Effects of Paternal Preconception Vapor Alcohol Exposure Paradigms on Behavioral Responses in Offspring



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
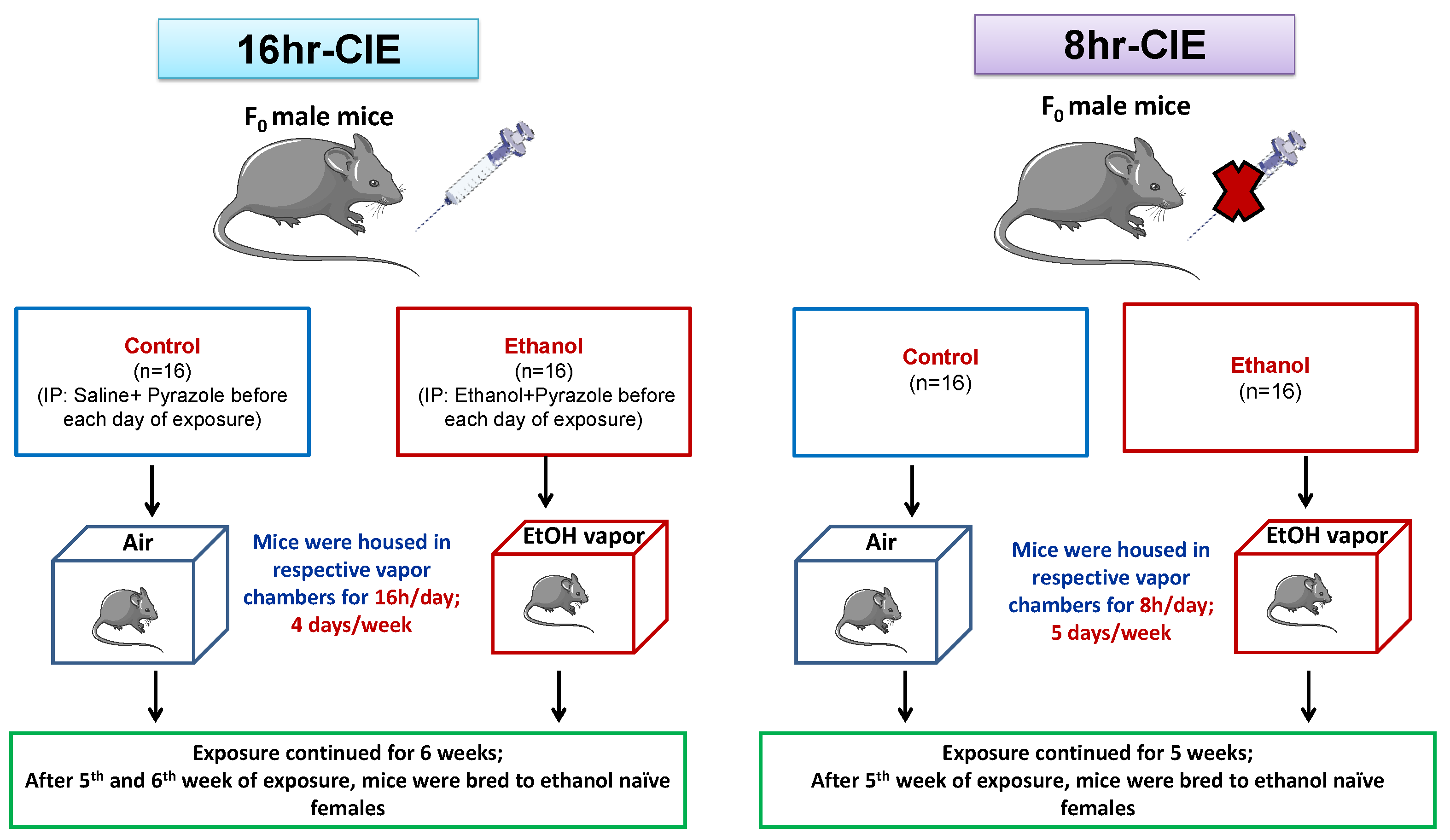
2.2. Chronic Intermittent Ethanol (CIE) Inhalation
2.3. Breeding Scheme
2.4. Behavioral Experiments
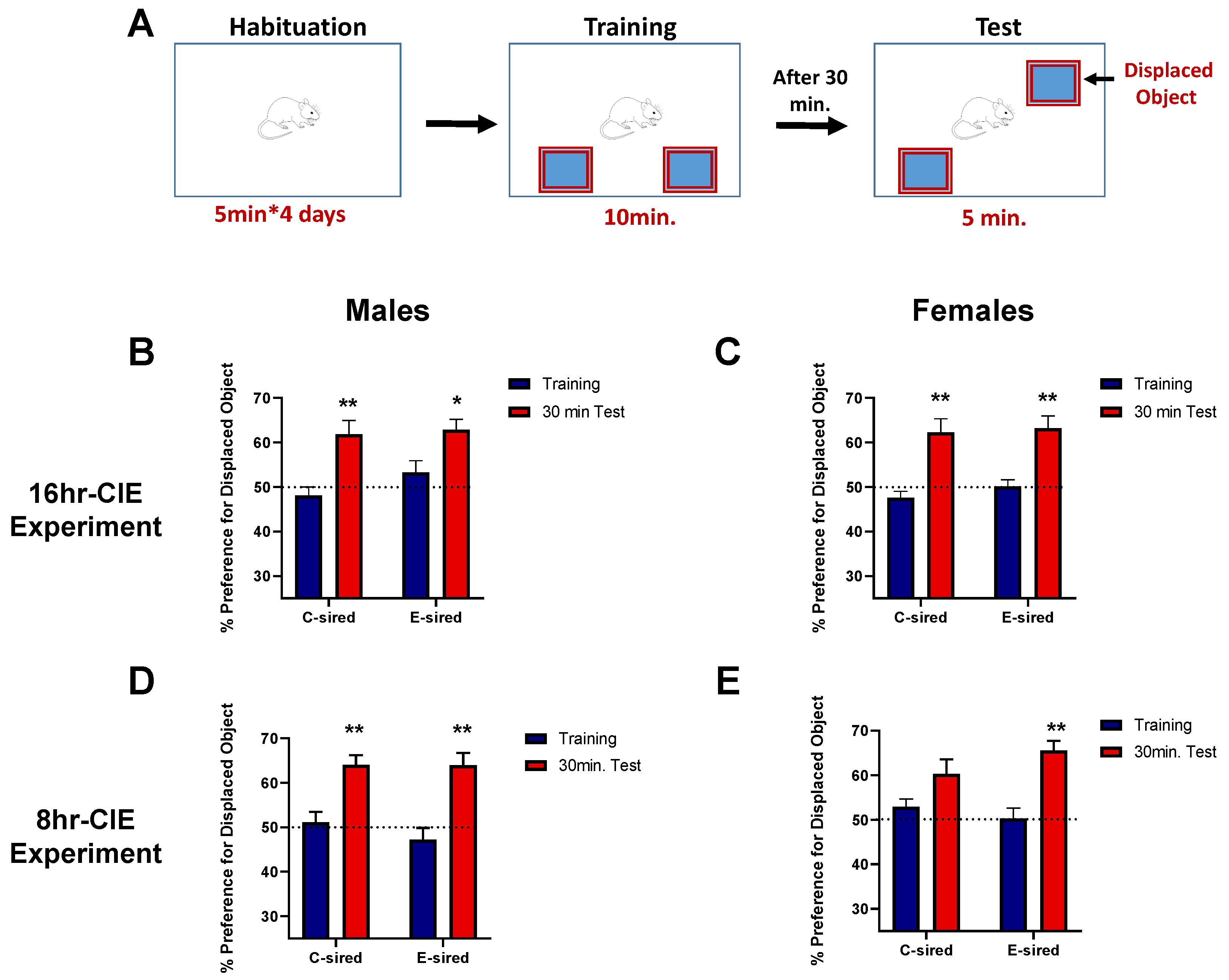
2.4.1. Object Location Memory Task
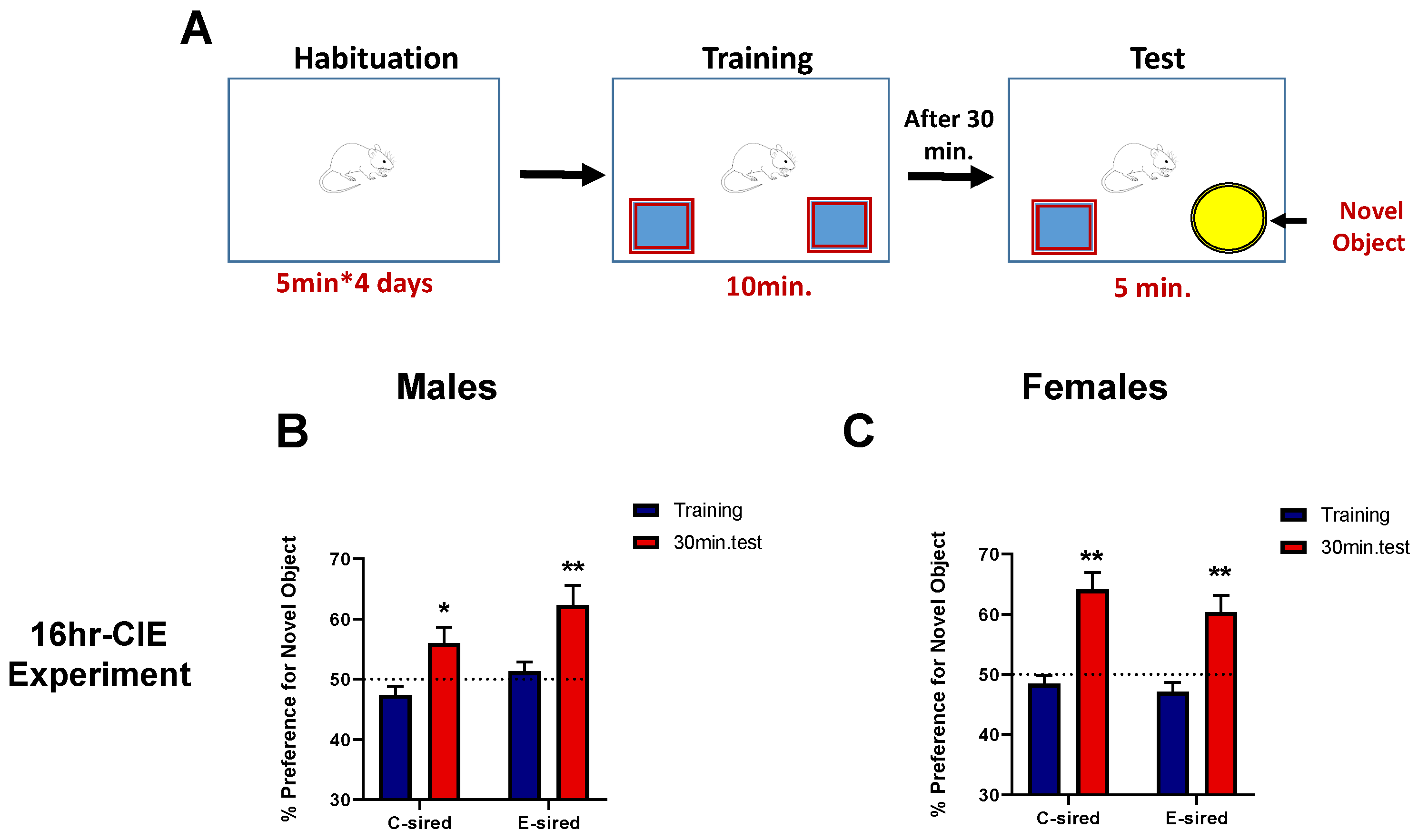
2.4.2. Novel Object Recognition Memory Task
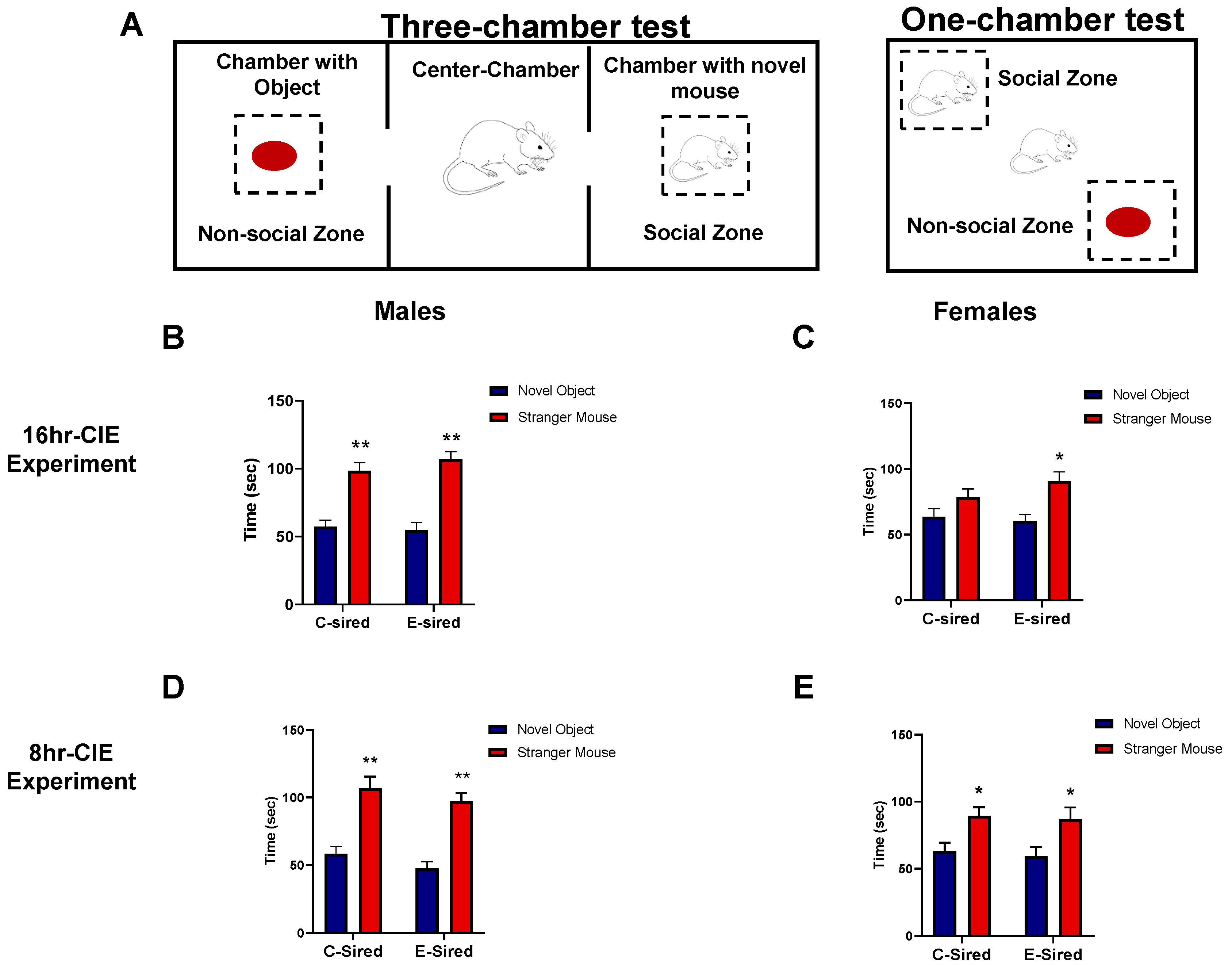
2.4.3. Social Interaction Test
2.4.4. Loss of Righting Response (LORR)
2.4.5. Acute Functional Tolerance Test (AFT)
2.4.6. Stress-Induced Hyperthermia (SIH)
2.4.7. Drinking in the Dark (DID) Assay
2.4.8. Every-Other-Day Two-Bottle Choice (EOD 2BC) Assay
2.5. Statistical Analysis
3. Results
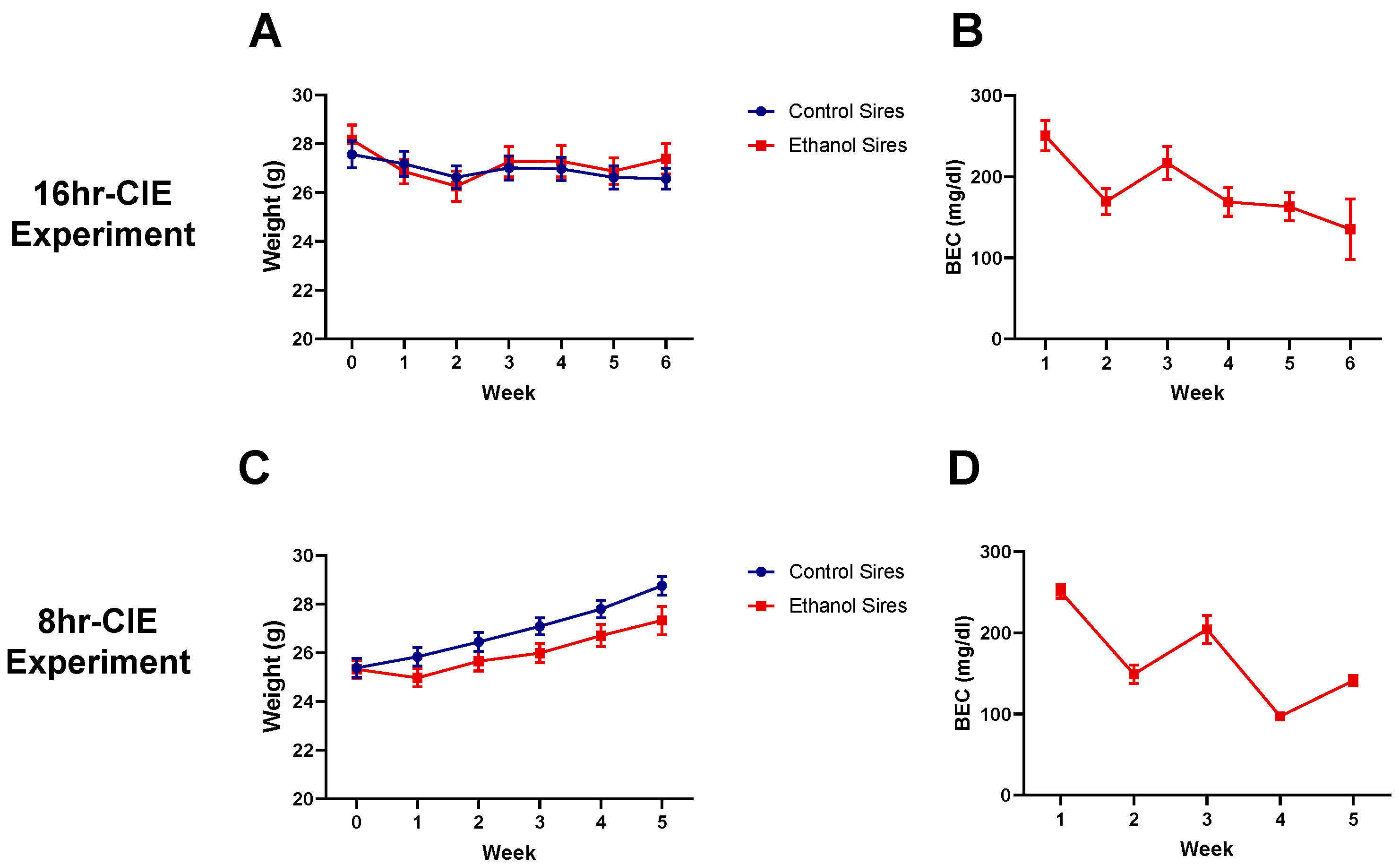
3.1. Paternal Preconception CIE Exposure
3.2. Breeding Results
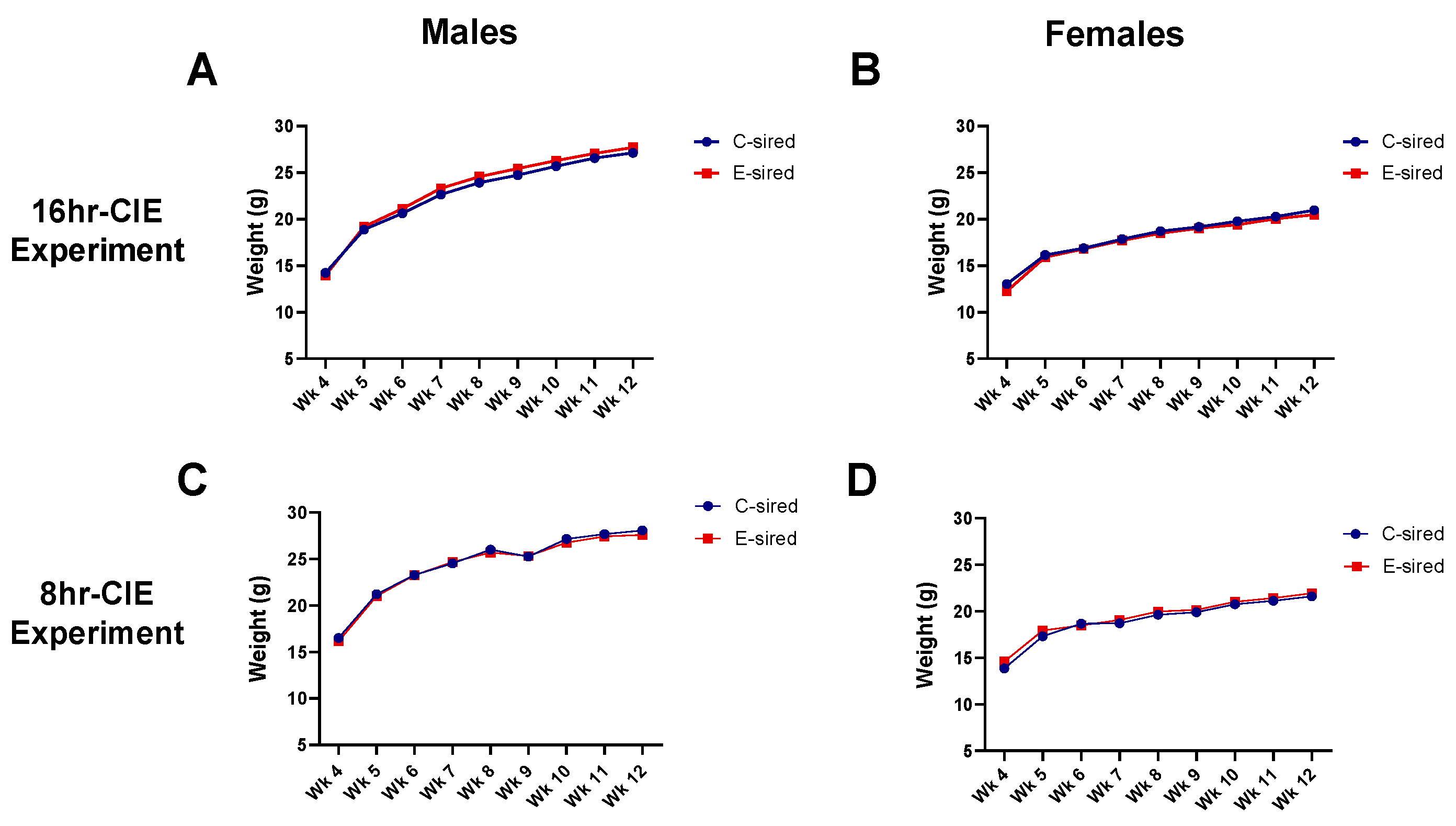
3.3. Body Weights of Offspring
3.4. Behavioral Assay Results of Adult Offspring
3.4.1. Object Location Memory Test (OLM)
3.4.2. Object Recognition Memory (ORM)
3.4.3. Social Interaction Test
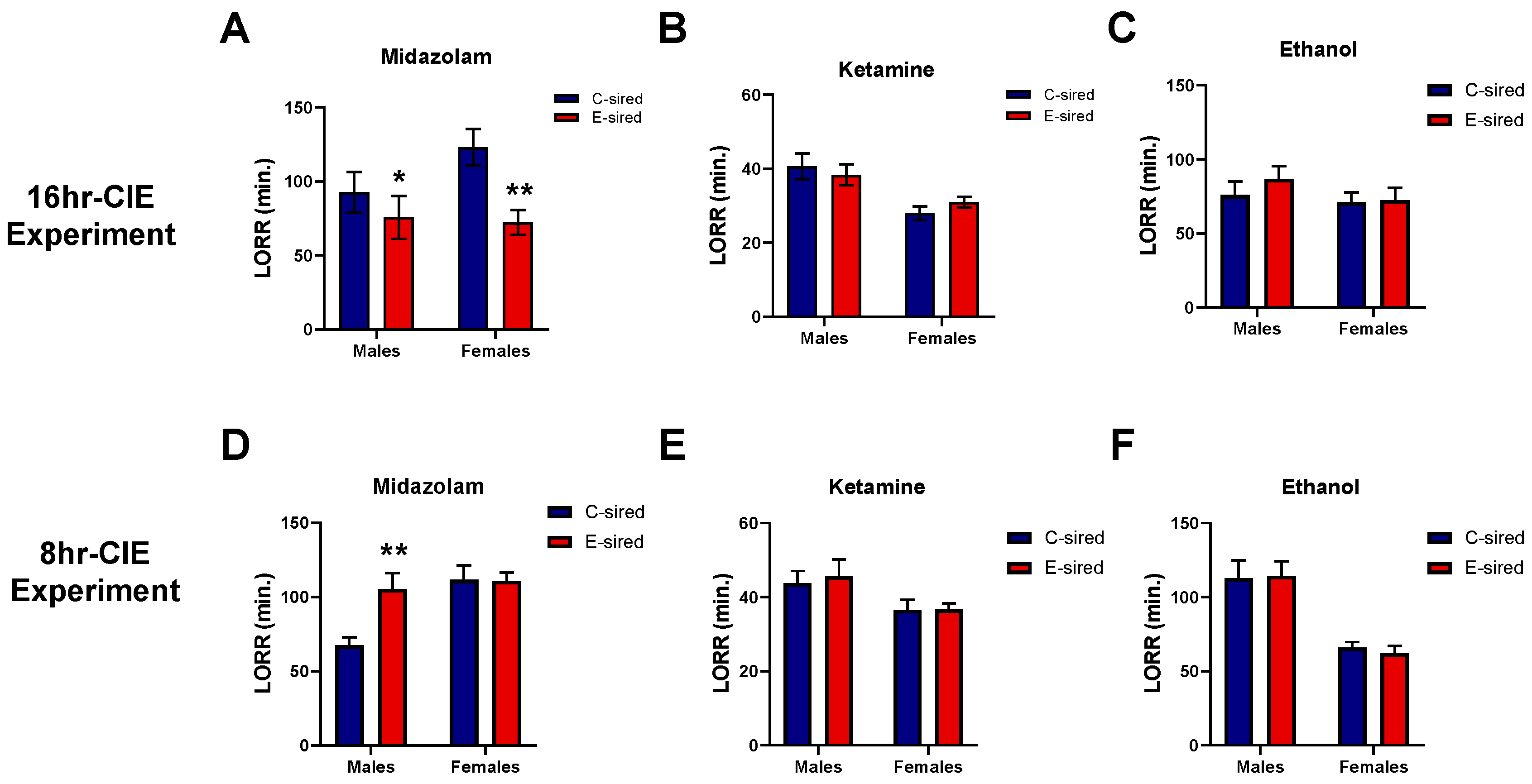
3.4.4. Loss of Righting Response (LORR)
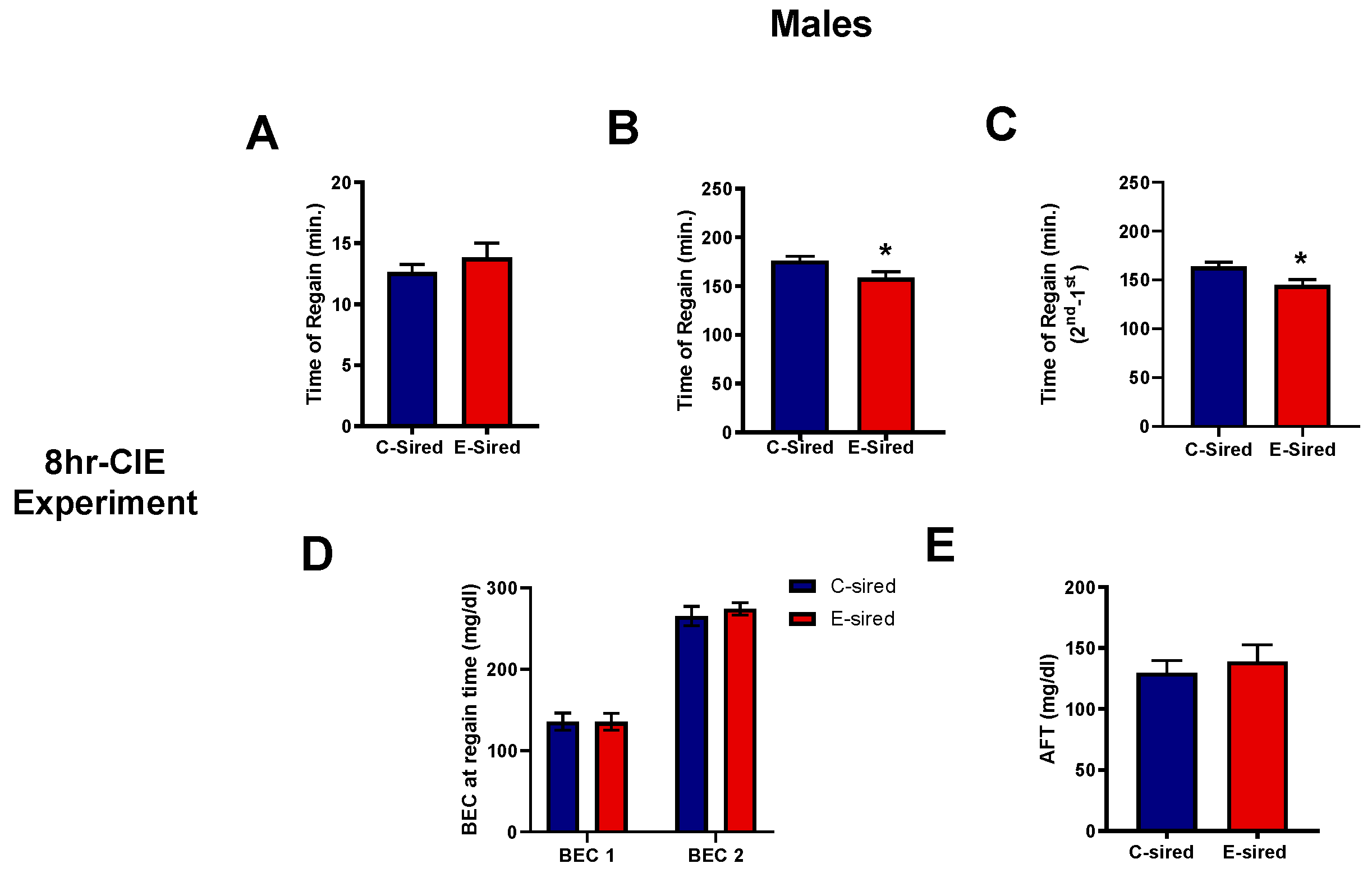
3.4.5. Acute Functional Tolerance Test (AFT)
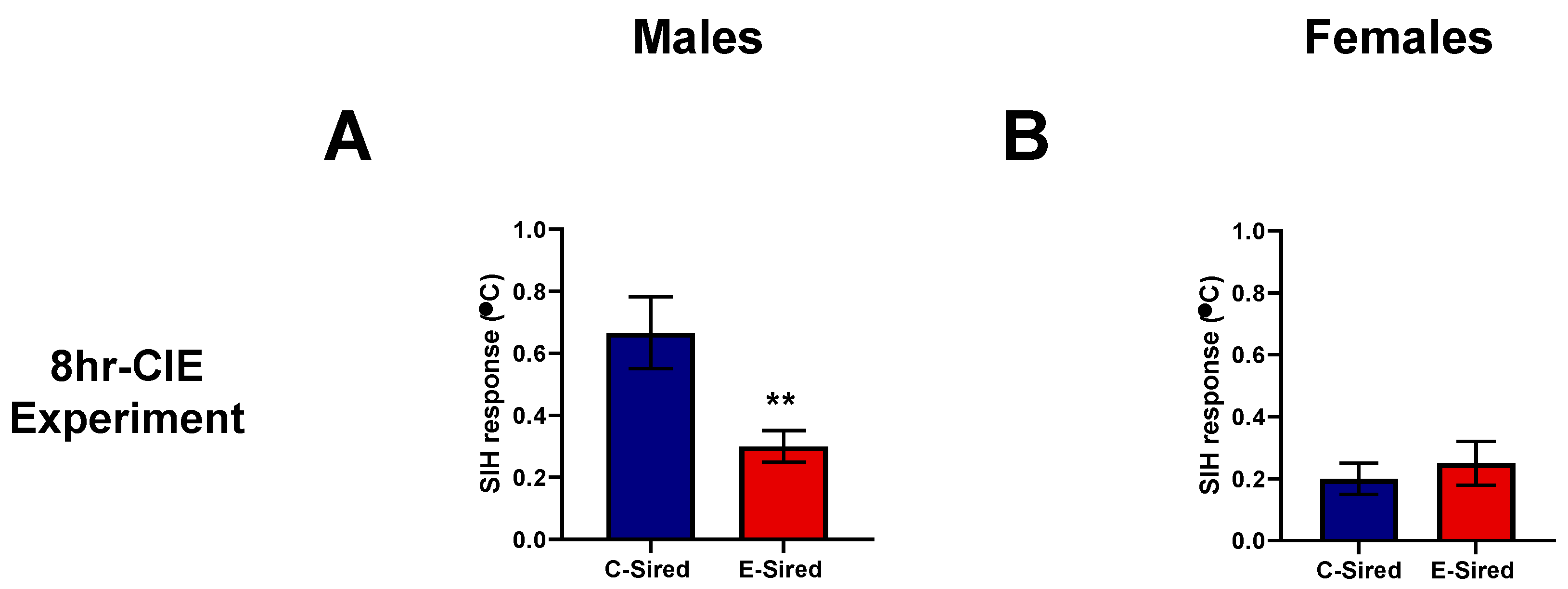
3.4.6. Stress-Induced Hyperthermia (SIH)
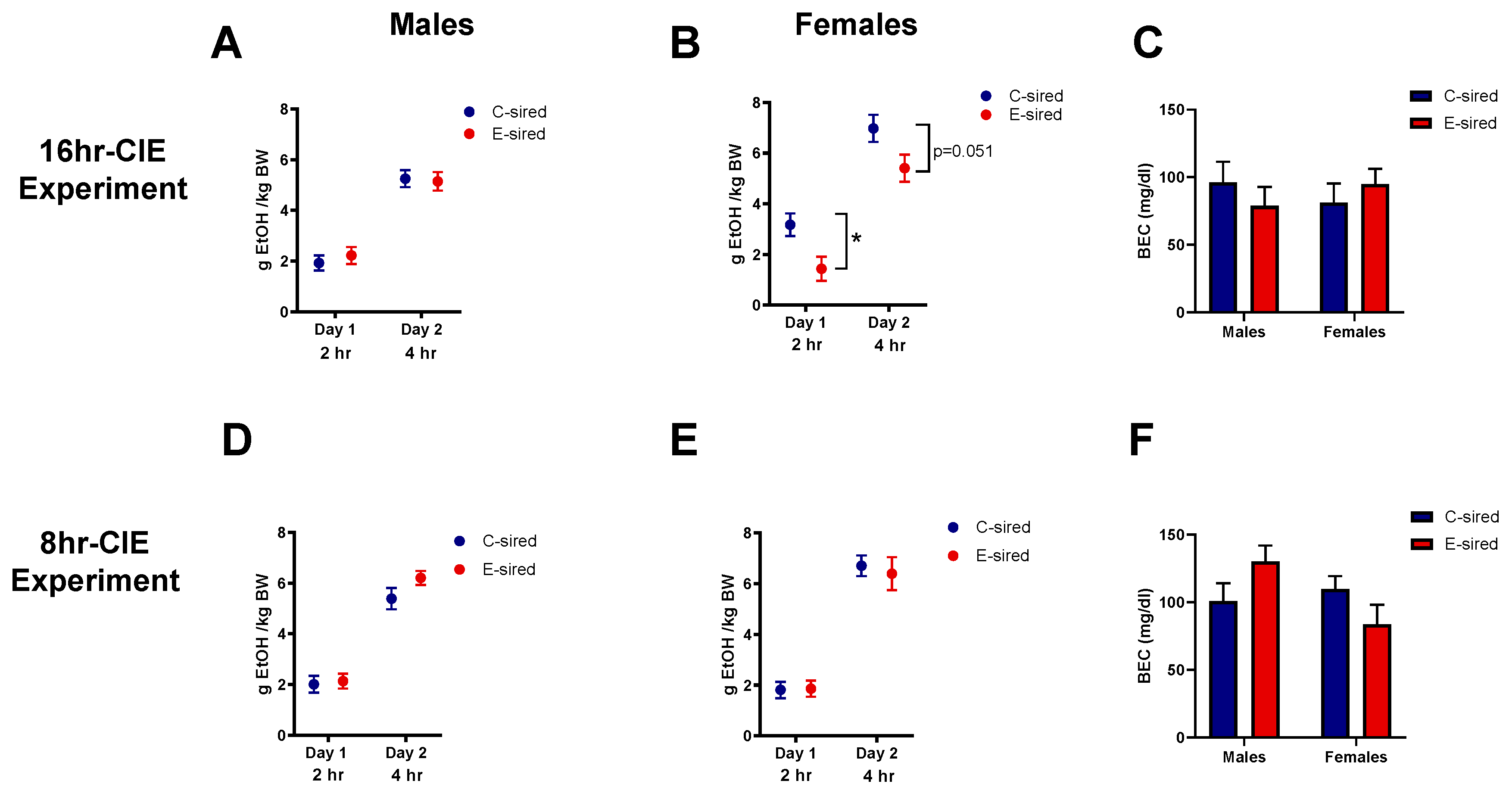
3.4.7. Drinking in the Dark (DID)
3.4.8. EOD 2BC Drinking Assay
3.4.9. Additional Behavioral Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mokdad, A.H.; Marks, J.S.; Stroup, D.F.; Gerberding, J.L. Actual causes of death in the United States, 2000. JAMA 2004, 291, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Substance Abuse and Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2018 National Survey on Drug Use and Health; HHS Publication No. PEP19-5068, NSDUH Series H-54; Center for Behavioral Health Statistics and Quality (CBHSQ): Rockville, MD, USA, 2019. [Google Scholar]
- Winslow, B.T.; Onysko, M.; Hebert, M. Medications for Alcohol Use Disorder. Am. Fam. Physician 2016, 93, 457–465. [Google Scholar] [PubMed]
- Kranzler, H.R.; Soyka, M. Diagnosis and Pharmacotherapy of Alcohol Use Disorder: A Review. JAMA 2018, 320, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Akbar, M.; Egli, M.; Cho, Y.E.; Song, B.J.; Noronha, A. Medications for alcohol use disorders: An overview. Pharmacol. Ther. 2018, 185, 64–85. [Google Scholar] [CrossRef]
- Day, J.; Savani, S.; Krempley, B.D.; Nguyen, M.; Kitlinska, J.B. Influence of paternal preconception exposures on their offspring: Through epigenetics to phenotype. Am. J. Stem Cells 2016, 5, 11–18. [Google Scholar]
- Marcho, C.; Oluwayiose, O.A.; Pilsner, J.R. The preconception environment and sperm epigenetics. Andrology 2020. [Google Scholar] [CrossRef]
- Martin-Calvo, N.; Minguez-Alarcon, L.; Gaskins, A.J.; Nassan, F.L.; Williams, P.L.; Souter, I.; Hauser, R.; Chavarro, J.E. Paternal preconception folate intake in relation to gestational age at delivery and birthweight of newborns conceived through assisted reproduction. Reprod. Biomed. Online 2019, 39, 835–843. [Google Scholar] [CrossRef]
- Yang, F.; Yuan, W.; Liang, H.; Song, X.; Yu, Y.; Gelaye, B.; Miao, M.; Li, J. Preconceptional paternal antiepileptic drugs use and risk of congenital anomalies in offspring: A nationwide cohort study. Eur. J. Epidemiol. 2019, 34, 651–660. [Google Scholar] [CrossRef]
- Wilsnack, R.W.; Wilsnack, S.C.; Kristjanson, A.F.; Vogeltanz-Holm, N.D.; Gmel, G. Gender and alcohol consumption: Patterns from the multinational GENACIS project. Addiction 2009, 104, 1487–1500. [Google Scholar] [CrossRef]
- Rompala, G.R.; Homanics, G.E. Intergenerational Effects of Alcohol: A Review of Paternal Preconception Ethanol Exposure Studies and Epigenetic Mechanisms in the Male Germline. Alcohol. Clin. Exp. Res. 2019, 43, 1032–1045. [Google Scholar] [CrossRef] [PubMed]
- Finegersh, A.; Rompala, G.R.; Martin, D.I.; Homanics, G.E. Drinking beyond a lifetime: New and emerging insights into paternal alcohol exposure on subsequent generations. Alcohol 2015, 49, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, R.C.; Wang, H.; Bedi, Y.; Golding, M.C. Preconception paternal alcohol exposure exerts sex-specific effects on offspring growth and long-term metabolic programming. Epigenet. Chromatin 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emanuele, N.V.; LaPaglia, N.; Steiner, J.; Colantoni, A.; Van Thiel, D.H.; Emanuele, M.A. Peripubertal paternal EtOH exposure. Endocrine 2001, 14, 213–219. [Google Scholar] [CrossRef]
- Ceccanti, M.; Coccurello, R.; Carito, V.; Ciafrè, S.; Ferraguti, G.; Giacovazzo, G.; Mancinelli, R.; Tirassa, P.; Chaldakov, G.N.; Pascale, E. Paternal alcohol exposure in mice alters brain NGF and BDNF and increases ethanol-elicited preference in male offspring. Addict. Biol. 2016, 21, 776–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finegersh, A.; Homanics, G.E. Paternal alcohol exposure reduces alcohol drinking and increases behavioral sensitivity to alcohol selectively in male offspring. PLoS ONE 2014, 9, e99078. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, D.F.; Cicero, T.J.; Kettinger, L., 3rd; Meyer, E.R. Paternal alcohol consumption in the rat impairs spatial learning performance in male offspring. Psychopharmacology 1991, 105, 289–302. [Google Scholar] [CrossRef]
- Liang, F.; Diao, L.; Liu, J.; Jiang, N.; Zhang, J.; Wang, H.; Zhou, W.; Huang, G.; Ma, D. Paternal ethanol exposure and behavioral abnormities in offspring: Associated alterations in imprinted gene methylation. Neuropharmacology 2014, 81, 126–133. [Google Scholar] [CrossRef]
- Kim, P.; Choi, C.S.; Park, J.H.; Joo, S.H.; Kim, S.Y.; Ko, H.M.; Kim, K.C.; Jeon, S.J.; Park, S.H.; Han, S.H. Chronic exposure to ethanol of male mice before mating produces attention deficit hyperactivity disorder-like phenotype along with epigenetic dysregulation of dopamine transporter expression in mouse offspring. J. Neurosci. Res. 2014, 92, 658–670. [Google Scholar] [CrossRef]
- Ledig, M.; Misslin, R.; Vogel, E.; Holownia, A.; Copin, J.C.; Tholey, G. Paternal alcohol exposure: Developmental and behavioral effects on the offspring of rats. Neuropharmacology 1998, 37, 57–66. [Google Scholar] [CrossRef]
- Rompala, G.R.; Finegersh, A.; Slater, M.; Homanics, G.E. Paternal preconception alcohol exposure imparts intergenerational alcohol-related behaviors to male offspring on a pure C57BL/6J background. Alcohol 2017, 60, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Beeler, E.; Nobile, Z.L.; Homanics, G.E. Paternal Preconception Every-Other-Day Ethanol Drinking Alters Behavior and Ethanol Consumption in Offspring. Brain Sci. 2019, 9, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollander, J.; McNivens, M.; Pautassi, R.M.; Nizhnikov, M.E. Offspring of male rats exposed to binge alcohol exhibit heightened ethanol intake at infancy and alterations in T-maze performance. Alcohol 2019, 76, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S.; DeCarli, L.M.; Sorrell, M.F. Experimental methods of ethanol administration. Hepatology 1989, 10, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, N.W.; Richardson, H.N.; Cole, M.; Koob, G.F. Vapor inhalation of alcohol in rats. Curr. Protoc. Neurosci. 2008. [Google Scholar] [CrossRef] [PubMed]
- Planeta, C.S. Animal models of alcohol and drug dependence. Braz. J. Psychiatry 2013, 35 (Suppl. 2), S140–S146. [Google Scholar] [CrossRef] [Green Version]
- Eisenhardt, M.; Hansson, A.C.; Spanagel, R.; Bilbao, A. Chronic intermittent ethanol exposure in mice leads to an up-regulation of CRH/CRHR1 signaling. Alcohol. Clin. Exp. Res. 2015, 39, 752–762. [Google Scholar] [CrossRef]
- Rompala, G.R.; Finegersh, A.; Homanics, G.E. Paternal preconception ethanol exposure blunts hypothalamic-pituitary-adrenal axis responsivity and stress-induced excessive fluid intake in male mice. Alcohol 2016, 53, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Vogel-Ciernia, A.; Wood, M.A. Examining object location and object recognition memory in mice. Curr. Protoc. Neurosci. 2014, 69, 8–31. [Google Scholar] [CrossRef] [Green Version]
- Park, S.M.; Plachez, C.; Huang, S. Sex-Dependent Motor Deficit and Increased Anxiety-Like States in Mice Lacking Autism-Associated Gene Slit3. Front. Behav. Neurosci. 2018, 12, 261. [Google Scholar] [CrossRef] [Green Version]
- Blednov, Y.A.; Borghese, C.M.; Ruiz, C.I.; Cullins, M.A.; Da Costa, A.; Osterndorff-Kahanek, E.A.; Homanics, G.E.; Harris, R.A. Mutation of the inhibitory ethanol site in GABAA rho1 receptors promotes tolerance to ethanol-induced motor incoordination. Neuropharmacology 2017, 123, 201–209. [Google Scholar] [CrossRef]
- Groenink, L.; Vinkers, C.; van Oorschot, R.; Olivier, B. Models of anxiety: Stress-induced hyperthermia (SIH) in singly housed mice. Curr. Protoc. Pharmacol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.J.; Flanagan, J.P.; Marchant, N.J.; Lawrence, A.J. Reduced alcohol-seeking in male offspring of sires exposed to alcohol self-administration followed by punishment-imposed abstinence. Pharmacol. Res. Perspect. 2018, 6, e00384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Alavez, M.; Alboni, S.; Conti, B. Sex- and age-specific differences in core body temperature of C57Bl/6 mice. Age 2011, 33, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rompala, G.R.; Mounier, A.; Wolfe, C.M.; Lin, Q.; Lefterov, I.; Homanics, G.E. Heavy Chronic Intermittent Ethanol Exposure Alters Small Noncoding RNAs in Mouse Sperm and Epididymosomes. Front. Genet. 2018, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, H.C.; Hale, R.L. Repeated episodes of ethanol withdrawal potentiate the severity of subsequent withdrawal seizures: An animal model of alcohol withdrawal “kindling”. Alcohol. Clin. Exp. Res. 1993, 17, 94–98. [Google Scholar] [CrossRef]
- Becker, H.C.; Lopez, M.F. Increased ethanol drinking after repeated chronic ethanol exposure and withdrawal experience in C57BL/6 mice. Alcohol. Clin. Exp. Res. 2004, 28, 1829–1838. [Google Scholar] [CrossRef]
- Becker, H.C. Alcohol dependence, withdrawal, and relapse. Alcohol Res. Health 2008, 31, 348–361. [Google Scholar]
- O’Dell, L.E.; Roberts, A.J.; Smith, R.T.; Koob, G.F. Enhanced alcohol self-administration after intermittent versus continuous alcohol vapor exposure. Alcohol. Clin. Exp. Res. 2004, 28, 1676–1682. [Google Scholar] [CrossRef]
- Goldstein, D.B.; Pal, N. Alcohol dependence produced in mice by inhalation of ethanol: Grading the withdrawal reaction. Science 1971, 172, 288–290. [Google Scholar] [CrossRef]
- Guthauser, B.; Boitrelle, F.; Plat, A.; Thiercelin, N.; Vialard, F. Chronic Excessive Alcohol Consumption and Male Fertility: A Case Report on Reversible Azoospermia and a Literature Review. Alcohol Alcohol. 2013, 49, 42–44. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.K.; Gottschau, M.; Madsen, J.O.B.; Andersson, A.-M.; Lassen, T.H.; Skakkebæk, N.E.; Swan, S.H.; Priskorn, L.; Juul, A.; Jørgensen, N. Habitual alcohol consumption associated with reduced semen quality and changes in reproductive hormones; a cross-sectional study among 1221 young Danish men. BMJ Open 2014, 4, e005462. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, R.A.; Calogero, A.E.; Vicari, E.; La Vignera, S. Chronic consumption of alcohol and sperm parameters: Our experience and the main evidences. Andrologia 2015, 47, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Muthusami, K.R.; Chinnaswamy, P. Effect of chronic alcoholism on male fertility hormones and semen quality. Fertil. Steril. 2005, 84, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Nizhnikov, M.E.; Popoola, D.O.; Cameron, N.M. Transgenerational Transmission of the Effect of Gestational Ethanol Exposure on Ethanol Use-Related Behavior. Alcohol. Clin. Exp. Res. 2016, 40, 497–506. [Google Scholar] [CrossRef]
- Olsen, R.W.; Liang, J. Role of GABAA receptors in alcohol use disorders suggested by chronic intermittent ethanol (CIE) rodent model. Mol. Brain 2017, 10, 45. [Google Scholar] [CrossRef]
- Kumar, S.; Ren, Q.; Beckley, J.H.; O’Buckley, T.K.; Gigante, E.D.; Santerre, J.L.; Werner, D.F.; Morrow, A.L. Ethanol Activation of Protein Kinase A Regulates GABA(A) Receptor Subunit Expression in the Cerebral Cortex and Contributes to Ethanol-Induced Hypnosis. Front. Neurosci. 2012, 6, 44. [Google Scholar] [CrossRef] [Green Version]
- Aguayo, L.G.; Peoples, R.W.; Yeh, H.H.; Yevenes, G.E. GABA(A) receptors as molecular sites of ethanol action. Direct or indirect actions? Curr. Top. Med. Chem. 2002, 2, 869–885. [Google Scholar] [CrossRef]
- Tarter, R.E.; Hegedus, A.M.; Goldstein, G.; Shelly, C.; Alterman, A.I. Adolescent sons of alcoholics: Neuropsychological and personality characteristics. Alcohol. Clin. Exp. Res. 1984, 8, 216–222. [Google Scholar] [CrossRef]
- Hegedus, A.M.; Alterman, A.I.; Tarter, R.E. Learning achievement in sons of alcoholics. Alcohol. Clin. Exp. Res. 1984, 8, 330–333. [Google Scholar] [CrossRef]
- Wimmer, M.; Briand, L.; Fant, B.; Guercio, L.; Arreola, A.; Schmidt, H.; Sidoli, S.; Han, Y.; Garcia, B.; Pierce, R. Paternal cocaine taking elicits epigenetic remodeling and memory deficits in male progeny. Mol. Psychiatry 2017, 22, 1641. [Google Scholar] [CrossRef] [Green Version]
- Diaz, M.R.; Mooney, S.M.; Varlinskaya, E.I. Acute prenatal exposure to ethanol on gestational day 12 elicits opposing deficits in social behaviors and anxiety-like behaviors in Sprague Dawley rats. Behav. Brain Res. 2016, 310, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooney, S.M.; Varlinskaya, E.I. Enhanced sensitivity to socially facilitating and anxiolytic effects of ethanol in adolescent Sprague Dawley rats following acute prenatal ethanol exposure. Alcohol 2018, 69, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Holman, P.J.; Ellis, L.; Morgan, E.; Weinberg, J. Prenatal alcohol exposure disrupts male adolescent social behavior and oxytocin receptor binding in rodents. Horm. Behav. 2018, 105, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Conner, K.E.; Bottom, R.T.; Huffman, K.J. The Impact of Paternal Alcohol Consumption on Offspring Brain and Behavioral Development. Alcohol. Clin. Exp. Res. 2020, 44, 125–140. [Google Scholar] [CrossRef]
- Meek, L.R.; Myren, K.; Sturm, J.; Burau, D. Acute paternal alcohol use affects offspring development and adult behavior. Physiol. Behav. 2007, 91, 154–160. [Google Scholar] [CrossRef]
- Zou, H.; Xie, Q.; Zhang, M.; Zhang, C.; Zhao, G.; Jin, M.; Yu, L. Chronic alcohol consumption from adolescence-to-adulthood in mice--effect on growth and social behavior. Drug Alcohol Depend. 2009, 104, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Melendez, R.I. Intermittent (every-other-day) drinking induces rapid escalation of ethanol intake and preference in adolescent and adult C57BL/6J mice. Alcohol. Clin. Exp. Res. 2011, 35, 652–658. [Google Scholar] [CrossRef]
- Rodgers, A.B.; Morgan, C.P.; Leu, N.A.; Bale, T.L. Transgenerational epigenetic programming via sperm microRNA recapitulates effects of paternal stress. Proc. Natl. Acad. Sci. USA 2015, 112, 13699–13704. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.-H.; Peng, H.; Zhang, X.; Zhang, Y. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Benito, E.; Kerimoglu, C.; Ramachandran, B.; Pena-Centeno, T.; Jain, G.; Stilling, R.M.; Islam, M.R.; Capece, V.; Zhou, Q.; Edbauer, D.; et al. RNA-Dependent Intergenerational Inheritance of Enhanced Synaptic Plasticity after Environmental Enrichment. Cell Rep. 2018, 23, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, A.B.; Morgan, C.P.; Bronson, S.L.; Revello, S.; Bale, T.L. Paternal stress exposure alters sperm microRNA content and reprograms offspring HPA stress axis regulation. J. Neurosci. 2013, 33, 9003–9012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapp, K.; Jawaid, A.; Sarkies, P.; Bohacek, J.; Pelczar, P.; Prados, J.; Farinelli, L.; Miska, E.; Mansuy, I.M. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 2014, 17, 667–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapp, K.; van Steenwyk, G.; Germain, P.-L.; Matsushima, W.; Rudolph, K.L.; Manuella, F.; Roszkowski, M.; Vernaz, G.; Ghosh, T.; Pelczar, P. Alterations in sperm long RNA contribute to the epigenetic inheritance of the effects of postnatal trauma. Mol. Psychiatry 2018, 25, 2162–2174. [Google Scholar] [CrossRef] [Green Version]
- Bedi, Y.; Chang, R.C.; Gibbs, R.; Clement, T.M.; Golding, M.C. Alterations in sperm-inherited noncoding RNAs associate with late-term fetal growth restriction induced by preconception paternal alcohol use. Reprod. Toxicol. 2019, 87, 11–20. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 16hr-CIE | 8hr-CIE |
|---|---|
| Housing Room: Standard 7am ON-7pm OFF light cycle | |
| Cohort 1 | Cohort 1 |
| Marble Burying Test Object Location Test Social Interaction Test Object Recognition Test Blood Glucose Levels LORR (Ethanol) | Object Location Test Social Interaction Test Acute Functional Tolerance Test LORR (Ketamine, Midazolam, and Ethanol) |
| Housing Room: Reversed 10am OFF-10pm ON light cycle | |
| Cohort 2 | Cohort 2: Single-housed |
| Home Cage Activity Prepulse Inhibition Test Novelty Suppressed Feeding Test LORR (Ketamine and Midazolam) Drinking in the Dark | Stress-Induced Hyperthermia Bottle-Brush Test Drinking in the Dark EOD/2 bottle choice |
| 16hr CIE | 8hr CIE ** | |||
|---|---|---|---|---|
| Parameter | C-Sired | E-Sired | C-Sired | E-Sired |
| Number of males exposed | 16 | 16 | 15 | 14 |
| Number of males siring litters | 12 (75%) | 8 (50%) | 14 (93%) | 12 (86%) |
| Number of sires with viable offspring | 11 (69%) | 4 (25%) * | 12 (80%) | 8 (57%) |
| Average litter size of viable litters | 6.7 | 7.2 | 6.5 | 6.8 |
| 16hr-CIE | 8hr-CIE | |||
|---|---|---|---|---|
| F1 Males | F1 Females | F1 Males | F1 Females | |
| Object location memory | No change | No change | No change | No change |
| Object recognition memory | No change | No change | No change | No change |
| Social Interaction | No change | No change | No change | No change |
| Acute functional tolerance | Not performed | Not performed | Shorter recovery time after 2nd EtOH injection | No change |
| Loss of righting response (LORR) (midazolam) | Shorter LORR duration | Shorter LORR duration | Longer LORR duration | No change |
| LORR (ketamine and ethanol) | No change | No change | No change | No change |
| Stress-induced hyperthermia | Not performed | Not performed | Reduced SIH response | No change |
| Drinking in the dark | No change | Reduced consumption | No change | No change |
| Every-other-day two-bottle choice | No change | No change | No change | No change |
| Marble burying test | No change | No change | No change | No change |
| Novelty suppressed feeding test | No change | No change | No change | No change |
| Home cage activity | No change | No change | No change | No change |
| Prepulse inhibition of acoustic startle | No change | No change | No change | No change |
| Bottle-brush test | No change | No change | No change | No change |
| Blood glucose levels | No change | No change | No change | No change |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rathod, R.S.; Ferguson, C.; Seth, A.; Baratta, A.M.; Plasil, S.L.; Homanics, G.E. Effects of Paternal Preconception Vapor Alcohol Exposure Paradigms on Behavioral Responses in Offspring. Brain Sci. 2020, 10, 658. https://doi.org/10.3390/brainsci10090658
Rathod RS, Ferguson C, Seth A, Baratta AM, Plasil SL, Homanics GE. Effects of Paternal Preconception Vapor Alcohol Exposure Paradigms on Behavioral Responses in Offspring. Brain Sciences. 2020; 10(9):658. https://doi.org/10.3390/brainsci10090658
Chicago/Turabian StyleRathod, Richa S., Carolyn Ferguson, Amit Seth, Annalisa M. Baratta, Sonja L. Plasil, and Gregg E. Homanics. 2020. "Effects of Paternal Preconception Vapor Alcohol Exposure Paradigms on Behavioral Responses in Offspring" Brain Sciences 10, no. 9: 658. https://doi.org/10.3390/brainsci10090658