Potential Application of Optogenetic Stimulation in the Treatment of Pain and Migraine Headache: A Perspective from Animal Studies

{kind=link}
Abstract
:1. Introduction
2. The Principle of Optogenetic Manipulation
3. The Applications of Optogenetic Manipulation
3.1. Application in Pain Research
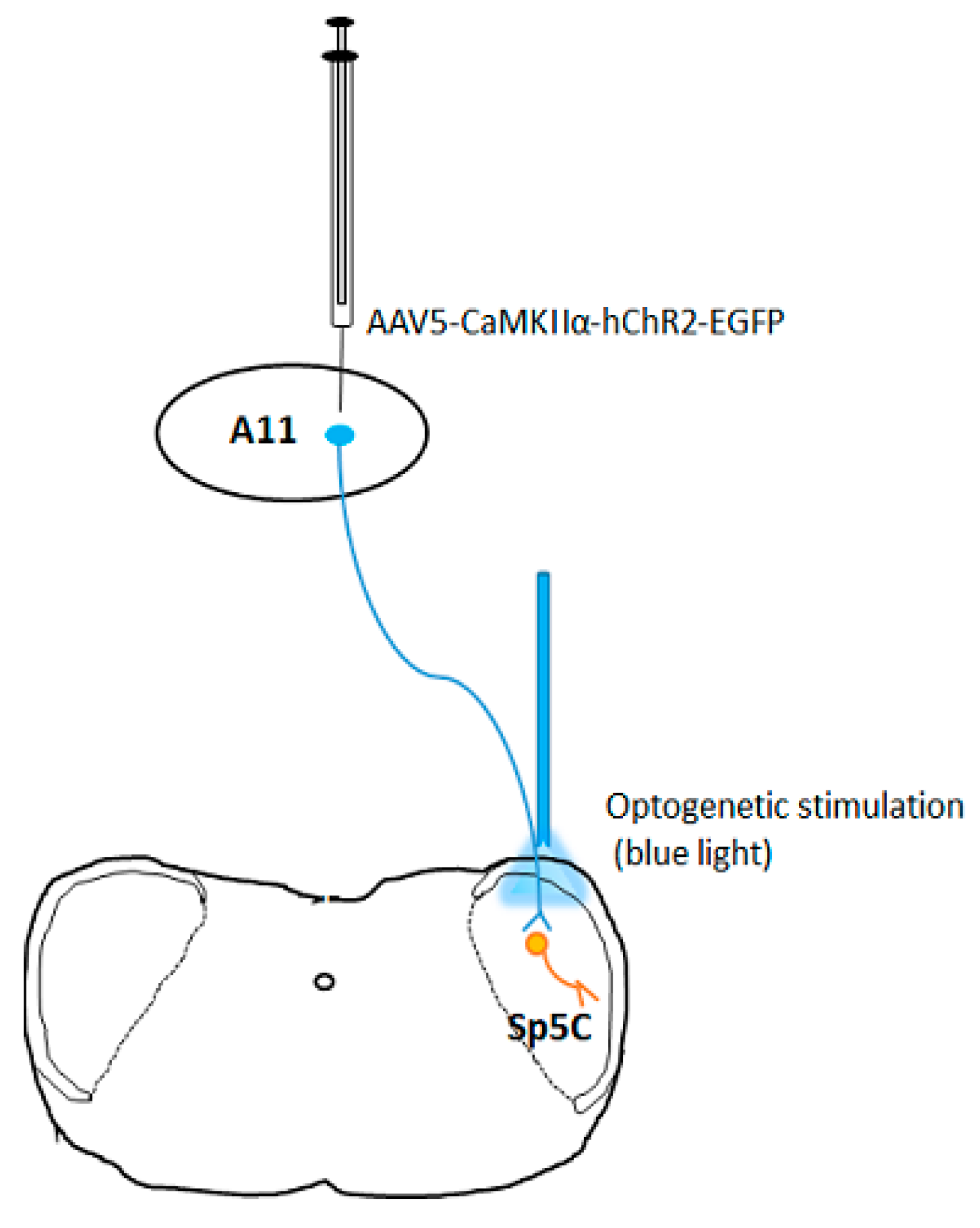
3.2. Potential Application in the Treatment of Migraine Headache
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, CSD, sensitization and modulation of pain. Pain 2013, 154 (Suppl. 1), S44–S53. [Google Scholar] [CrossRef]
- Behbehani, M.M.; Fields, H.L. Evidence that an excitatory connection between the periaqueductal gray and nucleus raphe magnus mediates stimulation produced analgesia. Brain Res. 1979, 170, 85–93. [Google Scholar] [CrossRef]
- Buckett, W.R. Pharmacological studies on stimulation-produced analgesia in mice. Eur. J. Pharmacol. 1981, 69, 281–290. [Google Scholar] [CrossRef]
- Mayer, D.J.; Liebeskind, J.C. Pain reduction by focal electrical stimulation of the brain: An anatomical and behavioral analysis. Brain Res. 1974, 68, 73–93. [Google Scholar] [CrossRef]
- Dasilva, A.F.; Mendonca, M.E.; Zaghi, S.; Lopes, M.; Dossantos, M.F.; Spierings, E.L.; Bajwa, Z.; Datta, A.; Bikson, M.; Fregni, F. tDCS-induced analgesia and electrical fields in pain-related neural networks in chronic migraine. Headache 2012, 52, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Lipton, R.B.; Dodick, D.W.; Silberstein, S.D.; Saper, J.R.; Aurora, S.K.; Pearlman, S.H.; Fischell, R.E.; Ruppel, P.L.; Goadsby, P.J. Single-pulse transcranial magnetic stimulation for acute treatment of migraine with aura: A randomised, double-blind, parallel-group, sham-controlled trial. Lancet Neurol. 2010, 9, 373–380. [Google Scholar] [CrossRef]
- Magis, D.; Schoenen, J. Advances and challenges in neurostimulation for headaches. Lancet Neurol. 2012, 11, 708–719. [Google Scholar] [CrossRef]
- Kalanithi, P.S.; Henderson, J.M. Optogenetic neuromodulation. Int. Rev. Neurobiol. 2012, 107, 185–205. [Google Scholar]
- Yizhar, O.; Fenno, L.E.; Davidson, T.J.; Mogri, M.; Deisseroth, K. Optogenetics in neural systems. Neuron 2011, 71, 9–34. [Google Scholar] [CrossRef]
- Berdyyeva, T.K.; Reynolds, J.H. The dawning of primate optogenetics. Neuron 2009, 62, 159–160. [Google Scholar] [CrossRef]
- Lin, J.Y. A user’s guide to channelrhodopsin variants: Features, limitations and future developments. Exp. Physiol. 2011, 96, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Feldbauer, K.; Zimmermann, D.; Pintschovius, V.; Spitz, J.; Bamann, C.; Bamberg, E. Channelrhodopsin-2 is a leaky proton pump. Proc. Natl. Acad. Sci. USA 2009, 106, 12317–12322. [Google Scholar] [CrossRef] [PubMed]
- Gradinaru, V.; Thompson, K.R.; Zhang, F.; Mogri, M.; Kay, K.; Schneider, M.B.; Deisseroth, K. Targeting and readout strategies for fast optical neural control in vitro and in vivo. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 14231–14238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradinaru, V.; Zhang, F.; Ramakrishnan, C.; Mattis, J.; Prakash, R.; Diester, I.; Goshen, I.; Thompson, K.R.; Deisseroth, K. Molecular and cellular approaches for diversifying and extending optogenetics. Cell 2010, 141, 154–165. [Google Scholar] [CrossRef]
- Li, N.; Downey, J.E.; Bar-Shir, A.; Gilad, A.A.; Walczak, P.; Kim, H.; Joel, S.E.; Pekar, J.J.; Thakor, N.V.; Pelled, G. Optogenetic-guided cortical plasticity after nerve injury. Proc. Natl. Acad. Sci. USA 2011, 108, 8838–8843. [Google Scholar] [CrossRef]
- Mei, Y.; Zhang, F. Molecular tools and approaches for optogenetics. Biol. Psychiatry 2012, 71, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Henckaerts, E.; Dutheil, N.; Zeltner, N.; Kattman, S.; Kohlbrenner, E.; Ward, P.; Clement, N.; Rebollo, P.; Kennedy, M.; Keller, G.M.; et al. Site-specific integration of adeno-associated virus involves partial duplication of the target locus. Proc. Natl. Acad. Sci. USA 2009, 106, 7571–7576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diester, I.; Kaufman, M.T.; Mogri, M.; Pashaie, R.; Goo, W.; Yizhar, O.; Ramakrishnan, C.; Deisseroth, K.; Shenoy, K.V. An optogenetic toolbox designed for primates. Nat. Neurosci. 2011, 14, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Airan, R.D.; Li, N.; Gilad, A.A.; Pelled, G. Genetic tools to manipulate MRI contrast. NMR Biomed. 2013, 26, 7. [Google Scholar] [CrossRef]
- Aravanis, A.M.; Wang, L.P.; Zhang, F.; Meltzer, L.A.; Mogri, M.Z.; Schneider, M.B.; Deisseroth, K. An optical neural interface: In vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J. Neural Eng. 2007, 4, S143–S156. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, E.; Saper, C.B. What optogenetic stimulation is telling us (and failing to tell us) about fast neurotransmitters and neuromodulators in brain circuits for wake-sleep regulation. Curr. Opin. Neurobiol. 2014, 29C, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, C.E.; Grinevich, V.P.; Gioia, D.; Day-Brown, J.D.; Bonin, K.D.; Stuber, G.D.; Weiner, J.L.; Budygin, E.A. Optogenetic stimulation of VTA dopamine neurons reveals that tonic but not phasic patterns of dopamine transmission reduce ethanol self-administration. Front. Behav. Neurosci. 2013, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Cardin, J.A.; Carlen, M.; Meletis, K.; Knoblich, U.; Zhang, F.; Deisseroth, K.; Tsai, L.H.; Moore, C.I. Targeted optogenetic stimulation and recording of neurons in vivo using cell-type-specific expression of Channelrhodopsin-2. Nat. Protoc. 2010, 5, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Stroh, A.; Diester, I. Optogenetics: A new method for the causal analysis of neuronal networks in vivo. e-Neuroforum 2012, 3, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Crock, L.W.; Kolber, B.J.; Morgan, C.D.; Sadler, K.E.; Vogt, S.K.; Bruchas, M.R.; Gereau, R.W. Central amygdala metabotropic glutamate receptor 5 in the modulation of visceral pain. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 14217–14226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barish, P.A.; Xu, Y.; Li, J.; Sun, J.; Jarajapu, Y.P.; Ogle, W.O. Design and functional evaluation of an optically active mu-opioid receptor. Eur. J. Pharmacol. 2013, 705, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo, O.; Friedrich, R.W. Optopharmacology: A light switch for pain. Nat. Chem. Biol. 2013, 9, 219–220. [Google Scholar] [CrossRef]
- Mourot, A.; Fehrentz, T.; Le Feuvre, Y.; Smith, C.M.; Herold, C.; Dalkara, D.; Nagy, F.; Trauner, D.; Kramer, R.H. Rapid optical control of nociception with an ion-channel photoswitch. Nat. Methods 2012, 9, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Kokel, D.; Cheung, C.Y.; Mills, R.; Coutinho-Budd, J.; Huang, L.; Setola, V.; Sprague, J.; Jin, S.; Jin, Y.N.; Huang, X.P.; et al. Photochemical activation of TRPA1 channels in neurons and animals. Nat. Chem. Biol. 2013, 9, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Chambers, J.J.; Banghart, M.R.; Trauner, D.; Kramer, R.H. Light-induced depolarization of neurons using a modified Shaker K(+) channel and a molecular photoswitch. J. Neurophysiol. 2006, 96, 2792–2796. [Google Scholar] [CrossRef]
- Fortin, D.L.; Dunn, T.W.; Fedorchak, A.; Allen, D.; Montpetit, R.; Banghart, M.R.; Trauner, D.; Adelman, J.P.; Kramer, R.H. Optogenetic photochemical control of designer K+ channels in mammalian neurons. J. Neurophysiol. 2011, 106, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Daou, I.; Tuttle, A.H.; Longo, G.; Wieskopf, J.S.; Bonin, R.P.; Ase, A.R.; Wood, J.N.; De Koninck, Y.; Ribeiro-da-Silva, A.; Mogil, J.S.; et al. Remote optogenetic activation and sensitization of pain pathways in freely moving mice. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 18631–18640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, S.M.; Montgomery, K.L.; Towne, C.; Lee, S.Y.; Ramakrishnan, C.; Deisseroth, K.; Delp, S.L. Virally mediated optogenetic excitation and inhibition of pain in freely moving nontransgenic mice. Nat. Biotechnol. 2014, 32, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Manders, T.R.; Eberle, S.E.; Su, C.; D’Amour, J.; Yang, R.; Lin, H.Y.; Deisseroth, K.; Froemke, R.C.; Wang, J. Activation of corticostriatal circuitry relieves chronic neuropathic pain. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 5247–5259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gadotti, V.M.; Chen, L.; Souza, I.A.; Stemkowski, P.L.; Zamponi, G.W. Role of Prelimbic GABAergic Circuits in Sensory and Emotional Aspects of Neuropathic Pain. Cell Rep. 2015, 12, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Kiritoshi, T.; Ji, G.; Neugebauer, V. Rescue of Impaired mGluR5-Driven Endocannabinoid Signaling Restores Prefrontal Cortical Output to Inhibit Pain in Arthritic Rats. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Q.; Cen, C.; Li, C.; Cao, S.; Wang, N.; Zhou, Z.; Liu, X.M.; Xu, Y.; Tian, N.X.; Zhang, Y.; et al. Deactivation of excitatory neurons in the prelimbic cortex via Cdk5 promotes pain sensation and anxiety. Nat. Commun. 2015, 6, 7660. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tang, Y.; Shu, H.; Tatum, D.; Bai, Q.; Crawford, J.; Xing, Y.; Lobo, M.K.; Bellinger, L.; Kramer, P.; et al. Dopamine receptor D2, but not D1, mediates descending dopaminergic pathway-produced analgesic effect in a trigeminal neuropathic pain mouse model. Pain 2018. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, M.; Dreier, J.P.; Fabricius, M.; Hartings, J.A.; Graf, R.; Strong, A.J. Clinical relevance of cortical spreading depression in neurological disorders: Migraine, malignant stroke, subarachnoid and intracranial hemorrhage; traumatic brain injury. J. Cerebral Blood Flow Metab. 2011, 31, 17–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayata, C.; Jin, H.; Kudo, C.; Dalkara, T.; Moskowitz, M.A. Suppression of cortical spreading depression in migraine prophylaxis. Ann. Neurol. 2006, 59, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Tozzi, A.; Rainero, I.; Cupini, L.M.; Calabresi, P.; Ayata, C.; Sarchielli, P. Cortical spreading depression as a target for anti-migraine agents. J. Headache Pain 2013, 14, 62. [Google Scholar] [CrossRef]
- Eikermann-Haerter, K.; Can, A.; Ayata, C. Pharmacological targeting of spreading depression in migraine. Expert Rev. Neurother. 2012, 12, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Arenkiel, B.R.; Peca, J.; Davison, I.G.; Feliciano, C.; Deisseroth, K.; Augustine, G.J.; Ehlers, M.D.; Feng, G. In vivo light-induced activation of neural circuitry in transgenic mice expressing channelrhodopsin-2. Neuron 2007, 54, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolner, E.A.; Houben, T.; Terwindt, G.M.; de Vries, B.; Ferrari, M.D.; van den Maagdenberg, A.M. From migraine genes to mechanisms. Pain 2015, 156, S64–S74. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Tang, Y.; Xing, Y.; Kramer, P.; Bellinger, L.; Tao, F. Potential Application of Optogenetic Stimulation in the Treatment of Pain and Migraine Headache: A Perspective from Animal Studies. Brain Sci. 2019, 9, 26. https://doi.org/10.3390/brainsci9020026
Liu S, Tang Y, Xing Y, Kramer P, Bellinger L, Tao F. Potential Application of Optogenetic Stimulation in the Treatment of Pain and Migraine Headache: A Perspective from Animal Studies. Brain Sciences. 2019; 9(2):26. https://doi.org/10.3390/brainsci9020026
Chicago/Turabian StyleLiu, Sufang, Yuanyuan Tang, Ying Xing, Phillip Kramer, Larry Bellinger, and Feng Tao. 2019. "Potential Application of Optogenetic Stimulation in the Treatment of Pain and Migraine Headache: A Perspective from Animal Studies" Brain Sciences 9, no. 2: 26. https://doi.org/10.3390/brainsci9020026