Which Effects on Neuroanatomy and Path-Integration Survive? Results of a Randomized Controlled Study on Intensive Balance Training

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Ethical Approval
2.2. Subjects
2.3. Study Design
2.4. Intervention
2.5. Behavioral Tests
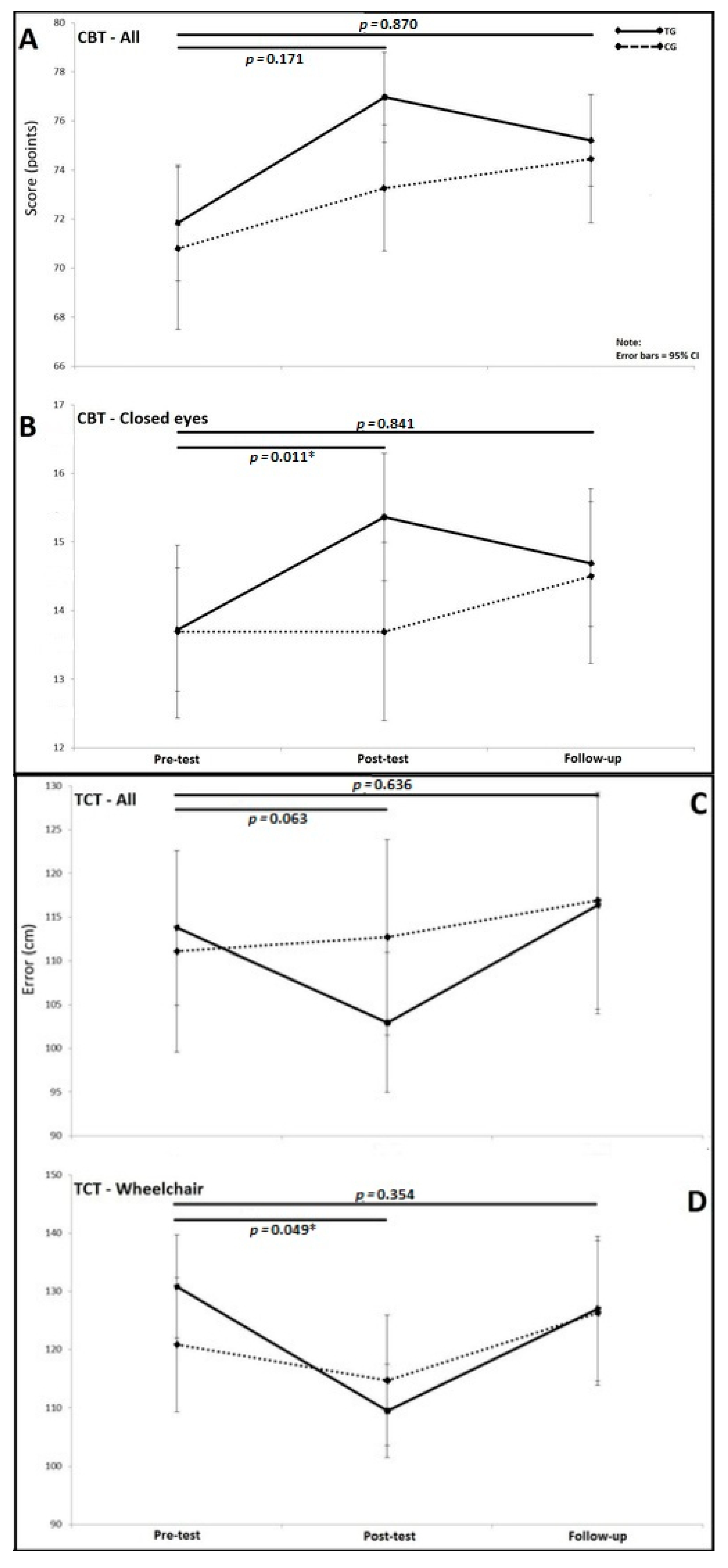
2.5.1. Clinical Balance Test (CBT)
- 3 points—stable stance, absence of body movements;
- 2 points—stable stance, attempted to stabilize body using arm movements;
- 1 point—stable stance, attempted to stabilize body using arm and leg movements;
- 0 points—loss of stance.
2.5.2. Triangle Completion Test (TCT)
2.6. MRI
2.7. Outcome Variables and Data Analysis
3. Results
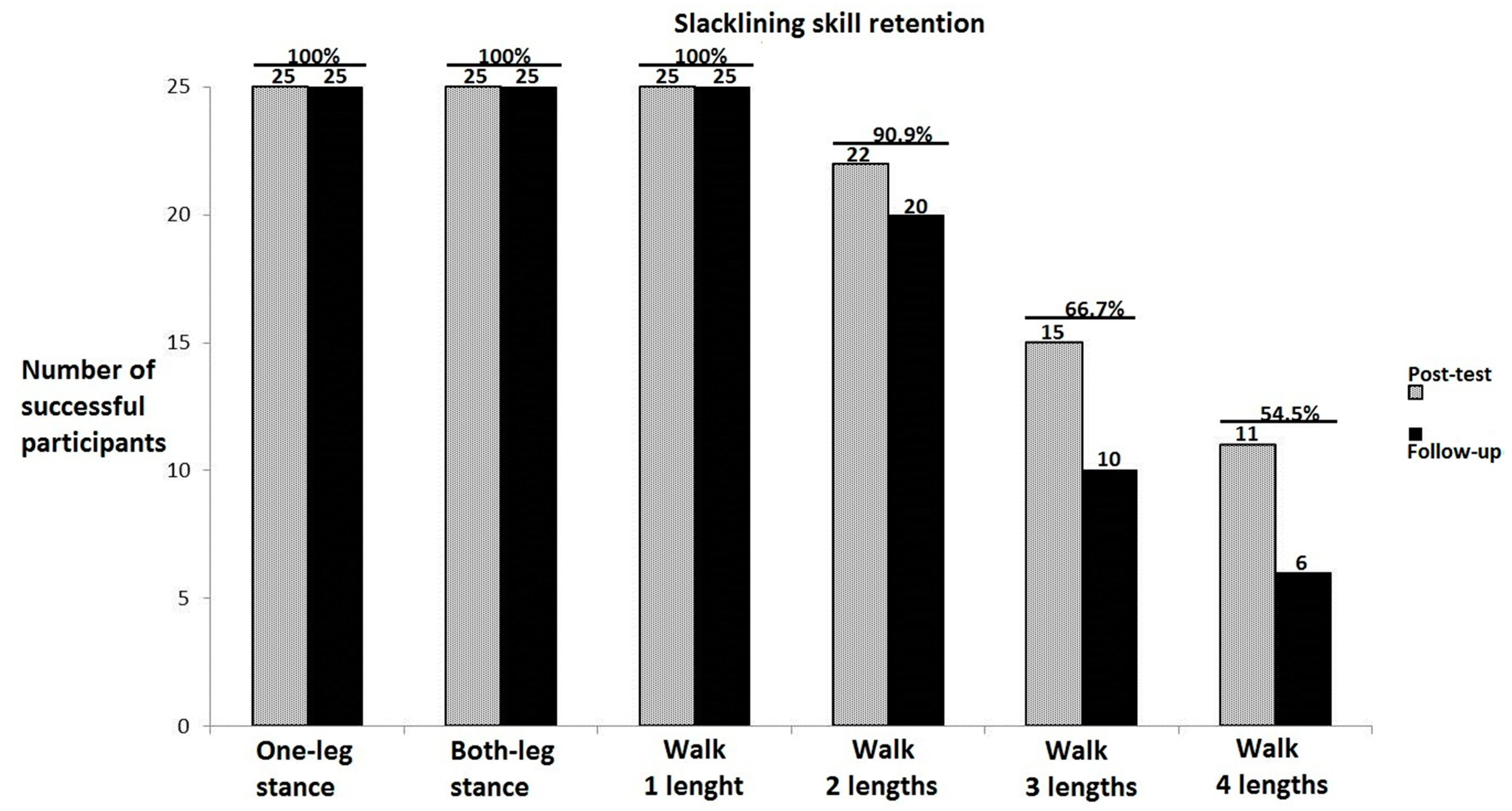
3.1. Training-Specific Skills
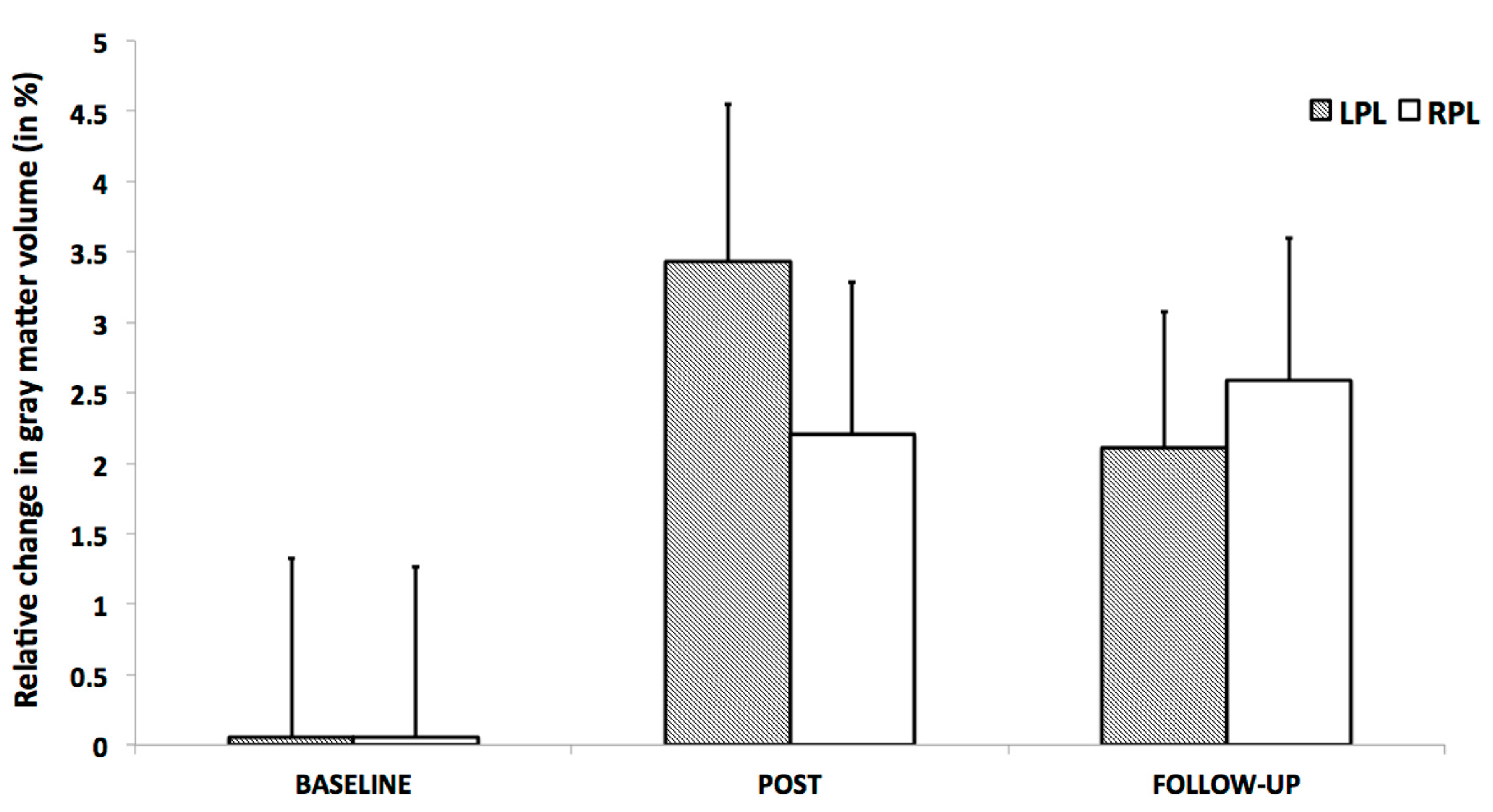
3.2. Triangle Completion Test (TCT)
3.3. Voxel-Based Morphometry (VBM)
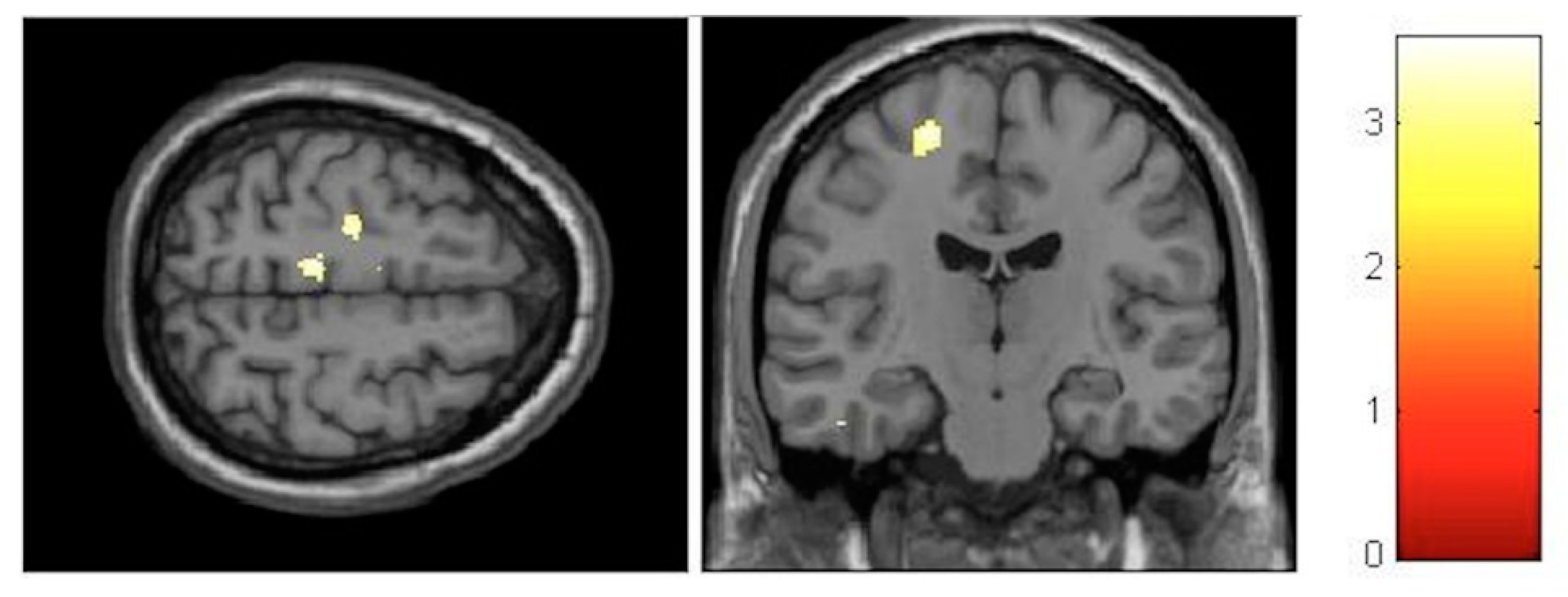
3.3.1. Interaction Effects
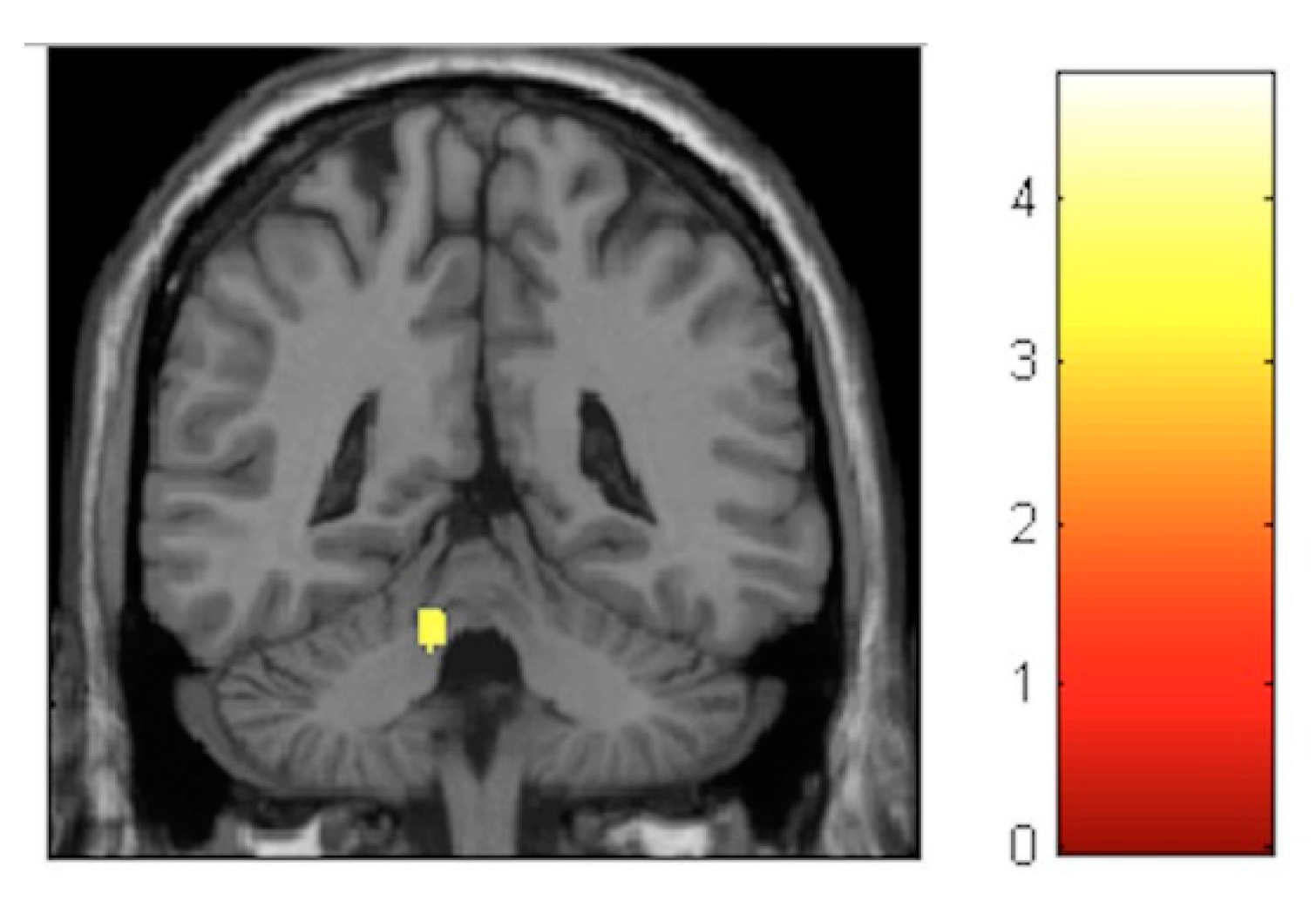
3.3.2. Within-Training-Group Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- Taubert, M.; Draganski, B.; Anwander, A.; Muller, K.; Horstmann, A.; Villringer, A.; Ragert, P. Dynamic Properties of Human Brain Structure: Learning-Related Changes in Cortical Areas and Associated Fiber Connections. J. Neurosci. 2010, 30, 11670–11677. [Google Scholar] [CrossRef] [Green Version]
- Taubert, M.; Mehnert, J.; Pleger, B.; Villringer, A. Rapid and specific gray matter changes in M1 induced by balance training. Neuroimage 2016, 133, 399–407. [Google Scholar] [CrossRef]
- Sehm, B.; Taubert, M.; Conde, V.; Weise, D.; Classen, J.; Dukart, J.; Draganski, B.; Villringer, A.; Ragert, P. Structural brain plasticity in parkinson’s disease induced by balance training. Neurobiol. Aging 2014, 35, 232–239. [Google Scholar] [CrossRef]
- Weber, B.; Koschutnig, K.; Schwerdtfeger, A.; Rominger, C.; Papousek, I.; Weiss, E.M.; Tilp, M.; Fink, A. Learning Unicycling Evokes Manifold Changes in Gray and White Matter Networks Related to Motor and Cognitive Functions. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Driemeyer, J.; Boyke, J.; Gaser, C.; Büchel, C.; May, A. Changes in gray matter induced by learning—Revisited. PLoS ONE 2008, 3, e2669. [Google Scholar] [CrossRef] [Green Version]
- Sampaio-Baptista, C.; Scholz, J.; Jenkinson, M.; Thomas, A.G.; Filippini, N.; Smit, G.; Douaud, G.; Johansen-Berg, H. Gray matter volume is associated with rate of subsequent skill learning after a long term training intervention. Neuroimage 2014, 96, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Seidel, O.; Carius, D.; Kenville, R.; Ragert, P. Motor learning in a complex balance task and associated neuroplasticity: A comparison between endurance athletes and non-athletes. J. Neurophysiol. 2017, 118, 1849–1860. [Google Scholar] [CrossRef]
- Müller, P.; Rehfeld, K.; Schmicker, M.; Hökelmann, A.; Dordevic, M.; Lessmann, V.; Brigadski, T.; Kaufmann, J.; Müller, N.G. Evolution of neuroplasticity in response to physical activity in old age: The case for dancing. Front. Aging Neurosci. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Dordevic, M.; Hökelmann, A.; Müller, P.; Rehfeld, K.; Müller, N.G. Improvements in Orientation and Balancing Abilities in Response to One Month of Intensive Slackline-Training. A Randomized Controlled Feasibility Study. Front. Hum. Neurosci. 2017, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Granacher, U.; Iten, N.; Roth, R.; Gollhofer, A. Slackline training for balance and strength promotion. Int. J. Sports Med. 2010, 31, 717–723. [Google Scholar] [CrossRef]
- Donath, L.; Roth, R.; Zahner, L.; Faude, O. Slackline Training (Balancing Over Narrow Nylon Ribbons) and Balance Performance: A Meta-Analytical Review. Sports Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.; Horak, F.B. The relevance of clinical balance assessment tools to differentiate balance deficits. Eur. J. Phys. Rehabil. Med. 2010, 46, 239–248. [Google Scholar] [PubMed]
- Karlsson, M.K.; Vonschewelov, T.; Karlsson, C.; Cöster, M.; Rosengen, B.E. Prevention of falls in the elderly: A review. Scand. J. Public Health 2013, 41, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Blanke, O. The thalamocortical vestibular system in animals and humans. Brain Res. Rev. 2011, 67, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, C.; Burgess, N. Neural mechanisms of self-location. Curr. Biol. 2014, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, G.; Van Schoor, N.M.; Cooper, R.; Tooth, L.; Kenny, R.A. Should prevention of falls start earlier? Coordinated analyses of harmonised data on falls in middle-aged adults across four population-based cohort studies. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Hitier, M.; Besnard, S.; Smith, P.F. Vestibular pathways involved in cognition. Front. Integr. Neurosci. 2014, 8, 59. [Google Scholar] [CrossRef]
- Stackman, R.W.; Clark, A.S.; Taube, J.S. Hippocampal spatial representations require vestibular input. Hippocampus 2002, 12, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Russell, N.A.; Horii, A.; Smith, P.F.; Darlington, C.L.; Bilkey, D.K. Long-term effects of permanent vestibular lesions on hippocampal spatial firing. J. Neurosci. 2003, 23, 6490–6498. [Google Scholar] [CrossRef]
- Brandt, T.; Schautzer, F.; Hamilton, D.A.; Br??ning, R.; Markowitsch, H.J.; Kalla, R.; Darlington, C.; Smith, P.; Strupp, M. Vestibular loss causes hippocampal atrophy and impaired spatial memory in humans. Brain 2005, 128, 2732–2741. [Google Scholar] [CrossRef]
- Hüfner, K.; Binetti, C.; Hamilton, D.A.; Stephan, T.; Flanagin, V.L.; Linn, J.; Labudda, K.; Markowitsch, H.; Glasauer, S.; Jahn, K.; et al. Structural and functional plasticity of the hippocampal formation in professional dancers and slackliners. Hippocampus 2011, 21, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Dordevic, M.; Schrader, R.; Taubert, M.; Müller, P.; Hökelmann, A.; Müller, N.G. Vestibulo-Hippocampal Function Is Enhanced and Brain Structure Altered in Professional Ballet Dancers. Front. Integr. Neurosci. 2018. [Google Scholar] [CrossRef] [PubMed]
- DiStefano, L.J.; Clark, M.A.; Padua, D.A. Evidence supporting balance training in healthy individuals: A systemic review. J. Strength Cond. Res. 2009, 23, 2718–2731. [Google Scholar] [CrossRef]
- Lesinski, M.; Hortobágyi, T.; Muehlbauer, T.; Gollhofer, A.; Granacher, U. Dose-Response Relationships of Balance Training in Healthy Young Adults: A Systematic Review and Meta-Analysis. Sport. Med. 2015, 45, 557–576. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Leckie, R.L.; Weinstein, A.M. Physical activity, fitness, and gray matter volume. Neurobiol. Aging 2014, 35. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.; Blanke, O.; Mast, F.W. The human vestibular cortex revealed by coordinate-based activation likelihood estimation meta-analysis. Neuroscience 2012, 212, 159–179. [Google Scholar] [CrossRef]
- Rogge, A.K.; Röder, B.; Zech, A.; Hötting, K. Exercise-induced neuroplasticity: Balance training increases cortical thickness in visual and vestibular cortical regions. Neuroimage 2018, 179, 471–479. [Google Scholar] [CrossRef]
- Hänggi, J.; Koeneke, S.; Bezzola, L.; Jäncke, L. Structural neuroplasticity in the sensorimotor network of professional female ballet dancers. Hum. Brain Mapp. 2010, 31, 1196–1206. [Google Scholar] [CrossRef]
- Makin, T.R.; Bensmaia, S.J. Stability of Sensory Topographies in Adult Cortex. Trends Cogn. Sci. 2017, 21, 195–204. [Google Scholar] [CrossRef]
- Georgopoulos, A.P.; Carpenter, A.F. Coding of movements in the motor cortex. Curr. Opin. Neurobiol. 2015, 33, 34–39. [Google Scholar] [CrossRef]
- Zu Eulenburg, P.; Caspers, S.; Roski, C.; Eickhoff, S.B. Meta-analytical definition and functional connectivity of the human vestibular cortex. Neuroimage 2012, 60, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Ferrè, E.R.; Day, B.L.; Bottini, G.; Haggard, P. How the vestibular system interacts with somatosensory perception: A sham-controlled study with galvanic vestibular stimulation. Neurosci. Lett. 2013, 550, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrè, E.R.; Kaliuzhna, M.; Herbelin, B.; Haggard, P.; Blanke, O. Vestibular-somatosensory interactions: Effects of passive whole-body rotation on somatosensory detection. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Ferrè, E.R.; Walther, L.E.; Haggard, P. Multisensory interactions between vestibular, visual and somatosensory signals. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.K.; Kim, H.J.; Lee, H.J. Changes in the gray matter volume during compensation after vestibular neuritis: A longitudinal VBM study. Restor. Neurol. Neurosci. 2014, 32, 663–673. [Google Scholar] [CrossRef]
- Helmchen, C.; Klinkenstein, J.; MacHner, B.; Rambold, H.; Mohr, C.; Sander, T. Structural changes in the human brain following vestibular neuritis indicate central vestibular compensation. Ann. N. Y. Acad. Sci. 2009, 1164, 104–115. [Google Scholar]
- Guldin, W.O.; Akbarian, S.; Grüsser, O.-J. Cortico-cortical connections and cytoarchitectonics of the primate vestibular cortex: A study in squirrel monkeys (Saimiri sciureus). J. Comp. Neurol. 1992, 326, 375–401. [Google Scholar] [CrossRef]
- Takakusaki, K. Functional Neuroanatomy for Posture and Gait Control. J. Mov. Disord. 2017, 10, 1–17. [Google Scholar] [CrossRef]
- Caligiore, D.; Pezzulo, G.; Baldassarre, G.; Bostan, A.C.; Strick, P.L.; Doya, K.; Helmich, R.C.; Dirkx, M.; Houk, J.; Jörntell, H.; et al. Consensus Paper: Towards a Systems-Level View of Cerebellar Function: The Interplay Between Cerebellum, Basal Ganglia, and Cortex. Cerebellum 2017, 16, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Blanke, O.; Perrig, S.; Thut, G.; Landis, T.; Seeck, M. Simple and complex vestibular responses induced by electrical cortical stimulation of the parietal cortex in humans. J. Neurol. Neurosurg. Psychiatry 2000, 69, 553–556. [Google Scholar] [CrossRef] [Green Version]
- Frank, S.M.; Greenlee, M.W. The parieto-insular vestibular cortex in humans: More than a single area? J. Neurophysiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Schindler, A.; Bartels, A. Integration of visual and non-visual self-motion cues during voluntary head movements in the human brain. Neuroimage 2018. [Google Scholar] [CrossRef] [PubMed]
- Uesaki, M.; Takemura, H.; Ashida, H. Computational neuroanatomy of human stratum proprium of interparietal sulcus. Brain Struct. Funct. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billington, J.; Billington, J. Neural mechanisms for discounting head-roll-induced retinal motion. J. Neurosci. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, S.M.; Pawellek, M.; Forster, L.; Langguth, B.; Schecklmann, M.; Greenlee, M.W. Attention networks in the parietooccipital cortex modulate activity of the human vestibular cortex during attentive visual processing. J. Neurosci. 2020. [Google Scholar] [CrossRef]
- Roberts, R.E.; Ahmad, H.; Arshad, Q.; Patel, M.; Dima, D.; Leech, R.; Seemungal, B.M.; Sharp, D.J.; Bronstein, A.M. Functional neuroimaging of visuo-vestibular interaction. Brain Struct. Funct. 2017. [Google Scholar] [CrossRef] [Green Version]
- Nigmatullina, Y.; Hellyer, P.J.; Nachev, P.; Sharp, D.J.; Seemungal, B.M. The neuroanatomical correlates of training-related perceptuo-reflex uncoupling in dancers. Cereb. Cortex 2015, 25, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Stoodley, C.J.; Limperopoulos, C. Structure–function relationships in the developing cerebellum: Evidence from early-life cerebellar injury and neurodevelopmental disorders. Semin. Fetal Neonatal Med. 2016, 21, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Chang, R. Anatomy of the vestibular system: A review. NeuroRehabilitation 2013, 32, 437–443. [Google Scholar] [CrossRef]
- Dieterich, M.; Bense, S.; Lutz, S.; Drzezga, A.; Stephan, T.; Bartenstein, P.; Brandt, T. Dominance for vestibular cortical function in the non-dominant hemisphere. Cereb. Cortex 2003, 13, 994–1007. [Google Scholar] [CrossRef] [Green Version]
- Grahn, J.A.; Parkinson, J.A.; Owen, A.M. The role of the basal ganglia in learning and memory: Neuropsychological studies. Behav. Brain Res. 2009, 199, 53–60. [Google Scholar] [CrossRef]
- Vitte, E.; Derosier, C.; Caritu, Y.; Berthoz, A.; Hasboun, D.; Soulié, D. Activation of the hippocampal formation by vestibular stimulation: A functional magnetic resonance imaging study. Exp. Brain Res. 1996, 112, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.F.; Zheng, Y. From ear to uncertainty: Vestibular contributions to cognitive function. Front. Integr. Neurosci. 2013, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Berthoz, A.; Israël, I.; Georges-François, P.; Grasso, R.; Tsuzuku, T. Spatial memory of body linear displacement: What is being stored? Science 1995, 269, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglói, K.; Doeller, C.F.; Berthoz, A.; Rondi-Reig, L.; Burgess, N. Lateralized human hippocampal activity predicts navigation based on sequence or place memory. Proc. Natl. Acad. Sci. USA 2010, 107, 14466–14471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.L.; Milner, B. The role of the right hippocampus in the recall of spatial location. Neuropsychologia 1981, 19, 781–793. [Google Scholar] [CrossRef]
- Healy, A.F.; Wohldmann, E.L.; Bourne, L.E. The Procedural Reinstatement Principle: Studies on Training, Retention, and Transfer. In Experimental Cognitive Psychology and Its Applications; APA: Washington, DC, USA, 2006; pp. 59–71. [Google Scholar]
- Dieterich, M.; Bense, S.; Stephan, T.; Yousry, T.A.; Brandt, T. FMRI signal increases and decreases in cortical areas during small-field optokinetic stimulation and central fixation. Exp. Brain Res. 2003, 148, 117–127. [Google Scholar] [CrossRef]
- Deutschländer, A.; Bense, S.; Stephan, T.; Schwaiger, M.; Brandt, T.; Dieterich, M. Sensory system interactions during simultaneous vestibular and visual stimulation in PET. Hum. Brain Mapp. 2002, 16, 92–103. [Google Scholar] [CrossRef]
- Kikuchi, M.; Naito, Y.; Senda, M.; Okada, T.; Shinohara, S.; Fujiwara, K.; Hori, S.-Y.; Tona, Y.; Yamazaki, H. Cortical activation during optokinetic stimulation—An fMRI study. Acta Otolaryngol. 2009, 129, 440–443. [Google Scholar] [CrossRef]
- Brandt, T.; Bartenstein, P.; Janek, A.; Dieterich, M. Reciprocal inhibitory visual-vestibular interaction. Visual motion stimulation deactivates the parieto-insular vestibular cortex. Brain 1998, 121, 1749–1758. [Google Scholar] [CrossRef] [Green Version]
- Lacour, M.; Helmchen, C.; Vidal, P.P. Vestibular compensation: The neuro-otologist’s best friend. J. Neurol. 2016, 263, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Ekstrom, A.D. Why vision is important to how we navigate. Hippocampus 2015, 25, 731–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shine, J.P.; Valdes-Herrera, J.P.; Hegarty, M.; Wolbers, T. The Human Retrosplenial Cortex and Thalamus Code Head Direction in a Global Reference Frame. J. Neurosci. 2016, 36, 6371–6381. [Google Scholar] [CrossRef] [PubMed]
- Callaert, D.V.; Ribbens, A.; Maes, F.; Swinnen, S.P.; Wenderoth, N. Assessing age-related gray matter decline with voxel-based morphometry depends significantly on segmentation and normalization procedures. Front. Aging Neurosci. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.G.; Marshall, S.W.; Batterham, A.M.; Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sports Exerc. 2009. [Google Scholar] [CrossRef] [Green Version]
- Gigerenzer, G. Mindless statistics. J. Socio. Econ. 2004. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Condition | Task | Points (Min = 0, Max = 3) | ||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | ||||
| 1. | Static—stable surface (floor) | Stand with feet together—open eyes | |||||
| 2. | Stand with feet together—closed eyes | ||||||
| 3. | One leg stance—left—open eyes | ||||||
| 4. | One leg stance—right—open eyes | ||||||
| 5. | One leg stance—left—closed eyes | ||||||
| 6. | One leg stance—right—closed eyes | ||||||
| 7. | Static—unstable surface (pad) | Stand normally (hip width stance)—open eyes | |||||
| 8. | Stand with feet together—open eyes | ||||||
| 9. | Stand normally (hip width stance)—closed eyes | ||||||
| 10. | Stand with feet together—closed eyes | ||||||
| 11. | One leg stance—left—open eyes | ||||||
| 12. | One leg stance—right—open eyes | ||||||
| 13. | One leg stance—left—closed eyes | ||||||
| 14. | One leg stance—right—closed eyes | ||||||
| 15. | Dynamic | Walk inside the zone (4 m × 30 cm) | Forward | ||||
| 16. | Turn (90°) | ||||||
| 17. | Backward | ||||||
| 18. | Walk on the line (4 m × 5 cm) | Forward | |||||
| 19. | Turn (90°) | ||||||
| 20. | Backward | ||||||
| 21. | Walk on the line with feet one after the other (4 m × 5 cm) | Forward | |||||
| 22. | Turn (90°) | ||||||
| 23. | Backward | ||||||
| 24. | Walk on the beam (4 m × 10 cm) | Forward | |||||
| 25. | Turn (90°) | ||||||
| 26. | Backward | ||||||
| 27. | Walk on the beam sideways (4 m × 10 cm) | Rightward | |||||
| 28. | Turn (90°) | ||||||
| 29. | Leftward | ||||||
| 30. | Walk on the line with closed eyes (4 m × 5 cm) | Forward | |||||
| Characteristic | Training (n = 25) | Control (n = 25) |
|---|---|---|
| Age (years) | 24.5 ± 2.7 | 23.2 ± 2.6 |
| Sex (females) | 11 (44%) | 12 (48%) |
| Weight (kg) | 69.1 ± 12.5 | 65.0 ± 10.0 |
| Height (cm) | 173.4 ± 9.2 | 170.3 ± 8.4 |
| Hours of activity (per week) | 3.0 ± 1.8 | 3.2 ± 2.5 |
| Handedness (right) | 24 (96%) | 23 (92%) |
| Profession (student) | 22 (88%) | 23 (92%) |
| Suffered a small injury (e.g., ankle sprain) | 5 (20%) | 5 (20%) |
| Ethnic origin | ||
| European | 20 (80%) | 19 (76%) |
| Asian (Indian) | 5 (20%) | 5 (20%) |
| Arabic | 0 (0%) | 1 (4%) |
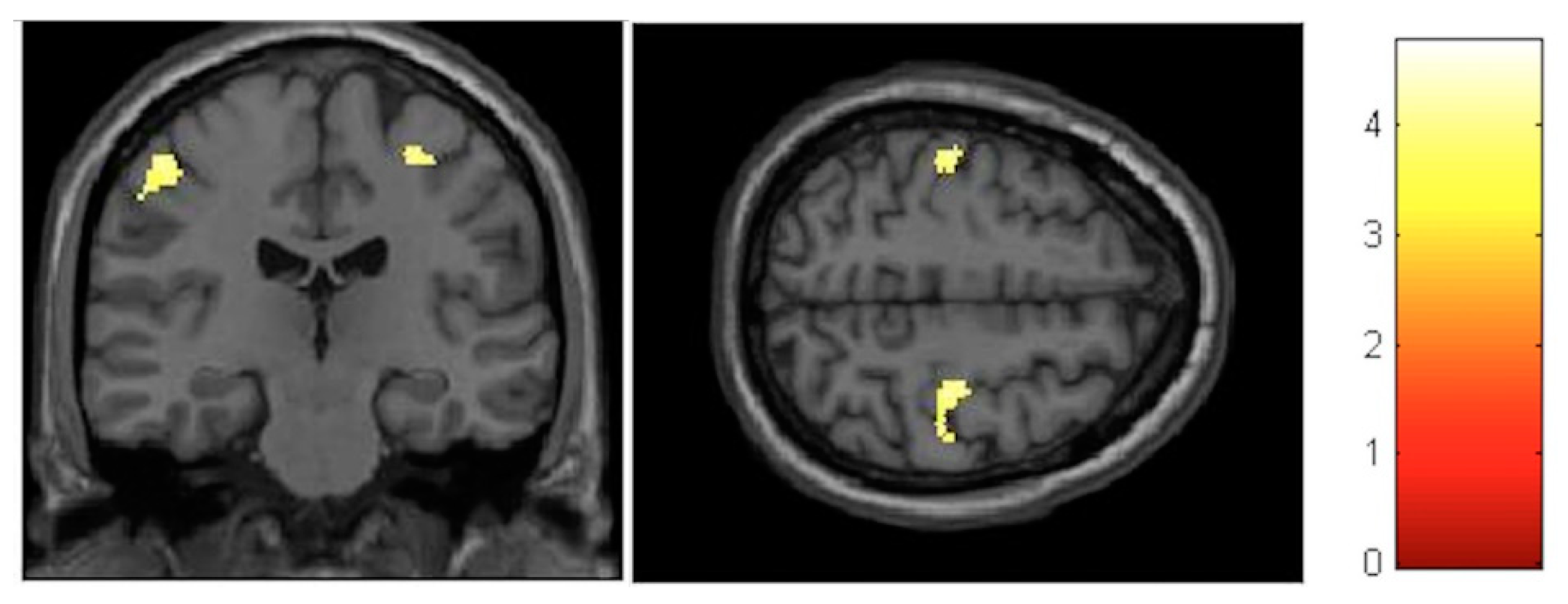
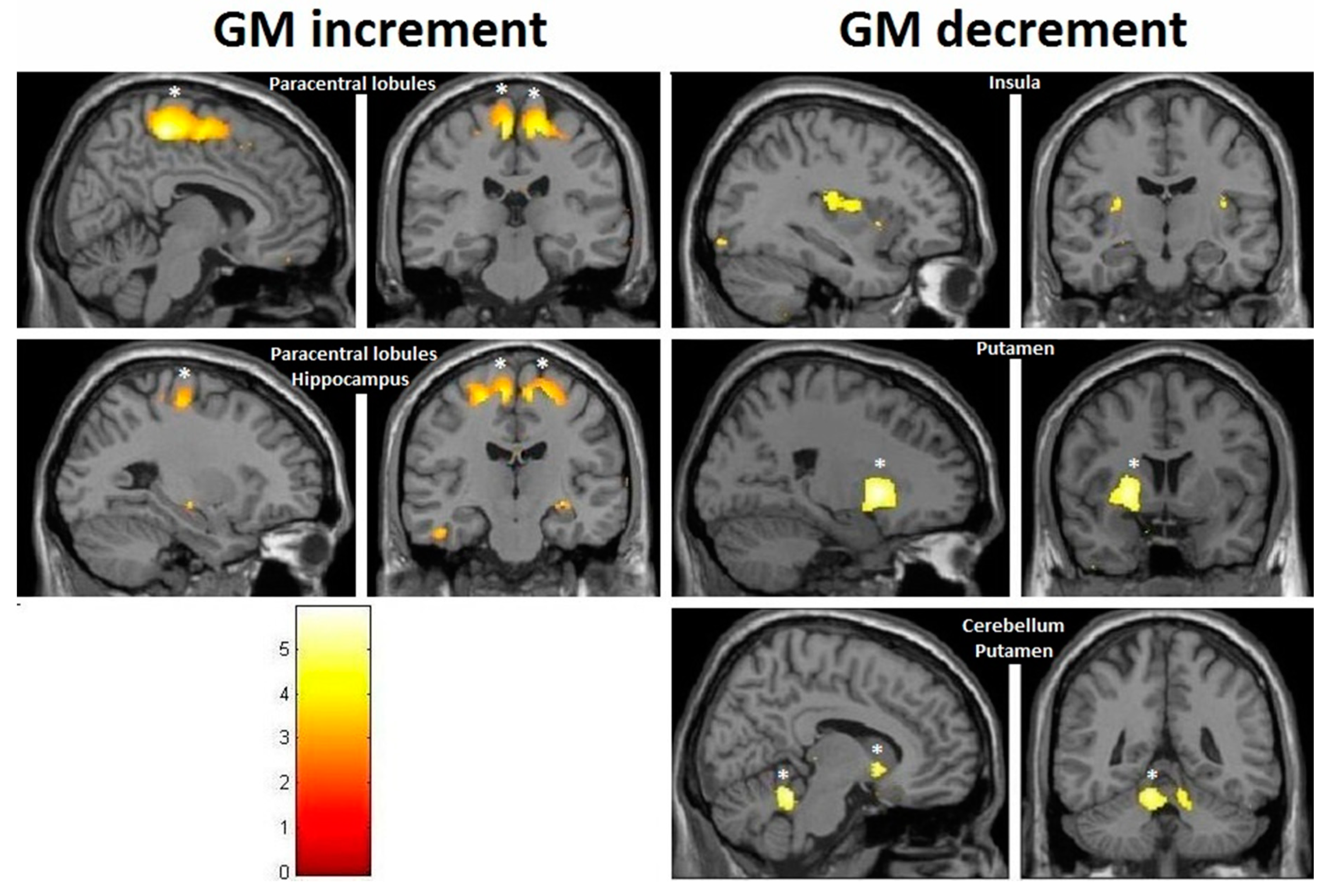
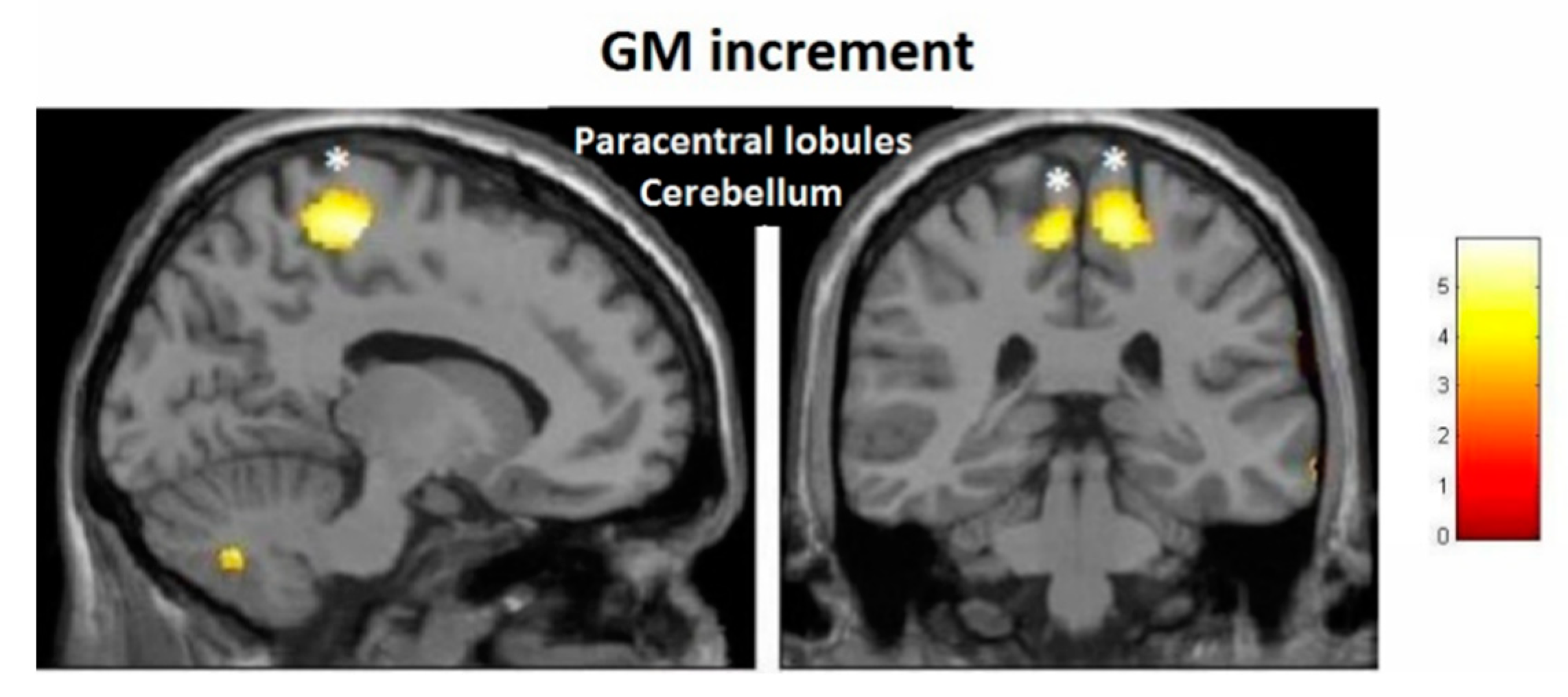
| Effect | Brain Region | Left Hemisphere MNI Coordinates (Cluster Size in Voxels) | T | d | Right Hemisphere MNI Coordinates (Cluster Size in Voxels) | T | d |
|---|---|---|---|---|---|---|---|
| Increment | Paracentral | −9, −32, 59 (1298) | 5.93 | 0.41 | 14, −36, 63 (1589) | 5.47 | 0.40 |
| Increment | Hippocampus | 34, −16, −11.5 (20) | 3.85 | 0.36 | |||
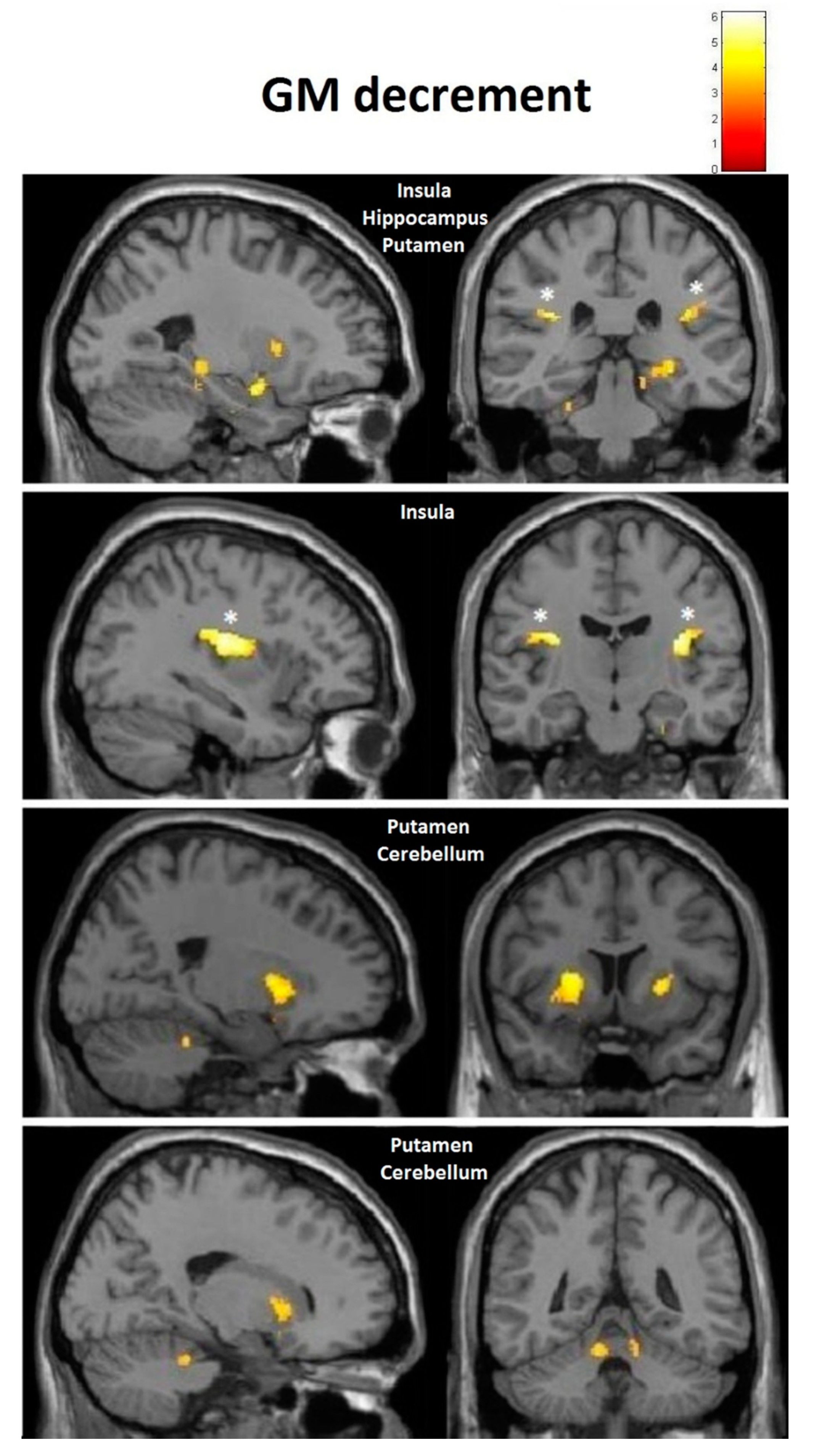
| Decrement | Insula | −32, −10, 12 (60) | 3.91 | 0.36 | 35, −20, 18 (261) | 4.36 | 0.38 |
| Decrement | Putamen | −20, 14, 0 (1879) | 5.45 | 0.40 | 24, 11, 2 (137) | 4.13 | 0.37 |
| Decrement | Cerebellum | −12, −47, −21 (511) | 5.33 | 0.40 | 11, −47, −23 (262) | 4.06 | 0.37 |
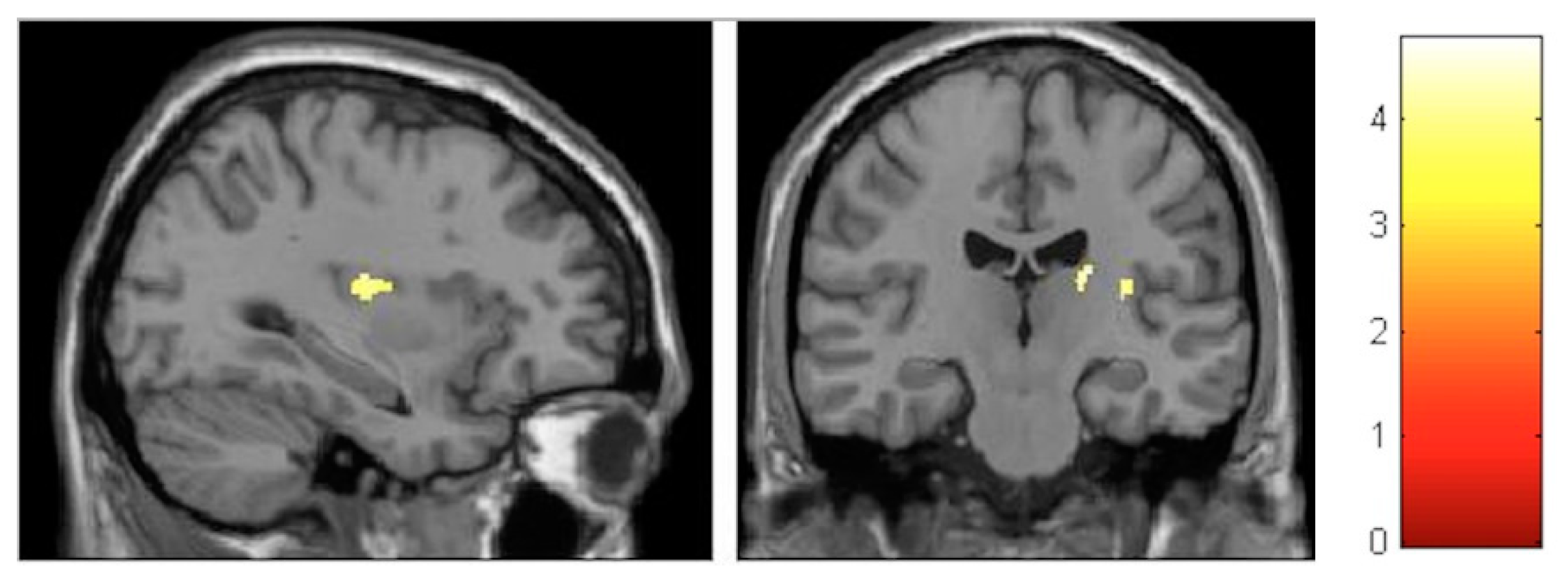
| Effect | Brain Region | Left Hemisphere MNI Coordinates (Cluster Size in Voxels) | T | d | Right Hemisphere MNI Coordinates (Cluster Size in Voxels) | T | d |
|---|---|---|---|---|---|---|---|
| Increment | Sensory-motor | −8, −32, 60 (314) | 5.17 | 0.39 | 11, −29, 60 (1087) | 6.09 | 0.41 |
| Decrement | Hippocampus | 27, −30, −6 (133) 23, −2, −17 (119) | 4.06 4.74 | 0.37 0.38 | |||
| Decrement | Insula | −33, −12, 18 (586) | 5.57 | 0.40 | 33, −17, 17 (797) | 6.17 | 0.41 |
| Decrement | Putamen | −26, 8, 6 (631) | 4.72 | 0.38 | 21, 11, 2 (134) | 4.22 | 0.38 |
| Decrement | Cerebellum | −11, −45, −23 (245) | 3.84 | 0.36 | 11, −65, −23 (224) | 3.99 | 0.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dordevic, M.; Taubert, M.; Müller, P.; Riemer, M.; Kaufmann, J.; Hökelmann, A.; Müller, N.G. Which Effects on Neuroanatomy and Path-Integration Survive? Results of a Randomized Controlled Study on Intensive Balance Training. Brain Sci. 2020, 10, 210. https://doi.org/10.3390/brainsci10040210
Dordevic M, Taubert M, Müller P, Riemer M, Kaufmann J, Hökelmann A, Müller NG. Which Effects on Neuroanatomy and Path-Integration Survive? Results of a Randomized Controlled Study on Intensive Balance Training. Brain Sciences. 2020; 10(4):210. https://doi.org/10.3390/brainsci10040210
Chicago/Turabian StyleDordevic, Milos, Marco Taubert, Patrick Müller, Martin Riemer, Jörn Kaufmann, Anita Hökelmann, and Notger G. Müller. 2020. "Which Effects on Neuroanatomy and Path-Integration Survive? Results of a Randomized Controlled Study on Intensive Balance Training" Brain Sciences 10, no. 4: 210. https://doi.org/10.3390/brainsci10040210