Effects of Ethanol Exposure and Withdrawal on Neuronal Morphology in the Agranular Insular and Prelimbic Cortices: Relationship with Withdrawal-Related Structural Plasticity in the Nucleus Accumbens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Chronic Ethanol Exposure and Withdrawal
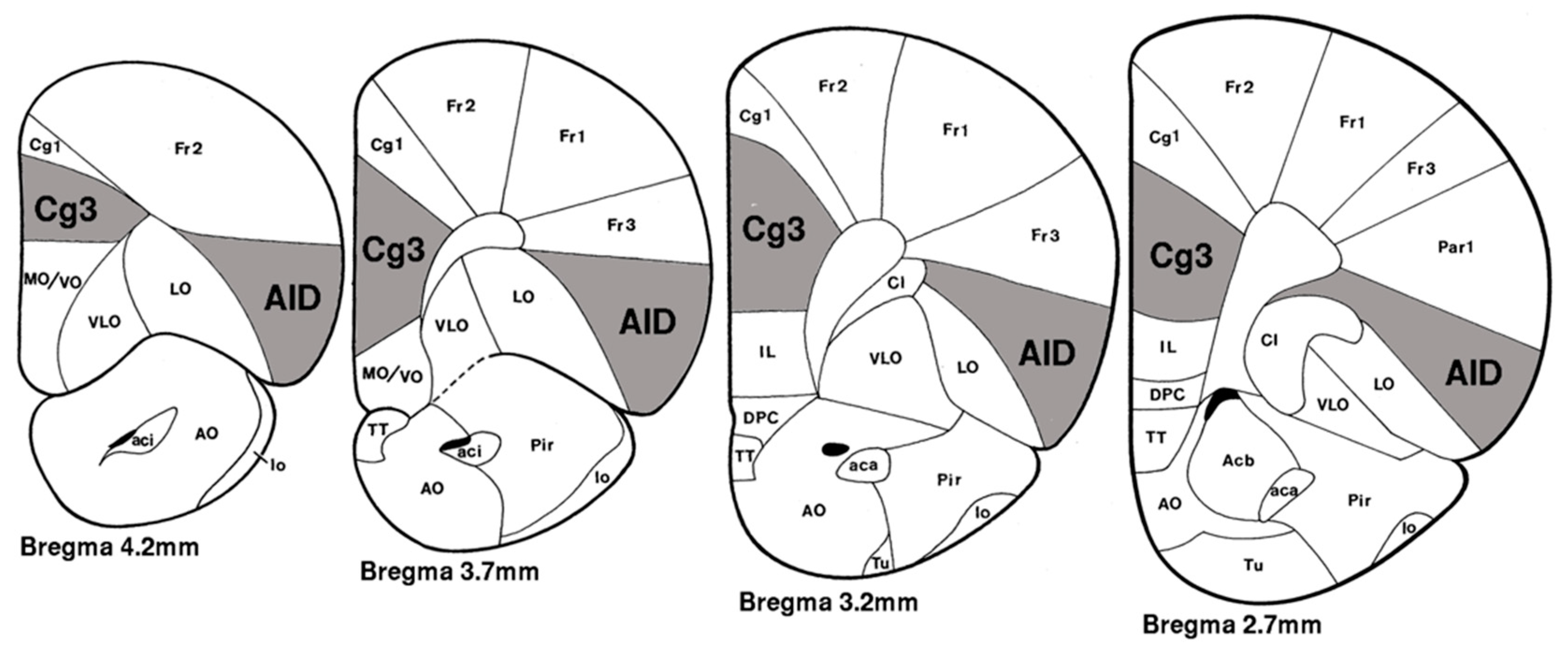
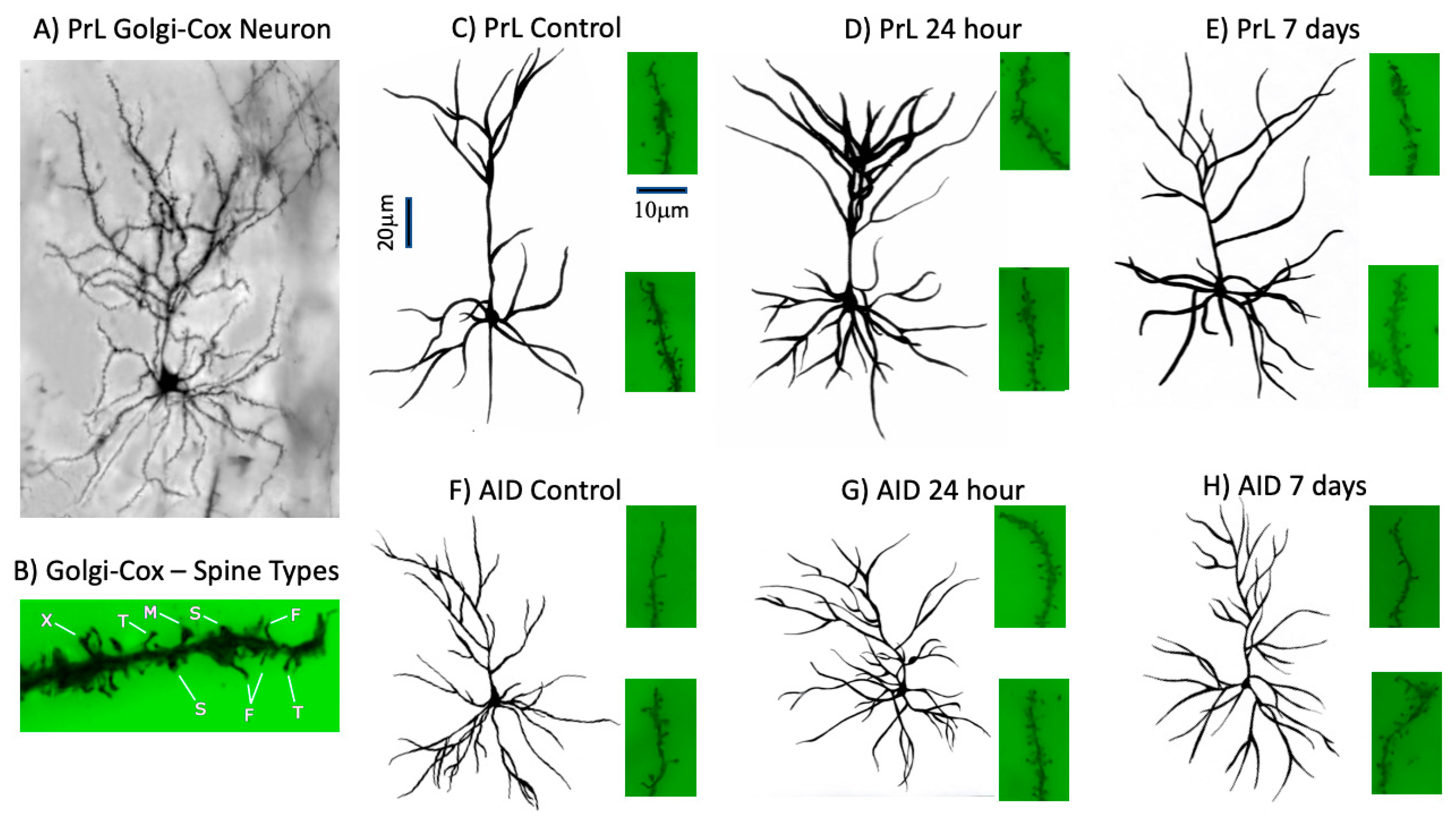
2.3. Golgi–Cox Staining and Analysis
2.4. Statistical Analyses
3. Results
3.1. Total Dendritic Length, Branching, and Spine Density in PrL and AID
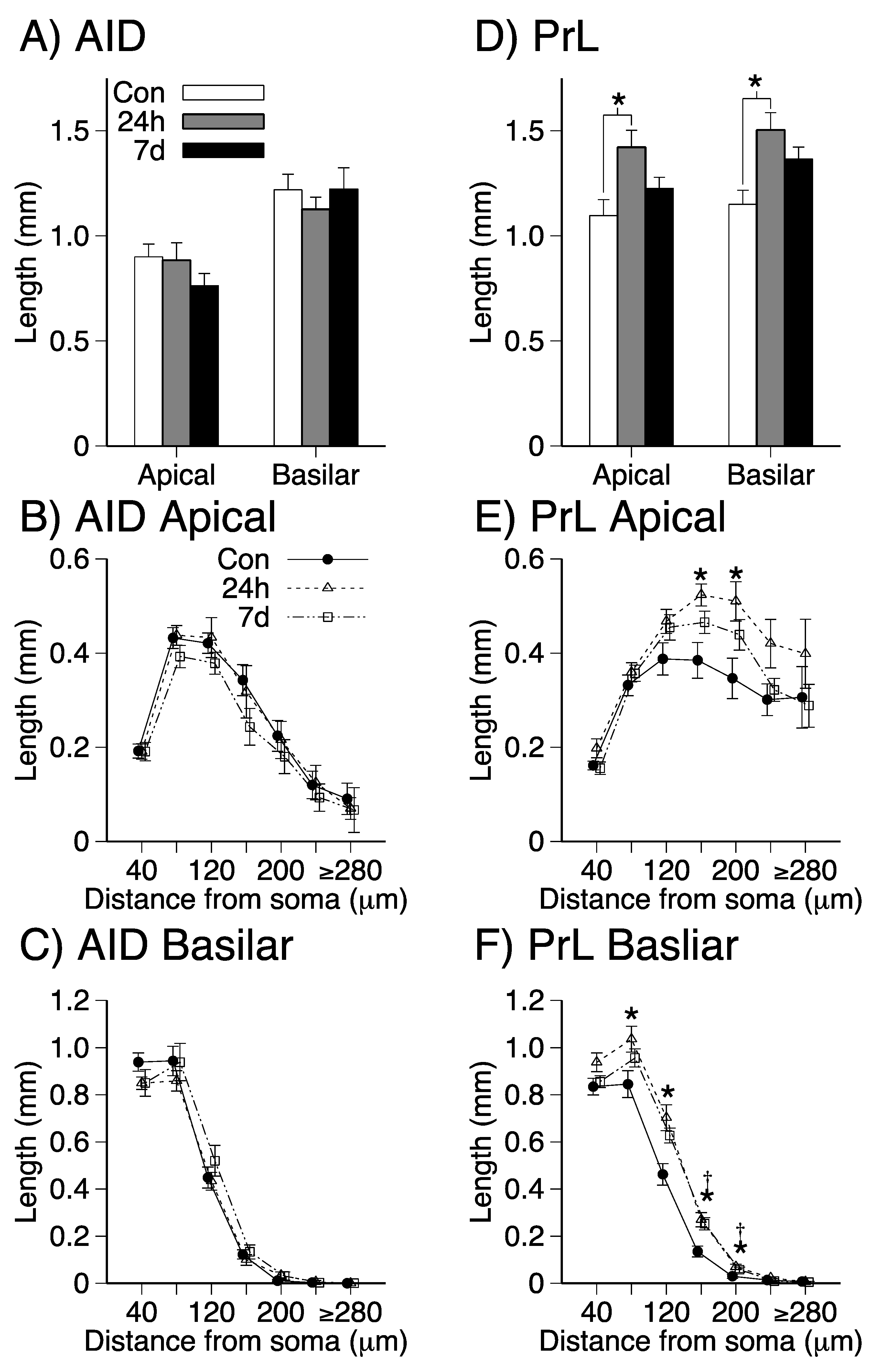
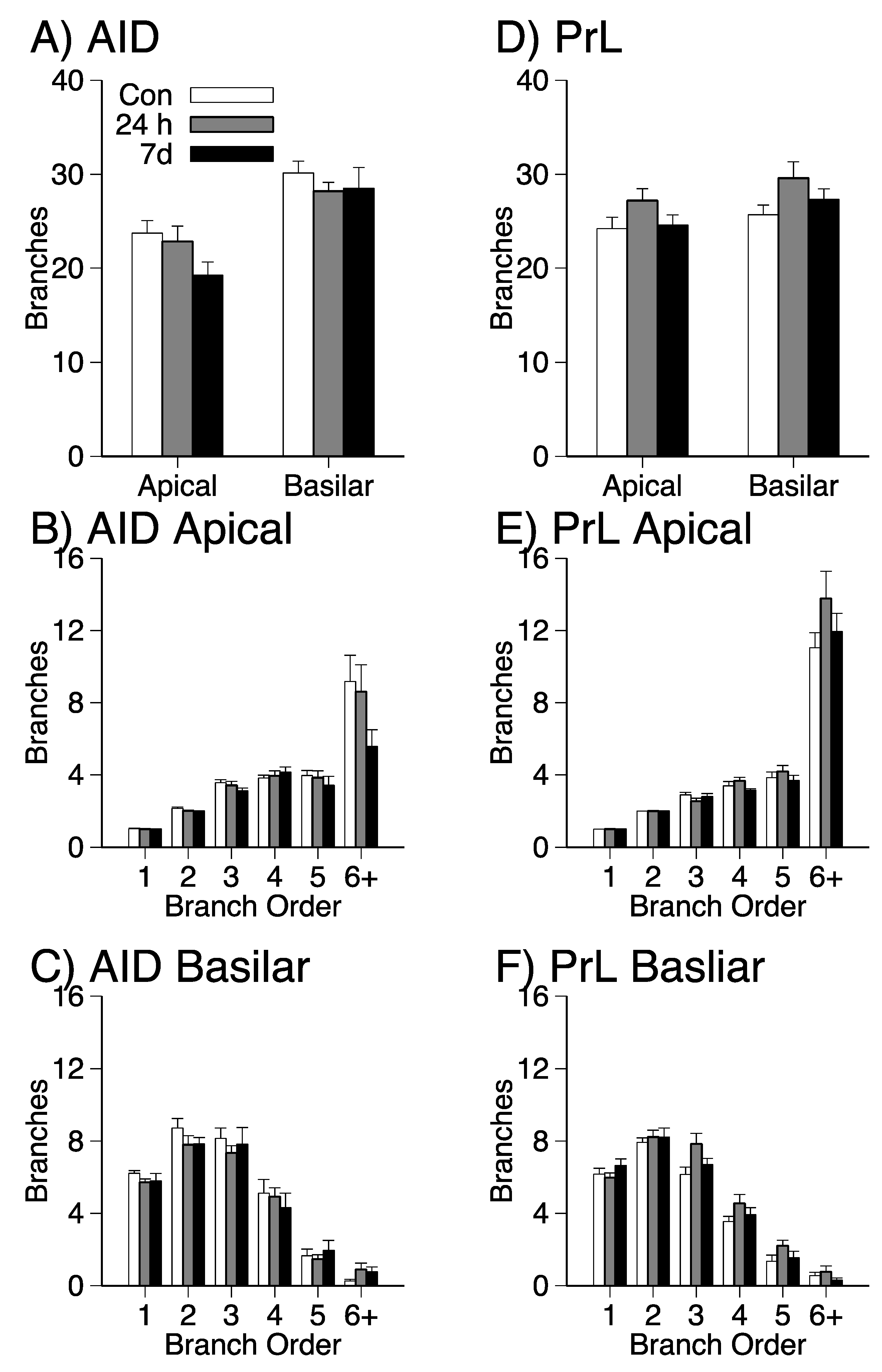
3.2. Dendritic Length and Branching in Apical and Basilar Dendritic Fields of PrL and AID
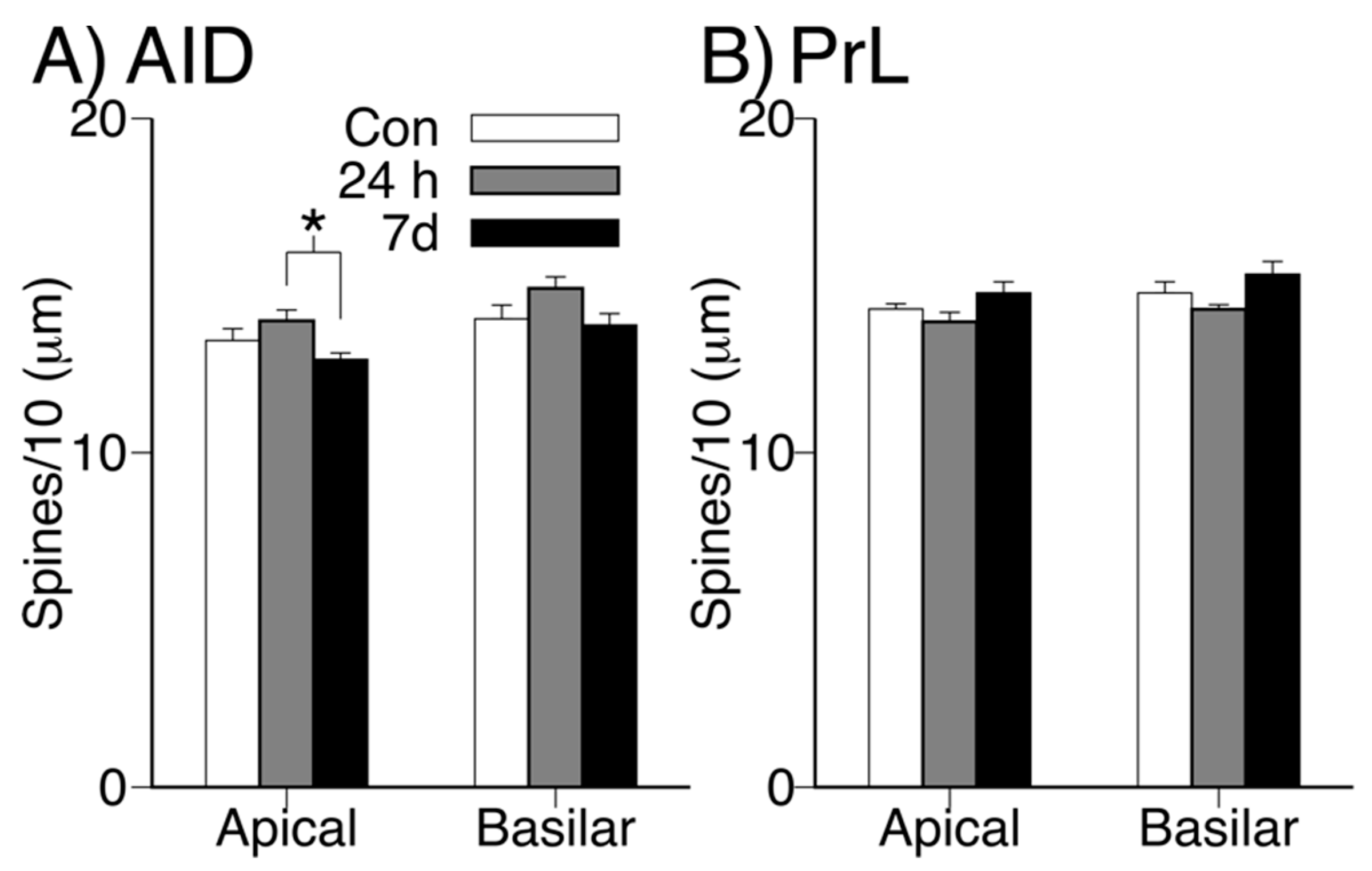
3.3. Dendritic Spine Density in Apical and Basilar Fields of PrL and AID
3.4. Spine Morphology in PrL and AID
3.5. Relationship between Nucleus Accumbens and Frontal Cortex Dendritic Morphology and Spine Density Following Ethanol Exposure and Withdrawal
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 24 h | 7 d | |
|---|---|---|---|
| NAc Core | |||
| Branches | 34.92 (1.252) | 32.150 (1.014) | # 35.463 (1.639) |
| Dendritic Length (mm) | 1.319 (0.071) | 1.278 (0.058) | # 1.456 (0.057) |
| Total Spine Density | 8.660 (0.290) | 9.715 (0.519) | 9.304 (0.547) |
| Mushroom Spine Density | 2.424 (0.161) | 3.123 (0.267) | 2.332 (0.300) |
| Stubby Spine Density | 2.813 (0.321) | 2.829 (0.245) | 3.940 (0.311) |
| NAc Shell | |||
| Branches | 34.33 (1.002) | 32.088 (1.189) | # 32.563 (1.566) |
| Dendritic Length (mm) | 1.333 (0.051) | 1.261 (0.055) | # 1.293 (0.056) |
| Total Spine Density | 8.976 (0.383) | 9.234 (0.497) | 9.355 (0.570) |
| Mushroom Spine Density | 2.589 (0.161) | 2.881 (0.105) | 2.299 (0.223) |
| Stubby Spine Density | 2.916 (0.403) | 3.002 (0.254) | 3.911 (0.284) |
References
- Peterson, V.L.; McCool, B.A.; Hamilton, D.A. Effects of ethanol exposure and withdrawal on dendritic morphology and spine density in the nucleus accumbens core and shell. Brain Res. 2015, 1594, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Spiga, S.; Talani, G.; Mulas, G.; Licheri, V.; Fois, G.R.; Muggironi, G.; Masala, N.; Cannizzaro, C.; Biggio, G.; Sanna, E.; et al. Hampered long-term depression and thin spine loss in the nucleus accumbens of ethanol-dependent rats. Proc. Natl. Acad. Sci. USA 2014, 111, E3745–E3754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiga, S.; Puddu, M.C.; Pisano, M.; Diana, M. Morphine withdrawal-induced morphological changes in the nucleus accumbens. Eur. J. Neurosci. 2005, 22, 2332–2340. [Google Scholar] [CrossRef] [PubMed]
- McMullen, P.A.; Stcyr, J.A.; Carlen, P.L. Morphological alterations in rat CA1 hippocampal pyramidal cell dendrites resulting from chronic ethanol-consumption and withdrawal. J. Comp. Neurol. 1984, 225, 111–118. [Google Scholar] [CrossRef] [PubMed]
- McGuier, N.S.; Padula, A.E.; Lopez, M.F.; Woodward, J.J.; Mulholland, P.J. Withdrawal from chronic intermittent alcohol, exposure increases dendritic spine density in the lateral orbitofrontal cortex of mice. Alcohol 2015, 49, 21–27. [Google Scholar] [CrossRef]
- Romero, A.M.; Renau-Piqueras, J.; Marin, M.P.; Timoneda, J.; Berciano, M.T.; Lafarga, M.; Esteban-Pretel, G. Chronic Alcohol Alters Dendritic Spine Development in Neurons in Primary Culture. Neurotoxicity Research 2013, 24, 532–548. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, D.A.; Kolb, B. Differential effects of nicotine and complex housing on subsequent experience-dependent structural plasticity in the nucleus accumbens. Behav. Neurosci. 2005, 119, 355–365. [Google Scholar] [CrossRef]
- Kolb, B.; Gorny, G.; Li, Y.L.; Samaha, A.N.; Robinson, T.E. Amphetamine or cocaine limits the ability of later experience to promote structural plasticity in the neocortex and nucleus accumbens. Proc. Natl. Acad. Sci. USA 2003, 100, 10523–10528. [Google Scholar] [CrossRef] [Green Version]
- Robinson, T.E.; Kolb, B. Structural plasticity associated with exposure to drugs of abuse. Neuropharmacology 2004, 47, 33–46. [Google Scholar] [CrossRef]
- Joyce, E.M.; Robbins, T.W. Frontal-Lobe Function In Korsakoff And Non-Korsakoff Alcoholics—Planning And Spatial Working Memory. Neuropsychologia 1991, 29, 709–723. [Google Scholar] [CrossRef]
- Ahveninen, J.; Jaaskelainen, I.P.; Pekkonen, E.; Hallberg, A.; Hietanen, M.; Naatanen, R.; Sillanaukee, P. Global field power of auditory N1 correlates with impaired verbal-memory performance in human alcoholics. Neurosci. Lett. 2000, 285, 131–134. [Google Scholar] [CrossRef]
- Loeber, S.; Duka, T.; Welzel, H.; Nakovics, H.; Heinz, A.; Flor, H.; Mann, K. Impairment of Cognitive Abilities and Decision Making after Chronic Use of Alcohol: The Impact of Multiple Detoxifications. Alcohol Alcohol. 2009, 44, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Ceccanti, M.; Coriale, G.; Hamilton, D.A.; Carito, V.; Coccurello, R.; Scalese, B.; Ciafre, S.; Codazzo, C.; Messina, M.P.; Chaldakov, G.N.; et al. Virtual Morris task responses in individuals in an abstinence phase from alcohol. Can. J. Physiol. Pharmacol. 2018, 96, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ceccanti, M.; Hamilton, D.; Coriale, G.; Carito, V.; Aloe, L.; Chaldakov, G.; Romeo, M.; Ceccanti, M.; Iannitelli, A.; Fiore, M. Spatial learning in men undergoing alcohol detoxification. Physiol. Behav. 2015, 149, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Zilles, K. The Cortex of the Rat: A Stereotaxic Atlas; Springer: Berlin, Germany, 1985. [Google Scholar]
- Crombag, H.S.; Gorny, G.; Li, Y.L.; Kolb, B.; Robinson, T.E. Opposite effects of amphetamine self-administration experience on dendritic spines in the medial and orbital prefrontal cortex. Cereb. Cortex 2005, 15, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Gorny, G.; Savage, V.R.; Kolb, B. Widespread but regionally specific effects of experimenter- versus self-administered morphine on dendritic spines in the nucleus accumbens, hippocampus, and neocortex of adult rats. Synapse 2002, 46, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liston, C.; Miller, M.M.; Goldwater, D.S.; Radley, J.J.; Rocher, A.B.; Hof, P.R.; Morrison, J.H.; McEwen, B.S. Stress-induced alterations in prefrontal cortical dendritic morphology predict selective impairments in perceptual attentional set-shifting. J. Neurosci. 2006, 26, 7870–7874. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Buck, C.L.; Cohen, A.; Edwards, S.; Park, P.E.; Schlosburg, J.E.; Schmeichel, B.; Vendruscolo, L.F.; Wade, C.L.; Whitfield, T.W.; et al. Addiction as a stress surfeit disorder. Neuropharmacology 2014, 76, 370–382. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. the neural basis of drug craving—An incentive-sensitization theory of addiction. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Ikemoto, S.; Panksepp, J. The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seeking. Brain Res. Rev. 1999, 31, 6–41. [Google Scholar] [CrossRef]
- Di Chiara, G. Nucleus accumbens shell and core dopamine: differential role in behavior and addiction. Behav. Brain Res. 2002, 137, 75–114. [Google Scholar] [CrossRef]
- McFarland, K.; Lapish, C.C.; Kalivas, P.W. Prefrontal glutamate release into the core of the nucleus accumbens mediates cocaine-induced reinstatement of drug-seeking behavior. J. Neurosci. 2003, 23, 3531–3537. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Robbins, T.W. Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481–1489. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of Addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, L.; Burkey, A.R.; Granato, A.; Ohara, P.T. Rostral agranular insular cortex and pain areas of the central nervous system: A tract-tracing study in the rat (vol 468, pg 311, 2004). J. Comp. Neurol. 2004, 480, 425–440. [Google Scholar] [CrossRef]
- Brog, J.S.; Salyapongse, A.; Deutch, A.Y.; Zahm, D.S. The patterns of afferent innervation of the core and shell in the “Accumbens” part of the rat ventral striatum: Immunohistochemical detection of retrogradely transported fluoro-gold. J. Comp. Neurol. 1993, 338, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Heimer, L.; Zahm, D.S.; Churchill, L.; Kalivas, P.W.; Wohltmann, C. Specificity in the projection patterns of accumbal core and shell in the rat. Neuroscience 1991, 41, 89–125. [Google Scholar] [CrossRef]
- Zahm, D.S.; Williams, E.; Wohltmann, C. Ventral striatopallidothalamic projection. 4. Relative involvements of neurochemically distinct subterritories in the ventral pallidum and adjacent parts of the rostroventral forebrain. J. Comp. Neurol. 1996, 364, 340–362. [Google Scholar] [CrossRef]
- Zahm, D.S.; Heimer, L. Specificity in the efferent projections of the nucleus-accumbens in the rat: Comparison of the rostral pole projection patterns with those of the core and shell. J. Comp. Neurol. 1993, 327, 220–232. [Google Scholar] [CrossRef]
- Zahm, D.S.; Heimer, L. Transpallidal pathways originating in the rat nucleus-accumbens. J. Comp. Neurol. 1990, 302, 437–446. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Berendse, H.W.; Haber, S.N. Organization of the output of the ventral striatopallidal system in the rat—ventral pallidal efferents. Neuroscience 1993, 57, 113–142. [Google Scholar] [CrossRef]
- Groenewegen, H.J. Organization of the afferent connections of the mediodorsal thalamic nucleus in the rat, related to the mediodorsal prefrontal topography. Neuroscience 1988, 24, 379–431. [Google Scholar] [CrossRef]
- Zhou, F.C.; Anthony, B.; Dunn, K.W.; Lindquist, W.B.; Xu, Z.C.; Deng, P. Chronic alcohol drinking alters neuronal dendritic spines in the brain reward center nucleus accumbens. Brain Res. 2007, 1134, 148–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uys, J.D.; McGuier, N.S.; Gass, J.T.; Griffin, W.C.; Ball, L.E.; Mulholland, P.J. Chronic intermittent ethanol exposure and withdrawal leads to adaptations in nucleus accumbens core postsynaptic density proteome and dendritic spines. Addict. Biol. 2016, 21, 560–574. [Google Scholar] [CrossRef] [PubMed]
- Varodayan, F.P.; Sidhu, H.; Kreifeldt, M.; Roberto, M.; Contet, C. Morphological and functional evidence of increased excitatory signaling in the prelimbic cortex during ethanol withdrawal. Neuropharmacology 2018, 133, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Lack, A.K.; Diaz, M.R.; Chappell, A.; DuBois, D.W.; McCool, B.A. Chronic ethanol and withdrawal differentially modulate pre- and postsynaptic function at glutamatergic synapses in rat basolateral amygdala. J. Neurophysiol. 2007, 98, 3185–3196. [Google Scholar] [CrossRef] [PubMed]
- Christian, D.T.; Alexander, N.J.; Diaz, M.R.; Robinson, S.; McCool, B.A. Chronic intermittent ethanol and withdrawal differentially modulate basolateral amygdala AMPA-type glutamate receptor function and trafficking. Neuropharmacology 2012, 62, 2430–2439. [Google Scholar] [CrossRef] [Green Version]
- Diaz, M.R.; Christian, D.T.; Anderson, N.J.; McCool, B.A. Chronic Ethanol and Withdrawal Differentially Modulate Lateral/Basolateral Amygdala Paracapsular and Local GABAergic Synapses. J. Pharmacol. Exp. Ther. 2011, 337, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Gibb, R.; Kolb, B. A method for vibratome sectioning of Golgi-Cox stained whole rat brain. J. Neurosci. Methods 1998, 79, 1–4. [Google Scholar] [CrossRef]
- Christian, D.T.; Alexander, N.J.; Diaz, M.R.; McCool, B.A. Thalamic glutamatergic afferents into the rat basolateral amygdala exhibit increased presynaptic glutamate function following withdrawal from chronic intermittent ethanol. Neuropharmacology 2013, 65, 134–142. [Google Scholar] [CrossRef]
- Glaser, E.M.; van der Loos, H. Analysis of thick brain sections by obverse-reverse computer microscopy: Application of a new, high clarity Golgi-Nissl stain. J. Neurosci. Methods 1981, 4, 117–125. [Google Scholar] [CrossRef]
- Hamilton, D.A.; Akers, K.G.; Rice, J.P.; Johnson, T.E.; Candelaria-Cook, F.T.; Maes, L.I.; Rosenberg, M.; Valenzuela, C.F.; Savage, D.D. Prenatal exposure to moderate levels of ethanol alters social behavior in adult rats: Relationship to structural plasticity and immediate early gene expression in frontal cortex. Behav. Brain Res. 2010, 207, 290–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, P.D.; Riesen, A.H. Environmental effects on cortical dendritic fields. I. Rearing in the dark. J. Anat. 1968, 102, 363–374. [Google Scholar] [PubMed]
- Sholl, D.A. The Organization of the Cerebral Cortex; Methuen: London, UK, 1981. [Google Scholar]
- Hering, H.; Sheng, M. Dendritic spines: Structure, dynamics and regulation. Nat. Rev. Neurosci. 2001, 2, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H.; Matsuzaki, M.; Noguchi, J.; Yasumatsu, N.; Nakahara, H. Structure-stability-function relationships of dendritic spines. Trends Neurosci. 2003, 26, 360–368. [Google Scholar] [CrossRef]
- Konur, S.; Rabinowitz, D.; Fenstermaker, V.L.; Yuste, R. Systematic regulation of spine sizes and densities in pyramidal neurons. J. Neurobiol. 2003, 56, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Spacek, J.; Harris, K.M. Three-dimensional organization of smooth endoplasmic reticulum in hippocampal CA1 dendrites and dendritic spines of the immature and mature rat. J. Neurosci. 1997, 17, 190–203. [Google Scholar] [CrossRef]
- Segal, M.; Vlachos, A.; Korkotian, E. The Spine Apparatus, Synaptopodin, and Dendritic Spine Plasticity. Neuroscientist 2010, 16, 125–131. [Google Scholar] [CrossRef]
- Griffin, W.C., III; Haun, H.L.; Hazelbaker, C.L.; Ramachandra, V.S.; Becker, H.C. Increased Extracellular Glutamate In the Nucleus Accumbens Promotes Excessive Ethanol Drinking in Ethanol Dependent Mice. Neuropsychopharmacology 2014, 39, 707–717. [Google Scholar] [CrossRef]
- Ding, Z.-M.; Rodd, Z.A.; Engleman, E.A.; Bailey, J.A.; Lahiri, D.K.; McBride, W.J. Alcohol drinking and deprivation alter basal extracellular glutamate concentrations and clearance in the mesolimbic system of alcohol-preferring (P) rats. Addict. Biol. 2013, 18, 297–306. [Google Scholar] [CrossRef]
- Faingold, C.; Li, Y.; Evans, M.S. Decreased GABA and increased glutamate receptor-mediated activity on inferior colliculus neurons in vitro are associated with susceptibility to ethanol withdrawal seizures. Brain Res. 2000, 868, 287–295. [Google Scholar] [CrossRef]
- Kash, T.L.; Baucum, A.J., II; Conrad, K.L.; Colbran, R.J.; Winder, D.G. Alcohol Exposure Alters NMDAR Function in the Bed Nucleus of the Stria Terminalis. Neuropsychopharmacology 2009, 34, 2420–2429. [Google Scholar] [CrossRef] [PubMed]
- Roberto, M.; Schweitzer, P.; Madamba, S.G.; Stouffer, D.G.; Parsons, L.H.; Siggins, G.R. Acute and chronic ethanol alter glutamatergic transmission in rat central amygdala: An in vitro and in vivo analysis. J. Neurosci. 2004, 24, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Fiala, J.C.; Feinberg, M.; Popov, V.; Harris, K.M. Synaptogenesis via dendritic filopodia in developing hippocampal area CA1. J. Neurosci. 1998, 18, 8900–8911. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M. Structure, development, and plasticity of dendritic spines. Curr. Opin. Neurobiol. 1999, 9, 343–348. [Google Scholar] [CrossRef]
- Nimchinsky, E.A.; Sabatini, B.L.; Svoboda, K. Structure and function of dendritic spines. Annu. Rev. Physiol. 2002, 64, 313–353. [Google Scholar] [CrossRef] [PubMed]
- Logrip, M.L.; Janak, P.H.; Ron, D. Dynorphin is a downstream effector of striatal BDNF regulation of ethanol intake. FASEB J. 2008, 22, 2393–2404. [Google Scholar] [CrossRef] [Green Version]
- McGough, N.N.H.; He, D.Y.; Logrip, M.L.; Jeanblanc, J.; Phamluong, K.; Luong, K.; Kharazia, V.; Janak, P.H.; Ron, D. RACK1 and brain-derived neurotrophic factor: A homeostatic pathway that regulates alcohol addiction. J. Neurosci. 2004, 24, 10542–10552. [Google Scholar] [CrossRef]
- Logrip, M.L.; Janak, P.H.; Ron, D. Escalating ethanol intake is associated with altered corticostriatal BDNF expression. J. Neurochem. 2009, 109, 1459–1468. [Google Scholar] [CrossRef]
- Swanger, S.A.; Yao, X.; Gross, C.; Bassell, G.J. Automated 4D analysis of dendritic spine morphology: applications to stimulus-induced spine remodeling and pharmacological rescue in a disease model. Molecular Brain 2011, 4. [Google Scholar] [CrossRef]
- Euston, D.R.; Gruber, A.J.; McNaughton, B.L. The Role of Medial Prefrontal Cortex in Memory and Decision Making. Neuron 2012, 76, 1057–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, T.M.; Cole, S.; Hamlin, A.S.; McNally, G.P. The Role of Prefrontal Cortex in Predictive Fear Learning. Behav. Neurosci. 2010, 124, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.F.; Watkins, L.R. Role of the medial prefrontal cortex in coping and resilience. Brain Res. 2010, 1355, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermann, A.; Schafer, A.; Walter, B.; Stark, R.; Vaitl, D.; Schienle, A. Emotion regulation in spider phobia: role of the medial prefrontal cortex. Soc. Cogn. Affect. Neurosci. 2009, 4, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, D.A.; Brigman, J.L. Behavioral flexibility in rats and mice: contributions of distinct frontocortical regions. Genes Brain Behav. 2015, 14, 4–21. [Google Scholar] [CrossRef]
- Kesner, R.P.; Gilbert, P.E. The role of the agranular insular cortex in anticipation of reward contrast. Neurobiol. Learn. Mem. 2007, 88, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffeen, U.; Ortega-Legaspi, J.M.; Lopez-Munoz, F.J.; Simon-Arceo, K.; Jaimes, O.; Pellicer, F. Insular cortex lesion diminishes neuropathic and inflammatory pain-like behaviours. Eur. J. Pain 2011, 15, 132–138. [Google Scholar] [CrossRef]
- McCool, B.A.; Christian, D.T.; Diaz, M.R.; Läck, A.K. Glutamate Plasticity in the Drunken Amygdala: The Making of an Anxious Synapse. In International Review of Neurobiology; Elsevier BV: Amsterdam, The Netherlands, 2010; Volume 91, pp. 205–233. [Google Scholar]
- Roberts, A.J.; Cole, M.; Koob, G.F. Intra-amygdala muscimol decreases operant ethanol self-administration in dependent rats. Alcohol. Clin. Exp. Res. 1996, 20, 1289–1298. [Google Scholar] [CrossRef]
- Macey, D.J.; Schulteis, G.; Heinrichs, S.C.; Koob, G.F. Time-dependent quantifiable withdrawal from ethanol in the rat: Effect of method of dependence induction. Alcohol 1996, 13, 163–170. [Google Scholar] [CrossRef]
- Roberts, A.J.; Heyser, C.J.; Cole, M.; Griffin, P.; Koob, G.F. Excessive ethanol drinking following a history of dependence: Animal model of allostasis. Neuropsychopharmacology 2000, 22, 581–594. [Google Scholar] [CrossRef]
- Buck, C.L.; Malavar, J.C.; George, O.; Koob, G.F.; Vendruscolo, L.F. Anticipatory 50 kHz ultrasonic vocalizations are associated with escalated alcohol intake in dependent rats. Behav. Brain Res. 2014, 271, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Rassnick, S.; Heinrichs, S.C.; Britton, K.T.; Koob, G.F. Microinjection of a corticotropin-releasing factor antagonist into the central nucleus of the amygdala reverses anxiogenic-like effects of ethanol withdrawal. Brain Res. 1993, 605, 25–32. [Google Scholar] [CrossRef]
- Valdez, G.R.; Sabino, V.; Koob, G.F. Increased anxiety-like behavior and ethanol self-administration in dependent rats: Reversal via corticotropin-releasing factor-2 receptor activation. Alcohol. Clin. Exp. Res. 2004, 28, 865–872. [Google Scholar] [CrossRef] [PubMed]
- McCool, B.A. Ethanol modulation of synaptic plasticity. Neuropharmacology 2011, 61, 1097–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.; Fitzgerald, P.J.; MacPherson, K.P.; DeBrouse, L.; Colacicco, G.; Flynn, S.M.; Masneuf, S.; Pleil, K.E.; Li, C.; Marcinkiewcz, C.A.; et al. Chronic alcohol remodels prefrontal neurons and disrupts NMDAR-mediated fear extinction encoding. Nat. Neurosci. 2012, 15, 1359. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.M.; Ye, Z.; Wang, H.; Hua, T.M.; Wen, Q.Y.; Zhang, C.Z. Theta-gamma coupling in the prelimbic area is associated with heroin addiction. Neurosci. Lett. 2019, 701, 26–31. [Google Scholar] [CrossRef]
- Susick, L.L.; Lowing, J.L.; Provenzano, A.M.; Hildebrandt, C.C.; Conti, A.C. Postnatal ethanol exposure simplifies the dendritic morphology of medium spiny neurons independently of adenylyl cyclase 1 and 8 activity in mice. Alcohol. Clin. Exp. Res. 2014, 38, 1339–1346. [Google Scholar] [CrossRef] [PubMed]







| 1.A. AID | |||
| Control | 24h | 7d | |
| Filopodial | 5.497 (0.132) | 5.497 (0.198) | 5.403 (0.135) |
| Apical | 5.466 (0.151) | 5.606 (0.273) | 5.400 (0.157) |
| Basilar | 5.528 (0.182) | 5.388 (0.146) | 5.406 (0.180) |
| Thin * | 5.969 (0.279) | 6.809 (0.456) | 5.519 (0.156) |
| Apical * | 5.984 (0.295) | 6.491 (0.429) | 5.306 (0.120) |
| Basilar # | 5.953 (0.373) | 7.128 (0.541) | 5.731 (0.248) |
| Stubby | 0.148 (0.020) | 0.130 (0.017) | 0.173 (0.044) |
| Apical | 0.153 (0.025) | 0.138 (0.030) | 0.156 (0.038) |
| Basilar | 0.144 (0.037) | 0.122 (0.023) | 0.191 (0.086) |
| Mushroom | 0.311 (0.034) | 0.283 (0.034) | 0.373 (0.027) |
| Apical | 0.250 (0.031) | 0.266 (0.043) | 0.388 (0.049) |
| Basilar | 0.372 (0.047) | 0.300 (0.029) | 0.359 (0.033) |
| Cup-Shaped | 0.102 (0.023) | 0.125 (0.030) | 0.153 (0.019) |
| Apical | 0.088 (0.023) | 0.059 (0.014) | 0.163 (0.032) |
| Basilar | 0.116 (0.032) | 0.191 (0.061) | 0.144 (0.025) |
| Multi-headed | 1.514 (0.162) | 1.544 (0.136) | 1.634 (0.077) |
| Apical | 1.322 (0.131) | 1.338 (0.177) | 1.322 (0.085) |
| Basilar | 1.706 (0.275) | 1.750 (0.137) | 1.947 (0.141) |
| Multi-headed mushroom * | 0.144 (0.044) | 0.053 (0.012) | 0.063 (0.007) |
| Apical | 0.097 (0.037) | 0.056 (0.014) | 0.084 (0.012) |
| Basilar | 0.191 (0.085) | 0.050 (0.023) | 0.041 (0.015) |
| 1.B. PrL | |||
| Control | 24h | 7d | |
| Filopodial | 5.230 (0.378) | 4.964 (0.469) | 5.281 (0.227) |
| Apical | 5.253 (0.148) | 4.931 (0.174) | 5.228 (0.103) |
| Basilar | 5.206 (0.148) | 4.997 (0.209) | 5.334 (0.108) |
| Thin | 6.255 (0.330) | 6.234 (0.484) | 6.139 (0.538) |
| Apical | 6.131 (0.089) | 6.244 (0.199) | 5.994 (0.253) |
| Basilar | 6.378 (0.167) | 6.225 (0.196) | 6.284 (0.156) |
| Stubby * | 0.183 (0.081) | 0.292 (0.099) | 0.322 (0.088) |
| Apical | 0.206 (0.035) | 0.294 (0.042) | 0.328 (0.045) |
| Basilar * | 0.159 (0.025) | 0.291 (0.044) | 0.316 (0.047) |
| Mushroom * | 0.497 (0.099) | 0.497 (0.094) | 0.689 (0.167) |
| Apical * | 0.481 (0.036) | 0.459 (0.037) | 0.772 (0.071) |
| Basilar | 0.513 (0.082) | 0.534 (0.052) | 0.606 (0.063) |
| Cup-Shaped | 0.177 (0.082) | 0.123 (0.052) | 0.181 (0.061) |
| Apical | 0.153 (0.030) | 0.122 (0.023) | 0.172 (0.027) |
| Basilar | 0.200 (0.047) | 0.125 (0.029) | 0.191 (0.045) |
| Multi-headed | 2.081 (0.462) | 1.905 (0.325) | 2.245 (0.513) |
| Apical | 1.956 (0.169) | 1.772 (0.109) | 2.094 (0.154) |
| Basilar | 2.206 (0.198) | 2.038 (0.141) | 2.397 (0.218) |
| Multi-headed mushroom | 0.119 (0.072) | 0.094 (0.130) | 0.200 (0.111) |
| Apical | 0.122 (0.055) | 0.103 (0.045) | 0.188 (0.041) |
| Basilar | 0.116 (0.042) | 0.084 (0.050) | 0.213 (0.082) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frost, M.E.; Peterson, V.L.; Bird, C.W.; McCool, B.; Hamilton, D.A. Effects of Ethanol Exposure and Withdrawal on Neuronal Morphology in the Agranular Insular and Prelimbic Cortices: Relationship with Withdrawal-Related Structural Plasticity in the Nucleus Accumbens. Brain Sci. 2019, 9, 180. https://doi.org/10.3390/brainsci9080180
Frost ME, Peterson VL, Bird CW, McCool B, Hamilton DA. Effects of Ethanol Exposure and Withdrawal on Neuronal Morphology in the Agranular Insular and Prelimbic Cortices: Relationship with Withdrawal-Related Structural Plasticity in the Nucleus Accumbens. Brain Sciences. 2019; 9(8):180. https://doi.org/10.3390/brainsci9080180
Chicago/Turabian StyleFrost, Madeline E., Veronica L. Peterson, Clark W. Bird, Brian McCool, and Derek A. Hamilton. 2019. "Effects of Ethanol Exposure and Withdrawal on Neuronal Morphology in the Agranular Insular and Prelimbic Cortices: Relationship with Withdrawal-Related Structural Plasticity in the Nucleus Accumbens" Brain Sciences 9, no. 8: 180. https://doi.org/10.3390/brainsci9080180