
Maintained Spatial Learning and Memory Functions in Middle-Aged α9 Nicotinic Receptor Subunit Knock-Out Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
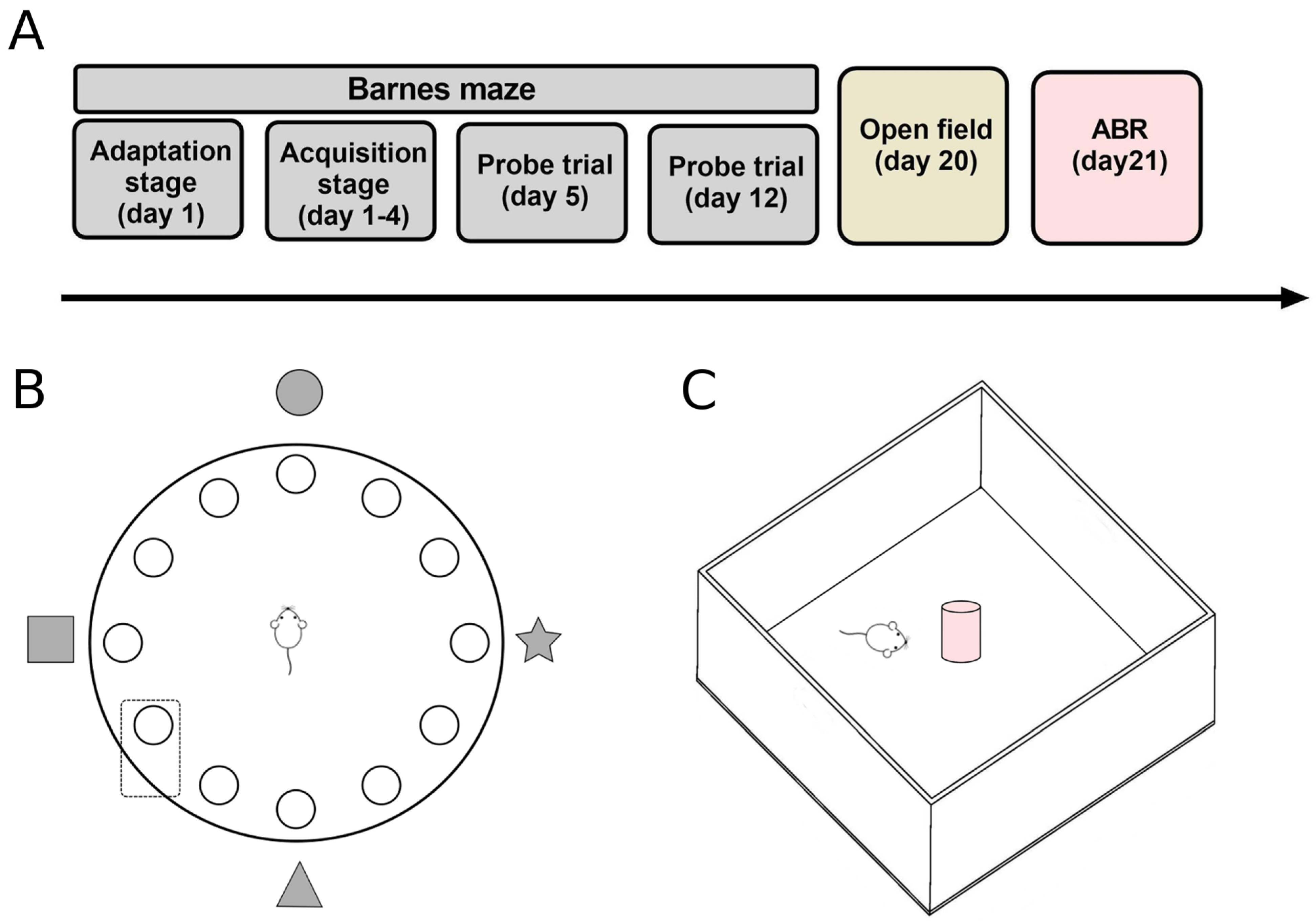
2.2. Barnes Maze
2.2.1. Adaptation Stage
2.2.2. Acquisition Stage
2.2.3. Probe Trial
2.3. Open Field Test
2.4. Electrophysiology
2.5. Immunostaining Analysis
2.6. Statistical Analysis
3. Results
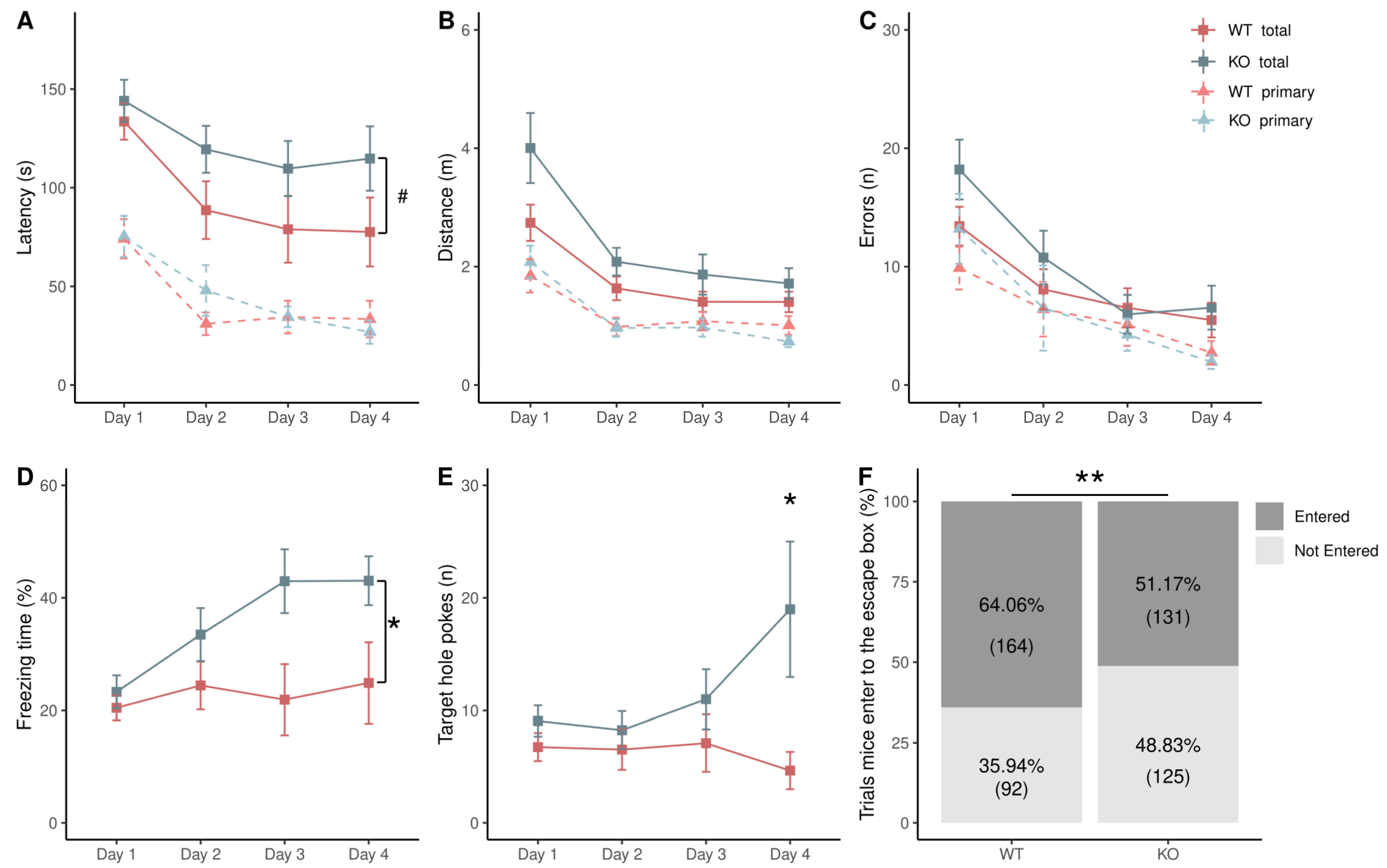
3.1. Difference in Acquisition Stage
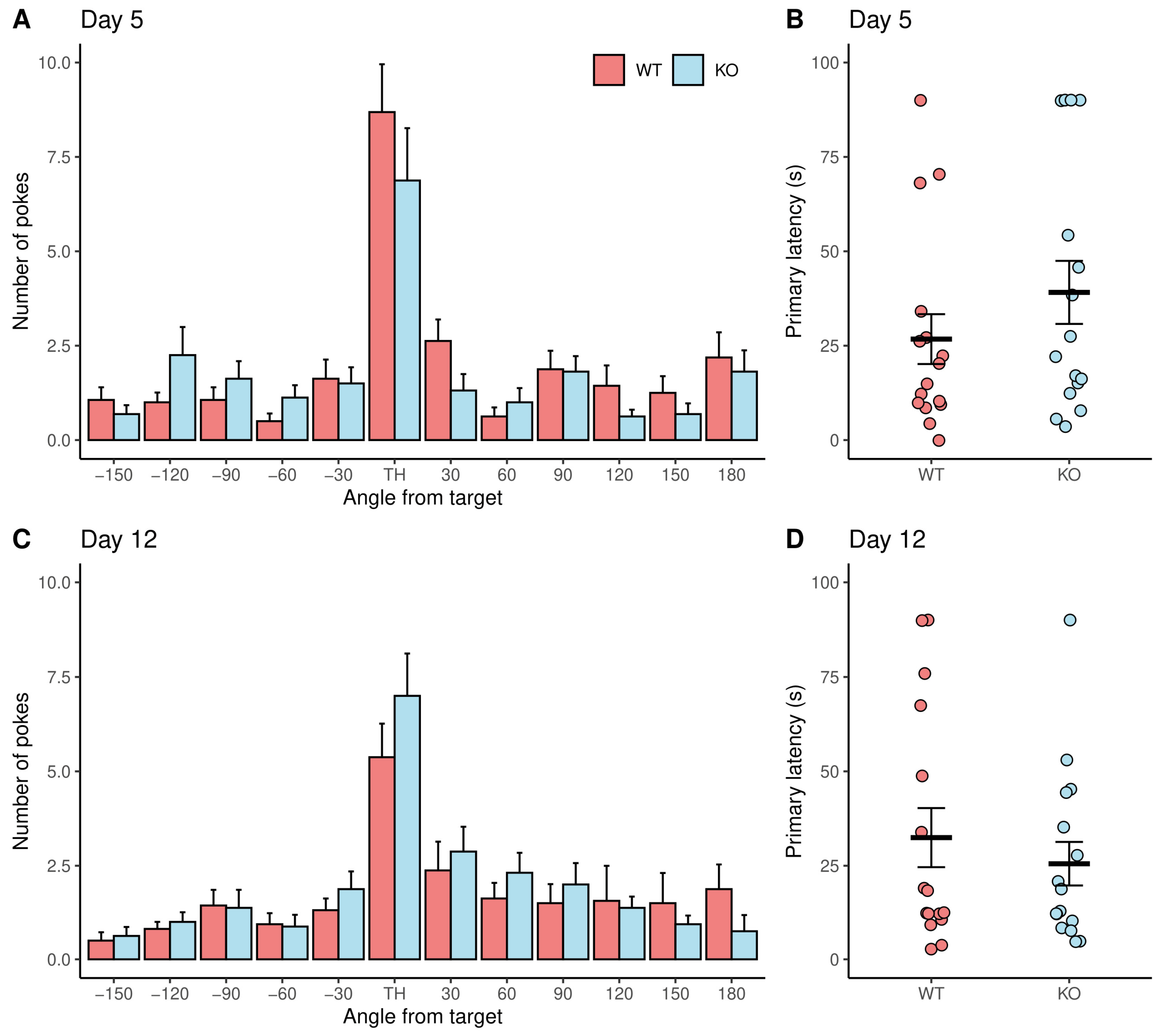
3.2. No Difference between Genotypes in Memory
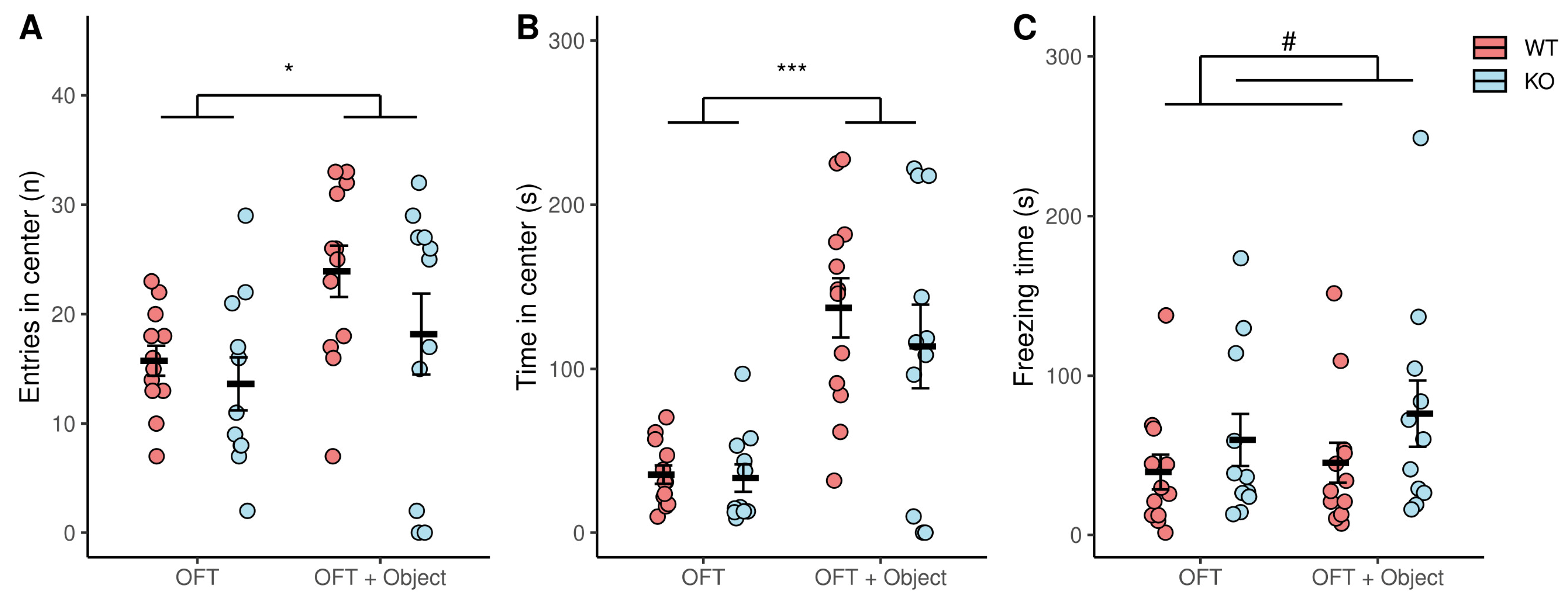
3.3. Novelty-Induced Behavior
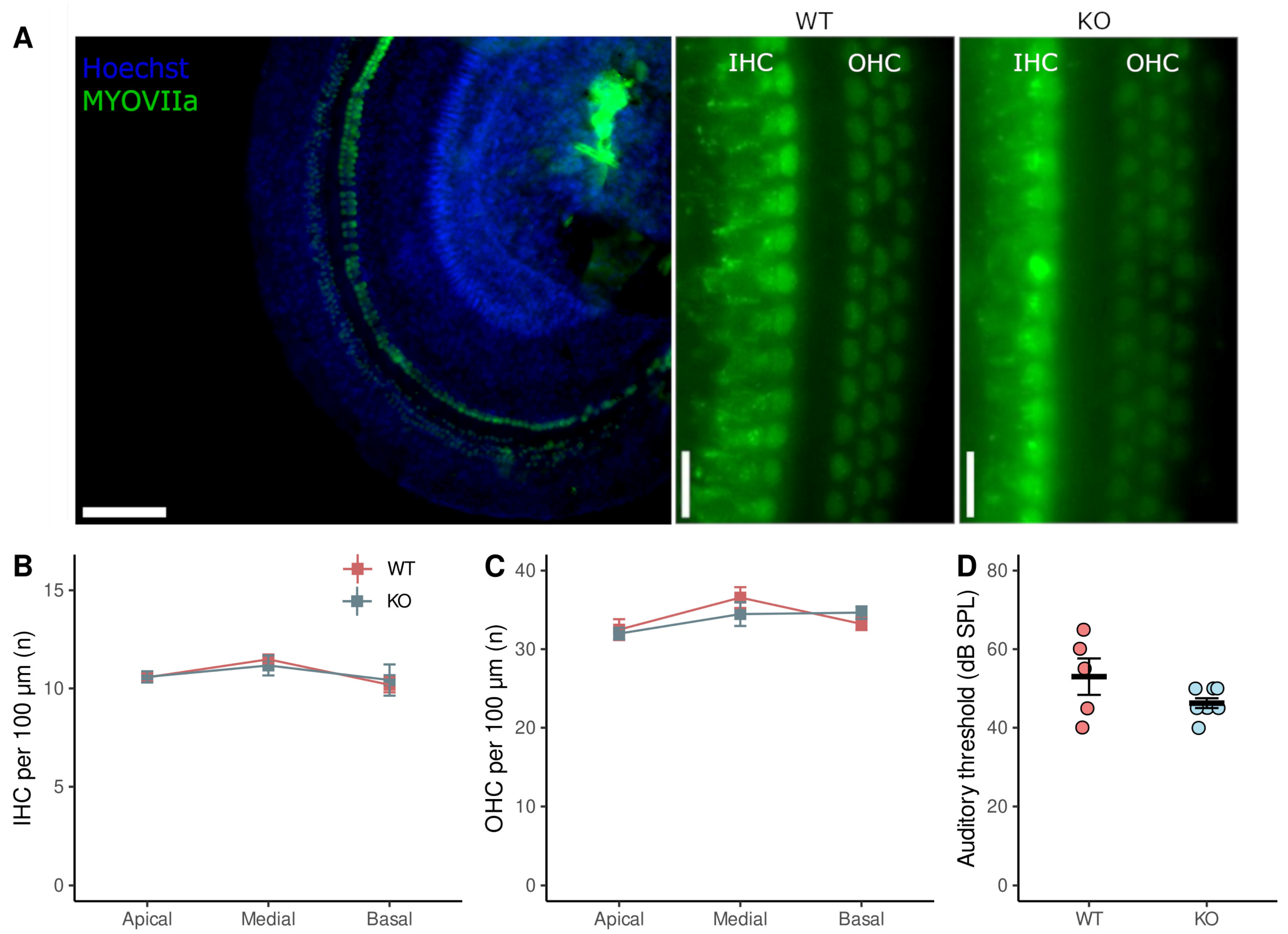
3.4. ABR Threshold and Hair Cells Number
4. Discussion
4.1. No Difference in Spatial Learning and Memory in α9 KO
4.2. Altered Response to Stress and Motivation for Reward in KO Mice
4.3. Auditory and Vestibular Function Related to Cognitive Impairment
4.4. Limitations in the Behavioral Study of KO Mice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. 2022 Alzheimer’s disease facts and figures. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2022, 18, 700–789. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, S.A.M.; van Boxtel, M.P.J.; van Hooren, S.A.H.; Bosma, H.; Beckers, H.J.M.; Ponds, R.W.H.M.; Jolles, J. Change in sensory functioning predicts change in cognitive functioning: Results from a 6-year follow-up in the Maastricht Aging Study. J. Am. Geriatr. Soc. 2005, 53, 374–380. [Google Scholar] [CrossRef]
- Lin, F.R.; Yaffe, K.; Xia, J.; Xue, Q.L.; Harris, T.B.; Purchase-Helzner, E.; Satterfield, S.; Ayonayon, H.N.; Ferrucci, L.; Simonsick, E.M.; et al. Hearing Loss and Cognitive Decline in Older Adults. Jama Intern. Med. 2013, 173, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.R. Hearing Loss and Cognition Among Older Adults in the United States. J. Gerontol. A Biol. 2011, 66, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, R.F.; Larson, E.B.; Rees, T.S.; Koepsell, T.D.; Duckert, L.G. Relationship of Hearing Impairment to Dementia and Cognitive Dysfunction in Older Adults. JAMA J. Am. Med. Assoc. 1989, 261, 1916–1919. [Google Scholar] [CrossRef]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, J.; Burns, A.; Cohen-Mansfield, J.; Cooper, C.; et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef]
- Panza, F.; Solfrizzi, V.; Seripa, D.; Imbimbo, B.P.; Capozzo, R.; Quaranta, N.; Pilotto, A.; Logroscino, G. Age-related hearing impairment and frailty in Alzheimer’s disease: Interconnected associations and mechanisms. Front. Aging Neurosci. 2015, 7, 113. [Google Scholar] [CrossRef]
- Huang, A.R.; Jiang, K.; Lin, F.R.; Deal, J.A.; Reed, N.S. Hearing Loss and Dementia Prevalence in Older Adults in the US. JAMA 2023, 329, 171–173. [Google Scholar] [CrossRef]
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120. [Google Scholar] [CrossRef]
- Taljaard, D.S.; Olaithe, M.; Brennan-Jones, C.G.; Eikelboom, R.H.; Bucks, R.S. The relationship between hearing impairment and cognitive function: A meta-analysis in adults. Clin. Otolaryngol. 2016, 41, 718–729. [Google Scholar] [CrossRef]
- Panza, F.; Lozupone, M.; Sardone, R.; Battista, P.; Piccininni, M.; Dibello, V.; La Montagna, M.; Stallone, R.; Venezia, P.; Liguori, A.; et al. Sensorial frailty: Age-related hearing loss and the risk of cognitive impairment and dementia in later life. Ther. Adv. Chronic Dis. 2019, 10, 2040622318811000. [Google Scholar] [CrossRef]
- Johnson, J.C.S.; Marshall, C.R.; Weil, R.S.; Bamiou, D.-E.; Hardy, C.J.D.; Warren, J.D. Hearing and dementia: From ears to brain. Brain 2021, 144, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, T.D.; Lad, M.; Kumar, S.; Holmes, E.; McMurray, B.; Maguire, E.A.; Billig, A.J.; Sedley, W. How can hearing loss cause dementia? Neuron 2020, 108, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Belkhiria, C.; Vergara, R.C.; Martinez, M.; Delano, P.H.; Delgado, C. Neural links between facial emotion recognition and cognitive impairment in presbycusis. Int. J. Geriatr. Psychiatry 2021, 36, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Delano, P.H.; Belkhiria, C.; Vergara, R.C.; Martínez, M.; Leiva, A.; Andrade, M.; Marcenaro, B.; Torrente, M.; Maass, J.C.; Delgado, C. Reduced suprathreshold auditory nerve responses are associated with slower processing speed and thinner temporal and parietal cortex in presbycusis. PLoS ONE 2020, 15, e0233224. [Google Scholar] [CrossRef] [PubMed]
- Belkhiria, C.; Vergara, R.C.; San Martin, S.; Leiva, A.; Martinez, M.; Marcenaro, B.; Andrade, M.; Delano, P.H.; Delgado, C. Insula and Amygdala Atrophy Are Associated with Functional Impairment in Subjects with Presbycusis. Front. Aging Neurosci. 2020, 12, 102. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, M.J.; Sikandaner, H.; Kim, D.K.; Yeo, S.W.; Park, S.N. A causal relationship between hearing loss and cognitive impairment. Acta Oto-Laryngol. 2016, 136, 480–483. [Google Scholar] [CrossRef]
- Liu, L.J.; Shen, P.; He, T.T.; Chang, Y.; Shi, L.J.; Tao, S.; Li, X.W.; Xun, Q.Y.; Guo, X.J.; Yu, Z.P.; et al. Noise induced hearing loss impairs spatial learning/memory and hippocampal neurogenesis in mice. Sci. Rep. 2016, 6, 20374. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, M.J.; Kim, H.L.; Kim, D.K.; Yeo, S.W.; Park, S.N. Cognitive decline and increased hippocampal p-tau expression in mice with hearing loss. Behav. Brain Res. 2018, 342, 19–26. [Google Scholar] [CrossRef]
- Paciello, F.; Rinaudo, M.; Longo, V.; Cocco, S.; Conforto, G.; Pisani, A.; Podda, M.V.; Fetoni, A.R.; Paludetti, G.; Grassi, C. Auditory sensory deprivation induced by noise exposure exacerbates cognitive decline in a mouse model of Alzheimer’s disease. eLife 2021, 10, e70908. [Google Scholar] [CrossRef]
- Yu, Y.F.; Zhai, F.; Dai, C.F.; Hu, J.J. The relationship between age-related hearing loss and synaptic changes in the hippocampus of C57BL/6J mice. Exp. Gerontol. 2011, 46, 716–722. [Google Scholar] [CrossRef]
- Liberman, M.C.; Liberman, L.D.; Maison, S.F. Efferent feedback slows cochlear aging. J. Neurosci. 2014, 34, 4599–4607. [Google Scholar] [CrossRef] [PubMed]
- Boero, L.E.; Castagna, V.C.; Terreros, G.; Moglie, M.J.; Silva, S.; Maass, J.C.; Fuchs, P.A.; Delano, P.H.; Elgoyhen, A.B.; Gómez-Casati, M.E. Preventing presbycusis in mice with enhanced medial olivocochlear feedback. Proc. Natl. Acad. Sci. USA 2020, 117, 11811–11819. [Google Scholar] [CrossRef] [PubMed]
- Vicencio-Jimenez, S.; Weinberg, M.M.; Bucci-Mansilla, G.; Lauer, A.M. Olivocochlear Changes Associated with Aging Predominantly Affect the Medial Olivocochlear System. Front. Neurosci. 2021, 15, 704805. [Google Scholar] [CrossRef]
- Terreros, G.; Jorratt, P.; Aedo, C.; Elgoyhen, A.B.; Delano, P.H. Selective Attention to Visual Stimuli Using Auditory Distractors Is Altered in Alpha-9 Nicotinic Receptor Subunit Knock-Out Mice. J. Neurosci. 2016, 36, 7198–7209. [Google Scholar] [CrossRef] [PubMed]
- Terreros, G.; Delano, P.H. Corticofugal modulation of peripheral auditory responses. Front. Syst. Neurosci. 2015, 9, 134. [Google Scholar] [CrossRef]
- Katz, E.; Elgoyhen, A.B. Short-term plasticity and modulation of synaptic transmission at mammalian inhibitory cholinergic olivocochlear synapses. Front. Syst. Neurosci. 2014, 8, 224. [Google Scholar] [CrossRef]
- Hiel, H.; Elgoyhen, A.B.; Drescher, D.G.; Morley, B.J. Expression of nicotinic acetylcholine receptor mRNA in the adult rat peripheral vestibular system. Brain Res. 1996, 738, 347–352. [Google Scholar] [CrossRef]
- Alvarez-Munoz, H.; Vicencio-Jimenez, S.; Jorratt, P.; Delano, P.H.; Terreros, G. Corticofugal and Brainstem Functions Associated With Medial Olivocochlear Cholinergic Transmission. Front. Neurosci. 2022, 16, 866161. [Google Scholar] [CrossRef]
- Jorratt, P.; Delano, P.H.; Delgado, C.; Dagnino-Subiabre, A.; Terreros, G. Difference in Perseverative Errors during a Visual Attention Task with Auditory Distractors in Alpha-9 Nicotinic Receptor Subunit Wild Type and Knock-Out Mice. Front. Cell. Neurosci. 2017, 11, 357. [Google Scholar] [CrossRef] [PubMed]
- Sunyer, B.; Patil, S.; Höger, H.; Lubec, G. Barnes maze, a useful task to assess spatial reference memory in the mice. Protoc. Exch. 2007. [Google Scholar] [CrossRef]
- Schuetze, S.; Manig, A.; Ribes, S.; Nau, R. Aged mice show an increased mortality after anesthesia with a standard dose of ketamine/xylazine. Lab. Anim. Res. 2019, 35, 8. [Google Scholar] [CrossRef]
- Rosenfeld, C.S.; Ferguson, S.A. Barnes Maze Testing Strategies with Small and Large Rodent Models. Jove J. Vis. Exp. 2014, 84, e51194. [Google Scholar]
- Harrison, F.E.; Reiserer, R.S.; Tomarken, A.J.; McDonald, M.P. Spatial and nonspatial escape strategies in the Barnes maze. Learn. Mem. 2006, 13, 809–819. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Filarowska, J.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodentsmethodological consideration. Naunyn-Schmiedebergs Arch. Pharmacol. 2019, 392, 1–18. [Google Scholar] [CrossRef]
- Grailhe, R.; Waeber, C.; Dulawa, S.C.; Hornung, J.P.; Zhuang, X.X.; Brunner, D.; Geyer, M.A.; Hen, R. Increased exploratory activity and altered response to LSD in mice lacking the 5-HT5A receptor. Neuron 1999, 22, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.A.; Burton, T.J.; Christie, M.J. alpha 9-nAChR knockout mice exhibit dysregulation of stress responses, affect and reward-related behaviour. Behav. Brain Res. 2017, 328, 105–114. [Google Scholar] [CrossRef]
- Harrison, F.E.; Hosseini, A.H.; McDonald, M.P. Endogenous anxiety and stress responses in water maze and Barnes maze spatial memory tasks. Behav. Brain Res. 2009, 198, 247–251. [Google Scholar] [CrossRef]
- Taranda, J.; Maison, S.F.; Ballestero, J.A.; Katz, E.; Savino, J.; Vetter, D.E.; Boulter, J.; Liberman, M.C.; Fuchs, P.A.; Elgoyhen, A.B. A Point Mutation in the Hair Cell Nicotinic Cholinergic Receptor Prolongs Cochlear Inhibition and Enhances Noise Protection. PLoS Biol. 2009, 7, 71–83. [Google Scholar] [CrossRef]
- Maison, S.F.; Liberman, M.C. Predicting vulnerability to acoustic injury with a noninvasive assay of olivocochlear reflex strength. J. Neurosci. 2000, 20, 4701–4707. [Google Scholar] [CrossRef] [PubMed]
- Boero, L.E.; Castagna, V.C.; Di Guilmi, M.N.; Goutman, J.D.; Elgoyhen, A.B.; Gomez-Casati, M.E. Enhancement of the Medial Olivocochlear System Prevents Hidden Hearing Loss. J. Neurosci. 2018, 38, 7440–7451. [Google Scholar] [CrossRef]
- Lauer, A.M. Minimal Effects of Age and Exposure to a Noisy Environment on Hearing in Alpha9 Nicotinic Receptor Knockout Mice. Front. Neurosci. 2017, 11, 304. [Google Scholar] [CrossRef]
- Schaette, R.; McAlpine, D. Tinnitus with a Normal Audiogram: Physiological Evidence for Hidden Hearing Loss and Computational Model. J. Neurosci. 2011, 31, 13452–13457. [Google Scholar] [CrossRef]
- Lips, K.S.; Pfeil, U.; Kummer, W. Coexpression of alpha 9 and alpha 10 nicotinic acetylcholine receptors in rat dorsal root ganglion neurons. Neuroscience 2002, 115, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Elgoyhen, A.B.; Johnson, D.S.; Boulter, J.; Vetter, D.E.; Heinemann, S. Alpha-9—An Acetylcholine-Receptor with Novel Pharmacological Properties Expressed in Rat Cochlear Hair-Cells. Cell 1994, 79, 705–715. [Google Scholar] [CrossRef]
- Colomer, C.; Olivos-Ore, L.A.; Vincent, A.; McIntosh, J.M.; Artalejo, A.R.; Guerineau, N.C. Functional Characterization of alpha 9-Containing Cholinergic Nicotinic Receptors in the Rat Adrenal Medulla: Implication in Stress-Induced Functional Plasticity. J. Neurosci. 2010, 30, 6732–6742. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.S.; Ferris, R.L.; Matthews, T.; Hiel, H.; Lopez-Albaitero, A.; Lustig, L.R. Characterization of the human nicotinic acetylcholine receptor subunit alpha (alpha) 9 (CHRNA9) and alpha (alpha) 10 (CHRNAIO) in lymphocytes. Life Sci. 2004, 76, 263–280. [Google Scholar] [CrossRef]
- Lykhmus, O.; Voytenko, L.P.; Lips, K.S.; Bergen, I.; Krasteva-Christ, G.; Vetter, D.E.; Kummer, W.; Skok, M. Nicotinic Acetylcholine Receptor alpha 9 and alpha 10 Subunits Are Expressed in the Brain of Mice. Front. Cell Neurosci. 2017, 11, 282. [Google Scholar] [CrossRef]
- Morley, B.J.; Whiteaker, P.; Elgoyhen, A.B. Commentary: Nicotinic Acetylcholine Receptor alpha 9 and alpha 10 Subunits Are Expressed in the Brain of Mice. Front. Cell Neurosci. 2018, 12, 104. [Google Scholar] [CrossRef]
- Sergeyenko, Y.; Lall, K.; Liberman, M.C.; Kujawa, S.G. Age-related cochlear synaptopathy: An early-onset contributor to auditory functional decline. J. Neurosci. 2013, 33, 13686–13694. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicencio-Jimenez, S.; Delano, P.H.; Madrid, N.; Terreros, G.; Maass, J.C.; Delgado, C.; Jorratt, P. Maintained Spatial Learning and Memory Functions in Middle-Aged α9 Nicotinic Receptor Subunit Knock-Out Mice. Brain Sci. 2023, 13, 794. https://doi.org/10.3390/brainsci13050794
Vicencio-Jimenez S, Delano PH, Madrid N, Terreros G, Maass JC, Delgado C, Jorratt P. Maintained Spatial Learning and Memory Functions in Middle-Aged α9 Nicotinic Receptor Subunit Knock-Out Mice. Brain Sciences. 2023; 13(5):794. https://doi.org/10.3390/brainsci13050794
Chicago/Turabian StyleVicencio-Jimenez, Sergio, Paul H. Delano, Natalia Madrid, Gonzalo Terreros, Juan C. Maass, Carolina Delgado, and Pascal Jorratt. 2023. "Maintained Spatial Learning and Memory Functions in Middle-Aged α9 Nicotinic Receptor Subunit Knock-Out Mice" Brain Sciences 13, no. 5: 794. https://doi.org/10.3390/brainsci13050794