Glutamatergic Activation of Neuronostatin Neurons in the Periventricular Nucleus of the Hypothalamus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Groups
2.3. Immunohistochemistry
2.4. Statistical Analysis
3. Results
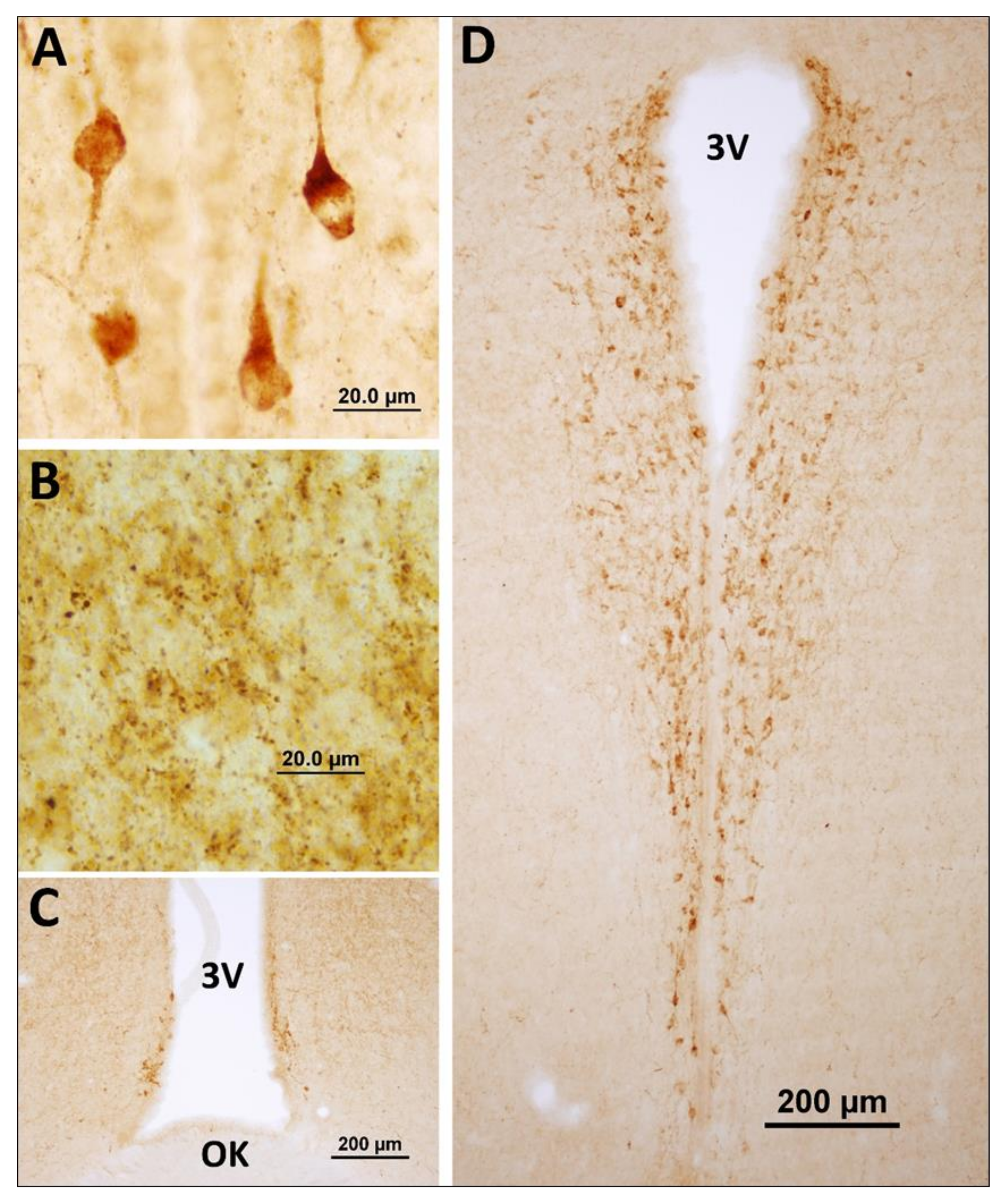
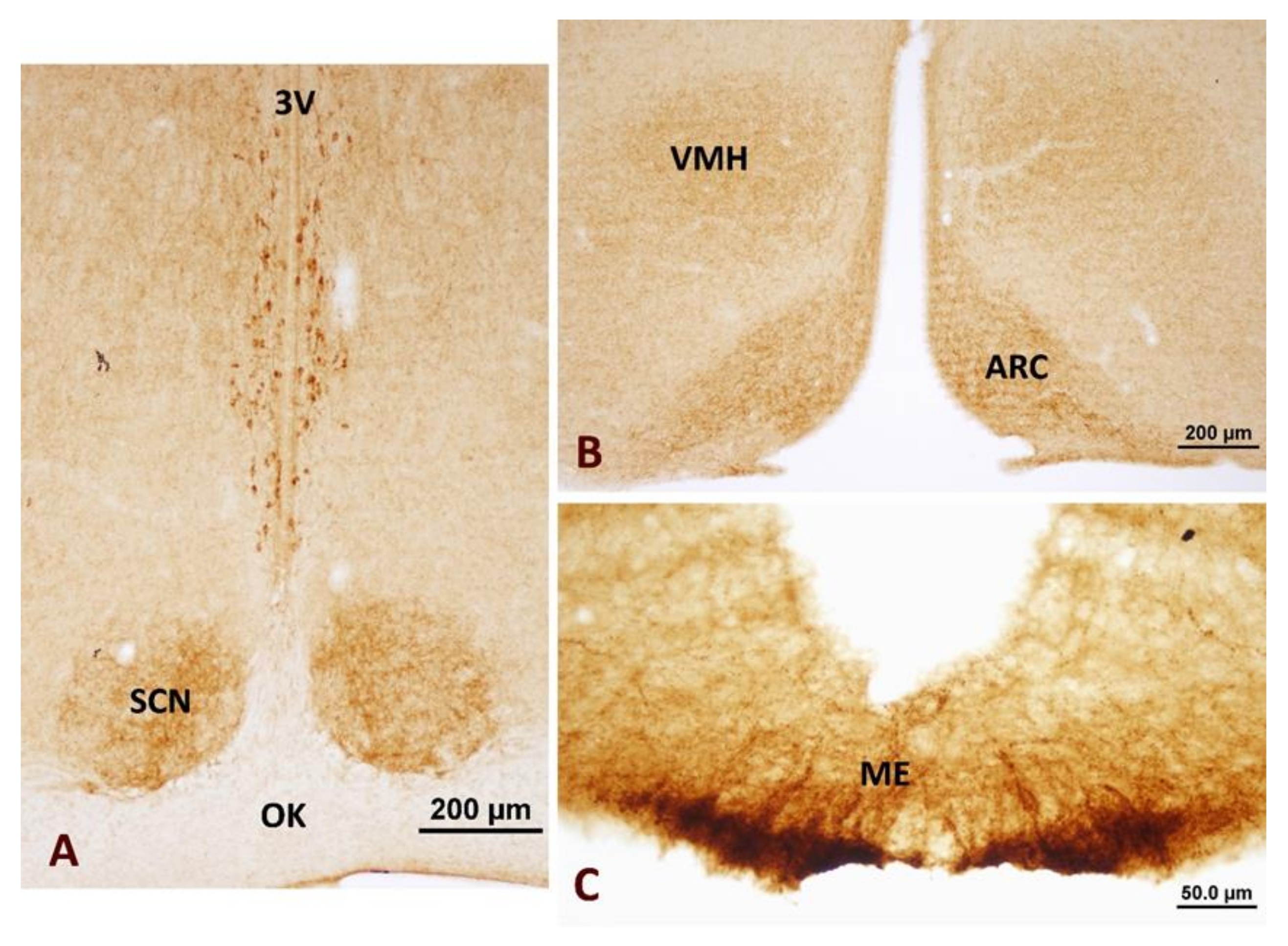
3.1. Neuronostatin Neuron Immunoreactivity in the Hypothalamus
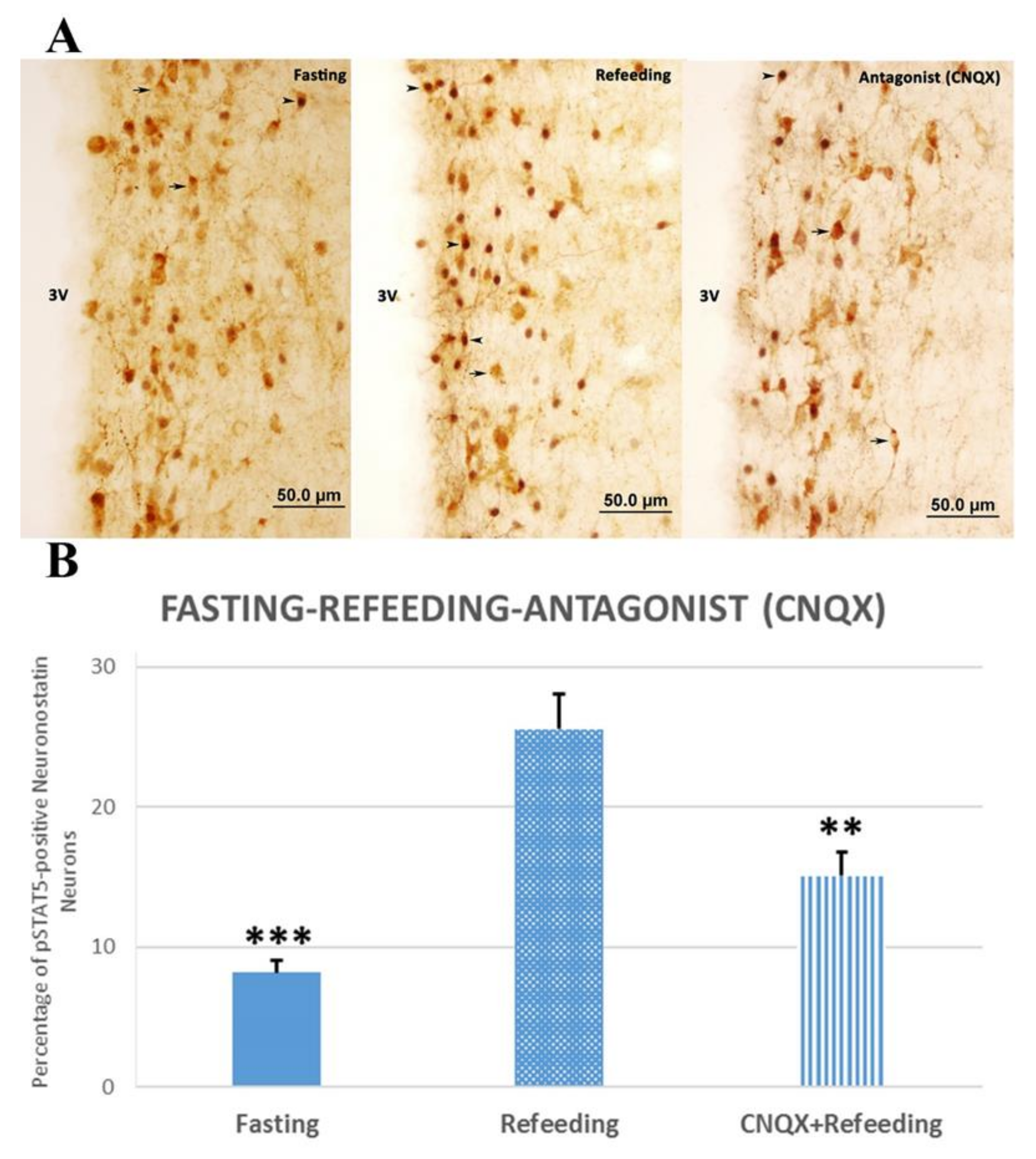
3.2. Physiological Stimulation of Neuronostatin Neurons and Investigation of the Effects of Glutamate Antagonists
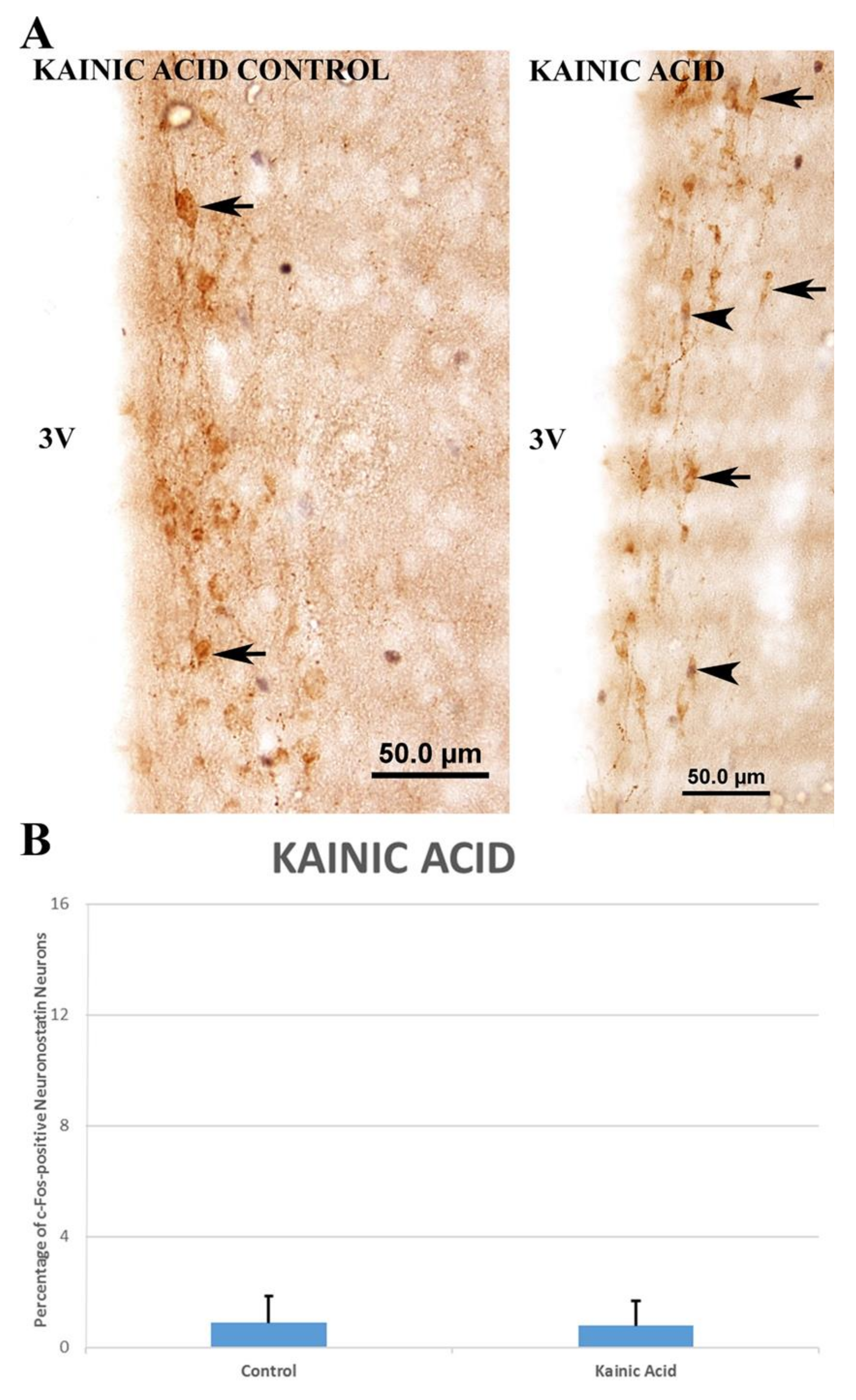
3.3. Investigation of the Glutamatergic System Effects and Activation Pathways in the Regulation of Neuronostatin Neurons
4. Discussion
4.1. Hypothalamic Distribution of Neuronostatin Neurons
4.2. Investigation of the Glutamatergic System Effects and Activation Pathways in the Regulation of Neuronostatin Neurons
4.3. Physiological Stimulation of Neuronostatin Neurons and Investigation of the Effects of Glutamate Antagonists
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Samson, W.K.; Zhang, J.V.; Avsian-Kretchmer, O.; Cui, K.; Yosten, G.L.C.; Klein, C.; Lyu, R.M.; Yong, X.W.; Xiang, Q.C.; Yang, J.; et al. Neuronostatin encoded by the somatostatin gene regulates neuronal, cardiovascular, and metabolic functions. J. Biol. Chem. 2008, 283, 31949–31959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dun, S.L.; Brailoiu, G.C.; Tica, A.A.; Yang, J.; Chang, J.K.; Brailoiu, E.; Dun, N.J. Neuronostatin is co-expressed with somatostatin and mobilizes calcium in cultured rat hypothalamic neurons. Neuroscience 2010, 166, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, A.; Baldassano, S.; Caldara, G.; Mulè, F. Neuronostatin: Peripheral site of action in mouse stomach. Peptides 2015, 64, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Su, S.F.; Yang, A.M.; Yang, S.B.; Wang, N.B.; Lu, S.S.; Wang, H.H.; Chen, Q. Intracerebroventricular administration of neuronostatin delays gastric emptying and gastrointestinal transit in mice. Peptides 2012, 35, 31–35. [Google Scholar] [CrossRef]
- Yosten, G.L.C.; Samson, W.K. The melanocortins, not oxytocin, mediate the anorexigenic and antidipsogenic effects of neuronostatin. Peptides 2010, 31, 1711–1714. [Google Scholar] [CrossRef] [Green Version]
- Brann, D.W.; Mahesh, V.B. Excitatory amino acids: Function and significance in reproduction and neuroendocrine regulation. Front. Neuroendocrinol. 1994, 15, 3–49. [Google Scholar] [CrossRef]
- Brann, D.W. Glutamate: A major excitatory transmitter in neuroendocrine regulation. Neuroendocrinology 1995, 61, 213–225. [Google Scholar] [CrossRef]
- Tse, Y.C.; Yung, K.K.L. Cellular expression of ionotropic glutamate receptor subunits in subpopulations of neurons in the rat substantia nigra pars reticulata. Brain Res. 2000, 854, 57–69. [Google Scholar] [CrossRef]
- Takamori, S. VGLUTs: “Exciting” times for glutamatergic research? Neurosci. Res. 2006, 55, 343–351. [Google Scholar] [CrossRef]
- Kaneko, T.; Fujiyama, F.; Hioki, H. Immunohistochemical localization of candidates for vesicular glutamate transporters in the rat brain. J. Comp. Neurol. 2002, 444, 39–62. [Google Scholar] [CrossRef]
- Hisano, S.; Nogami, H. Transporters in the neurohypophysial neuroendocrine system, with special reference to vesicular glutamate transporters (BNPI and DNPI): A review. Microsc. Res. Tech. 2002, 56, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Chung, K.F.; Adcock, I.M. Update on glucocorticoid action and resistance. J. Allergy Clin. Immunol. 2006, 117, 522–543. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D. Introduction to NF-κB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saklatvala, J. The p38 MAP kinase pathway as a therapeutic target in inflammatory disease. Curr. Opin. Pharmacol. 2004, 4, 372–377. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Lyo, D. Anatomical markers of activity in neuroendocrine systems: Are we all “fos-ed out”? J. Neuroendocrinol. 2002, 14, 259–268. [Google Scholar] [CrossRef]
- Zhao, J.B.; Zhang, Y.; Li, G.Z.; Su, X.F.; Hang, C.H. Activation of JAK2/STAT pathway in cerebral cortex after experimental traumatic brain injury of rats. Neurosci. Lett. 2011, 498, 147–152. [Google Scholar] [CrossRef]
- Balazs, R. Trophic Effect of Glutamate. Curr. Top. Med. Chem. 2006, 6, 961–968. [Google Scholar] [CrossRef]
- Altarejos, J.Y.; Montminy, M. CREB and the CRTC co-activators: Sensors for hormonal and metabolic signals. Nat. Rev. Mol. Cell Biol. 2011, 12, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: London, UK, 2009; pp. Plate 35–Plate 63. [Google Scholar]
- Eyigor, O.; Centers, A.; Jennes, L. Distribution of ionotropic glutamate receptor subunit mRNAs in the rat hypothalamus. J. Comp. Neurol. 2001, 434, 101–124. [Google Scholar] [CrossRef]
- Eyigor, O.; Minbay, Z.; Cavusoglu, I.; Jennes, L. Localization of kainate receptor subunit GluR5-immunoreactive cells in the rat hypothalamus. Mol. Brain Res. 2005, 136, 38–44. [Google Scholar] [CrossRef]
- Eyigor, O.; Minbay, Z.; Cavusoglu, I. Activation of orexin neurons through non-NMDA glutamate receptors evidenced by c-Fos immunohistochemistry. Endocrine 2010, 37, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Meeker, R.B.; McGinnis, S.; Greenwood, R.S.; Hayward, J.N. Increased hypothalamic glutamate receptors induced by water deprivation. Neuroendocrinology 1994, 60, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Van Del Pol, A.N.; Wuarin, J.P.; Dudek, F.E. Glutamate neurotransmission in the neuroendocrine hypothalamus. In Excitatory Amino AcidsTheir Role in Neuroendocrine Function; Brann, D.W., Mahesh, V.B., Eds.; CRC Press: Boca Rotan, FL, USA, 1996; pp. 1–54. [Google Scholar]
- Minbay, F.Z.; Eyigor, O.; Çavusoglu, I. Kainic acid activates oxytocinergic neurons through non-NMDA glutamate receptors. Int. J. Neurosci. 2006, 116, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Piekut, D.T.; Pretel, S.; Applegate, C.D. Activation of oxytocin-containing neurons of the paraventricular nucleus (PVN) following generalized seizures. Synapse 1996, 23, 312–320. [Google Scholar] [CrossRef]
- Gynther, M.; Petsalo, A.; Hansen, S.H.; Bunch, L.; Pickering, D.S. Blood–Brain Barrier permeability and brain uptake mechanism of kainic acid and dihydrokainic acid. Neurochem. Res. 2015, 40, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, C.; Tercero, I.; Pineda, A.; Burgos, J.S. Simvastatin is the statin that most efficiently protects against kainate-induced excitotoxicity and memory impairment. J. Alzheimers Dis. 2011, 24, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Hankir, M.K.; Parkinson, J.R.; Bloom, S.R.; Bell, J.D. The effects of glutamate receptor agonists and antagonists on mouse hypothalamic and hippocampal neuronal activity shown through manganese enhanced MRI. NeuroImage 2012, 59, 968–978. [Google Scholar] [CrossRef]
- Gok-Yurtseven, D.; Kafa, I.M.; Minbay, Z.; Eyigor, O. Glutamatergic activation of A1 and A2 noradrenergic neurons in the rat brain stem. Croat. Med. J. 2019, 60, 352–360. [Google Scholar] [CrossRef]
- Knapp, D.J.; Braun, C.J.; Duncan, G.E.; Qian, Y.; Fernandes, A.; Crews, F.T.; Breese, G.R. Regional specificity of ethanol and NMDA action in brain revealed with Fos-like immunohistochemistry and differential routes of drug administration. Alcohol. Clin. Exp. Res. 2001, 25, 1662–1672. [Google Scholar] [CrossRef]
- Shin, S.W.; Park, J.W.; Suh, M.H.; Suh, S.I.; Choe, B.K. Persistent expression of Fas/FasL mRNA in the mouse hippocampus after a single NMDA injection. J. Neurochem. 1998, 71, 1773–1776. [Google Scholar] [CrossRef]
- Mitsikostas, D.D.; Sanchez del Rio, M.; Waeber, C.; Huang, Z.; Cutrer, F.M.; Moskowitz, M.A. Non-NMDA glutamate receptors modulate capsaicin induced c-fos expression within trigeminal nucleus caudalis. Br. J. Pharmacol. 1999, 127, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yosten, G.L.C.; Pate, A.T.; Samson, W.K. Neuronostatin acts in brain to biphasically increase mean arterial pressure through sympatho-activation followed by vasopressin secretion: The role of melanocortin receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1194–R1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yosten, G.L.C.; Samson, W.K. Separating thirst from hunger. In Neurobiology of Body Fluid Homeostasis: Transduction and Integration; De Luca, L.A., Jr., Menani, J.V., Johnson, A.K., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; pp. 103–110. [Google Scholar]
- Zorrilla, E.P.; Inoue, K.; Fekete, É.M.; Tabarin, A.; Valdez, G.R.; Koob, G.F. Measuring meals: Structure of prandial food and water intake of rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1450–R1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakao, H.; Gouilleux, F.; Groner, B. Mammary gland factor (MGF) is a novel member of the cytokine regulated transcription factor gene family and confers the prolactin response. EMBO J. 1994, 13, 2182–2191. [Google Scholar] [CrossRef]
- Mütze, J.; Roth, J.; Gerstberger, R.; Hübschle, T. Nuclear translocation of the transcription factor STAT5 in the rat brain after systemic leptin administration. Neurosci. Lett. 2007, 417, 286–291. [Google Scholar] [CrossRef]
- Luo, S.X.; Huang, J.; Li, Q.; Mohammad, H.; Lee, C.-Y.; Krishna, K.; Maun-Yeng Kok, A.; Tan, Y.L.; Lim, J.Y.; Li, H.; et al. Regulation of feeding by somatostatin neurons in the tuberal nucleus. Science 2018, 361, 76–81. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serter Kocoglu, S.; Gok Yurtseven, D.; Cakir, C.; Minbay, Z.; Eyigor, O. Glutamatergic Activation of Neuronostatin Neurons in the Periventricular Nucleus of the Hypothalamus. Brain Sci. 2020, 10, 217. https://doi.org/10.3390/brainsci10040217
Serter Kocoglu S, Gok Yurtseven D, Cakir C, Minbay Z, Eyigor O. Glutamatergic Activation of Neuronostatin Neurons in the Periventricular Nucleus of the Hypothalamus. Brain Sciences. 2020; 10(4):217. https://doi.org/10.3390/brainsci10040217
Chicago/Turabian StyleSerter Kocoglu, Sema, Duygu Gok Yurtseven, Cihan Cakir, Zehra Minbay, and Ozhan Eyigor. 2020. "Glutamatergic Activation of Neuronostatin Neurons in the Periventricular Nucleus of the Hypothalamus" Brain Sciences 10, no. 4: 217. https://doi.org/10.3390/brainsci10040217