Effects of Andrographolide on Intracellular pH Regulation, Cellular Migration, and Apoptosis in Human Cervical Cancer Cells (Running Tittle: Effects of Andrographolide on pH Regulators and Apoptosis in Cervical Cancer)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
2.1. Resting and New Steady-State Intracellular pH Value of Cultured Cells of HeLa, End1, and Ect1
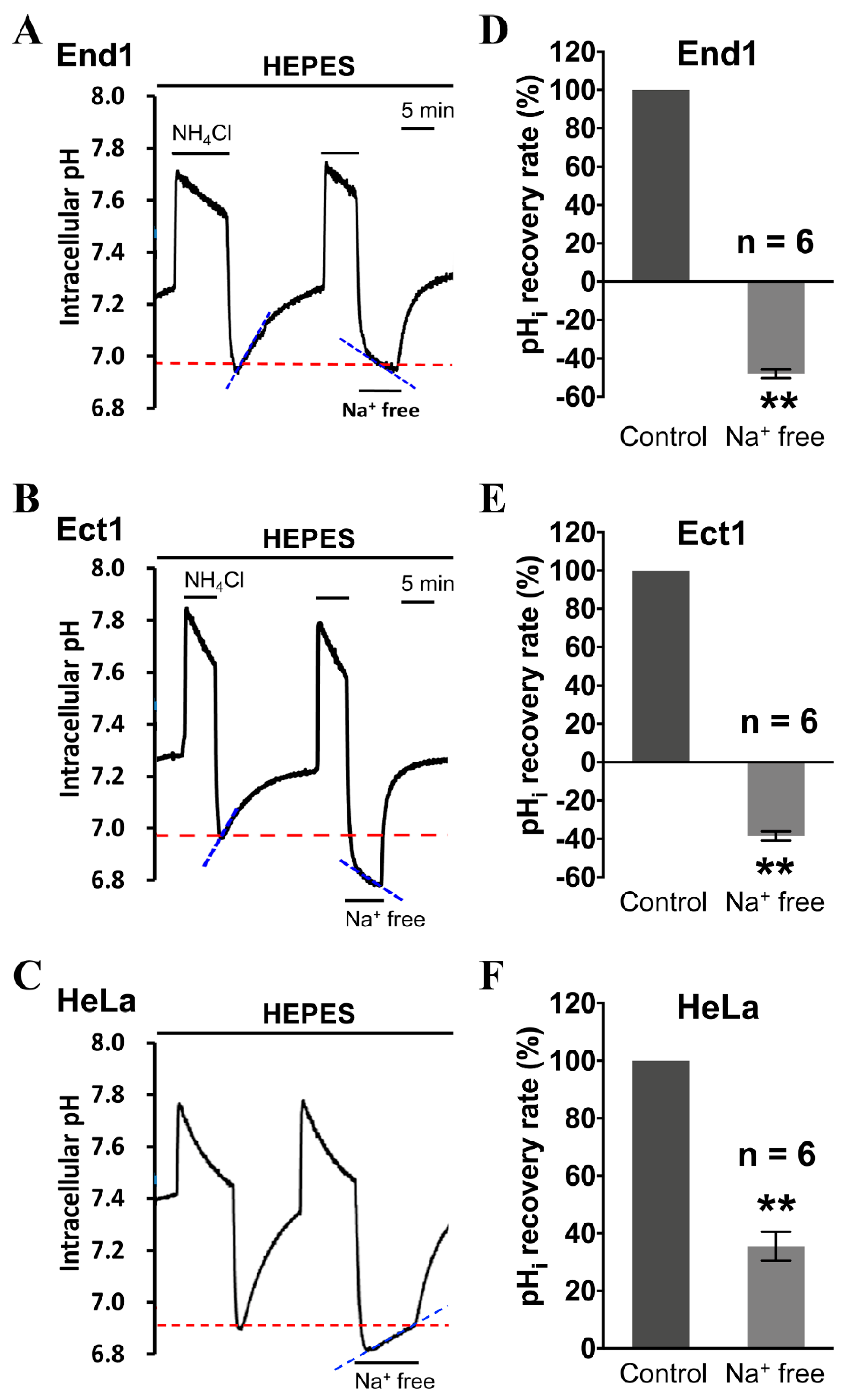
2.2. Functional Identification of Intracellular Acid Extruders—NHE and V-ATPase
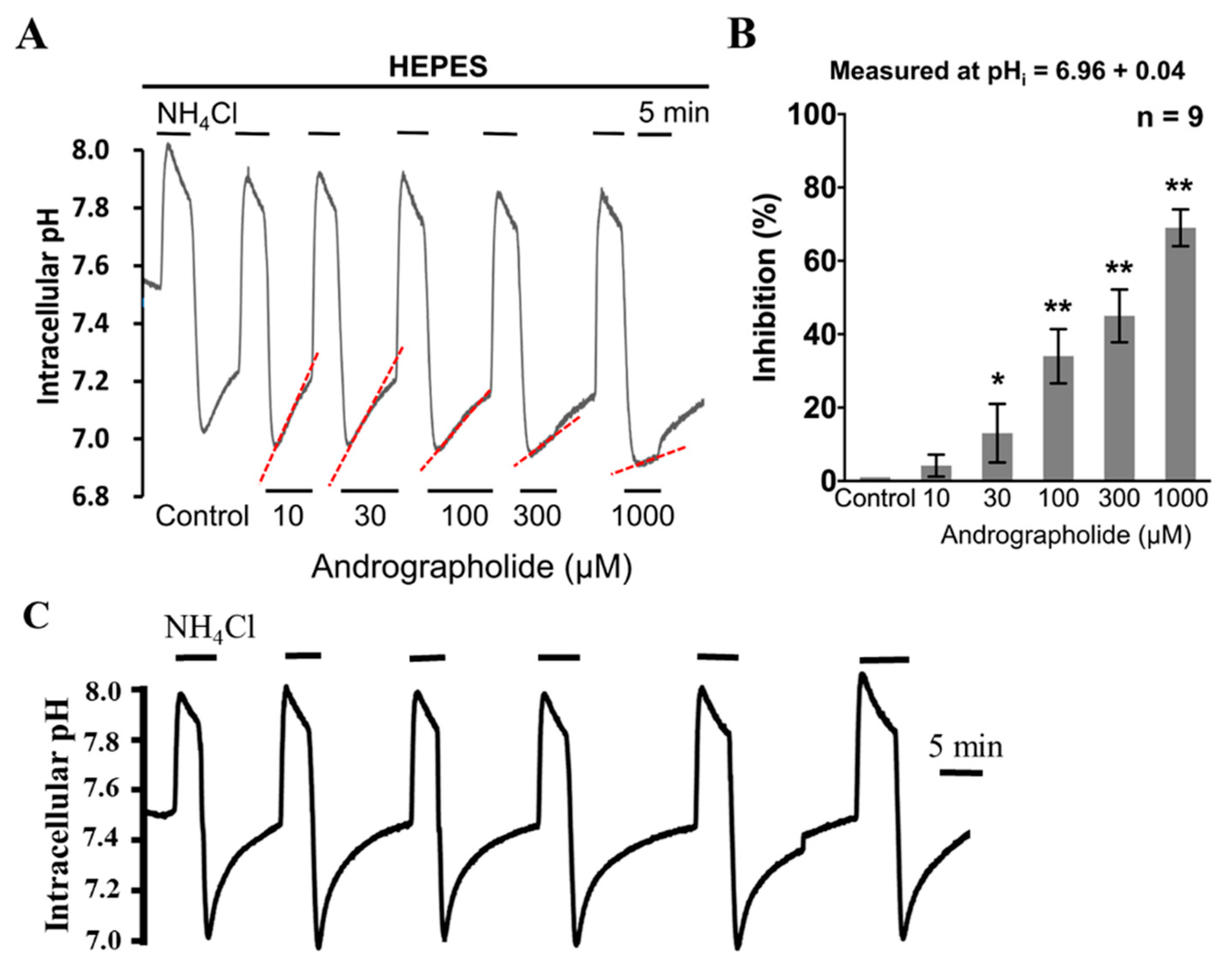
2.3. The Effect of Andrographolide on the Functional Activity of NHE and V-ATPase in HeLa Cells
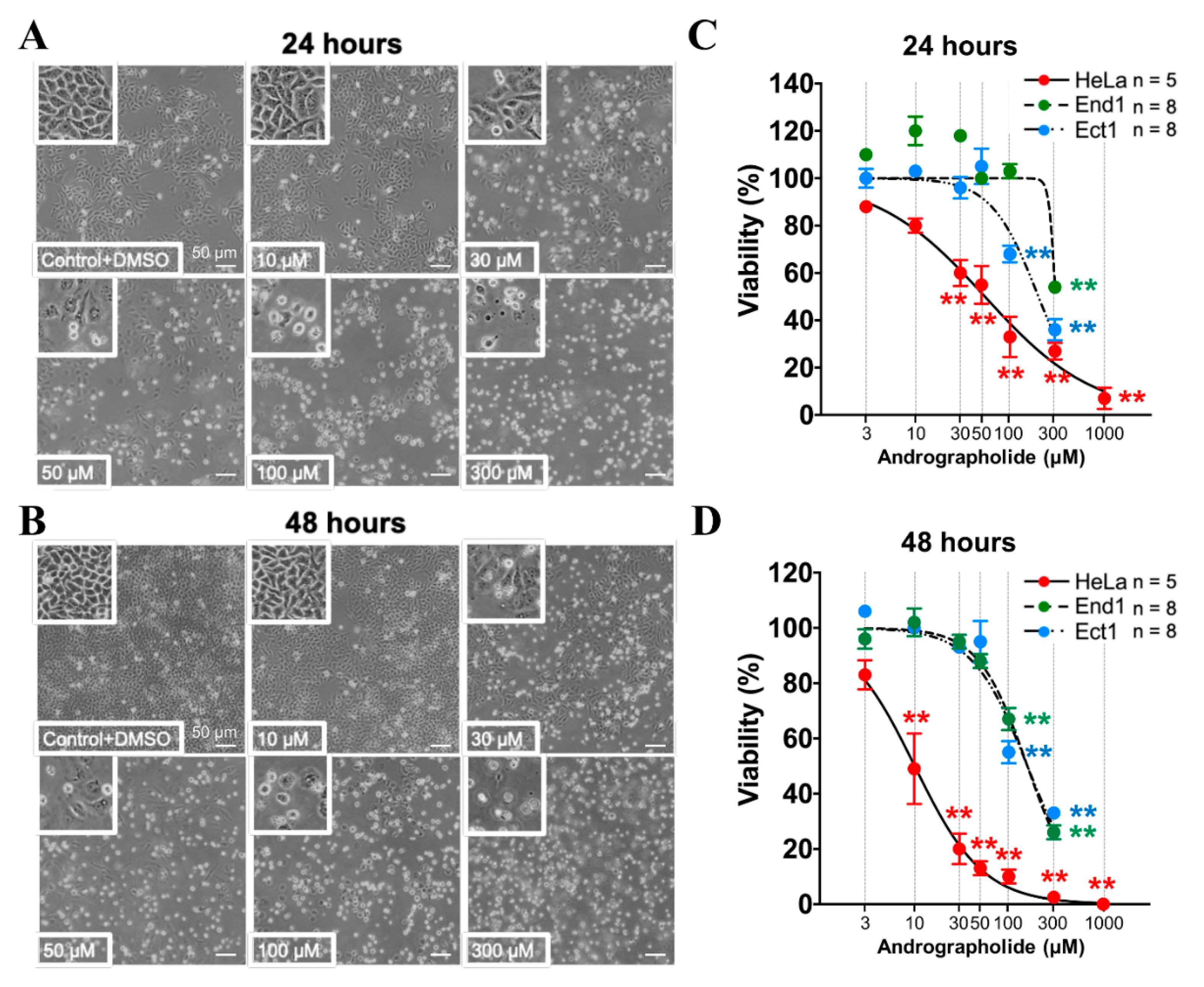
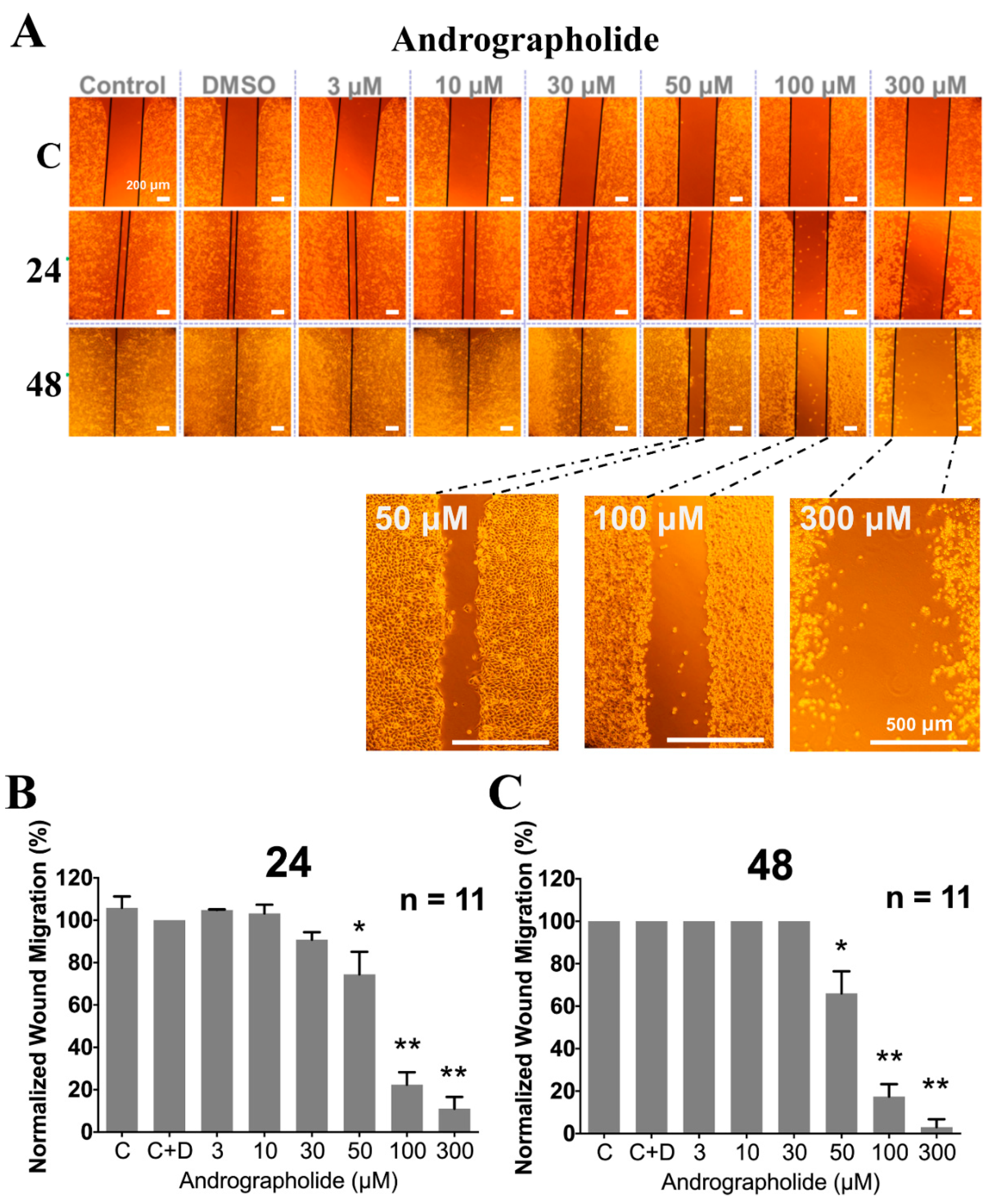
2.4. The Effect of Pretreating with Various Concentration of Andrographolide for 24 and 48 h on Proliferation/Viability and Cell Migration
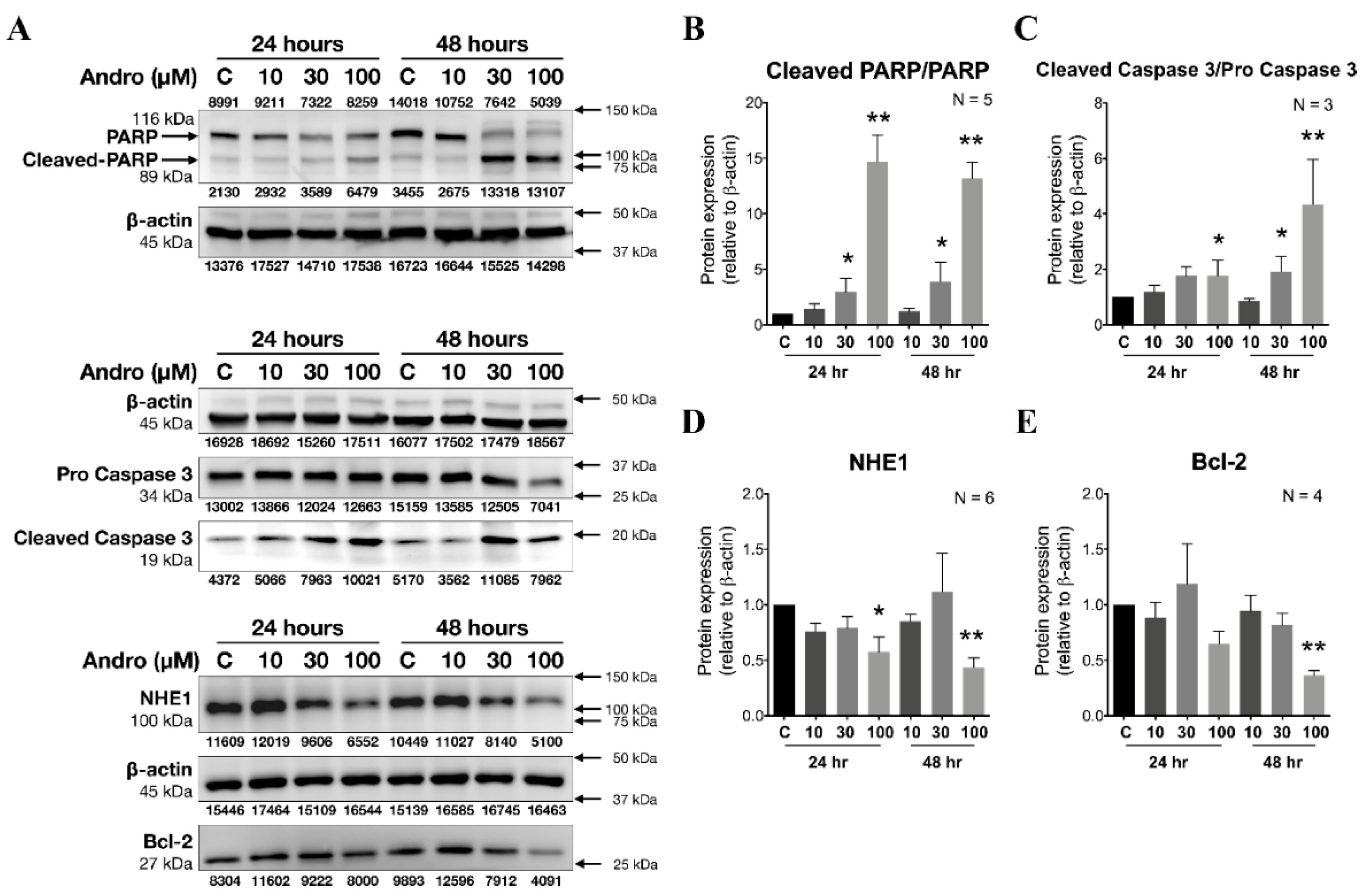
2.5. The Effect of Pretreating with Various Concentration of Andrographolide for 24 and 48 h on Protein Expression of NHE1, Bcl-2, PARP, Cleaved PARP, Pro-Caspase 3, and Cleaved Caspase 3
3. Discussion
3.1. The Resting pHi and New Steady-State pHi in Human Cervical Cancer Cells
3.2. Potential Role of Inhibitors/Activators of Isoforms of NHE and V-ATPase in a Clinical Setting
3.3. The Acute and Chronic Effect of Andrographolide on Intracellular pH Regulating Mechanism and Apoptosis in Cervical Cancer Cells
4. Materials and Methods
4.1. Cell Culture
4.2. Microspectrofluorometry and In Situ Calibration of Intracellular pH Fluorescent Dye BCECF
4.3. NH4Cl Prepulse Technique
4.4. Chemicals and Solutions
4.5. MTT (3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyltetrazolium Bromide) Assay
4.6. Wound Healing Assay
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability Statement
References
- Leem, C.H.; Lagadic-Gossmann, D.; Vaughan-Jones, R.D. Characterization of intracellular pH regulation in the guinea-pig ventricular myocyte. J. Physiol. 1999, 517, 159–180. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, F.A.; Kettunen, M.I.; Day, S.E.; Hu, D.E.; Ardenkjaer-Larsen, J.H.; Zandt, R.I.; Jensen, P.R.; Karlsson, M.; Golman, K.; Lerche, M.H.; et al. Magnetic resonance imaging of pH in vivo using hyperpolarized 13C-labelled bicarbonate. Nature 2008, 453, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Gillies, R.J.; Raghunand, N.; Karczmar, G.S.; Bhujwalla, Z.M. MRI of the tumor microenvironment. J. Magn. Reson Imaging 2002, 16, 430–450. [Google Scholar] [CrossRef]
- Stuwe, L.; Muller, M.; Fabian, A.; Waning, J.; Mally, S.; Noel, J.; Schwab, A.; Stock, C. pH dependence of melanoma cell migration: Protons extruded by NHE1 dominate protons of the bulk solution. J. Physiol. 2007, 585, 351–360. [Google Scholar] [CrossRef]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated pH: A perfect storm for cancer progression. Nat. Rev. Cancer 2011, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Rich, I.N.; Worthington-White, D.; Garden, O.A.; Musk, P. Apoptosis of leukemic cells accompanies reduction in intracellular pH after targeted inhibition of the Na(+)/H(+) exchanger. Blood 2000, 95, 1427–1434. [Google Scholar] [CrossRef]
- Moses, C.; Garcia-Bloj, B.; Harvey, A.R.; Blancafort, P. Hallmarks of cancer: The CRISPR generation. Eur. J. Cancer 2018, 93, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, S.; Llopis, J.; Deveraux, Q.L.; Tsien, R.Y.; Reed, J.C. Changes in intramitochondrial and cytosolic pH: Early events that modulate caspase activation during apoptosis. Nat. Cell Biology 2000, 2, 318–325. [Google Scholar] [CrossRef]
- Loh, S.H.; Jin, J.S.; Tsai, C.S.; Chao, C.M.; Chiung, C.H.; Chen, W.H.; Lin, C.I.; Chuang, C.H.; Wei, J. Functional evidence for intracellular acid extruders in human ventricular myocardium. Jpn. J. Physiol. 2002, 52, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Loh, S.H.; Lee, C.Y.; Tsai, Y.T.; Shih, S.J.; Chen, L.W.; Cheng, T.H.; Tsai, C.S. Intracellular Acid-extruding regulators and the effect of lipopolysaccharide in cultured human renal artery smooth muscle cells. PLoS ONE 2014, 9, e90273. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.T.; Liu, J.Y.; Lee, C.Y.; Tsai, C.S.; Chen, M.H.; Ou, C.C.; Chen, W.H.; Loh, S.H. Functional characterization of transmembrane intracellular pH regulators and mechanism of alcohol-induced intracellular acidosis in human umbilical cord blood stem cell-like cells. J. Cardiovasc. Pharmacol. 2011, 58, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.S.; Loh, S.H.; Liu, J.C.; Lin, J.W.; Chen, Y.L.; Chen, C.H.; Cheng, T.H. Urotensin II-induced endothelin-1 expression and cell proliferation via epidermal growth factor receptor transactivation in rat aortic smooth muscle cells. Atherosclerosis 2009, 206, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Longatto-Filho, A.; Azevedo-Silva, J.; Casal, M.; Schmitt, F.C.; Baltazar, F. Role of monocarboxylate transporters in human cancers: State of the art. J. Bioenerg. Biomembr. 2012, 44, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Stock, C.; Gassner, B.; Hauck, C.R.; Arnold, H.; Mally, S.; Eble, J.A.; Dieterich, P.; Schwab, A. Migration of human melanoma cells depends on extracellular pH and Na+/H+ exchange. J. Physiol. 2005, 567, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Reshkin, S.J.; Bellizzi, A.; Albarani, V.; Guerra, L.; Tommasino, M.; Paradiso, A.; Casavola, V. Phosphoinositide 3-kinase is involved in the tumor-specific activation of human breast cancer cell Na(+)/H(+) exchange, motility, and invasion induced by serum deprivation. J. Biol. Chem. 2000, 275, 5361–5369. [Google Scholar] [CrossRef] [Green Version]
- Busco, G.; Cardone, R.A.; Greco, M.R.; Bellizzi, A.; Colella, M.; Antelmi, E.; Mancini, M.T.; Dell’Aquila, M.E.; Casavola, V.; Paradiso, A.; et al. NHE1 promotes invadopodial ECM proteolysis through acidification of the peri-invadopodial space. FASEB J. 2010, 24, 3903–3915. [Google Scholar] [CrossRef]
- Abdelazeem, K.N.M.; Singh, Y.; Lang, F.; Salker, M.S. Negative Effect of Ellagic Acid on Cytosolic pH Regulation and Glycolytic Flux in Human Endometrial Cancer Cells. Cell Physiol. Biochem. 2017, 41, 2374–2382. [Google Scholar] [CrossRef] [Green Version]
- Moolenaar, W.H. Effects of growth factors on intracellular pH regulation. Annu. Rev. Physiol. 1986, 48, 363–376. [Google Scholar] [CrossRef]
- Pouyssegur, J.; Franchi, A.; L’Allemain, G.; Paris, S. Cytoplasmic pH, a key determinant of growth factor-induced DNA synthesis in quiescent fibroblasts. FEBS Lett. 1985, 190, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Belaud-Rotureau, M.A.; Leducq, N.; Macouillard Poulletier de Gannes, F.; Diolez, P.; Lacoste, L.; Lacombe, F.; Bernard, P.; Belloc, F. Early transitory rise in intracellular pH leads to Bax conformation change during ceramide-induced apoptosis. Apoptosis 2000, 5, 551–560. [Google Scholar] [CrossRef]
- Matsubara, K.; Kubota, M.; Adachi, S.; Kuwakado, K.; Hirota, H.; Wakazono, Y.; Akiyama, Y.; Mikawa, H. Different mode of cell death induced by calcium ionophore in human leukemia cell lines: Possible role of constitutive endonuclease. Exp. Cell Res. 1994, 210, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Lagadic-Gossmann, D.; Huc, L.; Lecureur, V. Alterations of intracellular pH homeostasis in apoptosis: Origins and roles. Cell Death Differ. 2004, 11, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Kim, S.J.; Bae, J.; Wang, Y.; Park, S.Y.; Min, Y.S.; Je, H.D.; Sohn, U.D. The p90rsk-mediated signaling of ethanol-induced cell proliferation in HepG2 cell line. Korean J. Physiol. Pharm. 2016, 20, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panossian, A.; Hovhannisyan, A.; Mamikonyan, G.; Abrahamian, H.; Hambardzumyan, E.; Gabrielian, E.; Goukasova, G.; Wikman, G.; Wagner, H. Pharmacokinetic and oral bioavailability of andrographolide from Andrographis paniculata fixed combination Kan Jang in rats and human. Phytomedicine 2000, 7, 351–364. [Google Scholar] [CrossRef]
- Trivedi, N.P.; Rawal, U.M. Hepatoprotective and antioxidant property of Andrographis paniculata (Nees) in BHC induced liver damage in mice. Indian J. Exp. Biol. 2001, 39, 41–46. [Google Scholar]
- Yu, B.C.; Hung, C.R.; Chen, W.C.; Cheng, J.T. Antihyperglycemic effect of andrographolide in streptozotocin-induced diabetic rats. Planta Med. 2003, 69, 1075–1079. [Google Scholar]
- Sheeja, K.; Kuttan, G. Activation of cytotoxic T lymphocyte responses and attenuation of tumor growth in vivo by Andrographis paniculata extract and andrographolide. Immunopharmacol. Immunotoxicol. 2007, 29, 81–93. [Google Scholar] [CrossRef]
- Yue, G.G.; Lee, J.K.; Li, L.; Chan, K.M.; Wong, E.C.; Chan, J.Y.; Fung, K.P.; Lui, V.W.; Chiu, P.W.; Lau, C.B. Andrographis paniculata elicits anti-invasion activities by suppressing TM4SF3 gene expression and by anoikis-sensitization in esophageal cancer cells. Am. J. Cancer Res. 2015, 5, 3570–3587. [Google Scholar]
- Kumar, R.A.; Sridevi, K.; Kumar, N.V.; Nanduri, S.; Rajagopal, S. Anticancer and immunostimulatory compounds from Andrographis paniculata. J. Ethnopharmacol. 2004, 92, 291–295. [Google Scholar] [CrossRef]
- Kandanur, S.G.; Golakoti, N.R.; Nanduri, S. Synthesis and in vitro cytotoxicity of novel C-12 substituted-14-deoxy-andrographolide derivatives as potent anti-cancer agents. Bioorganic Med. Chem. Lett. 2015, 25, 5781–5786. [Google Scholar] [CrossRef]
- Das, B.; Chowdhury, C.; Kumar, D.; Sen, R.; Roy, R.; Das, P.; Chatterjee, M. Synthesis, cytotoxicity, and structure-activity relationship (SAR) studies of andrographolide analogues as anti-cancer agent. Bioorganic Med. Chem. Lett. 2010, 20, 6947–6950. [Google Scholar] [CrossRef] [PubMed]
- Sirion, U.; Kasemsook, S.; Suksen, K.; Piyachaturawat, P.; Suksamrarn, A.; Saeeng, R. New substituted C-19-andrographolide analogues with potent cytotoxic activities. Bioorganic Med. Chem. Lett. 2012, 22, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, S.; Kumar, R.A.; Deevi, D.S.; Satyanarayana, C.; Rajagopalan, R. Andrographolide, a potential cancer therapeutic agent isolated from Andrographis paniculata. J. Exp. Ther. Oncol. 2003, 3, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, S.; Nyavanandi, V.K.; Thunuguntla, S.S.; Kasu, S.; Pallerla, M.K.; Ram, P.S.; Rajagopal, S.; Kumar, R.A.; Ramanujam, R.; Babu, J.M.; et al. Synthesis and structure-activity relationships of andrographolide analogues as novel cytotoxic agents. Bioorganic Med. Chem. Lett. 2004, 14, 4711–4717. [Google Scholar] [CrossRef]
- Varma, A.; Padh, H.; Shrivastava, N. Andrographolide: A new plant-derived antineoplastic entity on horizon. Evid. -Based Complement. Altern. Med. 2011, 2011, 815390. [Google Scholar] [CrossRef] [Green Version]
- Satyanarayana, C.; Deevi, D.S.; Rajagopalan, R.; Srinivas, N.; Rajagopal, S. DRF 3188 a novel semi-synthetic analog of andrographolide: Cellular response to MCF 7 breast cancer cells. BMC Cancer 2004, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Jayakumar, T.; Hsieh, C.-Y.; Lee, J.-J.; Sheu, J.-R. Experimental and Clinical Pharmacology of Andrographis paniculata and Its Major Bioactive Phytoconstituent Andrographolide. Evid. -Based Complement. Altern. Med. 2013, 2013, 16. [Google Scholar] [CrossRef] [Green Version]
- Gatti, L.; Zunino, F. Overview of tumor cell chemoresistance mechanisms. Methods Mol. Med. 2005, 111, 127–148. [Google Scholar]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Burow, M.E.; Weldon, C.B.; Tang, Y.; Navar, G.L.; Krajewski, S.; Reed, J.C.; Hammond, T.G.; Clejan, S.; Beckman, B.S. Differences in susceptibility to tumor necrosis factor alpha-induced apoptosis among MCF-7 breast cancer cell variants. Cancer Res. 1998, 58, 4940–4946. [Google Scholar]
- Li, J.; Cheung, H.Y.; Zhang, Z.; Chan, G.K.; Fong, W.F. Andrographolide induces cell cycle arrest at G2/M phase and cell death in HepG2 cells via alteration of reactive oxygen species. Eur. J. Pharmacol. 2007, 568, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Manikam, S.D.; Stanslas, J. Andrographolide inhibits growth of acute promyelocytic leukaemia cells by inducing retinoic acid receptor-independent cell differentiation and apoptosis. J. Pharm. Pharmacol. 2009, 61, 69–78. [Google Scholar] [CrossRef]
- Kim, Y.S.; Milner, J.A. Targets for indole-3-carbinol in cancer prevention. J. Nutr. Biochem. 2005, 16, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, H.Y.; Cheung, S.H.; Li, J.; Cheung, C.S.; Lai, W.P.; Fong, W.F.; Leung, F.M. Andrographolide isolated from Andrographis paniculata induces cell cycle arrest and mitochondrial-mediated apoptosis in human leukemic HL-60 cells. Planta Med. 2005, 71, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Ulmschneider, B.; Grillo-Hill, B.K.; Benitez, M.; Azimova, D.R.; Barber, D.L.; Nystul, T.G. Increased intracellular pH is necessary for adult epithelial and embryonic stem cell differentiation. J. Cell Biol. 2016, 215, 345–355. [Google Scholar] [CrossRef]
- Asgharzadeh, M.R.; Barar, J.; Pourseif, M.M.; Eskandani, M.; Jafari Niya, M.; Mashayekhi, M.R.; Omidi, Y. Molecular machineries of pH dysregulation in tumor microenvironment: Potential targets for cancer therapy. Bioimpacts 2017, 7, 115–133. [Google Scholar] [CrossRef] [Green Version]
- Hulikova, A.; Harris, A.L.; Vaughan-Jones, R.D.; Swietach, P. Regulation of intracellular pH in cancer cell lines under normoxia and hypoxia. J. Cell. Physiol. 2013, 228, 743–752. [Google Scholar] [CrossRef]
- Michl, J.; Park, K.C.; Swietach, P. Evidence-based guidelines for controlling pH in mammalian live-cell culture systems. Commun. Biol. 2019, 2, 144. [Google Scholar] [CrossRef]
- Loh, S.H.; Lee, C.Y.; Chen, G.S.; Wu, C.H.; Tsao, C.J.; Shih, S.J.; Tsai, C.S.; Tsai, Y.T. Effects of alcool on intracellular acid-extruders in human aorta smoot muscles. Alcohol. Clin. Exp. Res. 2015, 39, 2302–2312. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Lee, C.Y.; Hsu, C.C.; Hsueh, M.K.; Huang, E.Y.K.; Tsai, C.S.; Loh, S.H. Effects of urotensin II on intracellular pH regulation in cultured human internal mammary artery smooth muscle cells. Peptides 2014, 56, 173–182. [Google Scholar] [CrossRef]
- Loh, S.H.; Sun, B.; Vaughan-Jones, R.D. Effect of Hoe 694, a novel Na(+)-H+ exchange inhibitor, on intracellular pH regulation in the guinea-pig ventricular myocyte. Br. J. Pharmacol. 1996, 118, 1905–1912. [Google Scholar] [CrossRef] [Green Version]
- Loh, S.H.; Chen, W.H.; Chiang, C.H.; Hsueh, M.K.; Huang Eagle, Y.K.; Tsai, C.S.; Loh, S.H. Intracellular pH regulatory mechanism in human atrial myocardium: Functional evidence for Na(+)/H(+) exchanger and Na(+)/HCO(3)(−) symporter. J. Biomed. Sci. 2002, 9, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Feng, J.C.; Ke, L.J.; Xu, J.W.; Huang, Z.X.; Huang, J.; Zhu, Y.X.; Zhou, W.L. Mechanisms underlying the regulation of intracellular and luminal pH in vaginal epithelium. J. Cell. Physiol. 2019, 234, 15790–15799. [Google Scholar] [CrossRef]
- Donders, G.G.; Gonzaga, A.; Marconi, C.; Donders, F.; Michiels, T.; Eggermont, N.; Bellen, G.; Lule, J.; Byamughisa, J. Increased vaginal pH in Ugandan women: What does it indicate? Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, J.D.; Sennoune, S.R.; Maiti, D.; Bakunts, K.; Reuveni, M.; Sanka, S.C.; Martinez, G.M.; Seftor, E.A.; Meininger, C.J.; Wu, G.; et al. Vacuolar-type H+-ATPases at the plasma membrane regulate pH and cell migration in microvascular endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1147–H1157. [Google Scholar] [CrossRef] [Green Version]
- Fais, S.; De Milito, A.; You, H.; Qin, W. Targeting vacuolar H+-ATPases as a new strategy against cancer. Cancer Res. 2007, 67, 10627–10630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacFarlane, M.; Williams, A.C. Apoptosis and disease: A life or death decision. EMBO Rep. 2004, 5, 674–678. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, F.H.; Rahman, K.M.; Li, Y. Bax translocation to mitochondria is an important event in inducing apoptotic cell death by indole-3-carbinol (I3C) treatment of breast cancer cells. J. Nutr. 2003, 133, 2434s–2439s. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Yamaguchi, M.; Kawabata, S.; Murakami, M.; Nakashima, M.; Gotoh, M.; Yamamoto, T. Effects of Extracellular pH on Dental Pulp Cells In Vitro. J. Endod. 2016, 42, 735–741. [Google Scholar] [CrossRef]
- Yang, O.C.Y.; Loh, S.H. Acidic Stress Triggers Sodium-Coupled Bicarbonate Transport and Promotes Survival in A375 Human Melanoma Cells. Sci. Rep. 2019, 9, 6858. [Google Scholar] [CrossRef] [PubMed]
- Roos, A.; Boron, W.F. Intracellular pH. Physiol. Rev. 1981, 61, 296–434. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loh, S.-H.; Tsai, Y.-T.; Huang, S.-F.; Yu, T.-C.; Kuo, P.-C.; Chao, S.-C.; Chou, M.-F.; Tsai, C.-S.; Lee, S.-P. Effects of Andrographolide on Intracellular pH Regulation, Cellular Migration, and Apoptosis in Human Cervical Cancer Cells (Running Tittle: Effects of Andrographolide on pH Regulators and Apoptosis in Cervical Cancer). Cancers 2020, 12, 387. https://doi.org/10.3390/cancers12020387
Loh S-H, Tsai Y-T, Huang S-F, Yu T-C, Kuo P-C, Chao S-C, Chou M-F, Tsai C-S, Lee S-P. Effects of Andrographolide on Intracellular pH Regulation, Cellular Migration, and Apoptosis in Human Cervical Cancer Cells (Running Tittle: Effects of Andrographolide on pH Regulators and Apoptosis in Cervical Cancer). Cancers. 2020; 12(2):387. https://doi.org/10.3390/cancers12020387
Chicago/Turabian StyleLoh, Shih-Hurng, Yi-Ting Tsai, Shu-Fu Huang, Tien-Chieh Yu, Pei-Chun Kuo, Shih-Chi Chao, Mei-Fang Chou, Chien-Sung Tsai, and Shiao-Pieng Lee. 2020. "Effects of Andrographolide on Intracellular pH Regulation, Cellular Migration, and Apoptosis in Human Cervical Cancer Cells (Running Tittle: Effects of Andrographolide on pH Regulators and Apoptosis in Cervical Cancer)" Cancers 12, no. 2: 387. https://doi.org/10.3390/cancers12020387