; Addison, J.B.; Bondalapati, N.; Wan, Y.W.; Luo, D.; Qian, Y.; Castranova, V.; Ivanov, A.V.; Guo, N.L. The microRNA-200 family targets multiple non-small cell lung cancer prognostic markers in H1299 cells and BEAS-2B cells. Int. J. Oncol. 2013, 43, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, F.F.; Jamal, R.; Syafruddin, S.E.; Ab Mutalib, N.S.; Saidin, S.; MdZin, R.R.; Hossain Mollah, M.M.; Mokhtar, N.M. MicroRNA-200c and microRNA-31 regulate proliferation, colony formation, migration and invasion in serous ovarian cancer. J. Ovarian Res. 2015, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Scisciani, C.; Vossio, S.; Guerrieri, F.; Schinzari, V.; De Iaco, R.; D’Onorio de Meo, P.; Cervello, M.; Montalto, G.; Pollicino, T.; Raimondo, G.; et al. Transcriptional regulation of miR-224 upregulated in human HCCs by NFkappaB inflammatory pathways. J. Hepatol. 2012, 56, 855–861. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, L.; Li, Y.; Hao, B.; Wang, M.; Wang, J.; Gu, W.; Zhan, H.; Liu, G.; Wu, Q. miR3373p inhibits gastric tumor metastasis by targeting ARHGAP10. Mol. Med. Rep. 2020, 21, 705–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Wang, L.; Li, Z.; Wang, W.; Zhi, X.; Huang, X.; Zhang, Q.; Chen, Z.; Zhang, X.; He, Z.; et al. miR-3174 Contributes to Apoptosis and Autophagic Cell Death Defects in Gastric Cancer Cells by Targeting ARHGAP10. Mol. Ther. Nucleic Acids 2017, 9, 294–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanzaki, H.; Ito, S.; Hanafusa, H.; Jitsumori, Y.; Tamaru, S.; Shimizu, K.; Ouchida, M. Identification of direct targets for the miR-17-92 cluster by proteomic analysis. Proteomics 2011, 11, 3531–3539. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhou, Y.; Zhang, J.; Wong, C.C.; Li, W.; Kwan, J.S.H.; Yang, R.; Chan, A.K.Y.; Dong, Y.; Wu, F.; et al. SRGAP1, a crucial target of miR-340 and miR-124, functions as a potential oncogene in gastric tumorigenesis. Oncogene 2017, 37, 1159–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, S.; Martin, G.; Toussaint, L.G. MicroRNA-145 Promotes the Phenotype of Human Glioblastoma Cells Selected for Invasion. Anticancer Res. 2015, 35, 3209–3215. [Google Scholar]
- Chen, J.; Huang, X.; Wang, W.; Xie, H.; Li, J.; Hu, Z.; Zheng, Z.; Li, H.; Teng, L. LncRNA CDKN2BAS predicts poor prognosis in patients with hepatocellular carcinoma and promotes metastasis via the miR-153-5p/ARHGAP18 signaling axis. Aging 2018, 10, 3371–3381. [Google Scholar] [CrossRef]
- Howe, E.N.; Cochrane, D.R.; Richer, J.K. Targets of miR-200c mediate suppression of cell motility and anoikis resistance. Breast Cancer Res. 2011, 13, R45. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wei, W.Q.; Wu, Z.Y.; Wang, G.C. MicroRNA-590-5p regulates cell viability, apoptosis, migration and invasion of renal cell carcinoma cell lines through targeting ARHGAP24. Mol. Biosyst. 2017, 13, 2564–2573. [Google Scholar] [CrossRef]
- Wang, Q.; Hui, H.; Guo, Z.; Zhang, W.; Hu, Y.; He, T.; Tai, Y.; Peng, P.; Wang, L. ADAR1 regulates ARHGAP26 gene expression through RNA editing by disrupting miR-30b-3p and miR-573 binding. RNA 2013, 19, 1525–1536. [Google Scholar] [CrossRef] [Green Version]
- Bracken, C.P.; Li, X.; Wright, J.A.; Lawrence, D.M.; Pillman, K.A.; Salmanidis, M.; Anderson, M.A.; Dredge, B.K.; Gregory, P.A.; Tsykin, A.; et al. Genome-wide identification of miR-200 targets reveals a regulatory network controlling cell invasion. EMBO J. 2014, 33, 2040–2056. [Google Scholar] [CrossRef] [Green Version]
- D’Ippolito, E.; Plantamura, I.; Bongiovanni, L.; Casalini, P.; Baroni, S.; Piovan, C.; Orlandi, R.; Gualeni, A.V.; Gloghini, A.; Rossini, A.; et al. miR-9 and miR-200 Regulate PDGFRbeta-Mediated Endothelial Differentiation of Tumor Cells in Triple-Negative Breast Cancer. Cancer Res. 2016, 76, 5562–5572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Hu, W.; Li, G.; Guo, Y.; Wan, Z.; Yu, J. Inhibition of miR-9-5p suppresses prostate cancer progress by targeting StarD13. Cell. Mol. Biol. Lett. 2019, 24, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Zhang, R.; He, Y.; Zou, M.; Guo, L.; Xi, T. MicroRNA-125b induces metastasis by targeting STARD13 in MCF-7 and MDA-MB-231 breast cancer cells. PLoS ONE 2012, 7, e35435. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; He, S.; Qiu, G.; Lu, J.; Wang, J.; Liu, J.; Fan, L.; Zhao, W.; Che, X. MicroRNA-125b promotes invasion and metastasis of gastric cancer by targeting STARD13 and NEU1. Tumour Biol. 2016, 37, 12141–12151. [Google Scholar] [CrossRef] [PubMed]
- Xishan, Z.; Xianjun, L.; Ziying, L.; Guangxin, C.; Gang, L. The malignancy suppression role of miR-23a by targeting the BCR/ABL oncogene in chromic myeloid leukemia. Cancer Gene Ther. 2014, 21, 397–404. [Google Scholar] [CrossRef]
- Xishan, Z.; Ziying, L.; Jing, D.; Gang, L. MicroRNA-320a acts as a tumor suppressor by targeting BCR/ABL oncogene in chronic myeloid leukemia. Sci. Rep. 2015, 5, 12460. [Google Scholar] [CrossRef]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef]
- Furuta, M.; Kozaki, K.I.; Tanaka, S.; Arii, S.; Imoto, I.; Inazawa, J. miR-124 and miR-203 are epigenetically silenced tumor-suppressive microRNAs in hepatocellular carcinoma. Carcinogenesis 2010, 31, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Fowler, A.; Thomson, D.; Giles, K.; Maleki, S.; Mreich, E.; Wheeler, H.; Leedman, P.; Biggs, M.; Cook, R.; Little, N.; et al. miR-124a is frequently down-regulated in glioblastoma and is involved in migration and invasion. Eur. J. Cancer 2011, 47, 953–963. [Google Scholar] [CrossRef]
- Dong, P.; Ihira, K.; Xiong, Y.; Watari, H.; Hanley, S.J.; Yamada, T.; Hosaka, M.; Kudo, M.; Yue, J.; Sakuragi, N. Reactivation of epigenetically silenced miR-124 reverses the epithelial-to-mesenchymal transition and inhibits invasion in endometrial cancer cells via the direct repression of IQGAP1 expression. Oncotarget 2016, 7, 20260–20270. [Google Scholar] [CrossRef]
- Sun, G.; Liu, Y.; Wang, K.; Xu, Z. miR-506 regulates breast cancer cell metastasis by targeting IQGAP1. Int. J. Oncol. 2015, 47, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Pelossof, R.; Chow, O.S.; Fairchild, L.; Smith, J.J.; Setty, M.; Chen, C.T.; Chen, Z.; Egawa, F.; Avila, K.; Leslie, C.S.; et al. Integrated genomic profiling identifies microRNA-92a regulation of IQGAP2 in locally advanced rectal cancer. Genes Chromosomes Cancer 2016, 55, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Toste, P.A.; Li, L.; Kadera, B.E.; Nguyen, A.H.; Tran, L.M.; Wu, N.; Madnick, D.L.; Patel, S.G.; Dawson, D.W.; Donahue, T.R. p85alpha is a microRNA target and affects chemosensitivity in pancreatic cancer. J. Surg. Res. 2015, 196, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.X.; Liu, Y.H.; Xiang, J.W.; Wu, Q.N.; Xu, L.B.; Luo, X.L.; Zhu, X.L.; Liu, C.; Xu, F.P.; Luo, D.L.; et al. PIK3R1 targeting by miR-21 suppresses tumor cell migration and invasion by reducing PI3K/AKT signaling and reversing EMT, and predicts clinical outcome of breast cancer. Int. J. Oncol. 2016, 48, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Shen, Y.; Liu, M.; Bi, C.; Jiang, C.; Iqbal, J.; McKeithan, T.W.; Chan, W.C.; Ding, S.J.; Fu, K. Quantitative proteomics reveals that miR-155 regulates the PI3K-AKT pathway in diffuse large B-cell lymphoma. Am. J. Pathol. 2012, 181, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, R.M.; Xiao, S.; Lei, X.; Yang, H.; Fang, F.; Yang, L.Y. miRNA-487a Promotes Proliferation and Metastasis in Hepatocellular Carcinoma. Clin. Cancer Res. 2017, 23, 2593–2604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.K.; Verma, G.; Vig, S.; Srivastava, S.; Srivastava, A.K.; Datta, M. miR-29a levels are elevated in the db/db mice liver and its overexpression leads to attenuation of insulin action on PEPCK gene expression in HepG2 cells. Mol. Cell. Endocrinol. 2011, 332, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, J.H.; Ha, M.; Nam, J.W.; Kim, V.N. miR-29 miRNAs activate p53 by targeting p85 alpha and CDC42. Nat. Struct. Mol. Biol. 2009, 16, 23–29. [Google Scholar] [CrossRef]
- Okamoto, K.; Miyoshi, K.; Murawaki, Y. miR-29b, miR-205 and miR-221 enhance chemosensitivity to gemcitabine in HuH28 human cholangiocarcinoma cells. PLoS ONE 2013, 8, e77623. [Google Scholar] [CrossRef]
- Jiang, B.-H.; Chen, P.-H.; Cheng, C.-H.; Shih, C.-M.; Ho, K.-H.; Lin, C.-W.; Lee, C.-C.; Liu, A.-J.; Chang, C.-K.; Chen, K.-C. The Inhibition of microRNA-128 on IGF-1-Activating mTOR Signaling Involves in Temozolomide-Induced Glioma Cell Apoptotic Death. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, H.; Tang, H.; Fang, Z.; Wang, J.; Cui, S. miR-218 inhibits the invasion and migration of colon cancer cells by targeting the PI3K/Akt/mTOR signaling pathway. Int. J. Mol. Med. 2015, 35, 1301–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Yin, L.; Chen, H.; Yang, S.; Pan, C.; Lu, S.; Miao, M.; Jiao, B. miR-376a suppresses proliferation and induces apoptosis in hepatocellular carcinoma. FEBS Lett. 2012, 586, 2396–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.P.; Hou, J.; Shen, X.Y.; Huang, C.Y.; Zhang, X.H.; Xie, Y.A.; Luo, X.L. MicroRNA-486-5p, which is downregulated in hepatocellular carcinoma, suppresses tumor growth by targeting PIK3R1. FEBS J. 2015, 282, 579–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.; Wang, J.; Ouyang, T.; Lu, N.; Lu, J.; Shen, Y.; Bai, Y.; Xie, X.; Ge, Q. MiR-486-5p Serves as a Good Biomarker in Nonsmall Cell Lung Cancer and Suppresses Cell Growth with the Involvement of a Target PIK3R1. Front. Genet. 2019, 10, 688. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, L.; Zhang, Y.; Guan, H.; Wu, J.; Zhu, X.; Yuan, J.; Li, M. MiR-503 targets PI3K p85 and IKK-beta and suppresses progression of non-small cell lung cancer. Int. J. Cancer 2014, 135, 1531–1542. [Google Scholar] [CrossRef]
- Cai, J.; Zhao, J.; Zhang, N.; Xu, X.; Li, R.; Yi, Y.; Fang, L.; Zhang, L.; Li, M.; Wu, J.; et al. MicroRNA-542-3p Suppresses Tumor Cell Invasion via Targeting AKT Pathway in Human Astrocytoma. J. Biol. Chem. 2015, 290, 24678–24688. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Wang, F.; Wang, L.; Wong, S.C.; Cho, W.C.; Chan, L.W. MiR-30a-5p Overexpression May Overcome EGFR-Inhibitor Resistance through Regulating PI3K/AKT Signaling Pathway in Non-small Cell Lung Cancer Cell Lines. Front. Genet. 2016, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, D.; Xie, H.; Zhou, Z.; Chen, H.; Hu, T.; Bai, Y.; Shen, Y.; Yuan, W.; Jing, Q.; et al. Endothelial-specific intron-derived miR-126 is down-regulated in human breast cancer and targets both VEGFA and PIK3R2. Mol. Cell. Biochem. 2011, 351, 157–164. [Google Scholar] [CrossRef]
- Guo, C.; Sah, J.F.; Beard, L.; Willson, J.K.; Markowitz, S.D.; Guda, K. The noncoding RNA, miR-126, suppresses the growth of neoplastic cells by targeting phosphatidylinositol 3-kinase signaling and is frequently lost in colon cancers. Genes Chromosomes Cancer 2008, 47, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Nie, Z.C.; Weng, W.H.; Shang, Y.S.; Long, Y.; Li, J.; Xu, Y.T.; Li, Z. MicroRNA-126 is down-regulated in human esophageal squamous cell carcinoma and inhibits the proliferation and migration in EC109 cell via PI3K/AKT signaling pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 4745–4754. [Google Scholar]
- Liu, L.Y.; Wang, W.; Zhao, L.Y.; Guo, B.; Yang, J.; Zhao, X.G.; Hou, N.; Ni, L.; Wang, A.Y.; Song, T.S.; et al. Mir-126 inhibits growth of SGC-7901 cells by synergistically targeting the oncogenes PI3KR2 and Crk, and the tumor suppressor PLK2. Int. J. Oncol. 2014, 45, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.J.; Zhao, Z.F.; Kang, X.J.; Wang, H.J.; Zhao, J.; Pu, X.M. MicroRNA-126-3p suppresses cell proliferation by targeting PIK3R2 in Kaposi’s sarcoma cells. Oncotarget 2016, 7, 36614–36621. [Google Scholar]
- Choi, Y.C.; Yoon, S.; Byun, Y.; Lee, G.; Kee, H.; Jeong, Y.; Yoon, J.; Baek, K. MicroRNA library screening identifies growth-suppressive microRNAs that regulate genes involved in cell cycle progression and apoptosis. Exp. Cell Res. 2015, 339, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Georges, S.A.; Biery, M.C.; Kim, S.Y.; Schelter, J.M.; Guo, J.; Chang, A.N.; Jackson, A.L.; Carleton, M.O.; Linsley, P.S.; Cleary, M.A.; et al. Coordinated regulation of cell cycle transcripts by p53-Inducible microRNAs, miR-192 and miR-215. Cancer Res. 2008, 68, 10105–10112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalid, M.; Idichi, T.; Seki, N.; Wada, M.; Yamada, Y.; Fukuhisa, H.; Toda, H.; Kita, Y.; Kawasaki, Y.; Tanoue, K.; et al. Gene Regulation by Antitumor miR-204-5p in Pancreatic Ductal Adenocarcinoma: The Clinical Significance of Direct RACGAP1 Regulation. Cancers 2019, 11, 327. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.; Lu, M.; Ju, L.; Qian, K.; Wang, G.; Wu, C.L.; Liu, X.; Xiao, Y.; Wang, X. miR-4324-RACGAP1-STAT3-ESR1 feedback loop inhibits proliferation and metastasis of bladder cancer. Int. J. Cancer 2019, 144, 3043–3055. [Google Scholar] [CrossRef]
- Hu, Y.; Ou, Y.; Wu, K.; Chen, Y.; Sun, W. miR-143 inhibits the metastasis of pancreatic cancer and an associated signaling pathway. Tumor Biol. 2012, 33, 1863–1870. [Google Scholar] [CrossRef]
- Guo, B.; Hui, Q.; Zhang, Y.; Chang, P.; Tao, K. miR-194 is a negative regulator of GEF-H1 pathway in melanoma. Oncol. Rep. 2016, 36, 2412–2420. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Liu, X.; Kolokythas, A.; Yu, J.; Wang, A.; Heidbreder, C.E.; Shi, F.; Zhou, X. Downregulation of the Rho GTPase signaling pathway is involved in the microRNA-138-mediated inhibition of cell migration and invasion in tongue squamous cell carcinoma. Int. J. Cancer 2010, 127, 505–512. [Google Scholar] [CrossRef]
- Pellegrino, L.; Stebbing, J.; Braga, V.M.; Frampton, A.E.; Jacob, J.; Buluwela, L.; Jiao, L.R.; Periyasamy, M.; Madsen, C.D.; Caley, M.P.; et al. miR-23b regulates cytoskeletal remodeling, motility and metastasis by directly targeting multiple transcripts. Nucleic Acids Res. 2013, 41, 5400–5412. [Google Scholar] [CrossRef]
- Hemmesi, K.; Squadrito, M.L.; Mestdagh, P.; Conti, V.; Cominelli, M.; Piras, I.S.; Sergi, L.S.; Piccinin, S.; Maestro, R.; Poliani, P.L.; et al. miR-135a Inhibits Cancer Stem Cell-Driven Medulloblastoma Development by Directly Repressing Arhgef6 Expression. Stem Cells 2015, 33, 1377–1389. [Google Scholar] [CrossRef]
- Xiao, J.; Niu, S.; Zhu, J.; Lv, L.; Deng, H.; Pan, D.; Shen, D.; Xu, C.; Shen, Z.; Tao, T. miR223p enhances multichemoresistance by targeting NET1 in bladder cancer cells. Oncol. Rep. 2018, 39, 2731–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadimitriou, E.; Vasilaki, E.; Vorvis, C.; Iliopoulos, D.; Moustakas, A.; Kardassis, D.; Stournaras, C. Differential regulation of the two RhoA-specific GEF isoforms Net1/Net1A by TGF-beta and miR-24: Role in epithelial-to-mesenchymal transition. Oncogene 2012, 31, 2862–2875. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.M.; Muiwo, P.; Ramachandran, S.S.; Pandey, P.; Gupta, Y.K.; Kumar, L.; Kulshreshtha, R.; Bhattacharya, A. miR-22 regulates expression of oncogenic neuro-epithelial transforming gene 1, NET1. FEBS J. 2014, 281, 3904–3919. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, Z.; Luo, W.; Liu, J.; Lou, Y.; Xia, S. NET1 Enhances Proliferation and Chemoresistance in Acute Lymphoblastic Leukemia Cells. Oncol. Res. 2019, 27, 935–944. [Google Scholar] [CrossRef]
- Li, Y.; Ye, Z.; Chen, S.; Pan, Z.; Zhou, Q.; Li, Y.Z.; Shuai, W.D.; Kuang, C.M.; Peng, Q.H.; Shi, W.; et al. ARHGEF19 interacts with BRAF to activate MAPK signaling during the tumorigenesis of non-small cell lung cancer. Int. J. Cancer 2018, 142, 1379–1391. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Tao, Y.; Peng, C.; Gu, P.; Wang, W. miR-503 regulates metastatic function through Rho guanine nucleotide exchanger factor 19 in hepatocellular carcinoma. J. Surg. Res. 2014, 188, 129–136. [Google Scholar] [CrossRef]
- Shang, H.; Liu, Y.; Li, Z.; Liu, Q.; Cui, W.; Zhang, L.; Pang, Y.; Liu, C.; Li, F. MicroRNA-874 functions as a tumor suppressor in rhabdomyosarcoma by directly targeting GEFT. Am. J. Cancer Res. 2019, 9, 668–681. [Google Scholar] [PubMed]
- Jeansonne, D.; DeLuca, M.; Marrero, L.; Lassak, A.; Pacifici, M.; Wyczechowska, D.; Wilk, A.; Reiss, K.; Peruzzi, F. Anti-tumoral effects of miR-3189-3p in glioblastoma. J. Biol. Chem. 2015, 290, 8067–8080. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Dai, C.; Yu, X.; Yin, X.B.; Zhou, F. Upregulated microRNA-194 impairs stemness of cholangiocarcinoma cells through the Rho pathway via inhibition of ECT2. J. Cell. Biochem. 2020. [Google Scholar] [CrossRef]
- Xu, J.; Yao, Q.; Hou, Y.; Xu, M.; Liu, S.; Yang, L.; Zhang, L.; Xu, H. MiR-223/Ect2/p21 signaling regulates osteosarcoma cell cycle progression and proliferation. Biomed. Pharmacother. 2013, 67, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tong, Z.; Liu, H. MiR-223-3p targeting epithelial cell transforming sequence 2 oncogene inhibits the activity, apoptosis, invasion and migration of MDA-MB-468 breast cancer cells. Onco Targets Ther. 2019, 12, 7675–7684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lussier, Y.A.; Xing, H.R.; Salama, J.K.; Khodarev, N.N.; Huang, Y.; Zhang, Q.; Khan, S.A.; Yang, X.; Hasselle, M.D.; Darga, T.E.; et al. MicroRNA expression characterizes oligometastasis(es). PLoS ONE 2011, 6, e28650. [Google Scholar] [CrossRef] [PubMed]
- Ottman, R.; Levy, J.; Grizzle, W.E.; Chakrabarti, R. The other face of miR-17-92a cluster, exhibiting tumor suppressor effects in prostate cancer. Oncotarget 2016, 7, 73739–73753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Li, D.; Cheng, Q.; Ma, Z.; Jiang, B.; Peng, R.; Chen, R.; Cao, Y.; Wan, X. MicroRNA-203 inhibits the proliferation and invasion of U251 glioblastoma cells by directly targeting PLD2. Mol. Med. Rep. 2014, 9, 503–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fite, K.; Elkhadragy, L.; Gomez-Cambronero, J. A Repertoire of MicroRNAs Regulates Cancer Cell Starvation by Targeting Phospholipase D in a Feedback Loop That Operates Maximally in Cancer Cells. Mol. Cell. Biol. 2016, 36, 1078–1089. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Pan, M.; Han, L.; Lu, H.; Hao, X.; Dong, Q. miR-338-3p suppresses neuroblastoma proliferation, invasion and migration through targeting PREX2a. FEBS Lett. 2013, 587, 3729–3737. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Liu, L.; Yao, J.; Ma, R.; Chang, D.; Li, Z.; Song, T.; Huang, C. miR-338-3p suppresses gastric cancer progression through a PTEN-AKT axis by targeting P-REX2a. Mol. Cancer Res. 2014, 12, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Deng, D.; Xue, L.; Shao, N.; Qu, H.; Wang, Q.; Wang, S.; Xia, X.; Yang, Y.; Zhi, F. miR-137 acts as a tumor suppressor in astrocytoma by targeting RASGRF1. Tumour Biol. 2016, 37, 3331–3340. [Google Scholar] [CrossRef]
- Li, X.; Sanda, T.; Look, A.T.; Novina, C.D.; von Boehmer, H. Repression of tumor suppressor miR-451 is essential for NOTCH1-induced oncogenesis in T-ALL. J. Exp. Med. 2011, 208, 663–675. [Google Scholar] [CrossRef]
- Hong, S.; Yu, S.; Li, J.; Yin, Y.; Liu, Y.; Zhang, Q.; Guan, H.; Li, Y.; Xiao, H. MiR-20b Displays Tumor-Suppressor Functions in Papillary Thyroid Carcinoma by Regulating the MAPK/ERK Signaling Pathway. Thyroid 2016, 26, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Yang, L. miR-124 inhibits the growth of glioblastoma through the downregulation of SOS1. Mol. Med. Rep. 2013, 8, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, Y.; Taniguchi, K.; Tsujino, T.; Heishima, K.; Inamoto, T.; Takai, T.; Minami, K.; Azuma, H.; Miyata, K.; Hayashi, K.; et al. Anti-cancer Effects of a Chemically Modified miR-143 on Bladder Cancer by Either Systemic or Intravesical Treatment. Mol. Ther. Methods Clin. Dev. 2019, 13, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.L.; Ho, B.C.; Sher, S.; Yu, S.L.; Yang, P.C. miR-146a and miR-370 coordinate enterovirus 71-induced cell apoptosis through targeting SOS1 and GADD45beta. Cell. Microbiol. 2015, 17, 802–818. [Google Scholar] [CrossRef]
- Xie, Q.; Yu, Z.; Lu, Y.; Fan, J.; Ni, Y.; Ma, L. microRNA-148a-3p inhibited the proliferation and epithelial–mesenchymal transition progression of non-small-cell lung cancer via modulating Ras/MAPK/Erk signaling. J. Cell. Physiol. 2018, 234, 12786–12799. [Google Scholar] [CrossRef]
- Lin, C.; Gao, B.; Yan, X.; Lei, Z.; Chen, K.; Li, Y.; Zeng, Q.; Chen, Z.; Li, H. MicroRNA 628 suppresses migration and invasion of breast cancer stem cells through targeting SOS1. Onco Targets Ther. 2018, 11, 5419–5428. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, J.; Gao, J.; Li, Y. MicroRNA-4728 mediated regulation of MAPK oncogenic signaling in papillary thyroid carcinoma. Saudi J. Biol. Sci. 2018, 25, 986–990. [Google Scholar] [CrossRef]
- Chiyomaru, T.; Tatarano, S.; Kawakami, K.; Enokida, H.; Yoshino, H.; Nohata, N.; Fuse, M.; Seki, N.; Nakagawa, M. SWAP70, actin-binding protein, function as an oncogene targeting tumor-suppressive miR-145 in prostate cancer. Prostate 2011, 71, 1559–1567. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Jiang, X.; Yan, P.; Zhan, L.; Zhu, H.; Wang, T.; Wen, J. Tumor-suppressive microRNA-10a inhibits cell proliferation and metastasis by targeting Tiam1 in esophageal squamous cell carcinoma. J. Cell. Biochem. 2018. [Google Scholar] [CrossRef]
- Moriarty, C.H.; Pursell, B.; Mercurio, A.M. miR-10b targets Tiam1: Implications for Rac activation and carcinoma migration. J. Biol. Chem. 2010, 285, 20541–20546. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Xu, Y.; Pan, L.; Feng, Y.; Luo, K.; Mu, Q.; Luo, G. miR-10b Downregulated by DNA Methylation Acts as a Tumor Suppressor in HPV-Positive Cervical Cancer via Targeting Tiam1. Cell. Physiol. Biochem. 2018, 51, 1763–1777. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lei, H.; Luo, M.; Wang, Y.; Dong, L.; Ma, Y.; Liu, C.; Song, W.; Wang, F.; Zhang, J.; et al. DNA methylation downregulated mir-10b acts as a tumor suppressor in gastric cancer. Gastric Cancer 2015, 18, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Li, B.; Sun, H.; Zhang, H. The predicted target gene validation, function, and prognosis studies of miRNA-22 in colorectal cancer tissue. Tumour Biol. 2017, 39, 1010428317692257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Song, Y.; Liu, T.J.; Cui, Y.B.; Jiang, Y.; Xie, Z.S.; Xie, S.L. miRNA-22 suppresses colon cancer cell migration and invasion by inhibiting the expression of T-cell lymphoma invasion and metastasis 1 and matrix metalloproteinases 2 and 9. Oncol. Rep. 2013, 29, 1932–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liang, S.; Jin, H.; Xu, C.; Ma, D.; Lu, X. Tiam1, negatively regulated by miR-22, miR-183 and miR-31, is involved in migration, invasion and viability of ovarian cancer cells. Oncol. Rep. 2012, 27, 1835–1842. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Li, W.; Liu, H.; Yang, L.; Liao, Q.; Cui, S.; Wang, H.; Zhao, L. miR-29b suppresses tumor growth and metastasis in colorectal cancer via downregulating Tiam1 expression and inhibiting epithelial-mesenchymal transition. Cell Death Dis. 2014, 5, e1335. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Tang, L.L.; Sun, Y.; Cui, R.X.; Wang, H.Y.; Huang, B.J.; He, Q.M.; Jiang, W.; Ma, J. MiR-29c suppresses invasion and metastasis by targeting TIAM1 in nasopharyngeal carcinoma. Cancer Lett. 2013, 329, 181–188. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Y.; Huang, J.; Wang, S.; Ni, W.; Guan, J.; Li, Q.; Zhang, Y.; Ding, Y.; Chen, B.; et al. MiR-141 suppresses the migration and invasion of HCC cells by targeting Tiam1. PLoS ONE 2014, 9, e88393. [Google Scholar] [CrossRef]
- Hu, J.; Lv, G.; Zhou, S.; Zhou, Y.; Nie, B.; Duan, H.; Zhang, Y.; Yuan, X. The Downregulation of MiR-182 Is Associated with the Growth and Invasion of Osteosarcoma Cells through the Regulation of TIAM1 Expression. PLoS ONE 2015, 10, e0121175. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yu, X.; Wang, Y.; Shen, J.; Wu, W.K.; Liang, J.; Feng, F. By downregulating TIAM1 expression, microRNA-329 suppresses gastric cancer invasion and growth. Oncotarget 2015, 6, 17559–17569. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Lu, L.; Liu, C.; Shan, L.; Yuan, D. MicroRNA-377 suppresses cell proliferation and invasion by inhibiting TIAM1 expression in hepatocellular carcinoma. PLoS ONE 2015, 10, e0117714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.H.; Lin, Y.H.; Chi, H.C.; Liao, C.H.; Liao, C.J.; Wu, S.M.; Chen, C.Y.; Tseng, Y.H.; Tsai, C.Y.; Lin, S.Y.; et al. Thyroid hormone regulation of miR-21 enhances migration and invasion of hepatoma. Cancer Res. 2013, 73, 2505–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottonham, C.L.; Kaneko, S.; Xu, L. miR-21 and miR-31 converge on TIAM1 to regulate migration and invasion of colon carcinoma cells. J. Biol. Chem. 2010, 285, 35293–35302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.W.; Lin, C.C.; Chen, C.N.; Huang, H.C.; Juan, H.F. Integrative network analysis reveals active microRNAs and their functions in gastric cancer. BMC Syst. Biol. 2011, 5, 99. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, A.; Huck, B.; Keller, B.; Strotbek, M.; Schmid, S.; Boerries, M.; Busch, H.; Muller, D.; Olayioye, M.A. miR149 functions as a tumor suppressor by controlling breast epithelial cell migration and invasion. Cancer Res. 2014, 74, 5256–5265. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, N.; Li, S.; Fang, J.H.; Chen, M.X.; Yang, J.; Jia, W.H.; Yuan, Y.; Zhuang, S.M. MicroRNA-195 suppresses angiogenesis and metastasis of hepatocellular carcinoma by inhibiting the expression of VEGF, VAV2, and CDC42. Hepatology 2013, 58, 642–653. [Google Scholar] [CrossRef]
- Li, X.; Zhu, J.; Liu, Y.; Duan, C.; Chang, R.; Zhang, C. MicroRNA-331-3p inhibits epithelial-mesenchymal transition by targeting ErbB2 and VAV2 through the Rac1/PAK1/beta-catenin axis in non-small-cell lung cancer. Cancer Sci. 2019, 110, 1883–1896. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, Y.W.; Xing, A.Y.; Xiang, S.; Shi, D.B.; Liu, L.; Li, Y.X.; Gao, P. Suppression of SPIN1-mediated PI3K-Akt pathway by miR-489 increases chemosensitivity in breast cancer. J. Pathol. 2016, 239, 459–472. [Google Scholar] [CrossRef]
- Li, M.; Zhang, S.; Wu, N.; Wu, L.; Wang, C.; Lin, Y. Overexpression of miR-499-5p inhibits non-small cell lung cancer proliferation and metastasis by targeting VAV3. Sci. Rep. 2016, 6, 23100. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Li, H.; Yin, C.; Sun, X.; Zheng, S.; Zhang, C.; Shi, L.; Liu, Y.; Lu, S. Dock1 promotes the mesenchymal transition of glioma and is modulated by MiR-31. Neuropathol. Appl. Neurobiol. 2017, 43, 419–432. [Google Scholar] [CrossRef]
- Li, H.; Mou, Q.; Li, P.; Yang, Z.; Wang, Z.; Niu, J.; Liu, Y.; Sun, Z.; Lv, S.; Zhang, B.; et al. MiR-486-5p inhibits IL-22-induced epithelial-mesenchymal transition of breast cancer cell by repressing Dock1. J. Cancer 2019, 10, 4695–4706. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-H.; Lan, H.-Y.; Huang, C.-H.; Tai, S.-K.; Tzeng, C.-H.; Kao, S.-Y.; Wu, K.-J.; Hung, M.-C.; Yang, M.-H. RAC1 activation mediates Twist1-induced cancer cell migration. Nat. Cell Biol. 2012, 14, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Gao, G.; Chu, K.; Yang, X.; Ren, S.; Li, Y.; Wu, H.; Huang, Y.; Zhou, C. Inhibition of RAC1-GEF DOCK3 by miR-512-3p contributes to suppression of metastasis in non-small cell lung cancer. Int. J. Biochem. Cell Biol. 2015, 61, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Elgamal, O.A.; Park, J.K.; Gusev, Y.; Azevedo-Pouly, A.C.; Jiang, J.; Roopra, A.; Schmittgen, T.D. Tumor suppressive function of mir-205 in breast cancer is linked to HMGB3 regulation. PLoS ONE 2013, 8, e76402. [Google Scholar] [CrossRef]
- Li, X.; Jiang, M.; Chen, D.; Xu, B.; Wang, R.; Chu, Y.; Wang, W.; Zhou, L.; Lei, Z.; Nie, Y.; et al. miR-148b-3p inhibits gastric cancer metastasis by inhibiting the Dock6/Rac1/Cdc42 axis. J. Exp. Clin. Cancer Res. 2018, 37, 71. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Y.; Wang, H.; Zhang, G.; Ding, Y.; Zhao, L. Tumor suppressor miR-1 restrains epithelial-mesenchymal transition and metastasis of colorectal carcinoma via the MAPK and PI3K/ AKT pathway. J. Transl. Med. 2014, 12, 244. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; An, H.; Wang, B.; Liao, Q.; Li, W.; Jin, X.; Cui, S.; Zhang, Y.; Ding, Y.; Zhao, L. miR-133a represses tumour growth and metastasis in colorectal cancer by targeting LIM and SH3 protein 1 and inhibiting the MAPK pathway. Eur. J. Cancer 2013, 49, 3924–3935. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Su, Z.; Fei, H.; Liu, X.; Pan, Q. miR-25 promotes hepatocellular carcinoma cell migration and invasion by inhibiting RhoGDI1. Oncotarget 2015, 6, 36231–36244. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Huang, S.; Wu, S.; Zhao, Y.; Liang, L.; Yan, M.; Ge, C.; Yao, J.; Chen, T.; Wan, D.; et al. Gain of miR-151 on chromosome 8q24.3 facilitates tumour cell migration and spreading through downregulating RhoGDIA. Nat. Cell Biol. 2010, 12, 390–399. [Google Scholar] [CrossRef]
- Chiyomaru, T.; Yamamura, S.; Zaman, M.S.; Majid, S.; Deng, G.; Shahryari, V.; Saini, S.; Hirata, H.; Ueno, K.; Chang, I.; et al. Genistein suppresses prostate cancer growth through inhibition of oncogenic microRNA-151. PLoS ONE 2012, 7, e43812. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Li, F.L.; Liu, P.S. MiR-151 promotes ovarian cancer through activation of akt/mTOR signaling pathway by decreasing RhoGDIA. Int. J. Clin. Exp. Med. 2016, 9, 11222–11229. [Google Scholar]
- Fletcher, C.E.; Sulpice, E.; Combe, S.; Shibakawa, A.; Leach, D.A.; Hamilton, M.P.; Chrysostomou, S.L.; Sharp, A.; Welti, J.; Yuan, W.; et al. Androgen receptor-modulatory microRNAs provide insight into therapy resistance and therapeutic targets in advanced prostate cancer. Oncogene 2019, 38, 5700–5724. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Xu, Y.; Dong, Y.; Cao, L.; Tong, J.; Zhou, X. Ectopic expression of miR-34a enhances radiosensitivity of non-small cell lung cancer cells, partly by suppressing the LyGDI signaling pathway. J. Radiat. Res. 2013, 54, 611–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahai, E.; Marshall, C.J. RHO-GTPases and cancer. Nat. Rev. Cancer 2002, 2, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chardin, P.; Camonis, J.H.; Gale, N.W.; Van Aelst, L.; Schlessinger, J.; Wigler, M.H.; Bar-Sagi, D. Human Sos1: A Guanine Nucleotide Exchange Factor for Ras That Binds to GRB2. Science 1993, 260, 1338–1343. [Google Scholar] [CrossRef]
- Dillon, L.M.; Bean, J.R.; Yang, W.; Shee, K.; Symonds, L.K.; Balko, J.M.; McDonald, W.H.; Liu, S.; Gonzalez-Angulo, A.M.; Mills, G.B.; et al. P-REX1 creates a positive feedback loop to activate growth factor receptor, PI3K/AKT and MEK/ERK signaling in breast cancer. Oncogene 2015, 34, 3968–3976. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, M.D.M.; Dharmawardhane, S. Targeting Rac and Cdc42 GTPases in Cancer. Cancer Res. 2018, 78, 3101–3111. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.L.; von Delft, F.; Brennan, P.E. Targeting the Small GTPase Superfamily through Their Regulatory Proteins. Angew. Chem. Int. Ed. Engl. 2019. [Google Scholar] [CrossRef] [Green Version]
- Orgaz, J.L.; Herraiz, C.; Sanz-Moreno, V. Rho GTPases modulate malignant transformation of tumor cells. Small GTPases 2014, 5, e29019. [Google Scholar] [CrossRef]
- Aspenstrom, P. Activated Rho GTPases in Cancer-The Beginning of a New Paradigm. Int. J. Mol. Sci. 2018, 19, 3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Zheng, Y. Approaches of targeting Rho GTPases in cancer drug discovery. Expert Opin. Drug Discov. 2015, 10, 991–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, S.; Gosens, R.; Wieland, T.; Schmidt, M. Paving the Rho in cancer metastasis: Rho GTPases and beyond. Pharmacol. Ther. 2018, 183, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Talevi, A. Multi-target pharmacology: Possibilities and limitations of the “skeleton key approach” from a medicinal chemist perspective. Front. Pharmacol. 2015, 6, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, L.; Zhou, M.; Tang, Z.; Zhou, E.; Xie, P.; Yi, W. MicroRNA-3646 promotes cell proliferation, migration, and invasion by targeting RhoA in breast cancer. Int. J. Clin. Exp. Pathol. 2017, 10, 61–71. [Google Scholar]
- Chevalier, B.; Adamiok, A.; Mercey, O.; Revinski, D.R.; Zaragosi, L.E.; Pasini, A.; Kodjabachian, L.; Barbry, P.; Marcet, B. miR-34/449 control apical actin network formation during multiciliogenesis through small GTPase pathways. Nat. Commun. 2015, 6, 8386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Liu, R.; Zhang, D.; Deng, Q.; Liu, B.; Chao, H.P.; Rycaj, K.; Takata, Y.; Lin, K.; Lu, Y.; et al. MicroRNA-141 suppresses prostate cancer stem cells and metastasis by targeting a cohort of pro-metastasis genes. Nat. Commun. 2017, 8, 14270. [Google Scholar] [CrossRef]
- Gumireddy, K.; Young, D.D.; Xiong, X.; Hogenesch, J.B.; Huang, Q.; Deiters, A. Small-molecule inhibitors of microrna miR-21 function. Angew. Chem. Int. Ed. Engl. 2008, 47, 7482–7484. [Google Scholar] [CrossRef] [Green Version]
- Melo, S.; Villanueva, A.; Moutinho, C.; Davalos, V.; Spizzo, R.; Ivan, C.; Rossi, S.; Setien, F.; Casanovas, O.; Simo-Riudalbas, L.; et al. Small molecule enoxacin is a cancer-specific growth inhibitor that acts by enhancing TAR RNA-binding protein 2-mediated microRNA processing. Proc. Natl. Acad. Sci. USA 2011, 108, 4394–4399. [Google Scholar] [CrossRef] [Green Version]
- Watashi, K.; Yeung, M.L.; Starost, M.F.; Hosmane, R.S.; Jeang, K.T. Identification of small molecules that suppress microRNA function and reverse tumorigenesis. J. Biol. Chem. 2010, 285, 24707–24716. [Google Scholar] [CrossRef] [Green Version]
- Monroig Pdel, C.; Chen, L.; Zhang, S.; Calin, G.A. Small molecule compounds targeting miRNAs for cancer therapy. Adv. Drug Deliv. Rev. 2015, 81, 104–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jepsen, J.S.; Sørensen, M.D.; Wengel, J. Locked Nucleic Acid: A Potent Nucleic Acid Analog in Therapeutics and Biotechnology. Oligonucleotides 2004, 14, 130–146. [Google Scholar] [CrossRef] [PubMed]
- Elmen, J.; Lindow, M.; Schutz, S.; Lawrence, M.; Petri, A.; Obad, S.; Lindholm, M.; Hedtjarn, M.; Hansen, H.F.; Berger, U.; et al. LNA-mediated microRNA silencing in non-human primates. Nature 2008, 452, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Krutzfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of microRNAs in vivo with ‘antagomirs’. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Duan, H.; Gan, L.; Liu, X.; Chen, F.; Shen, X.; Tang, Y.Q.; Wang, S. MicroRNA-1291 promotes endometrial fibrosis by regulating the ArhGAP29-RhoA/ROCK1 signaling pathway in a murine model. Mol. Med. Rep. 2017, 16, 4501–4510. [Google Scholar] [CrossRef]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef]
- Lin, Q.; Ponnusamy, R.; Widagdo, J.; Choi, J.A.; Ge, W.; Probst, C.; Buckley, T.; Lou, M.; Bredy, T.W.; Fanselow, M.S.; et al. MicroRNA-mediated disruption of dendritogenesis during a critical period of development influences cognitive capacity later in life. Proc. Natl. Acad. Sci. USA 2017, 114, 9188–9193. [Google Scholar] [CrossRef] [Green Version]
- You, K.; Li, S.Y.; Gong, J.; Fang, J.H.; Zhang, C.; Zhang, M.; Yuan, Y.; Yang, J.; Zhuang, S.M. MicroRNA-125b Promotes Hepatic Stellate Cell Activation and Liver Fibrosis by Activating RhoA Signaling. Mol. Ther. Nucleic Acids 2018, 12, 57–66. [Google Scholar] [CrossRef]
- Haussecker, D. Current issues of RNAi therapeutics delivery and development. J. Control. Release 2014, 195, 49–54. [Google Scholar] [CrossRef]
- Tiemann, K.; Rossi, J.J. RNAi-based therapeutics-current status, challenges and prospects. EMBO Mol. Med. 2009, 1, 142–151. [Google Scholar] [CrossRef]
- Li, Y.; Humphries, B.; Yang, C.; Wang, Z. Nanoparticle-Mediated Therapeutic Agent Delivery for Treating Metastatic Breast Cancer-Challenges and Opportunities. Nanomaterials 2018, 8, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Chen, J.; Huang, Z. Recent progress in microRNA-based delivery systems for the treatment of human disease. ExRNA 2019, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control. Release 2013, 172, 962–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Humphries, B.; Wang, Z.; Lang, S.; Huang, X.; Xiao, H.; Jiang, Y.; Yang, C. Complex Coacervation-Integrated Hybrid Nanoparticles Increasing Plasmid DNA Delivery Efficiency in Vivo. ACS Appl. Mater. Interfaces 2016, 8, 30735–30746. [Google Scholar] [CrossRef] [PubMed]
- Warheit, D.B.; Brown, S.C. What is the impact of surface modifications and particle size on commercial titanium dioxide particle samples?—A review of in vivo pulmonary and oral toxicity studies—Revised 11-6-2018. Toxicol. Lett. 2019, 302, 42–59. [Google Scholar] [CrossRef]
- Rabanel, J.M.; Adibnia, V.; Tehrani, S.F.; Sanche, S.; Hildgen, P.; Banquy, X.; Ramassamy, C. Nanoparticle heterogeneity: An emerging structural parameter influencing particle fate in biological media? Nanoscale 2019, 11, 383–406. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
Figure 1.
Canonical microRNA (miRNA) biogenesis pathway. Canonically, the miRNAs are transcribed in the nucleus via their own promoters or their host gene promoters by RNA polymerase II or III. This results in the formation of a primary miRNA (pri-miRNA) transcript, which can range from hundreds to thousands of nucleotides long. After a polyadenylation and capping event, pri-miRNAs undergo a microprocessing cleavage event by a ribonuclease (RNase) III type enzyme, Drosha, and its binding partner DiGeorge syndrome critical region gene 8 (DGCR8) resulting in a ~60–120 nucleotide-long precursor miRNA transcript (pre-miRNA). Pre-miRNAs are then exported out of the nucleus by the karyopherin exportin-5 to the cytoplasm, where the RNase II enzyme, Dicer, processes the transcripts to form a miRNA duplex. The unwinding of the miRNA duplex occurs and one strand is usually degraded (miRNA* (star) strand), while the other (mature miRNA) is loaded into the RNA-induced silencing complex (RISC). The RISC probes for targets of the miRNA in the genome. Once bound to a target mRNA, the RISC may induce a negative expression of the mRNA in three ways: (1) mRNA destabilization and degradation, (2) mRNA translational inhibition, or (3) mRNA cleavage. The path at which the mRNA is regulated depends upon multiple factors of the mature miRNA.
Figure 1.
Canonical microRNA (miRNA) biogenesis pathway. Canonically, the miRNAs are transcribed in the nucleus via their own promoters or their host gene promoters by RNA polymerase II or III. This results in the formation of a primary miRNA (pri-miRNA) transcript, which can range from hundreds to thousands of nucleotides long. After a polyadenylation and capping event, pri-miRNAs undergo a microprocessing cleavage event by a ribonuclease (RNase) III type enzyme, Drosha, and its binding partner DiGeorge syndrome critical region gene 8 (DGCR8) resulting in a ~60–120 nucleotide-long precursor miRNA transcript (pre-miRNA). Pre-miRNAs are then exported out of the nucleus by the karyopherin exportin-5 to the cytoplasm, where the RNase II enzyme, Dicer, processes the transcripts to form a miRNA duplex. The unwinding of the miRNA duplex occurs and one strand is usually degraded (miRNA* (star) strand), while the other (mature miRNA) is loaded into the RNA-induced silencing complex (RISC). The RISC probes for targets of the miRNA in the genome. Once bound to a target mRNA, the RISC may induce a negative expression of the mRNA in three ways: (1) mRNA destabilization and degradation, (2) mRNA translational inhibition, or (3) mRNA cleavage. The path at which the mRNA is regulated depends upon multiple factors of the mature miRNA.

Figure 2.
GTPase-activating proteins (GAPs), guanine nucleotide exchange factors (GEFs) and guanosine diphosphate (GDP) dissociation factors (GDIs) regulate small Rho GTPases. Small Rho GTPases are known as molecular switches due to the fact they cycle between “on” (GTP-bound) and “off” (GDP-bound) states. In the GTP-bound state, small Rho GTPases are able to regulate intracellular signaling cascades by binding and activating effector molecules. This signaling can be terminated by the intrinsic GTPase capability (GTP to GDP) of small Rho GTPases, which is enhanced by the interaction with GTPase-activating proteins (GAPs). While in the GDP-bound state, Rho GTPases can also interact with guanosine nucleotide dissociation inhibitors (GDIs), which sequester the small Rho GTPase and do not allow for the GDP to be exchanged for GTP. In order for GDIs to release the small Rho GTPase, a release factor must be present. Conversely, in the GDP-bound state, small Rho GTPases are unable to regulate downstream signaling, but can be reactivated by exchanging GDP for GTP, facilitated by the interaction with a guanine nucleotide exchange factor (GEF). Current literature has demonstrated that miRNAs can directly bind and downregulate the expression of RhoGAPs, GEFs, and GDIs to regulate cancer progression.
Figure 2.
GTPase-activating proteins (GAPs), guanine nucleotide exchange factors (GEFs) and guanosine diphosphate (GDP) dissociation factors (GDIs) regulate small Rho GTPases. Small Rho GTPases are known as molecular switches due to the fact they cycle between “on” (GTP-bound) and “off” (GDP-bound) states. In the GTP-bound state, small Rho GTPases are able to regulate intracellular signaling cascades by binding and activating effector molecules. This signaling can be terminated by the intrinsic GTPase capability (GTP to GDP) of small Rho GTPases, which is enhanced by the interaction with GTPase-activating proteins (GAPs). While in the GDP-bound state, Rho GTPases can also interact with guanosine nucleotide dissociation inhibitors (GDIs), which sequester the small Rho GTPase and do not allow for the GDP to be exchanged for GTP. In order for GDIs to release the small Rho GTPase, a release factor must be present. Conversely, in the GDP-bound state, small Rho GTPases are unable to regulate downstream signaling, but can be reactivated by exchanging GDP for GTP, facilitated by the interaction with a guanine nucleotide exchange factor (GEF). Current literature has demonstrated that miRNAs can directly bind and downregulate the expression of RhoGAPs, GEFs, and GDIs to regulate cancer progression.

Figure 3.
miRNA targeting of the Rho GTPase regulators has distinct effects on the processes of cancer progression. The downregulation of small Rho GTPase regulators (RhoGAPs, RhoGEFs, and RhoGDIs) has different effects on the processes known to drive the cancer progression. Much of the literature agrees that the downregulation of RhoGEFs results in the inhibition of progression, whereas the downregulation of RhoGDIs promotes cancer progression. Currently, there are conflicting results on the effects of RhoGAPs downregulation on cancer progression, with about half demonstrating inhibition and the other half promoting progression. Arrowheads (→) signify a positive effect on cancer progression. (┴) signifies a negative effect. ( ![Cancers 12 01092 i001]() ) signifies that both positive and negative effects have been frequently reported in the literature.
) signifies that both positive and negative effects have been frequently reported in the literature.
 ) signifies that both positive and negative effects have been frequently reported in the literature.
) signifies that both positive and negative effects have been frequently reported in the literature.
Figure 3.
miRNA targeting of the Rho GTPase regulators has distinct effects on the processes of cancer progression. The downregulation of small Rho GTPase regulators (RhoGAPs, RhoGEFs, and RhoGDIs) has different effects on the processes known to drive the cancer progression. Much of the literature agrees that the downregulation of RhoGEFs results in the inhibition of progression, whereas the downregulation of RhoGDIs promotes cancer progression. Currently, there are conflicting results on the effects of RhoGAPs downregulation on cancer progression, with about half demonstrating inhibition and the other half promoting progression. Arrowheads (→) signify a positive effect on cancer progression. (┴) signifies a negative effect. ( ![Cancers 12 01092 i001]() ) signifies that both positive and negative effects have been frequently reported in the literature.
) signifies that both positive and negative effects have been frequently reported in the literature.
) signifies that both positive and negative effects have been frequently reported in the literature.

{kind=link}
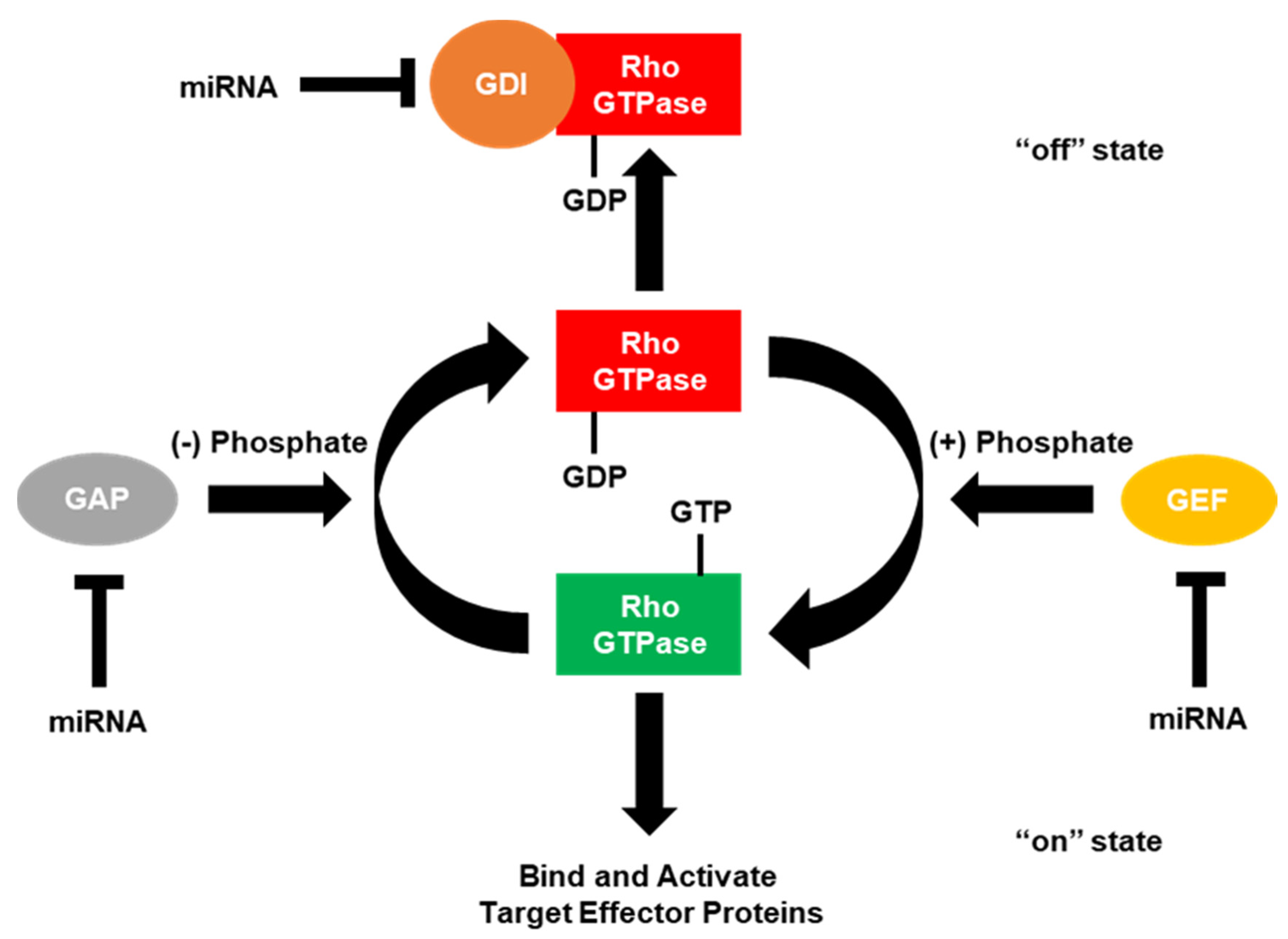{kind=link}
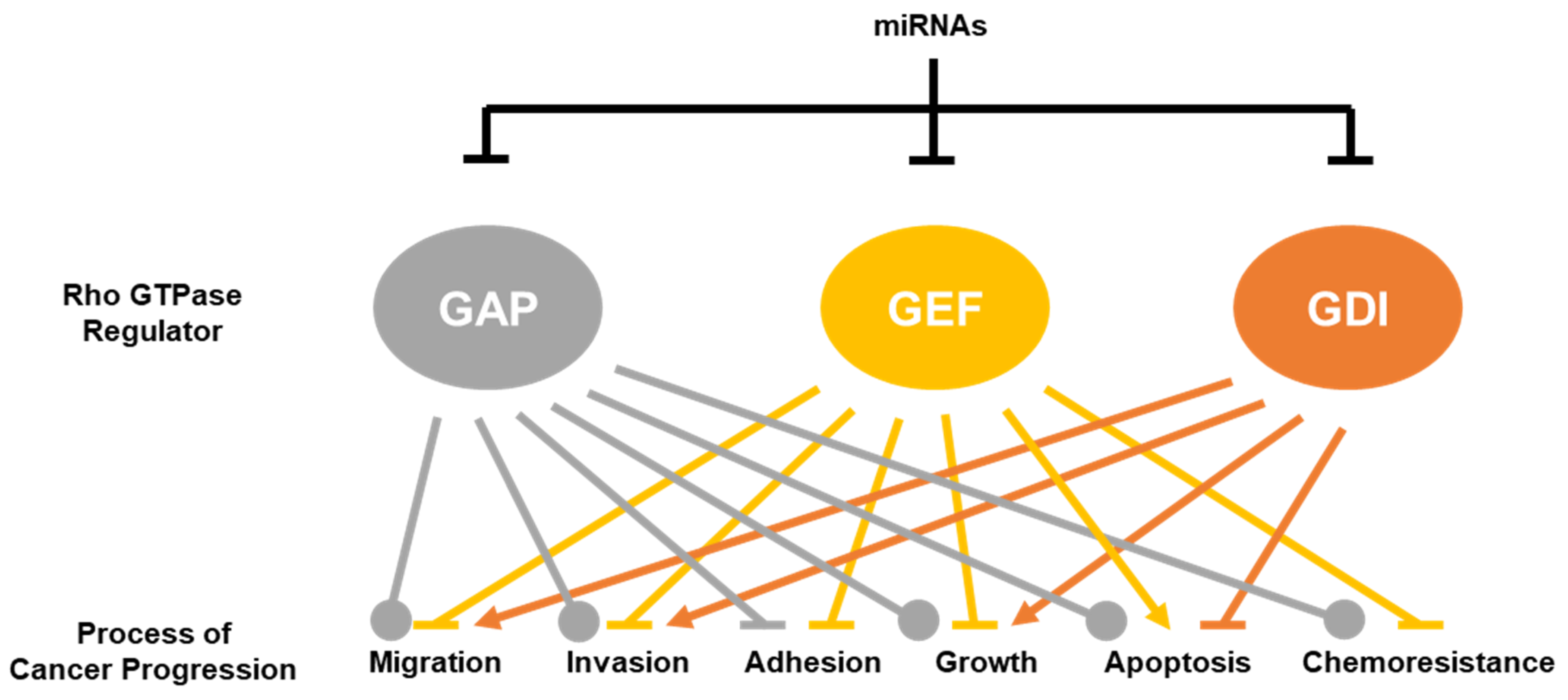{kind=link}
Table 1.
Mammalian Rho GTPases.
| GTPase (Alias) | Function | Citation |
|---|---|---|
| CDC42 (CDC42Hs) | Transduced signals to the actin cytoskeleton to initiate and maintain polarized growth and mitogen-activate protein morphogenesis; Responsible for the formation of filopodia in actin-based cell migration. | [32,33] |
| RAC1 | Transduced signals to the actin cytoskeleton to regulate the multiple signaling pathways that control actin cytoskeleton organization, transcription and proliferation; Responsible for the formation of lamellipodia in actin-based cell migration. | [34] |
| RAC2 | Transducer that localized to the plasma membrane, where it worked with Rac1 to regulate diverse processes, such as secretion, phagocytosis and cell polarization; Expressed primarily in hematopoietic cell lineages. | [35,36] |
| RAC3 | Involved in synaptic potentiation through regulating the actin cytoskeletal dynamics; Primarily expressed in the neurons of ganglia and the central nervous system. | [37,38] |
| RHOA | Involved in the regulation of cell adhesion and migration; Responsible for providing contractile force in cell migration through the formation of stress fibers and focal adhesions; Localized to the cytoplasm and to a certain degree the plasma membrane. | [39,40] |
| RHOB | Transducer involved in actin organization, cell migration, membrane and endosome trafficking, proliferation, DNA repair, and apoptosis; Thought to be an inhibitor of cancer progression; Localized to the endosomal membrane. | [28,41] |
| RHOBTB1 | Function not well known, did not play a major role in the organization of actin cytoskeleton dynamics; Not targetable by RhoGAPs, RhoGEFs, or RhoGDIs; Ubiquitously expressed, although high levels are found in skeletal muscle, placenta, stomach, kidney, testis, adrenal gland and uterus. | [42,43] |
| RHOBTB2 | Function not well known, did not play a major role in the organization of actin cytoskeleton dynamics; Not targetable by RhoGAPs, RhoGEFs, or RhoGDIs; Weakly expressed, although high levels were found in neural and cardiac tissues. | [42,43] |
| RHOBTB3 | Function not well known, did not play a major role in the organization of actin cytoskeleton dynamics; Not targetable by RhoGAPs, RhoGEFs, or RhoGDIs; Ubiquitously expressed, although high levels were found in placenta, testis, pancreas, adrenal and salivary gland, and neural and cardiac tissues. | [42,43] |
| RHOC | Responsible for actin cytoskeletal reorganization involved in promoting cell migration, proliferation, EMT, invasion, angiogenesis and metastasis; Localized to the cytoplasm and with undefined perinuclear structures. | [28,44] |
| RHOD | Controlled endocytic vesicle movement, Golgi homeostasis, and promoted actin stress fiber dissociation; Localized to the endosomal membrane. | [28,45] |
| RHOE (RND3) | Expressed ubiquitously; Inhibited contractility and the subsequent formation of actin stress fibers and focal adhesions; Drove cell rounding. GTPase-deficient, but constitutively bound to GTP in vivo. | [28] |
| RHOF (RIF) | Expressed ubiquitously; Promoted the formation of filopodia. | [28] |
| RHOG | Localized to caveolar vesicles; May have played a role in the inflammatory response; Involved in lamellipodia and filopodia formation, and membrane ruffling. | [28,38] |
| RHOH (TTF) | Expressed primarily in hematopoietic cell lineages; GTPase-deficient; Overexpression inhibited RAC1, RHOA, and CDC42 signaling; Not targetable by RhoGAPs, RhoGEFs, or RhoGDIs. | [28,36] |
| RHOJ (TCL) | Localized to the endosomal membrane; Promoted the formation of filopodia; Contributed to the focal adhesion turnover. | [1,28] |
| RHON (RND2) | Expressed primarily in testis, brain, and liver; GTPase-deficient; Involved in neurite outgrowth and cytokinesis. | [28,46] |
| RHOQ (TC10) | Localized to the endosomal membrane; Promoted the formation of filopodia; Implicated in receptor trafficking. | [28,46] |
| RHOS (RND1) | Expressed primarily in adult brain and liver; Inhibited contractility and the subsequent formation of actin stress fibers and focal adhesions; Drove cell rounding; GTPase-deficient. | [28,46] |
| RHOU (WRCH1) | Critical for Wnt signaling; Worked together with RAC1; Stimulated cell cycle progression; Promoted dissociation of stress fibers. | [1,28] |
| RHOV (WRCH2) | Promoted the formation of filopodia; Promoted the dissociation of stress fibers. | [28,46] |
RhoGAPs: Rho GTPase-activating proteins, RhoGEFs: Rho guanine nucleotide exchange factors, RhoGDIs: Rho guanosine diphosphate (GDP) dissociation inhibitors (GDIs).
Table 2.
Mammalian Rho GTPase regulators and their specificity for Rho GTPases in mammals.
| Function | Gene | Aliases | Rho GTPase Specificity | Citation |
|---|---|---|---|---|
| GTPase-activating protein (GAP) | ABR @ | MDB | RHOA, RAC1, RAC2, RAC3, CDC42 | [51,52] |
| ARAP1 | CENTD2 | RHOA, CDC42 | [53] | |
| ARAP2 | CENTD1, PARX | RHOA * | [54] | |
| ARAP3 | CENTD3, DRAG1 | RHOA, RAC1, CDC42 | [55,56] | |
| ARHGAP1 | CDC42GAP, p50RHOGAP, RHOGAP, RHOGAP1 | RHOA, RHOB, RHOC, RHOG, RHOJ, RHOQ, RAC1, RAC2, RAC3, RHOG, CDC42 | [51] | |
| ARHGAP2 | CHN1, n-CHIMERIN, DURS2, RHOGAP2 | RAC1, CDC42 | [57] | |
| ARHGAP3 | CHN2, β-CHIMERIN, RHOGAP3 | RAC1, CDC42 | [51] | |
| ARHGAP4 | SrGAP4, p115RHOGAP, RHOGAP4 | RHOA, RAC1, CDC42 | [58,59,60] | |
| ARHGAP5 | GFI2, p190BRHOGAP, RHOGAP5 | RHOA, RAC1, CDC42 | [61] | |
| ARHGAP6 | RHOGAPX-1, RHOGAP6 | RHOA, RAC3 | [62,63] | |
| ARHGAP7 | DLC1, STARD12, p122RHOGAP | RHOA, RHOB, RHOC, CDC42 | [51,64] | |
| ARHGAP8 | PP610, BPGAP1 | RHOA, RAC1, CDC42 | [65] | |
| ARHGAP9 | 10C, RGL1, MGC1295 | RHOA, RAC1, CDC42 | [66] | |
| ARHGAP10 | GRAF2, PSGAP | RHOA, CDC42 | [67] | |
| ARHGAP11A | MPGAP, GAP(1-12) | RHOA | [68] | |
| ARHGAP11B | B’-T, FAM7B1, GAP(1-8) | Not yet determined (Not RHOA) | [69] | |
| ARHGAP12 | RAC1, CDC42 | [70] | ||
| ARHGAP13 | SRGAP1, NMTC2 | RHOA, CDC42 | [71] | |
| ARHGAP14 | SRGAP3, WRP, MEGAP | RAC1, CDC42 | [72,73] | |
| ARHGAP15 | BM046 | RAC1 | [51] | |
| ARHGAP16P | Not yet determined | |||
| ARHGAP17 | RICH1, NADRIN, WBP15 | RHOA, RHOQ, RAC1, RAC2, CDC42 | [51,74] | |
| ARHGAP18 | MACGAP, SENEX | RHOA, RHOC | [75,76] | |
| ARHGAP19 | RHOA | [77] | ||
| ARHGAP20 | RARHOGAP | RHOA, RAC1, CDC42 | [78] | |
| ARHGAP21 | RHOA, RHOC, CDC42 | [48,79] | ||
| ARHGAP22 | RHOGAP22 | RAC1 | [80] | |
| ARHGAP23 | Not yet determined | |||
| ARHGAP24 | FILGAP, RCGAP72, p73RHOGAP | RAC1, CDC42 | [81,82] | |
| ARHGAP25 | HEL-S-308 | RAC1, CDC42 | [83] | |
| ARHGAP26 | GRAF1, OPHN1L | RHOA, RHOB, RHOC, RHOJ, RHOQ, RAC1, CDC42 | [51] | |
| ARHGAP27 | CAMGAP1, SH3D20 | RAC1, CDC42 | [82,84] | |
| ARHGAP28 | RHOA | [85] | ||
| ARHGAP29 | PARG1 | RHOA, RAC1, CDC42 | [86] | |
| ARHGAP30 | RHOA, RAC1 | [87] | ||
| ARHGAP31 | CDGAP, AOS1 | RAC1, CDC42 | [88] | |
| ARHGAP32 | GCGAP, GRIT, RICS, p200RHOGAP | RHOA, RAC1, CDC42 | [89] | |
| ARHGAP33 | TCGAP, NOMAGAP, SNX26 | RHOA, RHOQ, RAC1, CDC42 | [90] | |
| ARHGAP34 | SRGAP2, FNBP2 | RAC1 | [91,92,93] | |
| ARHGAP35 | GRLF1, p190RHOGAP | RHOA, RHOB, RHOC, RHOD, RHOJ, RHOQ, RAC1, RAC2, RAC3, CDC42 | [51] | |
| ARHGAP36 | RAC1 *,$, CDC42 *,$ | [94] | ||
| ARHGAP37 | STARD13, DLC2, GT650 | RHOA, CDC42 | [95,96] | |
| ARHGAP38 | STARD8, DLC3, STARTGAP3 | RHOA, CDC42 | [97] | |
| ARHGAP39 | CRGAP, VILSE | RAC1, CDC42 | [98] | |
| ARHGAP40 | Not yet determined | |||
| ARHGAP41 | OPHN1, OPN1, MRX60 | RHOA, RHOB, RHOC, RAC1, RAC2, RAC3, CDC42, RHOQ, RHOJ | [51] | |
| ARHGAP42 | TMEM133, GRAF3, AD031 | RHOA | [99] | |
| ARHGAP43 | SH3BP1 | RAC1, CDC42 | [100,101] | |
| ARHGAP44 | RICH2, NPC-A-10 | RAC1, CDC42 | [102,103] | |
| ARHGAP45 | HMHA1, HA-1, HLA-HA1 | RHOA, RAC1 | [104] | |
| ARHGAP46 | GMIP | RHOA | [105] | |
| ARHGAP47 | TAGAP, FKSG15, IDDM21 | RHOA, RHOH * | [106,107] | |
| ARHGAP48 | FAM13A | RHOA | [108] | |
| ARHGAP49 | FAM13B | Not yet determined | ||
| BCR @ | ALL, CML, PHL | RHOA, RAC1, RAC2, CDC42 | [51,52] | |
| INPP5B | 5PTase | RAC1, CDC42 | [109] | |
| INPP5F | OCRL, LOCR, DENT-2, NPHL2 | RHOA, RAC1, CDC42 | [110,111] | |
| IQGAP1 | SAR1, HUMORFA01, p195 | RHOA *, RHOB *, RAC1 *, CDC42 * | [112,113,114,115] | |
| IQGAP2 | RHOC *, RAC1 *, CDC42 * | [116,117] | ||
| IQGAP3 | RAC1 *, CDC42 * | [118] | ||
| MYO9B | MYR5, CELIAC4 | RHOA, RAC1, CDC42 | [119] | |
| PIK3R1 | p85ALPHA, p85α, AGM7, GRB1, IMD36 | RAC1, CDC42 | [120,121] | |
| PIK3R2 | p85BETA, p85β, MPPH | RAC1, CDC42 | [122] | |
| RACGAP1 | MGCRACGAP, CYK4, IDGAP | RHOA, RHOD, RAC1, RAC2, RAC3, CDC42 | [51,123,124] | |
| RALBP1 | RIP1, RLIP1, RLIP76 | RAC1, CDC42 | [125] | |
| Guanine nucleotide exchange factor (GEF) | ALS2 | ALS2CR6, ALSJ, IAHSP, PLSJ | RAC1 | [126] |
| ALS2CL | RN49018 | Not yet determined (Not RHOA, RAC1, or CDC42) | [127] | |
| ARHGEF1 | IMD62, LBCL2, LSC, p115RHOGEF | RHOA, RHOB, RHOC, RAC1$, CDC42$ | [94,128,129] | |
| ARHGEF2 | GEFH1, LFP40, LFC, GEF2 | RHOA, RHOB, RAC1 | [130,131,132] | |
| ARHGEF3 | XPLN, GEF3, STA3 | RHOA, RHOB | [133] | |
| ARHGEF4 | ASEF, GEF4, SMIM39, STM6 | RAC1, CDC42 | [134,135] | |
| ARHGEF5 | GEF5, TIM1 | RHOA, RHOB, RHOC, RHOG, RAC1, CDC42 | [136,137,138] | |
| ARHGEF6 | COOL2, MRX46, PIXA, ALPHAPIX | RAC1, CDC42 | [139] | |
| ARHGEF7 | BETAPIX, COOL1, PIXB | RAC1, CDC42 | [139] | |
| ARHGEF8 | NET1 | RHOA, RHOB | [140,141] | |
| ARHGEF9 | HPEM-2, PEM2 | CDC42 | [142] | |
| ARHGEF10 | GEF10, SNCV | RHOA, RHOB, RHOC | [143] | |
| ARHGEF10L | GRINCHGEF | RHOA, RHOB, RHOC | [144] | |
| ARHGEF11 | GTRAP48, PDZRHOGEF | RHOA, RHOB, RHOC | [129,145] | |
| ARHGEF12 | LARG, PRO2792 | RHOA, RHOB, RHOC | [129,146] | |
| ARHGEF13 | AKAP13, BRX, LBC, PRKA13 | RHOA, RHOB, RHOC | [147] | |
| ARHGEF14 | MCF2L, DBS, OST | RHOA, CDC42 | [148] | |
| ARHGEF15 | EPHEXIN5, VSMRHOGEF | RHOA, RAC1, CDC42 | [149,150,151] | |
| ARHGEF16 | GEF16, NBR | RHOG, RAC1 #, CDC42 | [152,153,154] | |
| ARHGEF17 | RHOGEF17, TEM4, p164RHOGEF | RHOA, RHOB, RHOC | [155,156,157] | |
| ARHGEF18 | RP78, SARHOGEF, p114RHOGEF | RHOA, RAC1 | [158,159] | |
| ARHGEF19 | WGEF | RHOA, RAC1, CDC42 | [160] | |
| ARHGEF21 | MCF2, DBL | RHOA, RHOB, RHOC, RHOG, CDC42 | [161,162] | |
| ARHGEF22 | MCF2L2 | RHOA, RAC1, CDC42 | [163,164] | |
| ARHGEF23 | TRIO, MEBAS, MRD44 | RHOA, RHOG, RAC1 | [165,166] | |
| ARHGEF24 | KALRN, DUO, CDH5, TRAD, HAPIP | RHOA #, RHOB #, RHOC, RHOG, RAC1, RAC2, CDC42 # | [167,168,169] | |
| ARHGEF25 | GEFT, p63RHOGEF | RHOA, RAC1, CDC42 | [170,171,172] | |
| ARHGEF26 | SGEF, CSGEF, HMFN1864 | RHOG | [173] | |
| ARHGEF27 | NGEF, EPHEXIN | RHOA, RAC1, CDC42 | [174] | |
| ARHGEF28 | RGNEF, RIP2, p190RHOGEF | RHOA, RHOB, RHOC, RAC1$ | [129,146,175] | |
| ARHGEF29 | SPATA13, ASEF2 | RHOA, RAC1, CDC42 | [176,177] | |
| ARHGEF30 | OBSCN, UNC89 | RHOA, RHOQ | [178,179] | |
| ARHGEF31 | ECT2 | RHOA, RHOB, RHOC, RHOG, RAC1, CDC42 | [141,180,181,182,183] | |
| ARHGEF32 | ECT2L, LFDH, FBXO49 | Not yet determined | ||
| ARHGEF33 | Not yet determined | |||
| ARHGEF34P | Not yet determined | |||
| ARHGEF35 | ARHGEF5L | Not yet determined | ||
| ARHGEF36 | DNMBP, TUBA, CTRCT48 | CDC42 | [184] | |
| ARHGEF37 | TUBA3 | CDC42$ | [184] | |
| ARHGEF38 | TUBA2 | CDC42$ | [184] | |
| ARHGEF39 | RAC1 | [185] | ||
| ARHGEF40 | SOLO | RHOA, RHOC, RAC1, RAC2, RAC3, CDC42 | [186,187] | |
| ARHGEF41 | PLEKHG1 | RAC1, CDC42 | [188] | |
| ARHGEF42 | PLEKHG2, CLG, LDAMD | RAC1, CDC42 | [189] | |
| ARHGEF43 | PLEKHG3 | RAC1, CDC42 | [190] | |
| ARHGEF44 | PLEKHG4, SCA4, PRTPHN1 | RHOA, RAC1, CDC42 | [191] | |
| DEF6 | IBP, SLAT, SWAP70L | RHOA, RAC1, CDC42 | [192,193] | |
| FARP1 | CDEP, PLEKHC2 | RHOA, RAC1, CDC42 | [94,194,195] | |
| FARP2 | FIR, FRG, PLEKHC3 | RAC1, CDC42 | [196,197] | |
| FGD1 | AAS, FGDY, MRXS16, ZFYVE3 | CDC42 | [198] | |
| FGD2 | ZFYVE4 | CDC42 | [199] | |
| FGD3 | ZFYVE5 | CDC42 | [200] | |
| FGD4 | CMT4H, FRABP, ZFYVE6 | CDC42 | [201] | |
| FGD5 | ZFYVE23 | RAC1, CDC42 | [202,203] | |
| FGD6 | ZFYVE24 | CDC42 | [204] | |
| ITSN1 | ITSN, SH3D1A, SH3P17 | CDC42 | [205] | |
| ITSN2 | PRO2015, SH3D1B, SH3P18, SWA, SWAP | CDC42 | [206] | |
| PLD2 | PLD1C | RHOA, RAC2 | [207,208] | |
| PLEKHG4B | Not yet determined | |||
| PLEKHG5 | CMTRIC, DSMA4, GEF720, SYX, TECH | RHOA | [209] | |
| PLEKHG6 | MYOGEF | RHOA, RHOC, RHOG, RAC1 | [210,211,212] | |
| PLEKHG7 | Not yet determined | |||
| PREX1 | P-REX1 | RHOA, RAC1, RAC2, CDC42 | [213] | |
| PREX2 | DEPDC2, P-REX2 | RHOG, RHOQ, RAC1, RAC2, RAC3, CDC42 | [214,215,216] | |
| RASGRF1 | GNRP, GRF1, CDC25, GRF55, CDC25L | RAC1, CDC42 | [217,218] | |
| RASGRF2 | GRF2 | RAC1, CDC42 | [218,219] | |
| RAP1GDS1 | GDS1, SMGGDS | RHOA, RHOC, CDC42 | [220,221] | |
| SOS1 | GF1, GGF1, GINGF, HGF, NS4 | RHOA, RAC1 | [222,223] | |
| SOS2 | NS9 | RAC1 | [222] | |
| SWAP70 | HSPC321 | RAC1, RAC2 | [224,225] | |
| TIAM1 | RHOA, RAC1, CDC42 | [226] | ||
| TIAM2 | STEF | RAC1, CDC42$ | [227,228,229] | |
| VAV1 | VAV | RHOA, RHOG, RAC1, CDC42 | [230,231,232] | |
| VAV2 | RHOA, RHOB, RHOG, RAC1, CDC42 | [230,231,232,233] | ||
| VAV3 | RHOA, RHOG, RAC1, CDC42 | [230,233,234] | ||
| DOCK1 | DOCK180, CED5 | RAC1, RAC2, RAC3 | [235,236,237] | |
| DOCK2 | IMD40 | RHOA, RAC1, RAC2 | [223,238,239] | |
| DOCK3 | MOCA, NEDIDHA, PBP | RAC1 | [240] | |
| DOCK4 | RAC1 | [241] | ||
| DOCK5 | RAC1, RAC2$, RAC3$ | [237,242] | ||
| DOCK6 | AOS2, ZIR1 | RAC1, CDC42 | [243] | |
| DOCK7 | EIEE23, ZIR2 | RAC1, RAC3, CDC42 | [244,245] | |
| DOCK8 | HEL-205, MRD2, ZIR8 | CDC42 | [246] | |
| DOCK9 | ZIZ1, ZIZIMIN1 | CDC42 | [247] | |
| DOCK10 | DRIP2, ZIZ3 | RHOJ$, RHOQ$, CDC42 | [248,249] | |
| DOCK11 | ACG, ZIZ2 | CDC42 | [249] | |
| GDP-dissociation inhibitor (GDI) | ARHGDIA | GDIA1, HEL-S-47e, NPHS8, RHOGDI | RHOA, RHOB$, RHOC, RHOG, RAC1, RAC2, CDC42 | [250,251,252,253,254] |
| ARHGDIB | D4, GDIA2, LYGDI, RAP1GN1, RHOGDI2 | RHOA, RAC1, CDC42 | [251,255,256] | |
| ARHGDIG | RHOGDI3 | RHOA, RHOB, RHOG, CDC42 | [257,258,259] |
* binds Rho GTPase, but does not stimulate the hydrolysis of GTP. # binds Rho GTPase, but does not facilitate the exchange of GDP for GTP. @ functions both as a GAP and a GEF. $ binding found, but no GTPase activity was analyzed; ABR: Active breakpoint cluster region-related protein, ALS2: Alsin rho guanine nucleotide exchange factor 2, ALS2CL: ALS2 C-terminal like, ARAP: ArfGAP with RhoGAP domain, ankyrin repeat and PH domain, ARHGAP: Rho GTPase activating protein, ARHGAP16P: Rho GTPase activating protein 16 pseudogene, ARHGEF: Rho guanine nucleotide exchange factor, ARHGEF10L: Rho guanine nucleotide exchange factor 10 like, ARHGDI: Rho GDP dissociation inhibitor, BCR: Breakpoint cluster region protein, DEF6: Differentially expressed in FDCP 6, DOCK: Dedicator of cytokinesis, FARP: FERM, ARH/RhoGEF and pleckstrin domain protein, FGD: FYVE, RhoGEF and PH domain containing, INPP: Inositol polyphosphate-1-phosphatase, IQGAP: IQ motif containing GTPase activating protein, ITSN: Intersectin, MYO9B: Myosin IXB, PIK3R: Phosphoinositide-3-kinase regulatory subunit, PLD2: Phospholipase D2, PLEKH: Pleckstrin homology and RhoGEF domain containing, PREX: Phosphatidylinositol-3,4,5-triphosphate dependent Rac exchange factor, RACGAP: Rac GTPase activating protein, RALBP: RalA binding protein, RAP1GDS1: Rap1 GTPase-GDP dissociation stimulator 1, RASGRF: Ras protein specific guanine nucleotide releasing factor, SOS: Son of sevenless, SWAP70: Switch associated protein 70, TIAM: T-lymphoma invasion and metastasis-inducing protein, VAV: Vav guanine nucleotide exchange factor.
Table 3.
RhoGAPs, RhoGEFs and RhoGDIs targeted by microRNAs in cancer.
| Rho GTPase Regulator | Direct Target of the miRNA | miRNA | miRNA is a Tumor Suppressor or Oncogene | Effect of microRNA Targeting | Reference |
|---|---|---|---|---|---|
| Rho GAPs | ABR | miR-762 | oncogene | Enhanced survival; Promoted gefitinib resistance; Promoted tumor formation. | [294] |
| ARHGAP1 | miR-19a | n/a | No effects on cells were assayed in this publication for ARHGAP1. | [295] | |
| ARHGAP1 | miR-34a/ miR-509 | tumor suppressor | Inhibited TGF-B-induced tumor cell invasion and metastasis; Inhibited growth, invasion and migration; Sensitized cells to cisplatin. | [296,297] | |
| ARHGAP1 | miR-130b/ miR-940 | oncogene | Enhanced CDC42 activity which led to enhanced AP-1-mediated growth, migration and invasion; Induced extensive osteoblastic lesions in calvarian tumors; drove osteogenic differentiation in mesenchymal stem cells. | [298,299] | |
| ARHGAP5 | miR-486 | tumor suppressor | Inhibited migration, invasion and metastasis. | [300] | |
| ARHGAP5 | miR-494 | oncogene | Enhanced the invasion of glioma cells through increased EGFR stabilization and the subsequent activation of ERK and Akt. | [301] | |
| ARHGAP5 | miR-744 | oncogene | Promoted cell migration and invasion; Directly interacted with the ARHGAP5 promoter to reduce expression. | [302,303] | |
| ARHGAP7 | miR-18a/ miR-106b/ miR-141/ miR-301a/ miR-429/ miR-483 | oncogene | Promoted growth, migration, invasion and EMT; Reduced cells in the G0/G1 phase of the cell cycle; Regulated by IGF2. | [304,305,306,307,308,309] | |
| ARHGAP7 | miR-200a/ miR-200b/ miR-200c | tumor suppressor | Reversed EMT and inhibited lung metastatic gene expression; Enhanced proliferation and colony formation, but reduced migration and invasion. | [310,311] | |
| ARHGAP9 | miR-224 | oncogene | Drove cell migration and invasion by the activation of upstream LPS, LTα, and TNFα inflammatory pathways. | [312] | |
| ARHGAP10 | miR-337 | tumor suppressor | Reduced migration, invasion and viability, but no effects on cell cycle. | [313] | |
| ARHGAP10 | miR-3174 | oncogene | Inhibited apoptosis and autophagy; Contributed to cisplatin resistance. | [314] | |
| ARHGAP12 | miR-20a | n/a | No effects on cells were assayed in this publication for ARHGAP12. | [315] | |
| ARHGAP13 | miR-124/ miR-340 | tumor suppressor | Negatively modulated ROCK1, MET and CTGF; Reduced proliferation, colony formation, migration and invasion; Promoted G1 cell cycle arrest and decreased cells in S-phase; Decreased phosphorylation of Rb. | [316] | |
| ARHGAP13 | miR-145 | oncogene | Drove an invasive phenotype. | [317] | |
| ARHGAP18 | miR-153/ miR-200b | tumor suppressor | Sponged (ceRNA) by CDKN2BAS to drive metastasis; Inhibited cell growth, migration, invasion, and metastasis; Enhanced stress fiber formation. | [50,318] | |
| ARHGAP19 | miR-200c | tumor suppressor | Suppressed anoikis resistance, migration, and EMT. | [319] | |
| ARHGAP21 | miR-224 | oncogene | Drove cell migration and invasion by the activation of upstream LPS, LTα and TNFα inflammatory pathways. | [312] | |
| ARHGAP24 | miR-590 | oncogene | Promoted cell viability, migration and invasion; Inhibited apoptosis. | [320] | |
| ARHGAP26 | miR-30b/ miR-573 | n/a | No effects on cells were assayed in this publication for ARHGAP26. | [321] | |
| ARHGAP29 | miR-200b | tumor suppressor | Inhibited migration and invasion; Drove actin cytoskeleton reorganization; Inhibited invadopodia formation. | [322] | |
| ARHGAP37 | miR-9/ miR-125b | oncogene | Promoted PDGFRB-induced angiogenic tube formation in vitro and vascular lacunae in vivo; Increased the proliferation, migration, invasion, EMT and metastasis; Drove the expression of a-SMA and vimentin through RHOA/ROCK signaling. | [323,324,325,326] | |
| BCR | miR-23a/ miR-320a | tumor suppressor | Inhibited growth and EMT; Drove cellular senescence; Blunted phosphorylation of PI3K, Akt, and NF-KB. | [327,328] | |
| IQGAP1 | miR-124 | n/a | Drove gene expression profile to that of the brain. | [329] | |
| IQGAP1 | miR-124/ miR-203/ miR-506 | tumor suppressor | Inhibited cell growth, migration, invasion, and adhesion; Induced cell cycle arrest at the G1-S checkpoint; Reversed EMT; Repressed ERK activation. | [330,331,332,333] | |
| IQGAP2 | miR-92a | n/a | No effects on cells were assayed in this publication for IQGAP2. | [334] | |
| PIK3R1 | miR-21/ miR-155/ miR-487a | oncogene | Promoted growth, invasion, EMT and metastasis; Increased MAPK and PI3K-Akt activation; Drove gemcitabine resistance. | [335,336,337,338] | |
| PIK3R1 | miR-29a | n/a | Inhibited Akt phosphorylation; Prevented the insulin-mediated inhibition of PEPCK. | [339] | |
| PIK3R1 | miR-29a/ miR-200b/ miR-200c/ miR-128/ miR-218/ miR-221/ miR-376a/ miR-486/ miR-503/ miR-542 | tumor suppressor | Suppressed growth, migration and invasion; Induced apoptosis; Blunted PI3K/Akt/mTOR and MMP9 signaling; Upregulated p53 expression; Enhanced chemosensitivity to gemcitabine and temozolomide. | [340,341,342,343,344,345,346,347,348] | |
| PIK3R2 | miR-30a/ miR-126/ miR-323/ miR-608 | tumor suppressor | Inhibited growth, migration and invasion; Decreased VEGF/PI3K/Akt signaling pathway activation; Suppressed G2/M cell cycle transition; Promoted EGFR-inhibitor sensitivity. | [349,350,351,352,353,354,355] | |
| RACGAP1 | miR-192/ miR-204/ miR-4324 | tumor suppressor | Inhibited growth, migration, invasion, and metastasis; Induced G0/G1 cell cycle arrest. | [356,357,358] | |
| Rho GEFs | ARHGEF1 | miR-143 | tumor suppressor | Inhibited migration, invasion, tumor growth and metastasis; Lowered the activities of RHOA, RAC1, and CDC42; Increased E-cadherin protein expression. | [359] |
| ARHGEF2 | miR-143/ miR-194 | tumor suppressor | Inhibited growth, migration, invasion, tumor growth and metastasis; Lowered the activities of RHOA, RAC1, and CDC42; Increased E-cadherin protein expression. | [359,360] | |
| ARHGEF3 | miR-138/ miR-200b | tumor suppressor | Suppressed migration and invasion; Reorganized the stress fibers to a more rounded shape; Inhibited invadopodia formation. | [322,361] | |
| ARHGEF6 | miR-23b/ miR-135a | tumor suppressor | Inhibited migration, invasion, spreading, adhesion, tumor growth, and metastasis; Inhibited lamellipodia formation; Blunted tumor initiation of CSCs. | [362,363] | |
| ARHGEF8 | miR-22/ miR-24 | oncogene | Increased colony formation, invasion, EMT, and chemoresistance; Decreased apoptosis. | [364,365] | |
| ARHGEF8 | miR-22/ miR-200b/ miR-206 | tumor suppressor | Decreased growth, migration, invasion, and chemoreistance; Inhibited stress fiber and invadopodia formation. | [322,366,367] | |
| ARHGEF19 | miR-29b/ miR-503 | tumor suppressor | Reduced growth, migration, invasion, tumor growth and metastasis; Blunted ERK signaling. | [368,369] | |
| ARHGEF25 | miR-874/ miR-3189 | tumor suppressor | Decreased growth, migration and invasion; Induced apoptosis. | [370,371] | |
| ARHGEF31 | miR-194/ miR-223/ | tumor suppressor | Decreased tumor growth, viability, migration, and invasion; Induced apoptosis; Induced p21, p27, and Rb expression. | [372,373,374] | |
| FGD1 | miR-200c | oncogene | Promoted lung metastasis. | [375] | |
| FGD4 | miR-17- 92a cluster | tumor suppressor | Decreased growth, EMT and migration; Delayed tumor onset; Reduced Akt and ERK activation. | [376] | |
| PLD2 | miR-203/ miR-887/ miR-3619 | tumor suppressor | Blunted growth, migration and invasion. | [377,378] | |
| PREX2 | miR-338 | tumor suppressor | Inhibited growth, migration and invasion; Induced G1 cell cycle arrest; Activated PTEN. | [379,380] | |
| RASGRF1 | miR-137/ miR-709 | tumor suppressor | Decreased growth, migration and invasion; Enhanced apoptosis; Reduced initiation and maintenance of leukemogenesis. | [381,382] | |
| SOS1 | miR-20b/ miR-124/ miR-143/ miR-146a/ miR-148a/ miR-628/ miR-4728 | tumor suppressor | Inhibited growth, viability, migration, invasion and EMT; Promoted apoptosis; Reduced ERK signaling. | [383,384,385,386,387,388,389] | |
| SWAP70 | miR-145 | tumor suppressor | Decreased growth, migration and invasion. | [390] | |
| TIAM1 | miR-10a/ miR-10b/ miR-22/ miR-29b/ miR-29c/ miR-31/ miR-141/ miR-182/ miR-183/ miR-329/ miR-377 | tumor suppressor | Inhibited growth, migration, invasion, EMT, tumor formation and metastasis; Induced apoptosis; Blocked Akt and ERK activation. | [391,392,393,394,395,396,397,398,399,400,401,402,403] | |
| TIAM1 | miR-21/ miR-31 | oncogene | Drove migration, invasion, and EMT; Increased B-catenin, vimentin and MMP2 expression. | [404,405] | |
| VAV2 | miR-148a/ miR-149/ miR-195/ miR-331 | tumor suppressor | Inhibited spreading, growth, adhesion, migration, invasion, EMT, tumor formation, angiogenesis, and metastasis. | [406,407,408,409] | |
| VAV3 | miR-489/ miR-499 | tumor suppressor | Blocked growth, migration and invasion; Induced apoptosis; Promoted sensitivity to chemotherapeutics. | [410,411] | |
| DOCK1 | miR-31/ miR-486 | tumor suppressor | Blunted migration, invasion, and metastasis; Suppressed N-cadherin protein expression. | [412,413] | |
| DOCK3 | let-7i/ miR-512 | tumor suppressor | Inhibited migration, invasion, adhesion, colonization and metastasis. | [414,415] | |
| DOCK4 | miR-205 | tumor suppressor | Reduced growth, migration and invasion; Decreased N-cadherin, vimentin, and ZEB1 protein expression. | [416] | |
| DOCK6 | miR-148b | tumor suppressor | Impaired migration, invasion and metastasis. | [417] | |
| Rho GDIs | ARHGDIA | miR-1/ miR-133a | tumor suppressor | Reversed EMT; Suppressed growth, migration, invasion and metastasis; Inhibited ERK and Akt phosphorylation. | [418,419] |
| ARHGDIA | miR-16/ miR-25/ miR-151/ miR-346/ miR-361 | oncogene | Promoted growth, migration, invasion, EMT and metastasis; Reduced apoptosis; Activated Akt signaling; Coordinated migration and invasion with FAK. | [288,420,421,422,423,424] | |
| ARHGDIB | miR-34a | tumor suppressor | Inhibited growth; Induced apoptosis; Enhanced sensitivity to radiation-induced apoptosis. | [420,425] |
α-SMA: Alpha smooth muscle actin, AP-1: Activator protein 1, CDKN2BAS: Cyclin dependent kinase inhibitor 2B antisense RNA, ceRNA: Competing endogenous RNA, CSC: Cancer stem cell, CTGF: Connective tissue growth factor, EGFR: Epidermal growth factor receptor, EMT: Epithelial-mesenchymal transition, ERK: Extracellular-signal-regulated kinase, FAK: Focal adhesion kinase, IGF2: Insulin like growth factor 2, LPS: Lipopolysaccharides, LTα: Lymphotoxin-alpha, MAPK: Mitogen-activated protein kinase, MET: Mesenchymal-epithelial transition factor, MMP2: Matrix metallopeptidase 2, MMP9: Matrix metallopeptidase 9, mTOR: Mammalian target of rapamycin, NF-ĸB: Nuclear factor kappa-light-chain-enhancer of activated B cells, PDGFRB: Platelet derived growth factor receptor beta, PEPCK: Phosphoenolpyruvate carboxykinase, PI3K: Phosphoinositide 3-kinase, PTEN: phosphatase and tensin homolog, Rb: Retinoblastoma protein, ROCK1: Rho associated coiled-coil containing protein kinase 1, TGF-β: Transforming growth factor beta, TNFa: Tumor necrosis factor alpha, VEGF: Vascular endothelial growth factor, ZEB1: Zinc finger E-box binding homeobox 1. n/a = not available.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Humphries, B.A.; Wang, Z.; Yang, C. MicroRNA Regulation of the Small Rho GTPase Regulators—Complexities and Opportunities in Targeting Cancer Metastasis. Cancers 2020, 12, 1092. https://doi.org/10.3390/cancers12051092
AMA Style
Humphries BA, Wang Z, Yang C. MicroRNA Regulation of the Small Rho GTPase Regulators—Complexities and Opportunities in Targeting Cancer Metastasis. Cancers. 2020; 12(5):1092. https://doi.org/10.3390/cancers12051092
Chicago/Turabian StyleHumphries, Brock A., Zhishan Wang, and Chengfeng Yang. 2020. "MicroRNA Regulation of the Small Rho GTPase Regulators—Complexities and Opportunities in Targeting Cancer Metastasis" Cancers 12, no. 5: 1092. https://doi.org/10.3390/cancers12051092
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.