Utilizing Experimental Mouse Model to Identify Effectors of Hepatocellular Carcinoma Induced by HBx Antigen

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
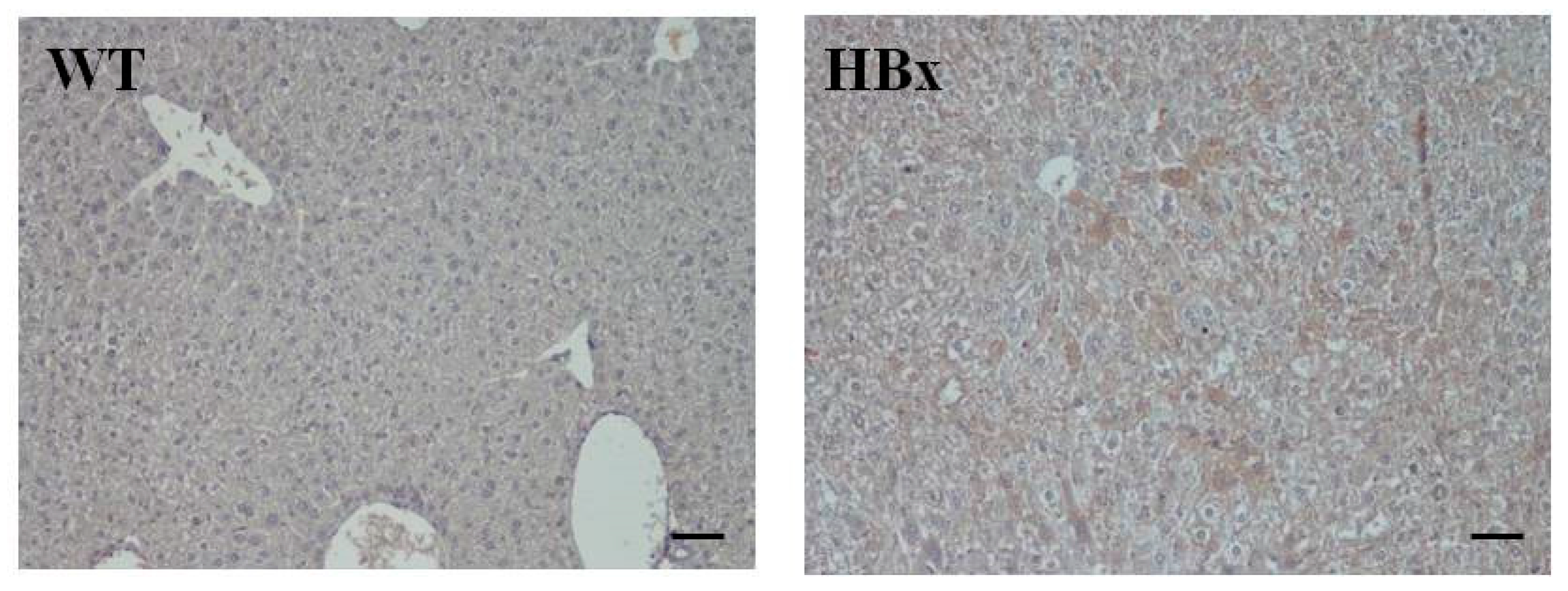
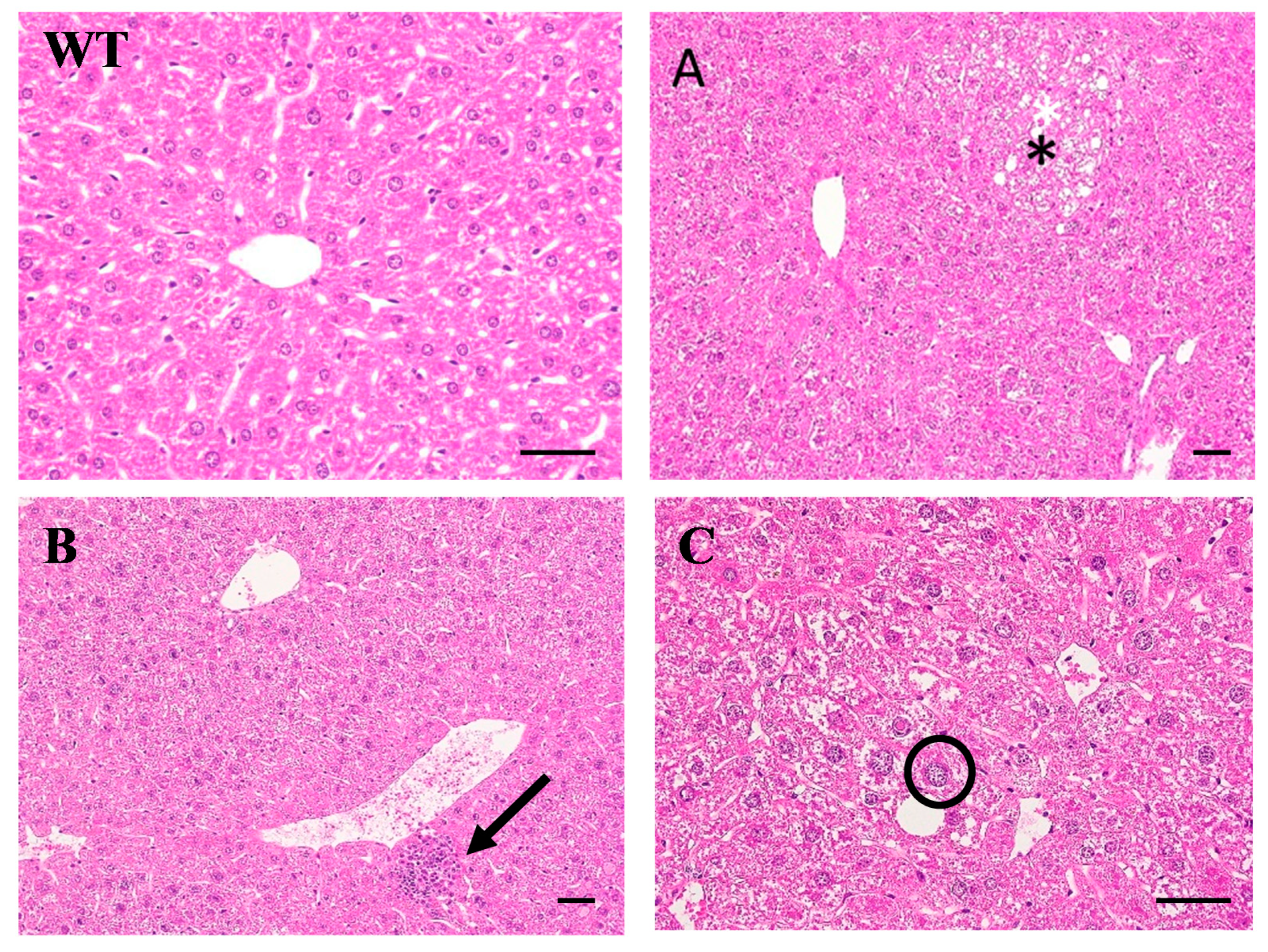
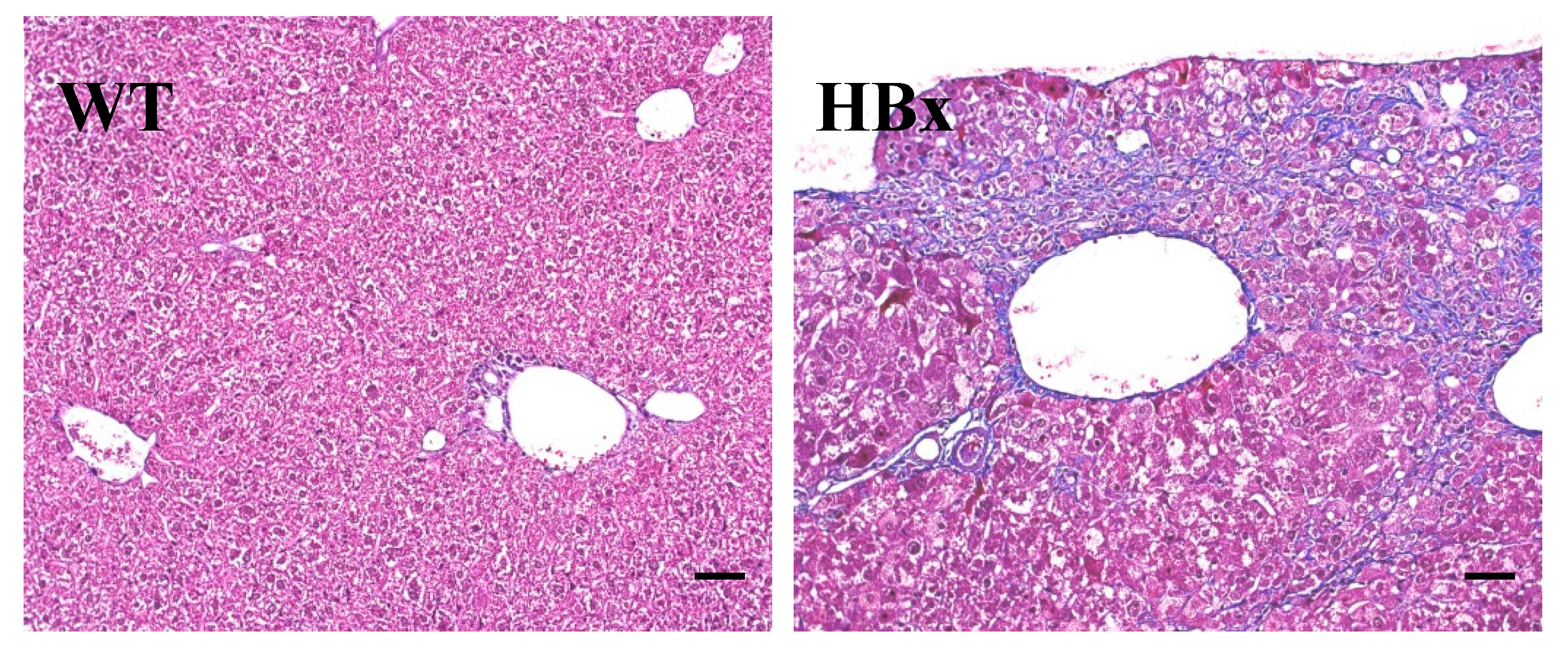
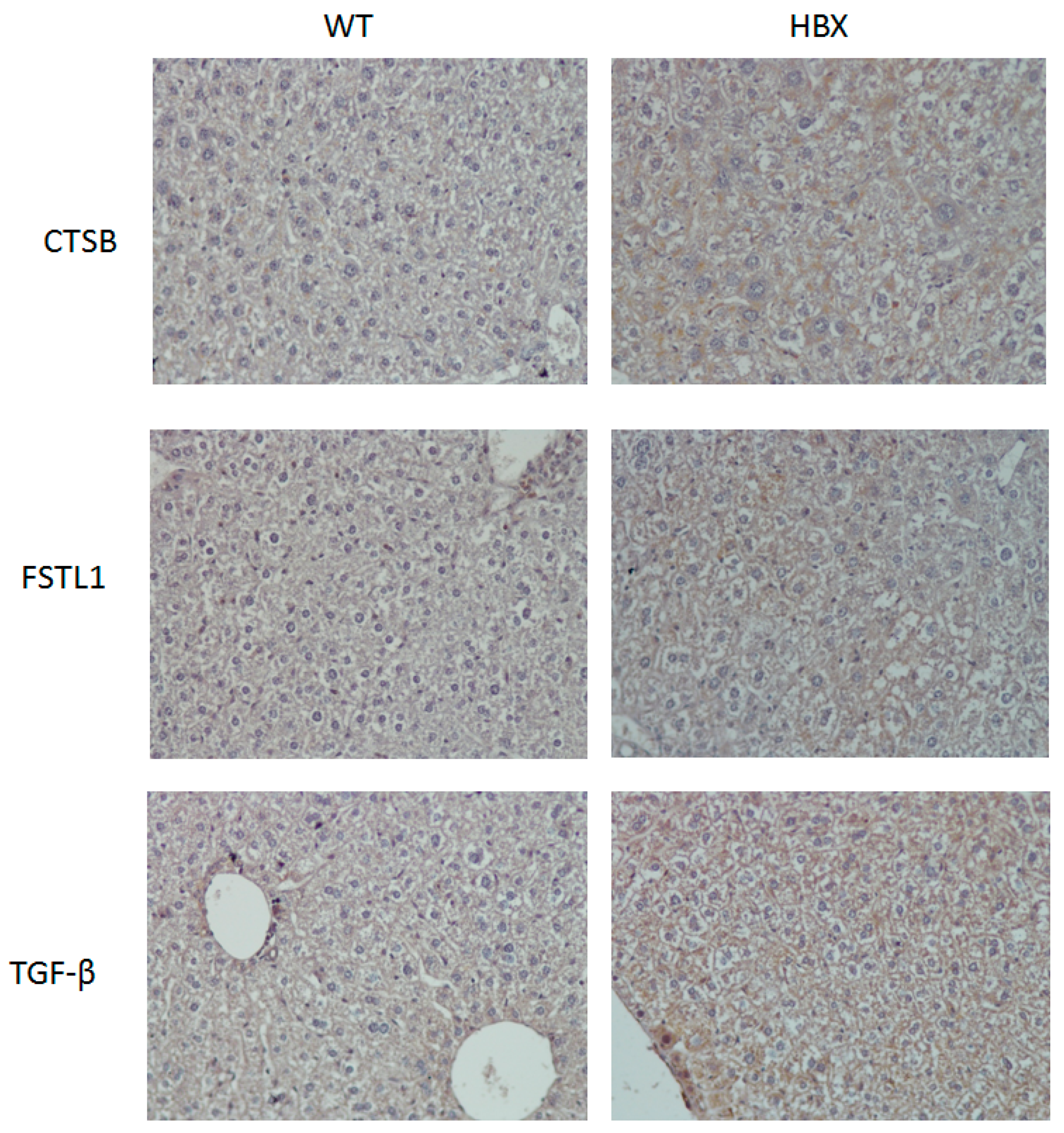
2.1. Pathological Findings in HBx Mice
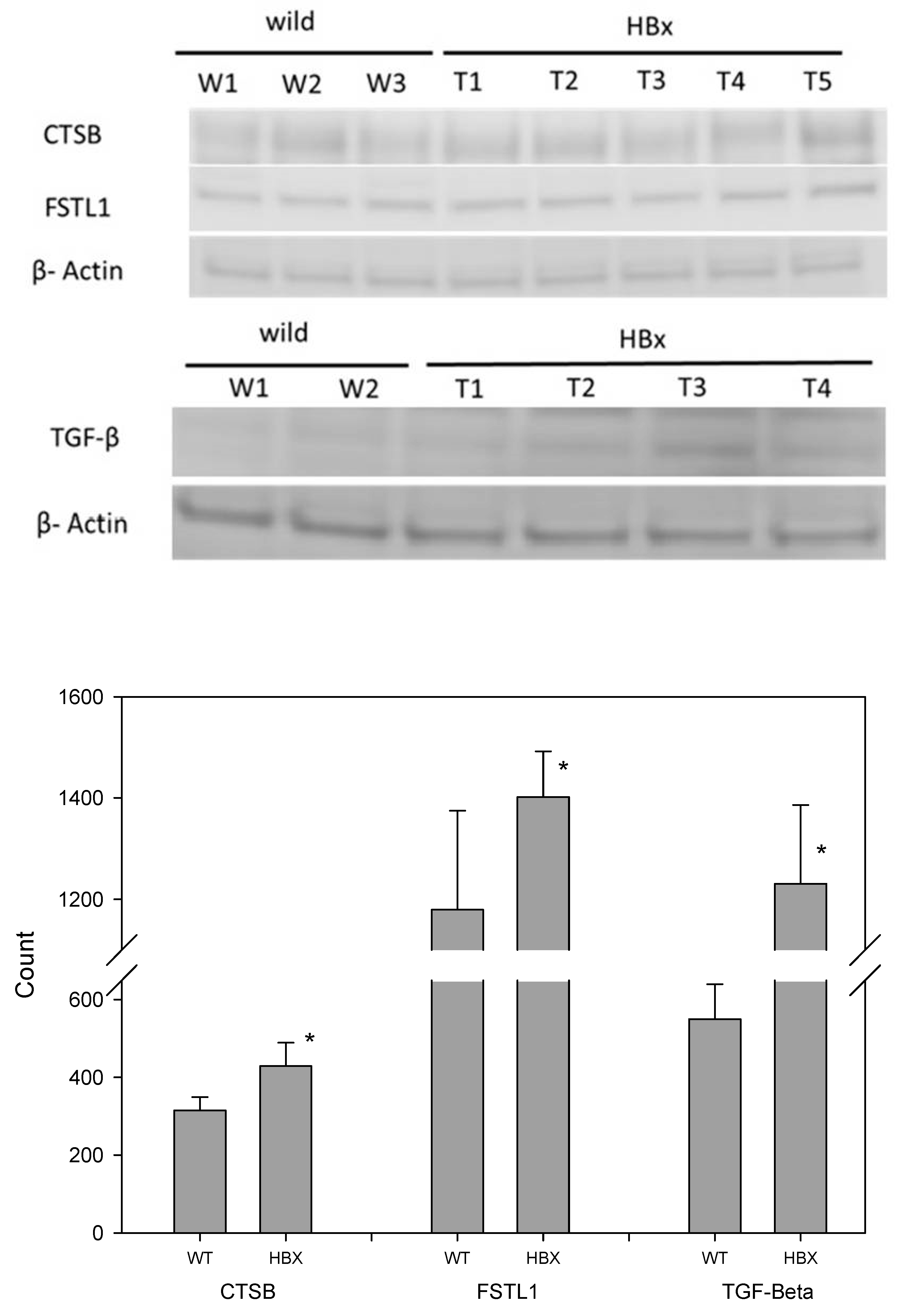
2.2. Proteomic Analysis
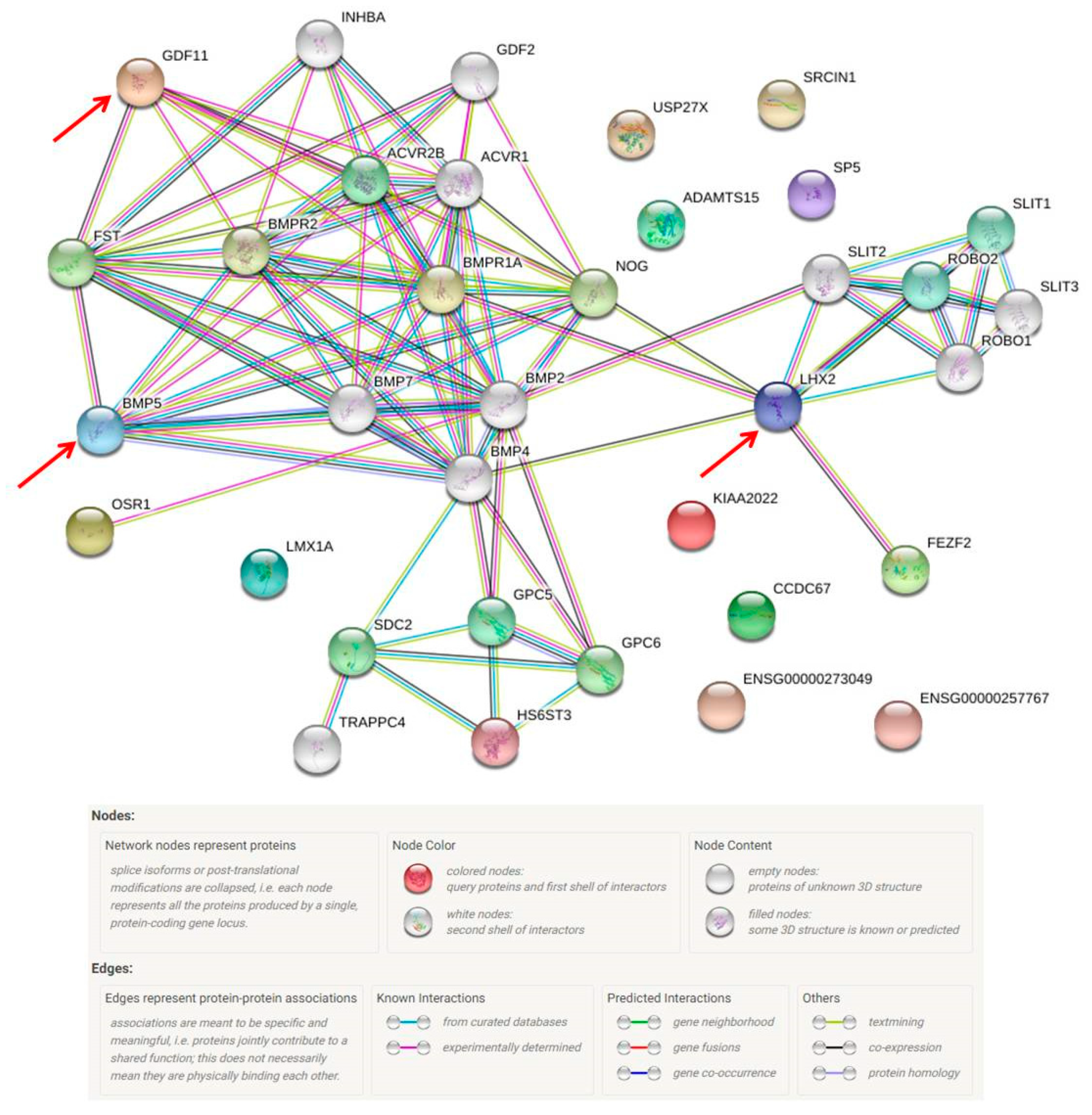
2.3. Missing Protein Analysis
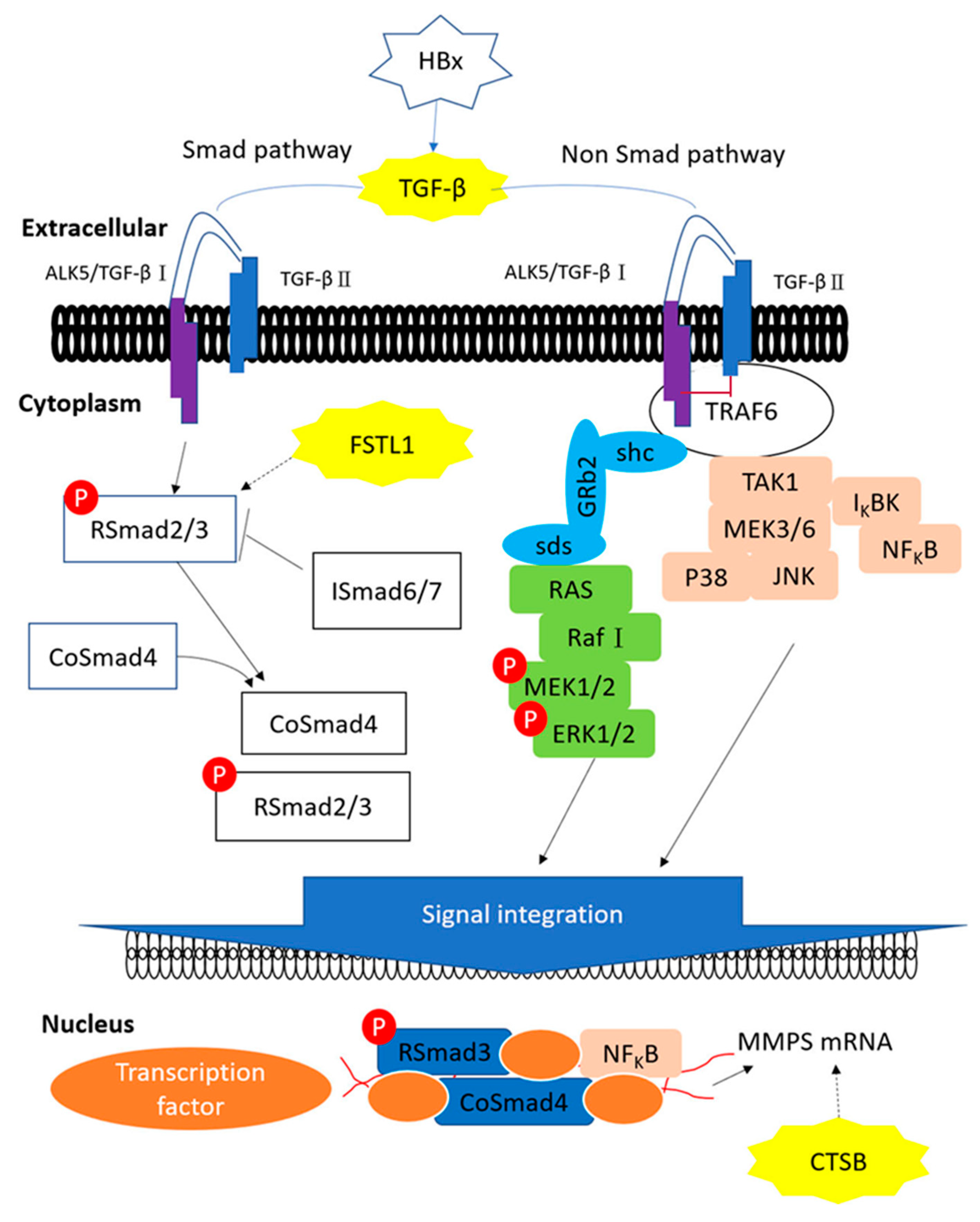
3. Discussion
4. Materials and Methods
4.1. Hepatocyte-Specific HBx Transgenic and Wild-Type Mice
4.2. Serum Alanine Aminotransferase and Aspartate Aminotransferase Tests
4.3. Histopathological Examination and Immunohistochemical Staining
4.4. Sample Digestion and Preparation
4.5. Immunoblotting
4.6. Proteomic Analysis
4.7. Protein Database Search and STRING Database for Protein–Protein Interaction Network Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Serag, H.B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 2012, 142, 1264–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, K.; Brunetto, M.; Seto, W.K.; Lim, Y.S.; Fung, S.; Marcellin, P.; Ahn, S.H.; Izumi, N.; Chuang, W.L.; Bae, H.; et al. 96weeks treatment of tenofovir alafenamide vs. tenofovir disoproxil fumarate for hepatitis B virus infection. J. Hepatol. 2018, 68, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.A.; Kowgier, M.; Hansen, B.E.; Brouwer, W.P.; Maan, R.; Wong, D.; Shah, H.; Khalili, K.; Yim, C.; Heathcote, E.J.; et al. Toronto HCC risk index: A validated scoring system to predict 10-year risk of HCC in patients with cirrhosis. J. Hepatol. 2018, 68, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Guerrieri, F.; Belloni, L.; Pediconi, N.; Levrero, M. Molecular mechanisms of HBV-associated hepatocarcinogenesis. Semin. Liver Dis. 2013, 33, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Nassal, M. HBV cccDNA: Viral persistence reservoir and key obstacle for a cure of chronic hepatitis B. Gut 2015, 64, 1972–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.D.; Trivedi, A.; Johnson, D.L. Regulation of RNA polymerase I-dependent promoters by the hepatitis B virus X protein via activated Ras and TATA-binding protein. Mol. Cell. Biol. 1998, 18, 7086–7094. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Huo, B.; Liu, J.; Huang, X.; Zhang, S.; Feng, T. Hepatitis B virus X reduces hepatocyte apoptosis and promotes cell cycle progression through the Akt/mTOR pathway In Vivo. Gene 2019, 691, 87–95. [Google Scholar] [CrossRef]
- Slagle, B.L.; Bouchard, M.J. Role of HBx in hepatitis B virus persistence and its therapeutic implications. Curr. Opin. Virol. 2018, 30, 32–38. [Google Scholar] [CrossRef]
- Murakami, S. Hepatitis B virus X protein: A multifunctional viral regulator. J. Gastroenterol. 2001, 36, 651–660. [Google Scholar] [CrossRef]
- Wu, B.K.; Li, C.C.; Chen, H.J.; Chang, J.L.; Jeng, K.S.; Chou, C.K.; Hsu, M.T.; Tsai, T.F. Blocking of G1/S transition and cell death in the regenerating liver of Hepatitis B virus X protein transgenic mice. Biochem. Biophys. Res. Commun. 2006, 340, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.F.; Fu, S.L.; Kao, C.H.; Yang, C.W.; Lin, C.H.; Hsu, M.T.; Tsai, T.F. Chemopreventive effect of silymarin on liver pathology in HBV X protein transgenic mice. Cancer Res. 2008, 68, 2033–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seibert, V.; Ebert, M.P.; Buschmann, T. Advances in clinical cancer proteomics: SELDI-ToF-mass spectrometry and biomarker discovery. Brief. Funct. Genom. Proteom. 2005, 4, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, E.; Van Hees, S.; Goethals, S.; Smits, E.; Huizing, M.; Francque, S.; De Winter, B.; Michielsen, P.; Vanwolleghem, T. Biobanking for Viral Hepatitis Research. Front. Med. (Lausanne) 2019, 6, 183. [Google Scholar] [CrossRef]
- Horvatovich, P.; Végvári, Á.; Saul, J.; Park, J.G.; Qiu, J.; Syring, M.; Pirrotte, P.; Petritis, K.; Tegeler, T.J.; Aziz, M.; et al. In Vitro Transcription/Translation System: A Versatile Tool in the Search for Missing Proteins. J. Proteome Res. 2015, 14, 3441–3451. [Google Scholar] [CrossRef]
- Adkins, J.N.; Varnum, S.M.; Auberry, K.J.; Moore, R.J.; Angell, N.H.; Smith, R.D.; Springer, D.L.; Pounds, J.G. Toward a human blood serum proteome: Analysis by multidimensional separation coupled with mass spectrometry. Mol. Cell. Proteom. 2002, 1, 947–955. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.H.; Chen, W.J.; Fu, Y.S.; Huang, B.; Tsai, W.C.; Chen, Y.M.; Lin, P.C.; Yuan, C.H.; Tyan, Y.C. Utilizing glycine N-methyltransferasegene knockout mice as a model for identification of missing proteins in hepatocellular carcinoma. Oncotarget 2018, 9, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Teng, C.F.; Yu, C.H.; Chang, H.Y.; Hsieh, W.C.; Wu, T.H.; Lin, J.H.; Wu, H.C.; Jeng, L.B.; Su, I.J. Chemopreventive effect of phytosomal curcumin on hepatitis b virus-related hepatocellular carcinoma in a transgenic mouse model. Sci. Rep. 2019, 9, 10338. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.J.; Chen, T.L.; Lee, T.S.; Wang, H.A.; Wang, C.K.; Liao, L.Y.; Liu, R.S.; Huang, S.F.; Chen, Y.M. Glycine N-methyltransferase deficiency affects Niemann-Pick type C2 protein stability and regulates hepatic cholesterol homeostasis. Mol. Med. 2012, 18, 412–422. [Google Scholar] [CrossRef]
- Li, C.H.; Yen, C.H.; Chen, Y.F.; Lee, K.J.; Fang, C.C.; Zhang, X.; Lai, C.C.; Huang, S.F.; Lin, H.K.; Chen, Y.M. Characterization of the GNMT-HectH9-PREX2 tripartite relationship in the pathogenesis of hepatocellular carcinoma. Int. J. Cancer 2017, 140, 2284–2297. [Google Scholar] [CrossRef]
- Ijaz, B.; Ahmad, W.; Javed, F.T.; Gull, S.; Hassan, S. Revised cutoff values of ALT and HBV DNA level can better differentiate HBeAg (-) chronic inactive HBV patients from active carriers. Virol. J. 2011, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.W.; Hsia, Y.; Yang, W.Y.; Lin, Y.I.; Li, C.C.; Tsai, T.F.; Chang, K.W.; Shieh, G.S.; Tsai, S.F.; Wang, H.D.; et al. Identification of the common regulators for hepatocellular carcinoma induced by hepatitis B virus X antigen in a mouse model. Carcinogenesis 2012, 33, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Debes, J.D.; Carrera, E.; Mattos, A.Z.; Prieto, J.E.; Boonstra, A.; ESCALON investigators. Hepatocellular carcinoma, a unique tumor with a lack of biomarkers. Ann. Hepatol. 2019, 18, 786–787. [Google Scholar] [CrossRef] [PubMed]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochim. Biophys. Acta 2012, 1824, 68–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keppler, D.; Sloane, B.F. Cathepsin B: Multiple enzyme forms from a single gene and their relation to cancer. Enzyme Protein 1996, 49, 94–105. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Bhargava, V.; Hansell, E.; Huling, S.; Kuwahara, T.; Matley, M.; Coussens, L.; Warren, R. A functional proteomics screen of proteases in colorectal carcinoma. Mol. Med. 2000, 6, 450–460. [Google Scholar] [CrossRef]
- Srisomsap, C.; Subhasitanont, P.; Otto, A.; Mueller, E.C.; Punyarit, P.; Wittmann-Liebold, B.; Svasti, J. Detection of cathepsin B up-regulation in neoplastic thyroid tissues by proteomic analysis. Proteomics 2002, 2, 706–712. [Google Scholar] [CrossRef]
- Wulfkuhle, J.D.; Sgroi, D.C.; Krutzsch, H.; McLean, K.; McGarvey, K.; Knowlton, M.; Chen, S.; Shu, H.; Sahin, A.; Kurek, R.; et al. Proteomics of human breast ductal carcinoma in situ. Cancer Res. 2002, 62, 6740–6749. [Google Scholar]
- Krueger, S.; Haeckel, C.; Buehling, F.; Roessner, A. Inhibitory effects of antisense cathepsin B cDNA transfection on invasion and motility in a human osteosarcoma cell line. Cancer Res. 1999, 59, 6010–6014. [Google Scholar]
- Ruan, J.; Zheng, H.; Rong, X.; Rong, X.; Zhang, J.; Fang, W.; Zhao, P.; Luo, R. Over-expression of cathepsin B in hepatocellular carcinomas predicts poor prognosis of HCC patients. Mol. Cancer. 2016, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Chen, J.; Wang, J.; Ye, J.; Tan, H.; Xu, L. Expression of cathepsin B in human hepatocellular carcinoma and its prognostic significance. Int. J. Clin. Exp. Pathol. 2016, 9, 1343–1350. [Google Scholar]
- Hambrock, H.O.; Kaufmann, B.; Müller, S.; Hanisch, F.; Nose, K.; Paulsson, M.; Maurer, P.; Hartmann, U. Structural characterization of TSC-36/Flik: Analysis of two charge isoforms. J. Biol. Chem. 2004, 279, 11727–11735. [Google Scholar] [CrossRef] [Green Version]
- Mattiotti, A.; Prakash, S.; Barnett, P.; van den Hoff, M.J.B. Follistatin-like 1 in development and human diseases. Cell. Mol. Life Sci. 2018, 75, 2339–2354. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wu, Y.; Wang, C.; Liu, Z.; Xu, M.; Zheng, X. FSTL1 contributes to tumor progression via attenuating apoptosis in a AKT/GSK-3beta-dependent manner in hepatocellular carcinoma. Cancer Biomark. 2017, 20, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.; Kale, V.P. TGF-beta signaling and its role in the regulation of hematopoietic stem cells. Syst. Synth. Boil. 2015, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fransvea, E.; Angelotti, U.; Antonaci, S.; Giannelli, G. Blocking transforming growth factor-beta up-regulates E-cadherin and reduces migration and invasion of hepatocellular carcinoma cells. Hepatology 2008, 47, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Mazzocca, A.; Fransvea, E.; Lavezzari, G.; Antonaci, S.; Giannelli, G. Inhibition of transforming growth factor beta receptor I kinase blocks hepatocellular carcinoma growth through neo-angiogenesis regulation. Hepatology 2009, 50, 1140–1151. [Google Scholar] [CrossRef]
- Fransvea, E.; Mazzocca, A.; Antonaci, S.; Giannelli, G. Targeting transforming growth factor (TGF)-betaRI inhibits activation of beta1 integrin and blocks vascular invasion in hepatocellular carcinoma. Hepatology 2009, 49, 839–850. [Google Scholar] [CrossRef]
- Bergamini, C.; Sgarra, C.; Trerotoli, P.; Lupo, L.; Azzariti, A.; Antonaci, S.; Giannelli, G. Laminin-5 stimulates hepatocellular carcinoma growth through a different function of alpha6beta4 and alpha3beta1 integrins. Hepatology 2007, 46, 1801–1809. [Google Scholar] [CrossRef]
- Li, X.Y.; Hu, S.Q.; Xiao, L. The cancer-associated fibroblasts and drug resistance. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 2112–2119. [Google Scholar]
- Porcelli, L.; Iacobazzi, R.M.; Di Fonte, R.; Serratì, S.; Intini, A.; Solimando, A.G.; Brunetti, O.; Calabrese, A.; Leonetti, F.; Azzariti, A.; et al. CAFs and TGF-β Signaling Activation by Mast Cells Contribute to Resistance to Gemcitabine/Nabpaclitaxel in Pancreatic Cancer. Cancers 2019, 11, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casadei Gardini, A.; Scarpi, E.; Faloppi, L.; Scartozzi, M.; Silvestris, N.; Santini, D.; de Stefano, G.; Marisi, G.; Negri, F.V.; Foschi, F.G.; et al. Immune inflammation indicators and implication for immune modulation strategies in advanced hepatocellular carcinoma patients receiving sorafenib. Oncotarget 2016, 7, 67142–67149. [Google Scholar] [PubMed] [Green Version]
- Argentiero, A.; De Summa, S.; Di Fonte, R.; Iacobazzi, R.M.; Porcelli, L.; Da Vià, M.; Brunetti, O.; Azzariti, A.; Silvestris, N.; Solimando, A.G. Gene Expression Comparison between the Lymph Node-Positive and -Negative Reveals a Peculiar Immune Microenvironment Signature and a Theranostic Role for WNT Targeting in Pancreatic Ductal Adenocarcinoma: A Pilot Study. Cancers 2019, 11, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinyol, R.; Sia, D.; Llovet, J.M. Immune Exclusion-Wnt/CTNNB1 Class Predicts Resistance to Immunotherapies in HCC. Clin. Cancer Res. 2019, 25, 2021–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labeur, T.A.; Ten Cate, D.W.G.; Bart Takkenberg, R.; Azahaf, H.; van Oijen, M.G.H.; van Delden, O.M.; de Man, R.A.; van Vugt, J.L.A.; IJzermans, J.N.M.; Eskens, F.A.L.M.; et al. Are we SHARP enough? The importance of adequate patient selection in sorafenib treatment for hepatocellular carcinoma. Acta Oncol. 2018, 57, 1467–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendry, S.A.; Farnsworth, R.H.; Solomon, B.; Achen, M.G.; Stacker, S.A.; Fox, S.B. The Role of the Tumor Vasculature in the Host Immune Response: Implications for Therapeutic Strategies Targeting the Tumor Microenvironment. Front. Immunol. 2016, 7, 621. [Google Scholar] [CrossRef]
- OLeone, P.; Di Lernia, G.; Solimando, A.G.; Cicco, S.; Saltarella, I.; Lamanuzzi, A.; Ria, R.; Frassanito, M.A.; Ponzoni, M.; Ditonno, P.; et al. Bone marrow endothelial cells sustain a tumor-specific CD8+ T cell subset with suppressive function in myeloma patients. Oncoimmunology 2018, 8, e1486949. [Google Scholar]
- Holmgaard, R.B.; Schaer, D.A.; Li, Y.; Castaneda, S.P.; Murphy, M.Y.; Xu, X.; Inigo, I.; Dobkin, J.; Manro, J.R.; Iversen, P.W.; et al. Targeting the TGFβ pathway with galunisertib, a TGFβRI small molecule inhibitor, promotes anti-tumor immunity leading to durable, complete responses, as monotherapy and in combination with checkpoint blockade. J. Immunother. Cancer. 2018, 6, 47. [Google Scholar] [CrossRef]
- Croucher, P.I.; McDonald, M.M.; Martin, T.J. Bone metastasis: The importance of the neighbourhood. Nat. Rev. Cancer 2016, 16, 373–386. [Google Scholar] [CrossRef]
- Hernanda, P.Y.; Chen, K.; Das, A.M.; Sideras, K.; Wang, W.; Li, J.; Cao, W.; Bots, S.J.; Kodach, L.L.; de Man, R.A.; et al. SMAD4 exerts a tumor-promoting role in hepatocellular carcinoma. Oncogene 2015, 34, 5055–5068. [Google Scholar] [CrossRef] [Green Version]
- Moles, A.; Tarrats, N.; Fernández-Checa, J.C.; Marí, M. Cathepsins B and D drive hepatic stellate cell proliferation and promote their fibrogenic potential. Hepatology 2009, 49, 1297–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manchanda, M.; Das, P.; Gahlot, G.P.S.; Singh, R.; Roeb, E.; Roderfeld, M.; Datta Gupta, S.; Saraya, A.; Pandey, R.M.; Chauhan, S.S. Cathepsin L and B as Potential Markers for Liver Fibrosis: Insights from Patients and Experimental Models. Clin. Transl. Gastroenterol. 2017, 8, e99. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, M.; Kakinuma, S.; Kamiya, A.; Tsunoda, T.; Tsuchiya, J.; Sato, A.; Kaneko, S.; Nitta, S.; Kawai-Kitahata, F.; Murakawa, M.; et al. LIM homeobox 2 promotes interaction between human iPS-derived hepatic progenitors and iPS-derived hepatic stellate-like cells. Sci Rep. 2019, 9, 2072. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Pan, L.H.; Pang, Y.; Yang, J.X.; Lv, M.J.; Liu, F.; Qu, X.F.; Chen, X.X.; Gong, H.J.; Liu, D.; et al. GDF11/BMP11 as a novel tumor marker for liver cancer. Exp. Ther. Med. 2018, 15, 3495–3500. [Google Scholar] [CrossRef] [PubMed]
- Plouhinec, J.L.; Zakin, L.; de Robertis, E.M. Systems control of BMP morphogen flow in vertebrate embryos. Curr. Opin. Genet. Dev. 2011, 21, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Mueller, T.D.; Nickel, J. Promiscuity and specificity in BMP receptor activation. FEBS Lett. 2012, 586, 1846–1859. [Google Scholar] [CrossRef]
- Deng, T.; Lin, D.; Zhang, M.; Zhao, Q.; Li, W.; Zhong, B.; Deng, Y.; Fu, X. Differential expression of bone morphogenetic protein 5 in human lung squamous cell carcinoma and adenocarcinoma (non-small-cell lung cancer (NSCLC)). Acta Biochim. Biophys. Sin. (Shanghai) 2015, 47, 557–563. [Google Scholar] [CrossRef] [Green Version]
- Romagnoli, M.; Belguise, K.; Yu, Z.; Wang, X.; Landesman-Bollag, E.; Seldin, D.C.; Chalbos, D.; Barillé-Nion, S.; Jézéquel, P.; Seldin, M.L.; et al. Epithelial-to-mesenchymal transition induced by TGF-β1 is mediated by Blimp-1-dependent repression of BMP-5. Cancer Res. 2012, 72, 6268–6278. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.S.; Jin, Y.; Cheng, K.I.; Yu, K.L.; Suwa, F.; Nakatsuji, I.; Makigusa, K.; Fang, Y.R. Transcriptional mRNA of bone morphogenetic proteins 2, 3, 4, and 5 in trigeminal nerve, benign and malignant peripheral nerve sheath tumors. Kaohsiung J. Med. Sci. 2001, 17, 16–24. [Google Scholar]
- Johnsen, I.K.; Kappler, R.; Auernhammer, C.J.; Beuschlein, F. Bone morphogenetic proteins 2 and 5 are down-regulated in adrenocortical carcinoma and modulate adrenal cell proliferation and steroidogenesis. Cancer Res. 2009, 69, 5784–5792. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.; Yang, F.; He, H.; Li, Q.; Zhang, W.; Xing, J.; Zhu, Z.; Jiang, J.; Wang, H.; Zhao, X.; et al. Alteration of tumor suppressor BMP5 in sporadic colorectal cancer: A genomic and transcriptomic profiling based study. Mol. Cancer 2018, 17, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.; Li, Q.; Wang, H.; Zhang, P.; Zhao, X.; Yang, F.; Yang, J. MiR-32 promotes tumorigenesis of colorectal cancer by targeting BMP5. Biomed. Pharmacother. 2018, 106, 1046–1051. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, S.; Alarmo, E.L.; Sandström, S.; Ampuja, M.; Kallioniemi, A. Bone morphogenetic protein -4 and -5 in pancreatic cancer—Novel bidirectional players. Exp. Cell Res. 2011, 317, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.; Sanchez, A.; Fabregat, I. BMPs and liver: More questions than answers. Curr. Pharm. Des. 2012, 18, 4114–4125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Tipoe, G.L.; Liong, E.C.; Lau, T.Y.; Fung, P.C.; Leung, K.M. Overexpression of BMP-2/4, -5 and BMPR-IA associated with malignancy of oral epithelium. Oral Oncol. 2001, 37, 225–233. [Google Scholar] [CrossRef]
- Chen, J.; Gingold, J.A.; Su, X. Immunomodulatory TGF-β Signaling in Hepatocellular Carcinoma. Trends Mol. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Swiss-Port TrEMBL Accession Number | Protein Name | MW a | Score | Match Queries | pI b | Sequence Coverage(%) | Matched Peptide |
|---|---|---|---|---|---|---|---|
| Q9CQ60 | 6-phosphogluconolactonase | 27237 | 20 | 3 | 5.55 | 23 | R.FALGLSGGSLVSMLAR.D + Oxidation (M); 2 Phospho (ST) R.ILEDKEGTLPAALVQPR.T + Deamidated (NQ) M.AAPAPSLISVFSSPQELGASLAQLVAQR.A + 2 Deamidated (NQ); 3 Phospho (ST) |
| P10605 | Cathepsin B | 37256 | 60 | 6 | 5.57 | 15 | K.HEAGDMMGGHAIR.I + Oxidation (M) K.HFGYTSYSVSNSVK.E + 4 Phospho (ST); Phospho (Y) K.NGPVEGAFTVFSDFLTYK.S K.KLCGTVLGGPKLPGR.V + Carboxymethyl (C); Phospho (ST) K.NGPVEGAFTVFSDFLTYK.S + Deamidated (NQ) R.DQGSCGSCWAFGAVEAISDR.T + Carboxymethyl (C) |
| Q62356 | Follistatin-related protein 1 | 34532 | 38 | 1 | 5.58 | 2 | K.RPVCGSNGK.T + Carboxymethyl (C); Deamidated (NQ) |
| P06745 | Glucose-6-phosphate isomerase | 62727 | 110 | 3 | 8.14 | 5 | K.YITKSGAR.V + Phospho (ST); Phospho (Y) K.ILLANFLAQTEALMK.G K.INYTENR.A + Deamidated (NQ) |
| P19157 | Glutathione S-transferase P 1 | 23594 | 561 | 10 | 7.68 | 40 | R.MLLADQGQSWK.E M.PPYTIVYFPVR.G K.YVTLIYTNYENGK.N + Deamidated (NQ) R.EAAQMDMVNDGVEDLR.G K.FEDGDLTLYQSNAILR.H R.EAAQMDMVNDGVEDLR.G + Oxidation (M) K.AFLSSPEHVNRPINGNGK.Q + 2 Deamidated (NQ) R.EAAQMDMVNDGVEDLR.G + 2 Deamidated (NQ); Oxidation (M) K.FEDGDLTLYQSNAILR.H + Deamidated (NQ) K.AFLSSPEHVNRPINGNGKQ.- + 2 Deamidated (NQ) |
| Q3UR70 | Transforming growth factor-beta receptor-associated protein 1 | 97241 | 38 | 5 | 6.24 | 5 | K.LKGNAVR.L + Deamidated (NQ) R.DSIHARR.T R.TTQVALGLAK.S R.YPNGGLVHTHCAASR.H + Carboxymethyl (C); Phospho (ST) R.VSTGGKDVEATETQAKLR.R + Deamidated (NQ); 3 Phospho (ST) |
| P54869 | Hydroxymethylglutaryl-CoA synthase | 56786 | 221 | 7 | 8.65 | 21 | K.TPMTSQKTFESLVDFCK.T + Carboxymethyl (C); Deamidated (NQ); Oxidation (M); Phospho (ST) K.TPMTSQK.T + Oxidation (M); 3 Phospho (ST) R.LLVPYLIEAVR.L R.QFLRSMSSSSSASAAAK.K + Deamidated (NQ); Oxidation (M); 2 Phospho (ST) R.QFLRSMSSSSSASAAAK.K + Oxidation (M); 4 Phospho (ST) R.SMSSSSSASAAAKK.I + Oxidation (M); 4 Phospho (ST) R.SMSSSSSASAAAK.K + Oxidation (M) |
| Name | Subcellular Location | Biological Process | Molecular Function | Protein Function |
|---|---|---|---|---|
| 6-phosphogluconolactonase | Carbohydrate metabolic process | |||
| Cathepsin B | Lysosome | Proteolysis epithelial cell differentiation collagen catabolic process | proteoglycan binding collagen binding | Thiol protease may participate in intracellular degradation and protein turnovers in addition to being involved in tumor invasion and metastasis. |
| Follistatin-related protein 1 | Secreted | Response to starvation | heparin binding | It may regulate some growth factors’ activities on cell proliferation and differentiation. |
| Glucose-6-phosphate isomerase | Cytoplasm | Positive regulation of immunoglobulin secretion response to immobilization stress | cytokine activity monosaccharide binding ubiquitin protein ligase binding | It is a glycolytic enzyme. It can also function as a tumor-secreted cytokine and an angiogenic factor (AMF) to stimulate endothelial cell motility. |
| Glutathione S-transferase P 1 | Cytoplasm Mitochondrion Nucleus | Negative regulation of fibroblast proliferationcellular response to cell-matrix adhesion | JUN kinase binding kinase regulator activity | |
| Transforming growth factor-beta receptor-associated protein 1 | Cytoplasm | Regulation of transcription | SMAD binding | It is involved in the TGF-beta/activin signaling pathway and linked to complexes of inactive heteromeric TGF-beta and activin receptor through the type II receptor. It can be released upon signaling activation. SMAD4 may play a role in the receptor complex and facilitate its interaction with receptor-regulated Smads, such as SMAD2. |
| Hydroxymethylglutaryl-CoA synthase | Mitochondrion | Cholesterol biosynthetic process |
| Swiss-port TrEMBL Accession Number | Protein Name | MW | Score | Match Queries | pI | Sequence Coverage (%) | Matched Peptide |
|---|---|---|---|---|---|---|---|
| Q9CQ60 | 6-phosphogluconolactonase | 27237 | 20 | 3 | 5.55 | 23 | R.FALGLSGGSLVSMLAR.D + Oxidation (M); 2 Phospho (ST) R.ILEDKEGTLPAALVQPR.T + Deamidated (NQ) M.AAPAPSLISVFSSPQELGASLAQLVAQR.A + 2 Deamidated (NQ); 3 Phospho (ST) |
| P62897 | Cytochrome c | 11598 | 71 | 2 | 9.61 | 16 | K.GITWGEDTLMEYLENPK.K K.TGQAAGFSYTDANK.N |
| O08749 | Dihydrolipoyl dehydrogenase | 54238 | 63 | 1 | 7.99 | 3 | K.IPNIYAIGDVVAGPMLAHK.A |
| O35382 | Exocyst complex component 4 | 110475 | 22 | 5 | 6.07 | 9 | R.LKEIICEQAAIK.Q + Carboxymethyl (C); Deamidated (NQ) K.VLGVQRPLLQSTIIVEK.T + Deamidated (NQ) -MAAEAAGGKYRSTVSK.S + Oxidation (M); 3 Phospho (ST); Phospho (Y) K.DISAMEEAMSASLQQHK.F + Deamidated (NQ); Oxidation (M); 3 Phospho (ST) K.QCPLREFLTVYIKSIFLNQVLAEINK.E + Phospho (Y) |
| P06745 | Glucose-6-phosphate isomerase | 62727 | 110 | 3 | 8.14 | 5 | K.YITKSGAR.V + Phospho (ST); Phospho (Y) K.ILLANFLAQTEALMK.G K.INYTENR.A + Deamidated (NQ) |
| P11352 | Glutathione peroxidase 1 | 22316 | 42 | 4 | 6.74 | 18 | K.YVRPGGGFEPNFTLFEK.C K.FLVGPDGVPVR.R R.DYTEMNDLQK.R + Oxidation (M); Phospho (ST); Phospho (Y) R.NALPTPSDDPTALMTDPK.Y + Deamidated (NQ); Oxidation (M); Phospho (ST) |
| P97494 | Glutamate--cysteine ligase catalytic subunit | 72525 | 44 | 4 | 5.59 | 6 | K.SLFFPDEAINK.H + Deamidated (NQ); Phospho (ST) -.MGLLSQGSPLSWEETQR.H + Deamidated (NQ); Oxidation (M) K.RASGELMTVAR.W + Oxidation (M); Phospho (ST) K.VQLLLNGGDVLETLQEKGER.T + Phospho (ST) |
| P19157 | Glutathione S-transferase P 1 | 23594 | 561 | 10 | 7.68 | 40 | R.MLLADQGQSWK.E M.PPYTIVYFPVR.G K.YVTLIYTNYENGK.N + Deamidated (NQ) R.EAAQMDMVNDGVEDLR.G K.FEDGDLTLYQSNAILR.H R.EAAQMDMVNDGVEDLR.G + Oxidation (M) K.AFLSSPEHVNRPINGNGK.Q + 2 Deamidated (NQ) R.EAAQMDMVNDGVEDLR.G + 2 Deamidated (NQ); Oxidation (M) K.FEDGDLTLYQSNAILR.H + Deamidated (NQ) K.AFLSSPEHVNRPINGNGKQ.- + 2 Deamidated (NQ) |
| P62983 | Ubiquitin-40S ribosomal protein S27a | 17939 | 52 | 6 | 9.68 | 26 | R.TLSDYNIQK.E K.IQDKEGIPPDQQR.L + 2 Deamidated (NQ) K.TITLEVEPSDTIENVK.A K.KSYTTPK.K + Phospho (ST) R.TLSDYNIQK.E R.TLSDYNIQKESTLHLVLR.L + Deamidated (NQ); 2 Phospho (ST); Phospho (Y) |
| Q3UR70 | Transforming growth factor-beta receptor-associated protein 1 | 97241 | 38 | 5 | 6.24 | 5 | K.LKGNAVR.L + Deamidated (NQ) R.DSIHARR.T R.TTQVALGLAK.S R.YPNGGLVHTHCAASR.H + Carboxymethyl (C); Phospho (ST) R.VSTGGKDVEATETQAKLR.R + Deamidated (NQ); 3 Phospho (ST) |
| Q9JIQ8 | Transmembrane protease serine 2 | 53491 | 27 | 2 | 8.1 | 4 | R.QSLMFYGSR.H + Deamidated (NQ); Oxidation (M); Phospho (ST) K.SSGALCTSKSK.K + Carboxymethyl (C); 2 Phospho (ST) |
| Name | Subcellular Location | Biological Process | Molecular Function | Function |
|---|---|---|---|---|
| 6-phosphogluconolactonase | carbohydrate metabolic process | |||
| Cytochrome c | Mitochondrion | apoptotic process hydrogen peroxide metabolic process mitochondrial electron transport | electron transporter heme binding | This protein has a role in apoptosis. If suppresses the anti-apoptotic members or activates the pro-apoptotic members of the Bcl-2 family, this will lead to altered mitochondrial membrane permeability and result in releasing cytochrome c into the cytosol. When cytochrome c binds to Apaf-1, the activation of caspase-9 will be triggered and then accelerating apoptosis by activating other caspases |
| Dihydrolipoyl dehydrogenase | Mitochondrion matrix Nucleus | mitochondrial electron transport cell redox homeostasis lipoate metabolic process regulation of membrane potential | electron transfer activity | |
| Exocyst complex component 4 | chemical synaptic transmission protein targeting to membrane | Ral GTPase binding protein N-terminus binding | It is part of the exocyst complex involved in the docking of exocytic vesicles and may have fusion sites on the plasma membrane. | |
| Glucose-6-phosphate isomerase | Cytoplasm | positive regulation of immunoglobulin secretion response to immobilization stress | cytokine activity monosaccharide binding ubiquitin protein ligase binding | It is a glycolytic enzyme. It can also function as a tumor-secreted cytokine and an angiogenic factor (AMF) to stimulate endothelial cell motility. |
| Glutathione peroxidase 1 | Cytoplasm | response to hydroperoxide regulation of neuron apoptotic process protein oxidation triglyceride metabolic process | SH3 domain binding | The oxidative breakdown to the hemoglobin in erythrocytes can be protected by this protein. |
| Glutamate--cysteine ligase catalytic subunit | Cytosol | cysteine metabolic process negative regulation of extrinsic apoptotic signaling pathway negative regulation of hepatic stellate cell activation | ADP binding ATP binding coenzyme binding | |
| Glutathione S-transferase P 1 | Cytoplasm Mitochondrion Nucleus | negative regulation of fibroblast proliferationcellular response to cell-matrix adhesion | JUN kinase binding kinase regulator activity | |
| Ubiquitin-40S ribosomal protein S27a | Cytoplasm Nucleus | translation | metal ion binding structural constituent of ribosome | |
| Transforming growth factor-beta receptor-associated protein 1 | Cytoplasm | regulation of transcription | SMAD binding | It is involved in the TGF-beta/activin signaling pathway and linked to complexes of inactive heteromeric TGF-beta and activin receptor through the type II receptor. It can be released upon signaling activation. SMAD4 may play a role in the receptor complex and facilitate its interaction with receptor-regulated Smads, such as SMAD2. |
| Transmembrane protease serine 2 | Cell membrane | protein autoprocessing | scavenger receptor activity | It proteolytically cleaves and activates the glycoproteins of viral spikes and then facilitates virus-cell membrane fusions. Because such spike proteins are synthesized and maintained in the intermediate folding states, this proteolysis permits the refolding and energy release. As a result, this creates stable virus-cell linkages and membrane coalescence. |
| Swiss-port TrEMBL Accession Number | Protein Name | MW | Score | Match Queries | pI | Sequence Coverage(%) | Matched Peptide |
|---|---|---|---|---|---|---|---|
| P11352 | Glutathione peroxidase 1 | 22316 | 42 | 4 | 6.74 | 18 | K.YVRPGGGFEPNFTLFEK.C K.FLVGPDGVPVR.R R.DYTEMNDLQK.R + Oxidation (M); Phospho (ST); Phospho (Y) R.NALPTPSDDPTALMTDPK.Y + Deamidated (NQ); Oxidation (M); Phospho (ST) |
| P16627 | Heat shock 70 kDa protein 1-like | 70593 | 98 | 15 | 5.91 | 12 | K.VMVSYKGEK.K + Phospho (Y) K.VMVSYKGEK.K + Oxidation (M); Phospho (Y) K.VEIIANDQGNR.T R.KELENMCNPIITKLYQSGCTGPTCTPGYTPGR.A + 2 Carboxymethyl (C); 2 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); Phospho (Y) K.AFYPEEISSMVLTKMK.E + Phospho (ST) R.KELENMCNPIITKLYQSGCTGPTCTPGYTPGR.A + 2 Deamidated (NQ); Oxidation (M); 5 Phospho (ST); 2 Phospho (Y) K.MDKAKIHDIVLVGGSTR.I + Oxidation (M); Phospho (ST) K.LWPFQVINEAGKPKVMVSYKGEK.K + 2 Deamidated (NQ); Oxidation (M); Phospho (Y) K.VMVSYKGEK.K + Phospho (ST) K.ITITNDKGRLSK.E + Deamidated (NQ); 2 Phospho (ST) K.NQVAMNPQNTVFDAK.R + Phospho (ST) K.NALESYAFNMK.S + Deamidated (NQ); Oxidation (M); Phospho (ST); Phospho (Y) R.TTPSYVAFTDTER.L K.AFYPEEISSMVLTK.M + Oxidation (M) K.SINPDEAVAYGAAVQAAILMGDK.S + Phospho (ST) |
| P11499 | Heat shock protein HSP 90-beta | 83229 | 175 | 21 | 4.97 | 18 | R.YESLTDPSK.L K.YIDQEELNK.T K.ADLINNLGTIAK.S R.ELISNASDALDK.I K.EDQTEYLEER.R K.EISDDEAEEEK.G + Phospho (ST) K.IEDVGSDEEDDSGK.D R.GVVDSEDLPLNISR.E K.NIVKKCLELFSELAEDK.E + Carboxymethyl (C); Phospho (ST) K.VTISNRLVSSPCCIVTSTYGWTANMER.I + 2 Carboxymethyl (C); 2 Deamidated (NQ); Oxidation (M); 3 Phospho (ST); Phospho (Y) R.EMLQQSK.I + Phospho (ST) K.LMKEILDKK.V R.DNSTMGYMMAK.K + Oxidation (M) K.AQALRDNSTMGYMMAK.K + 2 Deamidated (NQ); 2 Oxidation (M); Phospho (ST) K.VTISNRLVSSPCCIVTSTYGWTANMER.I + Oxidation (M); Phospho (ST) K.FENLCKLMK.E R.IMKAQALRDNSTMGYMMAK. K + Deamidated (NQ); 3 Oxidation (M); Phospho (ST) R.TLTLVDTGIGMTKADLINNLGTIAK.S + Oxidation (M); 4 Phospho (ST) R.TLTLVDTGIGMTK.A + Oxidation (M); 2 Phospho (ST) K.IEDVGSDEEDDSGK.D + Phospho (ST) K.AQALRDNSTMGYMMAKK.H + Deamidated (NQ); Oxidation (M); 2 Phospho (ST); Phospho (Y) |
| P63017 | Heat shock cognate 71 kDa protein | 70827 | 101 | 20 | 5.37 | 16 | K.VEIIANDQGNR.T K.STAGDTHLGGEDFDNR.M K.STGKENKITITNDK.G + Deamidated (NQ); 3 Phospho (ST) K.NSLESYAFNMKATVEDEK.L + Oxidation (M); Phospho (ST) K.NSLESYAFNMKATVEDEK.L + Deamidated (NQ); Oxidation (M); Phospho (Y) K.SINPDEAVAYGAAVQAAILSGDK.S K.TVTNAVVTVPAYFNDSQRQATK.D + 4 Deamidated (NQ); Phospho (ST) M.SKGPAVGIDLGTTYSCVGVFQHGK.V + 2 Phospho (ST); Phospho (Y) M.SKGPAVGIDLGTTYSCVGVFQHGK.V + Deamidated (NQ); 2 Phospho (ST); Phospho (Y) K.LQGKINDEDKQK.I + 2 Deamidated (NQ) K.STGKENKITITNDK.G + 3 Phospho (ST) K.ITITNDKGRLSK.E + Deamidated (NQ); 2 Phospho (ST) K.TVTNAVVTVPAYFNDSQR.Q + 3 Deamidated (NQ); 4 Phospho (ST) K.EIAEAYLGK.T + Phospho (Y) K.QTQTFTTYSDNQPGVLIQVYEGER.A + 3 Deamidated (NQ); Phospho (ST) K.EIAEAYLGK.T K.LLQDFFNGK.E + 2 Deamidated (NQ) R.AMTKDNNLLGK.F + 2 Deamidated (NQ); Phospho (ST) M.SKGPAVGIDLGTTYSCVGVFQHGK.V + Deamidated (NQ); 3 Phospho (ST); Phospho (Y) R.TTPSYVAFTDTER.L |
| Q3UR70 | Transforming growth factor-beta receptor-associated protein 1 | 97241 | 38 | 5 | 6.24 | 5 | K.LKGNAVR.L + Deamidated (NQ) R.DSIHARR.T R.TTQVALGLAK.S R.YPNGGLVHTHCAASR.H + Carboxymethyl (C); Phospho (ST) R.VSTGGKDVEATETQAKLR.R + Deamidated (NQ); 3 Phospho (ST) |
| Q9JIQ8 | Transmembrane protease serine 2 | 53491 | 27 | 2 | 8.1 | 4 | R.QSLMFYGSR.H + Deamidated (NQ); Oxidation (M); Phospho (ST) K.SSGALCTSKSK.K + Carboxymethyl (C); 2 Phospho (ST) |
| P54869 | Hydroxymethylglutaryl-CoA synthase | 56786 | 221 | 7 | 8.65 | 21 | K.TPMTSQKTFESLVDFCK.T + Carboxymethyl (C); Deamidated (NQ); Oxidation (M); Phospho (ST) K.TPMTSQK.T + Oxidation (M); 3 Phospho (ST) R.LLVPYLIEAVR.L R.QFLRSMSSSSSASAAAK.K + Deamidated (NQ); Oxidation (M); 2 Phospho (ST) R.QFLRSMSSSSSASAAAK.K + Oxidation (M); 4 Phospho (ST) R.SMSSSSSASAAAKK.I + Oxidation (M); 4 Phospho (ST) R.SMSSSSSASAAAK.K + Oxidation (M) |
| Name | Subcellular Location | Biological Process | Molecular Function | Function |
|---|---|---|---|---|
| Glutathione peroxidase 1 | Cytoplasm | Response to hydroperoxide regulation of neuron apoptotic process protein oxidation triglyceride metabolic process | SH3 domain binding | Protects the hemoglobin in erythrocytes from oxidative breakdown. |
| Heat shock 70 kDa protein 1-like | Mitochondrion Nucleus | Cell differentiation protein refolding | ATP binding unfolded protein binding | It may play a pivotal role in the protein quality control system, ensuring the correct folding of proteins, the re-folding of misfolded proteins and controlling the targeting of proteins for subsequent degradation. |
| Heat shock protein HSP 90-beta | Cell membrane Cytoplasm Membrane Nucleus | Positive regulation of telomerase activity positive regulation of protein binding | ATP binding GTP binding protein kinase binding | Molecular chaperone that promotes the maturation, structural maintenance and proper regulation of specific target proteins involved for instance in cell cycle control and signal transduction. |
| Heat shock cognate 71 kDa protein | Cell membrane Cytoplasm Nucleus | ATP metabolic process regulation of cell cycle protein folding | ATP binding enzyme binding | Molecular chaperone implicated in a wide variety of cellular processes, including protection of the proteome from stress, folding and transport of newly synthesized polypeptides, activation of proteolysis of misfolded proteins and the formation and dissociation of protein complexes. Plays a pivotal role in the protein quality control system, ensuring the correct folding of proteins, the re-folding of misfolded proteins and controlling the targeting of proteins for subsequent degradation. This is achieved through cycles of ATP binding, ATP hydrolysis and ADP release, mediated by co-chaperones. |
| Transforming growth factor-beta receptor-associated protein 1 | Cytoplasm | Regulation of transcription | SMAD binding | It may plays a role in the TGF-beta/activin signaling pathway. It associates with inactive heteromeric TGF-beta and activin receptor complexes, mainly through the type II receptor, and is released upon activation of signaling. May recruit SMAD4 to the vicinity of the receptor complex and facilitate its interaction with receptor-regulated Smads, such as SMAD2. |
| Transmembrane protease serine 2 | Cell membrane | Protein autoprocessing | scavenger receptor activity | Serine protease that proteolytically cleaves and activates the viral spike glycoproteins which facilitate virus-cell membrane fusions. The spike proteins are synthesized and maintained in precursor intermediate folding states and proteolysis permits the refolding and energy release required to create stable virus-cell linkages and membrane coalescence. |
| Hydroxymethylglutaryl-CoA synthase | Mitochondrion | Cholesterol biosynthetic process | hydroxymrthylglutaryl-CoA synthase activity | This enzyme condenses acetyl-CoA with acetoacetyl-CoA to form HMG-CoA, which is the substrate for HMG-CoA reductase. |
| Serial No. | Chromosome No. | Gene Name | Gene ID | Chromosome Position | Transcript ID | Protein Class | Protein Evidence | Matched Sequence |
|---|---|---|---|---|---|---|---|---|
| 1 | 7 | AC068533.7 | ENSG00000249319 | 66087761–66152277 | ENST00000450043 | Predicted intracellular proteins | Evidence at transcript level | RCAGLLMTLKGLPSTYNKD |
| RQAHEASGKA | ||||||||
| KNPDSLELIR | ||||||||
| RCAGLLMTLK | ||||||||
| RDFVAEFLFWASLCMTHLSR | ||||||||
| 2 | 17 | BAHCC1-201 | ENSG00000266074 | 1399740–81466332 | ENST00000307745 | Predicted intracellular proteins | Evidence at transcript level | KHLTSCLLNTKV |
| SPTPPPR | ||||||||
| RSPARRGPGRPRK | ||||||||
| KLDHEGVTSPKN | ||||||||
| KHLTSCLLNTK | ||||||||
| KWSGNPTQR | ||||||||
| KWSGNPTQRR | ||||||||
| RRGSPLLSWSAVAQTK | ||||||||
| RSPARRGPGRPR | ||||||||
| KEALSFSKAKELSR | ||||||||
| KERQGLLGACR | ||||||||
| KLDHEGVTSPK | ||||||||
| 3 | 4 | CCSER1-003 | ENSG00000184305 | 90127535–90923530 | ENST00000505073 | Predicted intracellular proteins | Evidence at transcript level | SSSEGTAGSSR |
| ITRSLTEDFEREK | ||||||||
| TNSPR | ||||||||
| LRSSSEGTAGSS | ||||||||
| SSSEGTAGSS | ||||||||
| 4 | 13 | KLHL1 | ENSG00000150361 | 69700594–70108493 | ENST00000377844 | Predicted intracellular proteins | Evidence at transcript level | KSTVGTLYAVGGMDNNKGATTIEK |
| STVGTLYAVGGMDNNK | ||||||||
| LQQGAPGQGTQQPA | ||||||||
| NDLECQ | ||||||||
| 5 | 3 | KLHL24-001 | ENSG00000114796 | 183635568–183684477 | ENST00000242810 | Predicted intracellular proteins | Evidence at transcript level | DVWIYNSQLNIWIR |
| INSRDVWIYNSQLNIWIR | ||||||||
| QENCGMSVCNG | ||||||||
| VQSYDPETNSWLL | ||||||||
| 6 | 12 | RP11-162P23.2 | ENSG00000257767 | 111753890–111791418 | ENST00000546840 | Predicted intracellular proteins | Evidence at transcript level | RTYLAALETLDNGKPYVISYLVDLDMVLKC |
| KTIPIDGDFFSYTRH | ||||||||
| KLGPALATGNVVVMKV | ||||||||
| KVAEQTPLTALYVANLIKE | ||||||||
| RAAFQLGSPWRR | ||||||||
| KLGPALATGNVVVMK | ||||||||
| KTIPIDGDFFSYTR | ||||||||
| KVAEQTPLTALYVANLIK | ||||||||
| RAAFQLGSPWR | ||||||||
| RLADLIER | ||||||||
| KTIPIDGDFFSYTR | ||||||||
| 7 | 17 | SRCIN1-001 | ENSG00000277363 | 38530016–38605930 | ENST00000617146 | Predicted intracellular proteins | Evidence at protein level | MEAMEKQIASLTGLVQSALLR |
| TGEVVVTSK | ||||||||
| SRLSYAGGRPPSYAGSPVHHAAER | ||||||||
| RCRGVTDTLAQI | ||||||||
| REMVYAS | ||||||||
| 8 | 11 | ADAMTS15 | ENSG00000166106 | 130448974–130476641 | ENST00000299164 | Enzymes, predicted secreted proteins | Evidence at transcript level | SFREEQCEAFNGYNHSTNR |
| EEQCEAFNGYNHSTNR | ||||||||
| 9 | 12 | ANO4-002 | ENSG00000151572 | 100717526–101128641 | ENST00000392977 | Predicted intracellular proteins, predicted membrane proteins | Evidence at transcript level | DLRDRMR |
| LINRWR | ||||||||
| 10 | 6 | BMP5 | ENSG00000112175 | 55753645–55875564 | ENST00000370830 | Predicted intracellular proteins | Evidence at transcript level | LNAISVLYFDDSSNVILK |
| TPLTTQSPPLASLHDTNFLNDADMVMSFVNLVER | ||||||||
| 11 | 11 | CCDC67-002 | ENSG00000165325 | 93329990–93415200 | ENST00000530862 | Predicted intracellular proteins, predicted membrane proteins | Evidence at protein level | SELQSRDDLLR |
| MYQRQCQAMEAGLSEVK | ||||||||
| 12 | 3 | FEZF2-004 | ENSG00000153266 | 62369681–62374324 | ENST00000486811 | Predicted intracellular proteins, transcription factors | Evidence at transcript level | IIHTQEKPHKCNQCGK |
| KLLNLCSPLPCMIPLQPLGYEVPSK | ||||||||
| 13 | 15 | GABRG3-001 | ENSG00000182256 | 26971282–27541991 | ENST00000615808 | FDA approved drug targets Predicted intracellular proteins, predicted membrane proteins Predicted secreted proteins | Evidence at protein level | IWNDGK |
| KNSVEAADQK | ||||||||
| 14 | 12 | GDF11-001 | ENSG00000135414 | 55743280–55757278 | ENST00000257868 | Predicted secreted proteins | Evidence at protein level | RNLGLDCDEHSSESR |
| SLKIELHSR | ||||||||
| 15 | 13 | HS6ST3 | ENSG00000185352 | 96090839–96839562 | ENST00000376705 | Predicted intracellular proteins, predicted secreted proteins | Evidence at transcript level | HLVKNIR |
| RHLVKNIRL | ||||||||
| 16 | 23X | KIAA2022-001 | ENSG00000050030 | 74732849–74925447 | ENST00000055682 | Disease related genes, predicted intracellular proteins | Evidence at transcript level | DCSRYMARDTNSGSSSSQQNYGLR |
| YMARDTNSGSSSSQQNYGLR | ||||||||
| APFAIMEPAGMSALNGDCLMQPSRTCLGCFMESK | ||||||||
| HKSSSK | ||||||||
| 17 | 9 | LHX2-004 | ENSG00000106689 | 124001670–124015361 | ENST00000560961 | Predicted intracellular proteins, transcription factors | Evidence at transcript level | CARCHLGISASEMVMR |
| CHLGISASEMVMRAR | ||||||||
| 18 | 1 | LMX1A-001 | ENSG00000162761 | 165201867–165356715 | ENST00000342310 | Predicted intracellular proteins, transcription factors | Evidence at protein level | EPLETTCFYRDKK |
| KVRETLAAETGLSVR | ||||||||
| 19 | 19 | LRFN3-001 | ENSG00000126243 | 35935550–35945767 | ENST00000588831 | Predicted intracellular proteins, transcription factors | Evidence at transcript level | LDMTSNR |
| LARLDMTSNR | ||||||||
| LDMTSNRLTTIPPDPLFSRLPLLARPR | ||||||||
| 20 | 6 | MSH5-SAPCD1-001 | ENSG00000255152 | 31740020–31764851 | ENST00000493662 | Plasma proteins, predicted intracellular proteins, predicted secreted proteins | Evidence at protein level | DQETLLMYQLQCQVLAR |
| IHSCESISLGLSTFMIDLNQVAK | ||||||||
| 21 | 2 | OSR1 | ENSG00000143867 | 19351485–19358653 | ENST00000272223 | Enzymes, predicted intracellular proteins | Evidence at protein level | EFVCKFCGR |
| TSKIKC | ||||||||
| GFCQSRTLAVHKTLHSQVK | ||||||||
| 22 | 17 | RASL10B | ENSG00000270885 | 35731649–35743521 | ENST00000603017 | Predicted intracellular proteins | Evidence at protein level | KTWKCGYVECSAK |
| WNVSHLVRKTWK | ||||||||
| 23 | 2 | RNF103-CHMP3-002 | ENSG00000249884 | 86505668–86721122 | ENST00000604011 | Predicted intracellular proteins | Evidence at transcript level | NQLAVLRVAGSLQK |
| AHMNSVLMGMKNQLAVLR | ||||||||
| 24 | 12 | RP11-834C11.12 | ENSG00000273049 | 53985845–54034888 | ENST00000513209 | Predicted intracellular proteins | Evidence at transcript level | TSYTRYQTLELEK |
| SDSQTPSPNEIK | ||||||||
| 25 | 2 | SP5 | ENSG00000204335 | 170715351–170718078 | ENST00000375281 | Predicted intracellular proteins, transcription factors | Evidence at transcript level | NDSLQAFLQDRTPSASPDLGK |
| FACPECGK | ||||||||
| 26 | 2 | SP9 | ENSG00000217236 | 174334946–174338492 | ENST00000394967 | Predicted intracellular proteins, transcription factors | Evidence at transcript level | THNGGGGGKKGSDSDTDASNLETPR |
| LGPAGASLRRK | ||||||||
| RYSGRATCDCPNCQEAER | ||||||||
| MATSILGEEPRFGTTPLAMLAATCNK | ||||||||
| 27 | 1 | TSSK3-001 | ENSG00000162526 | 32362197–32364312 | ENST00000373534 | Enzymes, predicted intracellular proteins | Evidence at protein level | MEDFLLSNGYQLGKTIGEGTYSK |
| TLDHKNIIQVYEMLESADGK | ||||||||
| 28 | 23X | USP27X | ENSG00000273820 | 49879948–49882565 | ENST00000621775 | Enzymes, predicted intracellular proteins | Evidence at transcript level | CGSCQSYQESTK |
| ITTYISFPLELDMTPFMASSK | ||||||||
| 29 | 23Y | USP9Y | ENSG00000279327 | 12709448–12859416 | ENST00000625141 | Disease related genes, enzymes, potential drug targets, predicted intracellular proteins | Evidence at protein level | FFRDGLTISFTK |
| ASWTNASKK | ||||||||
| LYSVVSQLIR | ||||||||
| MDDDEEMK |
| UniProtKBEntry | Gene Name | UniProt Protein Name | Subcellular Location | Biological Process | Molecular Function | Protein Function |
|---|---|---|---|---|---|---|
| Q7M6Y5 | CCDC67-002 | Deuterosome assembly protein 1 | Cytoplasm | Cilium biogenesis/degradation | Identical protein binding | Key structural component of the deuterosome, a structure that promotes de novo centriole amplification in multiciliated cells. Deuterosome-mediated centriole amplification occurs in terminally differentiated multiciliated cells and can generate more than 100 centrioles. Probably sufficient for the specification and formation of the deuterosome inner core. Interacts with CEP152 and recruits PLK4 to activate centriole biogenesis. |
| Q9Z1W4 | GDF11-001 | Growth/differentiation factor 11 | Secreted | Animal organ morphogenesis, cell development, negative regulation of cell differentiation | Cytokine, Growth factor | Secreted signal that acts globally to specify positional identity along the anterior/posterior axis during development (PubMed:10391213). May play critical roles in patterning both mesodermal and neural tissues and in establishing the skeletal pattern. Signals through activin receptors type-2, ACVR2A and ACVR2B, and activin receptors type-1, ACVR1B, ACVR1C and TGFBR1 leading to the phosphorylation of SMAD2 and SMAD3 (PubMed:16845371, PubMed:12414726). |
| Q9Z0S2 | LHX2-004 | LIM/homeobox protein Lhx2 | Nucleus | Activator, DNA-binding | Transcription, Transcription regulation | Acts as a transcriptional activator. Stimulates the promoter of the alpha-glycoprotein gene. Transcriptional regulatory protein involved in the control of cell differentiation in developing lymphoid and neural cell types. |
| P49003 | BMP5 | Bone morphogenetic protein 5 | Secreted | Chondrogenesis, Differentiation, Osteogenesis | Cytokine, Developmental protein, Growth factor | Induces cartilage and bone formation. |
| Q9ESP5 | FEZF2-004 | Fez family zinc finger protein 2 | Nucleus | Differentiation, Neurogenesis, Transcription, Transcription regulation | Developmental protein, DNA-binding, Repressor | Transcription repressor. Required for the specification of corticospinal motor neurons and other subcerebral projection neurons. May play a role in layer and neuronal subtype-specific patterning of subcortical projections and axonal fasciculation. Controls the development of dendritic arborization and spines of large layer V pyramidal neurons. Plays a role in rostro-caudal patterning of the diencephalon and in prethalamic formation. |
| Q9QYK4 | HS6ST3 | Heparan-sulfate 6-O-sulfotransferase 3 | Membrane Single-pass type II membrane protein | Blastocyst hatching, glycosaminoglycan biosynthetic process | Transferase | 6-O-sulfation enzyme which catalyzes the transfer of sulfate from3’-phosphoadenosine 5’-phosphosulfate (PAPS) to position 6 of the N-sulfoglucosamine residue (GlcNS) of heparan sulfate. |
| Q5DTT1 | KIAA2022-001 | Neurite extension and migration factor | Nucleus Cytoplasm | Neurogenesis, Transcription, Transcription regulation | Developmental protein | Involved in neurite outgrowth by regulating cell-cell adhesion via the N-cadherin signaling pathway. May act by regulating expression of protein-coding genes, such as N-cadherins and integrin beta-1 (ITGB1). |
| Q9JHX2 | SP5 | Transcription factor Sp5 | Nucleus | Transcription, Transcription regulation | Activator, DNA-binding | Binds to GC boxes promoters elements. Probable transcriptional activator that has a role in the coordination of changes in transcription required to generate pattern in the developing embryo. |
| P59384 | ADAMTS15 | A disintegrin and metalloproteinase with thrombospondin motifs 15 | extracellular marix | Metalloendopeptidase activity | Hydrolase, Metalloprotease, Protease | This gene encodes a member of the ADAMTS (a disintegrin and metalloproteinase with thrombospondin motifs) protein family. ADAMTS family members share several distinct protein modules, including a propeptide region, a metalloproteinase domain, a disintegrin-like domain, and a thrombospondin type 1 (TS) motif. Individual members of this family differ in the number of C-terminal TS motifs, and some have unique C-terminal domains. The encoded preproprotein is proteolytically processed to generate the mature enzyme, which may play a role in versican processing during skeletal muscle development. This gene may function as a tumor suppressor in colorectal and breast cancers. |
| Q9WVG7 | OSR1 | Protein odd-skipped-related 1 | Nucleus | Transcription, Transcription regulation | Developmental protein, DNA-binding | Transcription factor that plays a role in the regulation of embryonic heart and urogenital development. |
| Q8CEG8 | USP27X | Ubiquitin carboxyl-terminal hydrolase 27 | Cytosol Nucleus | Ubl conjugation pathway | Hydrolase, Protease, Thiol protease | Deubiquitinase that can reduce the levels of BCL2L11/BIM ubiquitination and stabilize BCL2L11 in response to the RAF-MAPK-degradation signal. By acting on BCL2L11 levels, may counteract the anti-apoptotic effects of MAPK activity. |
| Q9JKU8 | LMX1A-001 | LIM homeobox transcription factor 1-alpha | Nucleus | Activator, Developmental protein, DNA-binding | Transcription, Transcription regulation | Acts as a transcriptional activator by binding to an A/T-rich sequence, the FLAT element, in the insulin gene promoter. Required for development of the roof plate and, in turn, for specification of dorsal cell fates in the CNS and developing vertebrae. |
| Q9Z0S2 | LHX2-004 | LIM/homeobox protein Lhx2 | Nucleus | Transcription, Transcription regulation | Activator, DNA-binding | Acts as a transcriptional activator. Stimulates the promoter of the alpha-glycoprotein gene. Transcriptional regulatory protein involved in the control of cell differentiation in developing lymphoid and neural cell types. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.-H.; Chen, M.; Mo, H.-H.; Tsai, W.-C.; Chang, Y.-C.; Chang, C.-C.; Chen, K.-C.; Wu, H.-Y.; Yuan, C.-H.; Lee, C.-H.; et al. Utilizing Experimental Mouse Model to Identify Effectors of Hepatocellular Carcinoma Induced by HBx Antigen. Cancers 2020, 12, 409. https://doi.org/10.3390/cancers12020409
Yang M-H, Chen M, Mo H-H, Tsai W-C, Chang Y-C, Chang C-C, Chen K-C, Wu H-Y, Yuan C-H, Lee C-H, et al. Utilizing Experimental Mouse Model to Identify Effectors of Hepatocellular Carcinoma Induced by HBx Antigen. Cancers. 2020; 12(2):409. https://doi.org/10.3390/cancers12020409
Chicago/Turabian StyleYang, Ming-Hui, Marcelo Chen, Hsiao-Hsuan Mo, Wan-Chi Tsai, Yu-Chi Chang, Chin-Chuan Chang, Ko-Chin Chen, Hsin-Yi Wu, Cheng-Hui Yuan, Che-Hsin Lee, and et al. 2020. "Utilizing Experimental Mouse Model to Identify Effectors of Hepatocellular Carcinoma Induced by HBx Antigen" Cancers 12, no. 2: 409. https://doi.org/10.3390/cancers12020409