Genotoxic Effect of Dicyclopropanated 5-Vinyl-2-Norbornene

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Culture Medium and Growth Conditions31
2.3. Measurement of Bioluminescence
2.4. Data Processing
2.5. P1 Transduction
2.6. Chemicals
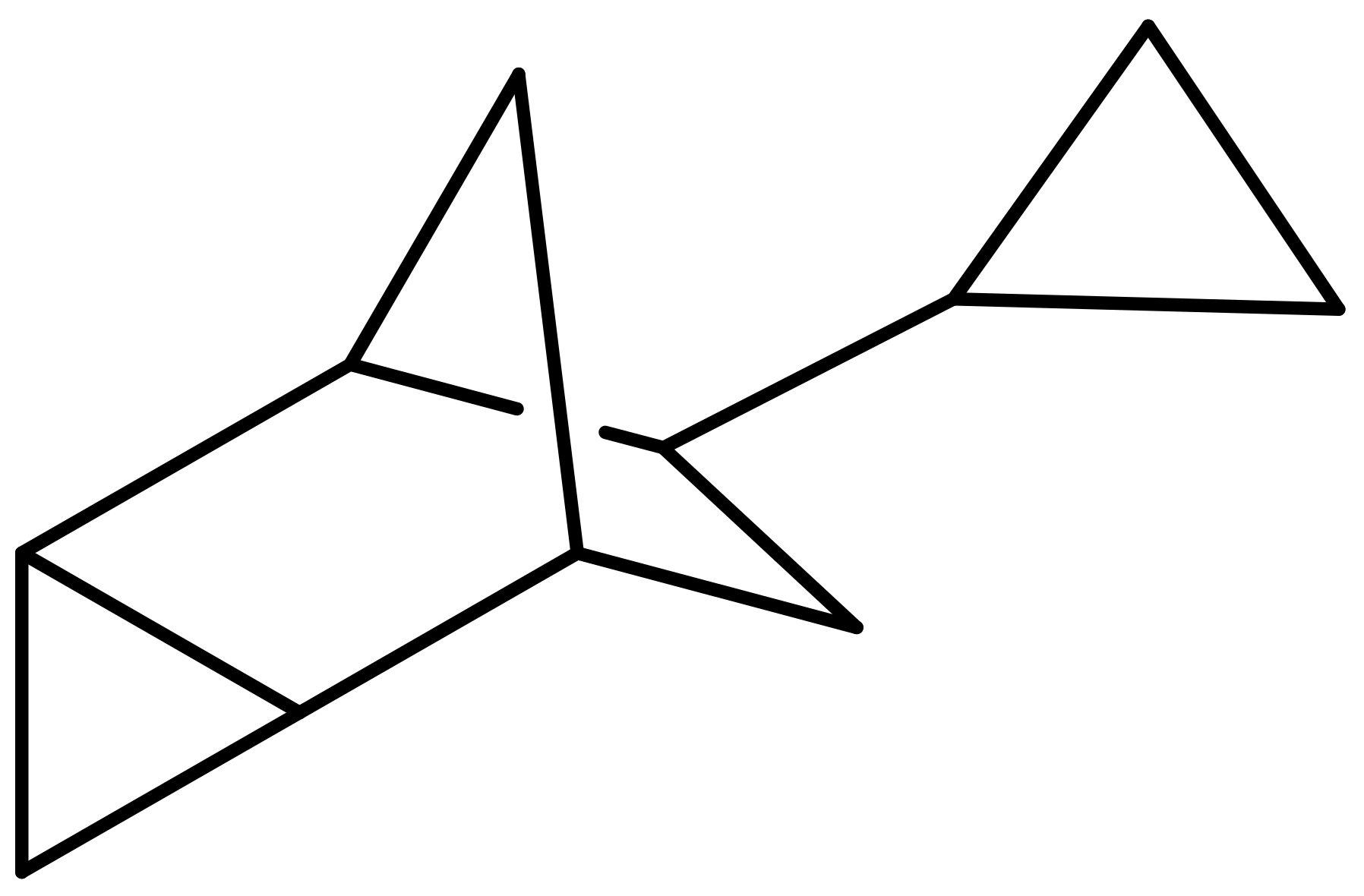
2.7. Synthesis of Dicyclopropanated 5-Vinyl-2-Norbornene (dcpVNB)
3. Results
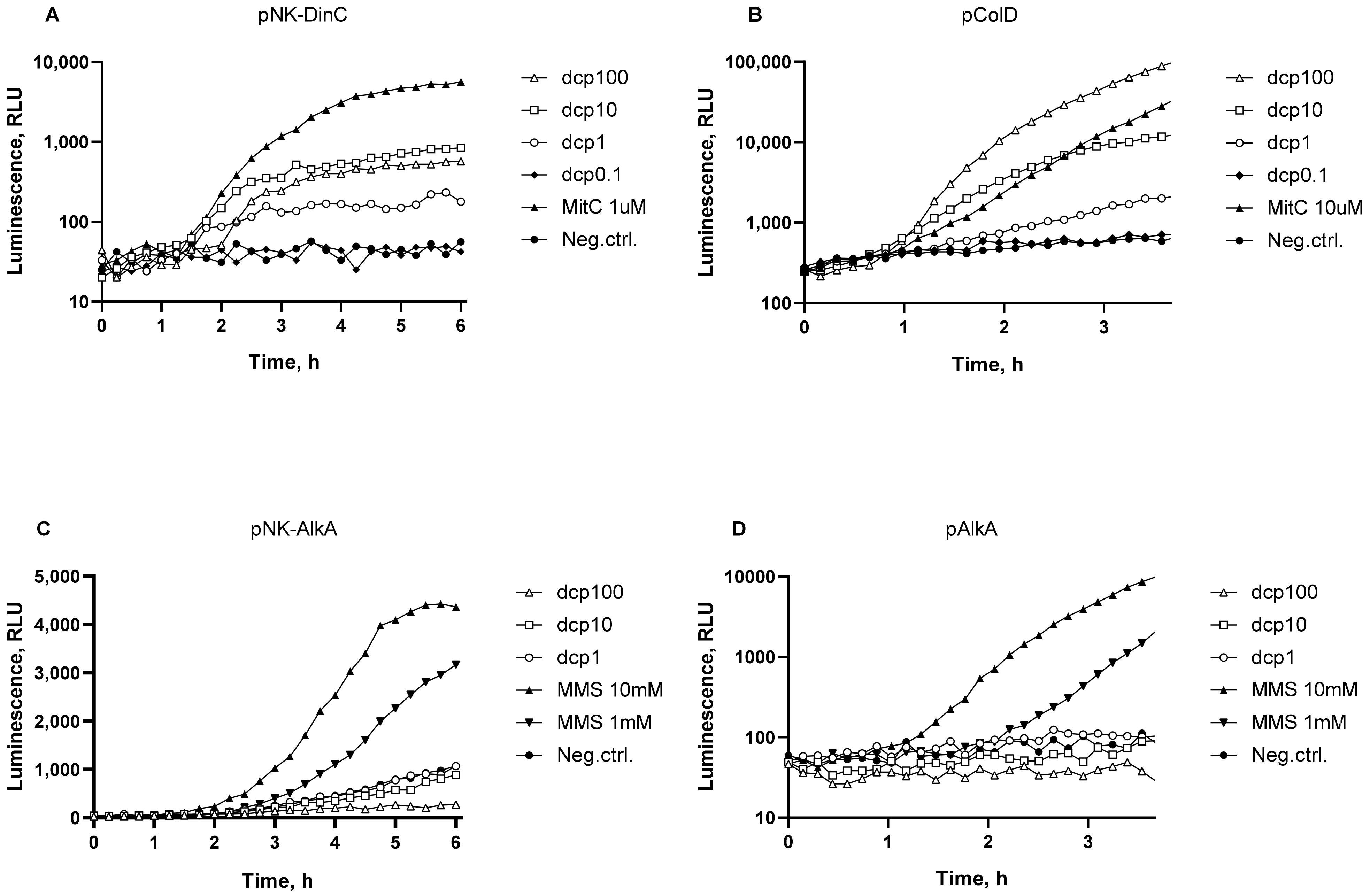
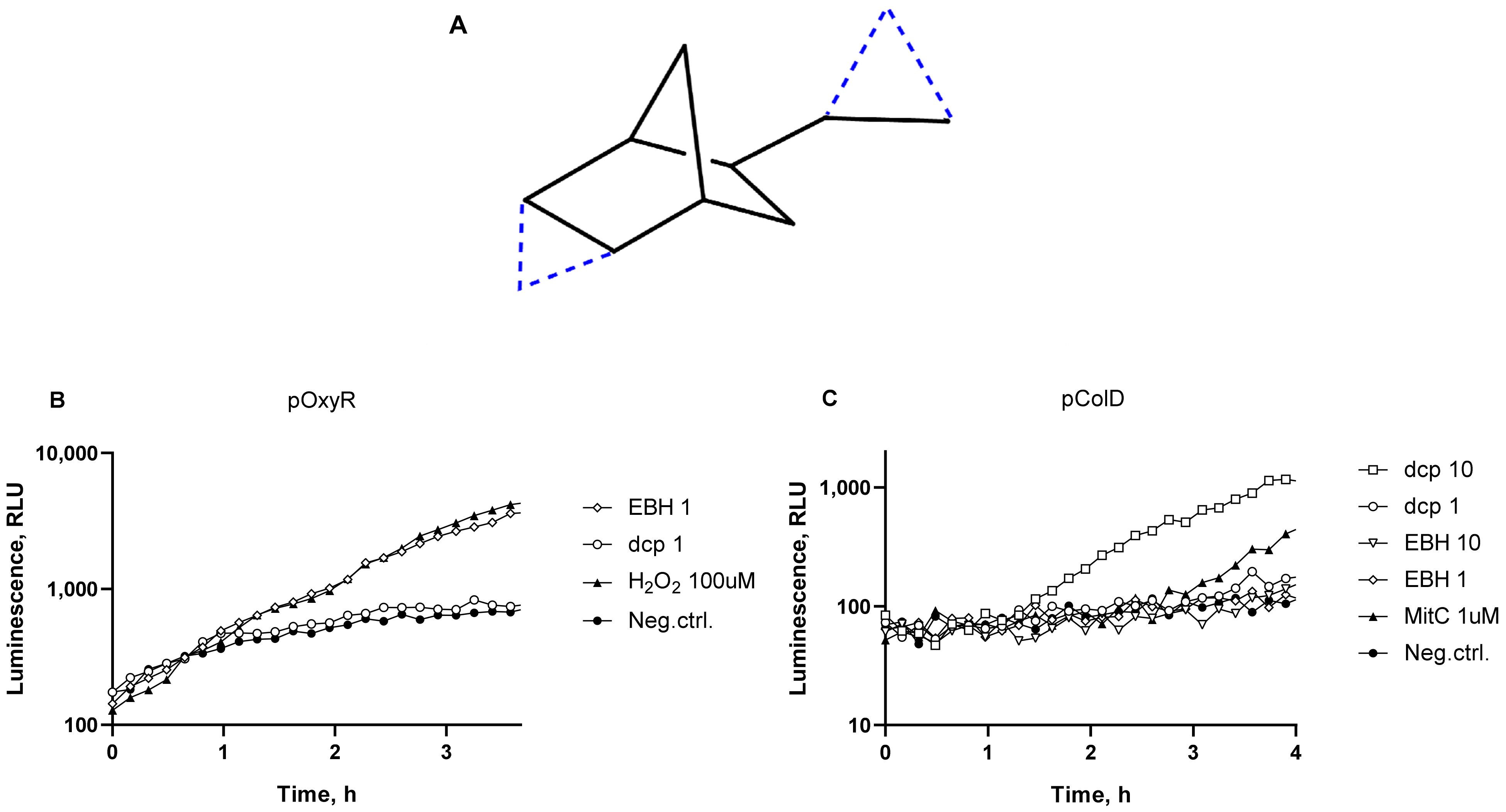
3.1. DNA Damage: SOS Response and DNA Alkylation
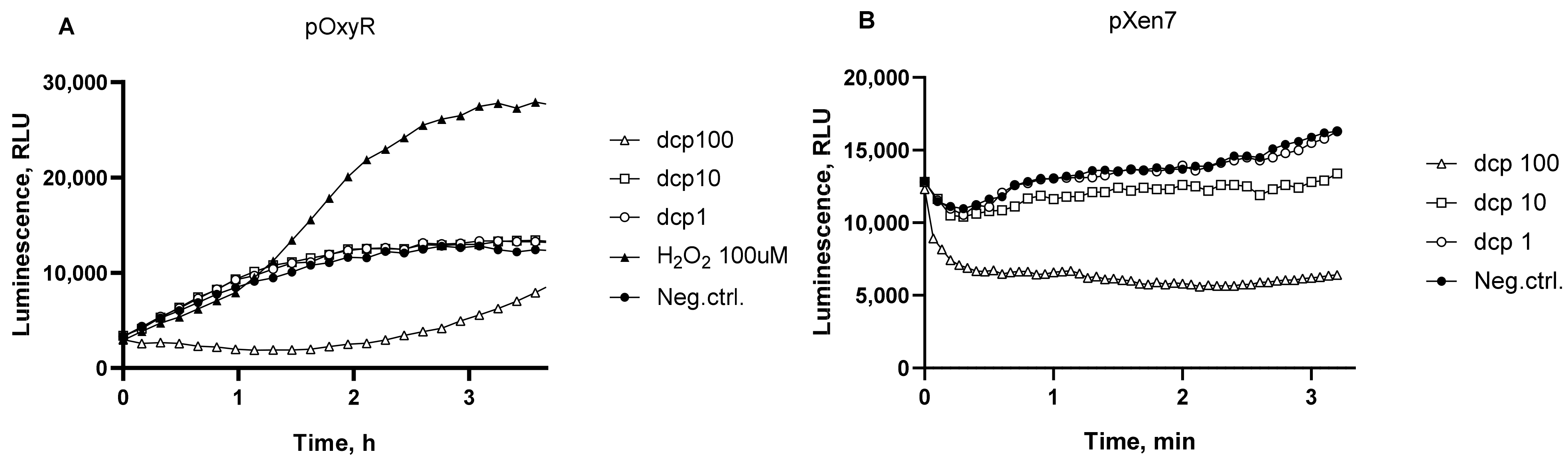
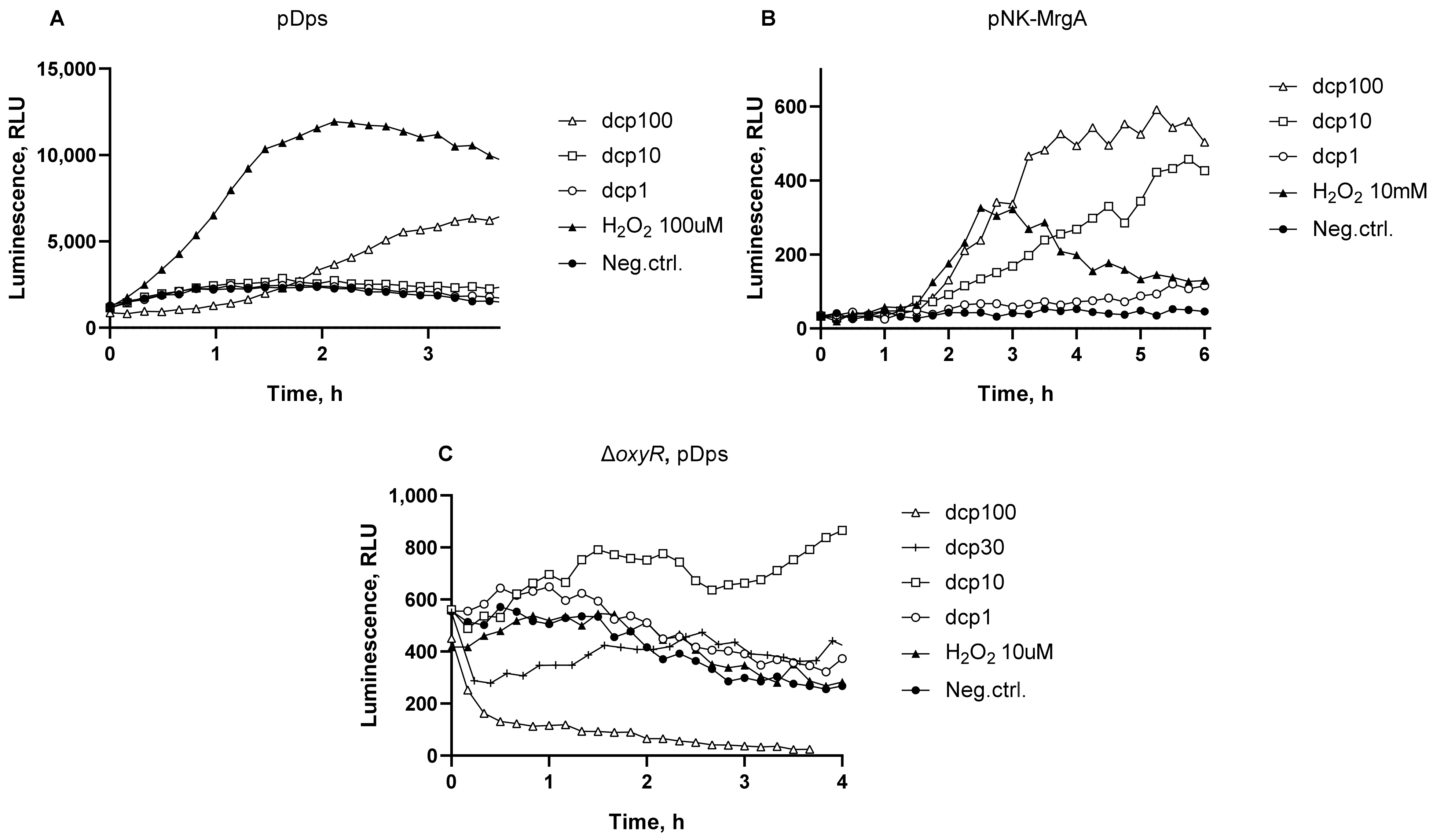
3.2. Oxidative Stress
3.3. Comparison of SOS Response and Oxidative Stress in Cells from dcpVNB and EBH
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sirjean, B.; Glaude, P.A.; Ruiz-Lopez, M.F.; Fournet, R. Detailed Kinetic Study of the Ring Opening of Cycloalkanes by CBS-QB3 Calculations. J. Phys. Chem. A 2006, 110, 12693–12704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Back, K.C.; Carter, V.L.; Thomas, A.A. Occupational Hazards of Missile Operations with Special Regard to the Hydrazine Propellants. Aviat Space Environ. Med. 1978, 49, 591–598. [Google Scholar] [PubMed]
- Hu, C.; Zhang, Y.; Zhou, Y.; Liu, Z.F.; Feng, X.S. Unsymmetrical Dimethylhydrazine and Related Compounds in the Environment: Recent Updates on Pretreatment, Analysis, and Removal Techniques. J. Hazard Mater. 2022, 432, 128708. [Google Scholar] [CrossRef] [PubMed]
- Kenessov, B.; Batyrbekova, S. Actual Directions in Study of Ecological Consequences of a Highly Toxic 1,1-Dimethylhydrazine-Based Rocket Fuel Spills. Chem. Bull. Kazakh Natl. Univ. 2012, 66, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Kenessov, B.; Batyrbekova, S.; Nauryzbayev, M.; Bekbassov, T.; Alimzhanova, M.; Carlsen, L. GC-MS Determination of 1-Methyl-1H-1,2,4-Triazole in Soils Affected by Rocket Fuel Spills in Central Kazakhstan. Chromatographia 2008, 67, 421–424. [Google Scholar] [CrossRef]
- Carlsen, L.; Kenesova, O.A.; Batyrbekova, S.E. A Preliminary Assessment of the Potential Environmental and Human Health Impact of Unsymmetrical Dimethylhydrazine as a Result of Space Activities. Chemosphere 2007, 67, 1108–1116. [Google Scholar] [CrossRef]
- Carlsen, L.; Kenessov, B.N.; Batyrbekova, S.Y. A QSAR/QSTR Study on the Human Health Impact of the Rocket Fuel 1,1-Dimethyl Hydrazine and Its Transformation Products. Multicriteria Hazard Ranking Based on Partial Order Methodologies. Environ. Toxicol Pharm. 2009, 27, 415–423. [Google Scholar] [CrossRef]
- Goryanin, I.; Kotova, V.; Krasnopeeva, E.; Manukhov, I.; Chubukov, P.; Balabanov, V.; Chalkin, S.; Shatrov, T.; Zavilgelsky, G. Genotoxic Action of the 1,1-Dimethylhydrazine Determined by Alkylating Compounds Appearing in the Result of Oxidation and Hydrogen Peroxide. Tr. MIPT 2013, 5, 103–111. [Google Scholar]
- Kessenikh, A.; Manukhov, I.; Vagapova, E.; Vysokikh, M.; Konopleva, M.; Kotova, V.; Gorbunov, M.; Chalkin, S.; Zavilgelsky, G. Lux-Biosensors Set for Detection of Toxic Products of Incomplete Oxidation of Non-Symmetric Dimethylhydrazine in Medium. Patent RU2626569, 17 December 2015. [Google Scholar]
- Manukhov, I.; Gorbunov, M.; Degtev, I.; Zavilgelsky, G.; Kessenikh, A.; Konopleva, M.; Kotova, V.; Krasnopeeva, E.; Motovilov, K.; Osetrova, M.; et al. The Set of Lux-Biosensors for Determination of Genotoxic Products of Incomplete Oxidation of Unsymmetrical Dimethyl Hydrazine in the Environment. Patent RU2569156, 18 December 2014. [Google Scholar]
- Zavilgelsky, G.; Kotova, V.; Manukhov, I.; Kondratyev, A.; Sambros, V.; Shatrov, Y.; Chalkin, S. Kit of Lux-Biosensors for Heptyl Detection in Medium. Patent RU229745, 1 December 2007. [Google Scholar]
- Zavilgelsky, G.B.; Kotova, V.Y.; Manukhov, I.V. Action of 1,1-Dimethylhydrazine on Bacterial Cells Is Determined by Hydrogen Peroxide. Mutat. Res. Genet. Toxicol. Environ. Mutagen 2007, 634, 172–176. [Google Scholar] [CrossRef]
- Dóbé, S.; Turányi, T.; Bérces, T.; Márta, F. The Kinetics of Hydroxyl Radical Reactions with Cyclopropane and Cyclobutane. Proc. Indian Acad. Sci. Chem. Sci. 1991, 103, 499–503. [Google Scholar] [CrossRef]
- Kessenikh, A.G.; Manukhov, I.V.; Yaguzhinsky, L.S.; Bermeshev, M.V.; Zisman, M.A.; Pevgov, V.G.; Samoilov, V.O.; Shorunov, S.V.; Maksimov, A.L. Toxic Effect of 2-Ethy l(Bicyclo[2.2.1] Heptane) on Bacterial Cells. Biotekhnologiya 2019, 35, 67–72. [Google Scholar]
- Kessenikh, A.; Gnuchikh, E.; Bazhenov, S.; Bermeshev, M.; Pevgov, V.; Samoilov, V.; Shorunov, S.; Maksimov, A.; Yaguzhinsky, L.; Manukhov, I. Genotoxic Effect of 2,2’-Bis(Bicyclo[2.2.1] Heptane) on Bacterial Cells. PLoS ONE 2020, 15, e0228525. [Google Scholar] [CrossRef]
- Kessenikh, A.G.; Novoyatlova, U.S.; Bazhenov, S.V.; Stepanova, E.A.; Khrulnova, S.A.; Gnuchikh, E.Y.; Kotova, V.Y.; Kudryavtseva, A.A.; Bermeshev, M.V.; Manukhov, I. V Constructing of Bacillus subtilis-Based Lux-Biosensors with the Use of Stress-Inducible Promoters. Int. J. Mol. Sci. 2021, 22, 9571. [Google Scholar] [CrossRef] [PubMed]
- Kotova, V.Y.; Manukhov, I.v.; Zavilgelskii, G.B. Lux-Biosensors for Detection of SOS Response, Heat Shock, and Oxidative Stress. Appl. Biochem. Microbiol. 2010, 46, 781–788. [Google Scholar] [CrossRef]
- Zhu, Y.; Elcin, E.; Jiang, M.; Li, B.; Wang, H.; Zhang, X.; Wang, Z. Use of Whole-Cell Bioreporters to Assess Bioavailability of Contaminants in Aquatic Systems. Front. Chem. 2022, 10, 1018124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, B.; Schillereff, D.N.; Chiverrell, R.C.; Tefsen, B.; Wells, M. Whole-Cell Biosensors for Determination of Bioavailable Pollutants in Soils and Sediments: Theory and Practice. Sci. Total Environ. 2022, 811, 152178. [Google Scholar] [CrossRef]
- Zavilgelsky, G.B.; Zarubina, A.P.; Manukhov, I.v. Sequencing and Comparative Analysis of the Lux Operon of Photorhabdus Luminescens Strain Zm1: ERIC Elements as Putative Recombination Hot Spots. Mol. Biol. 2002, 36, 637–647. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006-0008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnuchikh, E.Y.; Manukhov, I.V.; Zavilgelsky, G. Biosensors for the Determination of Promoters and Chaperones Activity in Bacillus subtilis Cells. Biotekhnologiya 2020, 36, 68–77. [Google Scholar] [CrossRef]
- Van Dyk, T.K.; Rosson, R.A. Photorhabdus Luminescens LuxCDABE Promoter Probe Vectors. Methods Mol. Biol. 1998, 102, 85–95. [Google Scholar]
- Melkina, O.E.; Goryanin, I.I.; Zavilgelsky, G.B. The DNA-Mimic Antirestriction Proteins ArdA ColIB-P9, Arn T4, and Ocr T7 as Activators of H-NS-Dependent Gene Transcription. Microbiol. Res. 2016, 192, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Thomason, L.C.; Costantino, N.; Court, D.L. E. coli Genome Manipulation by P1 Transduction. Curr. Protoc. Mol. Biol. 2007, 79, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Langsdorf, L.; Burtovyy, O. Cycloalkylnorbornene Monomers, Polymers Suitable for Use as Pervaporation Membrane Films. U.S. Patent WO25735, 13 February 2014. [Google Scholar]
- Bermeshev, M.V.; Chapala, P.P. Addition Polymerization of Functionalized Norbornenes as a Powerful Tool for Assembling Molecular Moieties of New Polymers with Versatile Properties. Prog. Polym. Sci. 2018, 84, 1–46. [Google Scholar] [CrossRef]
- Ushakov, N.V. Selective Hydrogenation of 5-Vinylnorborn-2-Ene and Other Methods for the Synthesis of 2-Vinylnorbornane. Russ. J. Appl. Chem. 2018, 91, 728–745. [Google Scholar] [CrossRef]
- Zarezin, D.P.; Rudakova, M.A.; Shorunov, S.V.; Sultanova, M.U.; Samoilov, V.O.; Maximov, A.L.; Bermeshev, M.V. Design and Preparation of Liquid Polycyclic Norbornanes as Potential High Performance Fuels for Aerospace Propulsion. Fuel Process. Technol. 2022, 225, 107056. [Google Scholar] [CrossRef]
- Shorunov, S.V.; Zarezin, D.P.; Samoilov, V.O.; Rudakova, M.A.; Borisov, R.S.; Maximov, A.L.; Bermeshev, M.V. Synthesis and Properties of High-Energy-Density Hydrocarbons Based on 5-Vinyl-2-Norbornene. Fuel 2021, 283, 118935. [Google Scholar] [CrossRef]
- Almiron, M.; Link, A.J.; Furlong, D.; Kolter, R. A Novel DNA-Binding Protein with Regulatory and Protective Roles in Starved Escherichia coli. Genes Dev. 1992, 6, 2646–2654. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; James, L.P.; Helmann, J.D. Metalloregulation in Bacillus subtilis: Isolation and Characterization of Two Genes Differentially Repressed by Metal Ions. J. Bacteriol. 1993, 175, 5428–5437. [Google Scholar] [CrossRef] [Green Version]
- Altuvia, S.; Almiron, M.; Huisman, G.; Kolter, R.; Storz, G. The Dps Promoter Is Activated by OxyR during Growth and by IHF and Σs in Stationary Phase. Mol. Microbiol. 1994, 13, 265–272. [Google Scholar] [CrossRef]
- Helmann, J.D.; Wu, M.F.W.; Gaballa, A.; Kobel, P.A.; Morshedi, M.M.; Fawcett, P.; Paddon, C. The Global Transcriptional Response of Bacillus subtilis to Peroxide Stress Is Coordinated by Three Transcription Factors. J. Bacteriol. 2003, 185, 243. [Google Scholar] [CrossRef] [Green Version]
- Sudarev, V.V.; Dolotova, S.M.; Bukhalovich, S.; Bazhenov, S.V.; Ryzhykau, Y.L.; Uversky, V.N.; Bondarev, N.A.; Osipov, S.D.; Mikhailov, A.E.; Kuklina, D.D.; et al. Ferritin Self-Assembly, Structure, Function, and Biotechnological Applications. Int. J. Biol. Macromol. 2022, 224, 319–343. [Google Scholar] [CrossRef] [PubMed]
- Manukhov, I.; Balabanov, V.; Kotova, V.; Khrulnova, S.; Melkina, O.; Kraynov, A.; Pustovoit, K.; Krechetov, P.; Koroleva, T.; Shatrov, T. Use of Lux Biosensors for Detection of UDMH in Soil. Dual Technol. 2008, 44, 50–56. [Google Scholar]
- Møller, P.; Scholten, R.H.; Roursgaard, M.; Krais, A.M. Inflammation, Oxidative Stress and Genotoxicity Responses to Biodiesel Emissions in Cultured Mammalian Cells and Animals. Crit. Rev. Toxicol. 2020, 50, 383–401. [Google Scholar] [CrossRef] [PubMed]
- Sazykin, I.; Makarenko, M.; Khmelevtsova, L.; Seliverstova, E.; Rakin, A.; Sazykina, M. Cyclohexane, Naphthalene, and Diesel Fuel Increase Oxidative Stress, CYP153, SodA, and RecA Gene Expression in Rhodococcus erythropolis. Microbiologyopen 2019, 8, e00855. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Source | |
|---|---|---|---|
| Strains | |||
| E. coli MG1655 | F- ilvG rfb-50 rph 1 | (VKPM, Russia) | |
| E. coli BW25113 ΔoxyR | Δ(araD-araB)567 ΔlacZ4787(::rrnB-3) λ- rph-1 Δ(rhaD-rhaB)568 hsdR514 ΔoxyR::Kmr | [21] | |
| E. coli MK2022 | ΔoxyR::Kmr (obtained by transfer of gene deletion ΔoxyR from BW25113 ΔoxyR to MG1655 using P1 transduction) | This study | |
| B. subtilis 168 | trpC2 | (VKPM, Russia) | |
| Plasmids | |||
| pNK-AlkA | pPL_ABCDExen vector [22] with insertion of the B. subtilis PalkA promoter; PalkA is transcriptionally fused with luxCDABE P. luminescens. Trimethoprim (Tpr), chloramphenicol (Cmr), and ampicillin (Apr) resistance. | [16] | |
| pNK-DinC | As pNK-AlkA, but B. subtilis PdinC promoter is used. Tpr, Cmr, Apr | [16] | |
| pNK-MrgA | As pNK-AlkA, but B. subtilis PmrgA promoter is used. Tpr, Cmr, Apr | [16] | |
| pAlkA-lux | pDEW201 [23] vector with insertion of E. coli PalkA promoter transcriptionally fused with luxCDABE P. luminescens. Apr | [8] | |
| pDps | As pAlkA-lux, but E. coli Pdps promoter is used. Apr | [24] | |
| pOxyR-lux | As pAlkA-lux, but E. coli PoxyS promoter with the gene oxyR is used. Apr | [17] | |
| pColD-lux | As pAlkA-lux, but Pcda from plasmid ColD-CA23 is used. Apr | [17] | |
| pXen7 | pUC18-based plasmid constitutively expressing luxCDABE genes. Apr | [20] | |
| Biosensors | dcpVNB, mM | EBH, mM | UDMH *, mM | Note |
|---|---|---|---|---|
| E. coli pAlkA-lux | nd ** | nd | 2*10−2 | Alkylation of DNA |
| B. subtilis pNK-AlkA | nd | ne *** | ne | |
| E. coli pColD-lux | 2.3 ± 0.7 | 5.6 ± 1.9 | 8*10−3 | DNA damage, leading to SOS response |
| B. subtilis pNK-DinC | 3.5 ± 1.2 | ne | ne | |
| E. coli pDpS | 54 ± 8 | ne | ne | Oxidative stress |
| B. subtilis pNK-MrgA | 6.0 ± 2.1 | ne | ne | |
| E. coli pOxyR-lux | 470 ± 50 | 4.2 ± 1.8 | 3*10−3 | Oxidation by hydrogen peroxide |
| E. coli pXen7 | 67 ± 15 | 43 ± 7 | 2 | Total toxicity, decrease in luminescence correlates with the number of living cells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novoyatlova, U.S.; Kessenikh, A.G.; Kononchuk, O.V.; Bazhenov, S.V.; Fomkin, A.A.; Kudryavtseva, A.A.; Shorunov, S.V.; Bermeshev, M.V.; Manukhov, I.V. Genotoxic Effect of Dicyclopropanated 5-Vinyl-2-Norbornene. Biosensors 2023, 13, 57. https://doi.org/10.3390/bios13010057
Novoyatlova US, Kessenikh AG, Kononchuk OV, Bazhenov SV, Fomkin AA, Kudryavtseva AA, Shorunov SV, Bermeshev MV, Manukhov IV. Genotoxic Effect of Dicyclopropanated 5-Vinyl-2-Norbornene. Biosensors. 2023; 13(1):57. https://doi.org/10.3390/bios13010057
Chicago/Turabian StyleNovoyatlova, Uliana S., Andrei G. Kessenikh, Olga V. Kononchuk, Sergey V. Bazhenov, Alexander A. Fomkin, Anna A. Kudryavtseva, Sergey V. Shorunov, Maxim V. Bermeshev, and Ilya V. Manukhov. 2023. "Genotoxic Effect of Dicyclopropanated 5-Vinyl-2-Norbornene" Biosensors 13, no. 1: 57. https://doi.org/10.3390/bios13010057