Extracellular Vesicles (EVs) and Pancreatic Cancer: From the Role of EVs to the Interference with EV-Mediated Reciprocal Communication



Abstract
:1. Introduction
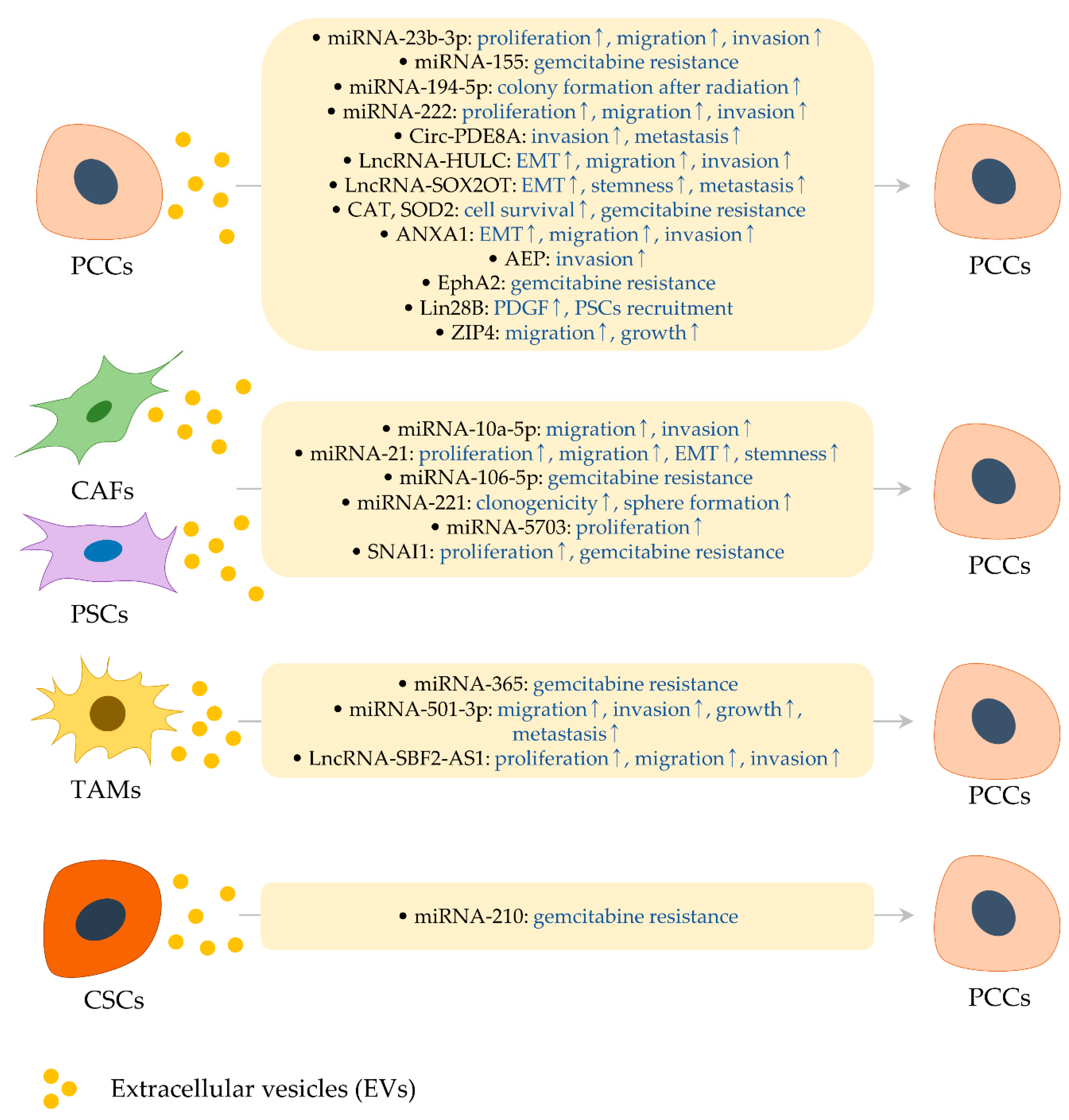
2. Effects of EVs and Their Cargo Molecules on Pancreatic Cancer
2.1. RNA Cargo in PCC-Derived EVs
2.1.1. MiRNA-23b-3p and miRNA-222
2.1.2. MiRNA-155 and ROS Detoxification Genes
2.1.3. MiRNA-194-5p
2.1.4. Circ-PDE8A
2.1.5. LncRNAs
2.2. Protein Cargo in PCC-Derived EVs
2.2.1. AEP
2.2.2. ANXA1
2.2.3. EphA2
2.2.4. Lin28B
2.2.5. ZIP4
2.3. RNA Cargo in EVs from PSCs and CAFs
2.3.1. MiRNA-10a-5p
2.3.2. MiRNA-21 and miRNA-221
2.3.3. MiRNA-106-5p
2.3.4. MiRNA-5703
2.3.5. SNAI1
2.4. RNA Cargo in EVs from TAMs and NKCs
2.4.1. MiRNA-365
2.4.2. MiRNA-501-3p
2.4.3. LncRNA-SBF2-AS1
2.5. RNA Cargo in EVs from CSCs
MiRNA-210

{kind=link}
{kind=link}
| Cargo | Source of EVs | Type of Study | Major Function of Cargo Molecules | Ref. |
|---|---|---|---|---|
| miRNAs | ||||
| miRNA-10a-5p | CAFs isolated from human pancreatic cancer tissues | In vitro | Support the aggressiveness of PANC-1 and SW1990 cells | [64] |
| miRNA-21 | PSCs (human PSC21-S/T cell line), CAFs (human CAF-19 cell line) | In vitro | Reinforce the proliferation, migration and EMT process of PANC-1 and SUIT-2 cells; augment clonogenicity and sphere formation of Colo-357 cells | [66,67] |
| miRNA-23b-3p | PCCs (human PANC-1 cells) | In vitro | Increase the proliferation, migration and invasion of PANC-1 cells | [11] |
| miRNA-106-5p | CAFs isolated from human pancreatic cancer tissues | In vitro | Confer gemcitabine resistance in AsPC-1 cells | [68] |
| miRNA-155 | PCCs (gemcitabine-treated human MIAPaCa-2 and Colo-357 cells, gemcitabine-resistant human PANC-1 cells) | In vitro, In vivo | Inhibit gemcitabine-induced apoptosis in MIAPaCa-2 and Colo-357 cells in vitro; confer gemcitabine resistance in PANC-1 cells in vivo | [15,16] |
| miRNA-194-5p | Irradiated human PANC-1 and SW1990 cells | In vitro, In vivo | Augment the survival of SW1990 cells following radiation in vitro | [20] |
| miRNA-210 | CSCs derived from gemcitabine-resistant human BxPC-3 cells | In vitro, In vivo | Inhibit gemcitabine-induced apoptosis in BxPC-3 and PANC-1 cells in vitro; confer gemcitabine resistance in BxPC-3 cells in vivo | [88] |
| miRNA-221 | CAFs (human CAF-19 cell line), PSCs isolated from human pancreatic cancer tissues | In vitro | Stimulate the clonogenicity and sphere formation of Colo-357 cells | [66] |
| miRNA-222 | PCCs (human Hs 766 T-L3 cells) | In vitro, In vivo | Enhance the proliferation, migration and invasion of CAPAN-1 and Hs 766 T-L3 cells in vitro; promote cancer progression in vivo | [12] |
| miRNA-365 | TAMs (M2-polarized murine peritoneal macrophages) | In vitro, In vivo | Attenuate the gemcitabine efficacy in K989 murine cells | [76] |
| miRNA-501-3p | TAMs (M2-polarized human THP-1 cells) | In vitro, In vivo | Enhance the migration and invasion of PANC-1 and BxPC-3 cells in vitro; promote cancer growth and metastasis in vivo | [80] |
| miRNA-5703 | PSCs isolated from human pancreatic cancer tissues | In vitro | Promote the proliferation of Patu8988 and T3M4 cells | [73] |
| Other non-coding RNAs | ||||
| Circ-PDE8A | PCCs (human Hs 766 T-L2 cells) | In vitro, In vivo | Facilitate the invasion of BxPC-3 and CAPAN-1 cells in vitro; enhance liver metastasis in vivo | [24] |
| LncRNA-HULC | PCCs (human PANC-1 cells) | In vitro, In vivo | Trigger migration, invasion and EMT process in PANC-1 and MIAPaCa-2 cells in vitro; promote cancer progression in vivo | [30] |
| LncRNA-SBF2-AS1 | TAMs (M2-polarized human THP-1 cells) | In vitro, In vivo | Enhance the proliferation, migration and invasion of PANC-1 cells in vitro; force the tumorigenic ability of PANC-1 cells in vivo | [81] |
| LncRNA-SOX2OT | PCCs (human Hs 766 T and Hs 766 T-L2 cells) | In vitro, In vivo | Promote EMT and stemness in Hs 766 T cells in vitro; trigger EMT, stemness and metastasis in vivo | [33] |
| mRNAs | ||||
| CAT and SOD2 | PCCs (gemcitabine-treated human MIAPaCa-2 and Colo-357 cells) | In vitro | Protect cell death induced by ROS in gemcitabine-treated MIAPaCa-2 cells | [16] |
| SNAI1 | CAFs isolated from human pancreatic cancer tissues | In vitro | Promote the proliferation and gemcitabine resistance in AsPC-1 cells | [75] |
| Proteins | ||||
| AEP | PCCs (human BxPC-3 cells) | In vitro | Aggravate the invasion ability of BxPC-3 and AsPC-1 cells | [38] |
| ANXA1 | PCCs (human MIAPaCa-2 cells) | In vitro | Facilitate the EMT, migration and invasion in MIAPaCa-2 cells | [43] |
| EphA2 | PCCs (gemcitabine-resistant human PANC-1 cells) | In vitro | Develop gemcitabine resistance in MIAPaCa-2 and BxPC-3 cells | [48] |
| Lin28B | PCCs (human PANC-1 and MIAPaCa-2 cells) | In vitro, In vivo | Increase the levels of PDGF in PANC-1 and MIAPaCa-2 cells, ultimately enhancing PSCs recruitment to the metastatic site | [52] |
| ZIP4 | PCCs (hamster PC-1.0 cells) | In vitro, In vivo | Promote the proliferation and migration of PC-1.0 cells in vitro; enhance the growth of cancer in vivo | [59] |
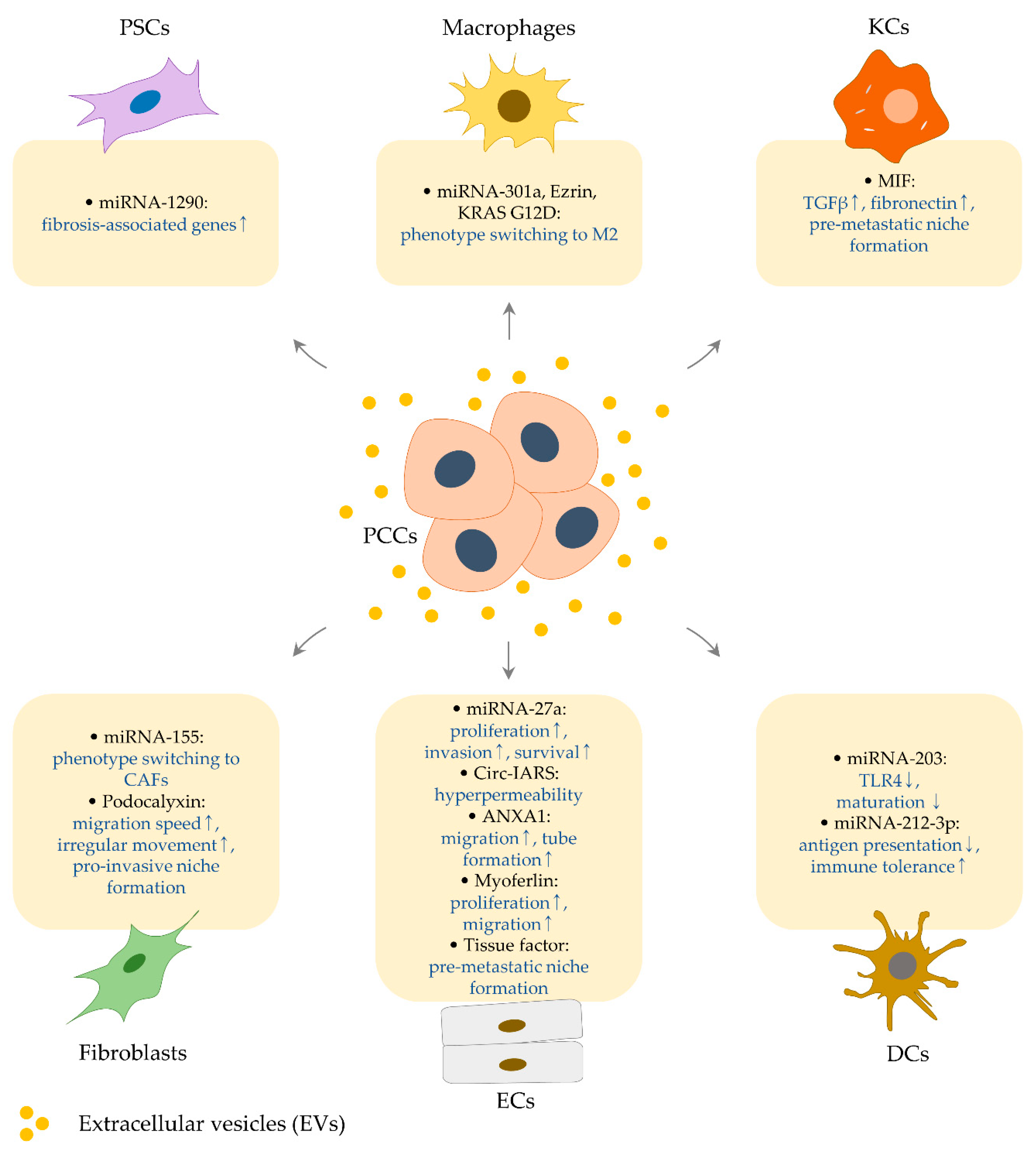
3. Effects of PCC-Derived EVs on the Cellular Components in Pancreatic Cancer Microenvironment and Metastatic Site
3.1. Regulation of Endothelial Cells by PCC-Derived EVs
3.1.1. MiRNA-27a
3.1.2. Circ-IARS
3.1.3. ANXA1
3.1.4. Myoferlin
3.1.5. Tissue Factor
3.2. Regulation of Fibroblasts and Stellate Cells by PCC-Derived EVs
3.2.1. MiRNA-155
3.2.2. Podocalyxin
3.2.3. MiRNA-1290
3.3. Effects of PCC-derived EVs on Kupffer Cells
MIF
3.4. Effects of PCC-Derived EVs on Macrophages
3.4.1. MiRNA-301a
3.4.2. Ezrin
3.4.3. KRAS G12D
3.5. Effects of PCC-Derived EVs on Dendritic Cells
3.5.1. MiRNA-203
3.5.2. MiRNA-212-3p
| Cargo | Source of EVs | Type of Study | Major Function of Cargo Molecules | Ref. |
|---|---|---|---|---|
| miRNAs | ||||
| miRNA-27a | PCCs (human PANC-1 cells) | In vitro, In vivo | Enhance the proliferation, invasion and survival of human endothelial cells in vitro; promote cancer growth and angiogenesis in vivo | [91] |
| miRNA-155 | PCCs (human BxPC-3 and SW1990 cells) | In vitro | Participate in the conversion from primary mouse fibroblasts to CAFs phenotypes | [110] |
| miRNA-203 | PCCs (human PANC-1 cells) | In vitro | Interrupt the maturation of human dendritic cells | [133] |
| miRNA-212-3p | PCCs (human PANC-1 cells) | In vitro | Repress the levels of MHC class II in human dendritic cells | [134] |
| miRNA-301a | Hypoxic PCCs (human PANC-1 cells) | In vitro, In vivo | Convert human bone marrow–derived macrophages into M2 types in vitro; facilitate lung metastasis in vivo | [125] |
| miRNA-1290 | PCCs (human PANC-1 cells) | In vitro | Activate human primary stellate cells; induce fibrogenic genes | [118] |
| Non-coding RNA | ||||
| Circ-IARS | PCCs (human Hs 766 T and Hs 766 T-L2 cells) | In vitro, In vivo | Disrupt the barrier integrity of human endothelial cells in vitro; promote invasion and metastasis in vivo | [97] |
| Proteins | ||||
| ANXA1 | PCCs (human MIAPaCa-2 cells) | In vitro | Mediate VEGF-induced migration and formation of the tube structure in human endothelial cells | [43] |
| Ezrin | PCCs (PC080 and PC084 cells derived from human pancreatic cancer tissues) | In vitro, In vivo | Promote M2 polarization of THP-1/U937-derived macrophages in vitro; facilitate liver metastasis along with a high M2/M1 ratio in vivo | [128] |
| KRAS G12D | Ferroptotic dying PCCs (human PANC-1 and AsPC-1 cells, primary PCCs from human pancreatic cancer tissues) | In vitro, In vivo | Promote M2 polarization of human mononuclear cell-derived macrophages in vitro; macrophage-mediated cancer growth is retarded by blocking KRAS G12D release and uptake in vivo | [130] |
| MIF | PCCs (murine PAN02 cells) | In vitro, In vivo | Activate human Kupffer cells in vitro; enhance the formation of liver pre-metastatic niche in vivo | [120] |
| Myoferlin | PCCs (human BxPC-3 cells) | In vitro | Enhance the proliferation and migration of human endothelial cells | [104] |
| Podocalyxin | PCCs isolated from human pancreatic cancer tissues | In vitro | Contribute to the generation of pro-invasive niche via regulating the migration of immortalized human dermal fibroblasts | [115] |
| Tissue Factor | PCCs (human BxPC-3 and CAPAN-1 cells) | In vitro | Activate human endothelial cells by upregulating E-selectin and IL-8 levels in a PAR-1 dependent manner | [109] |
4. Interference with EV-Based Conversation between Cells: Possibilities for Pancreatic Cancer Therapy
4.1. Cellular Factors Affecting EVs Biogenesis and Secretion
4.1.1. ANXA1
4.1.2. GIPC
4.1.3. PAFR
4.1.4. PAR2
4.1.5. RAB27
4.1.6. SMPD3
4.1.7. MiRNA-155 and lncRNA-PVT1
4.1.8. Other Possible Factors and Their Inhibitors
4.2. Regulation of EVs Uptake
4.2.1. AGER
4.2.2. ANXA6
4.2.3. TSPAN8
4.2.4. Dynamin-Dependent Endocytosis
4.2.5. Macropinocytosis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khalaf, N.; El-Serag, H.B.; Abrams, H.R.; Thrift, A.P. Burden of Pancreatic Cancer: From Epidemiology to Practice. Clin. Gastroenterol. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Adamska, A.; Domenichini, A.; Falasca, M. Pancreatic Ductal Adenocarcinoma: Current and Evolving Therapies. Int. J. Mol. Sci. 2017, 18, 1338. [Google Scholar] [CrossRef] [PubMed]
- Weniger, M.; Honselmann, K.C.; Liss, A.S. The Extracellular Matrix and Pancreatic Cancer: A Complex Relationship. Cancers 2018, 10, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, M.; Zhu, J.; Wu, J.; Cuneo, K.C.; Lubman, D.M. Circulating Microvesicles from Pancreatic Cancer Accelerate the Migration and Proliferation of PANC-1 Cells. J. Proteome Res. 2018, 17, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yang, L.; Baddour, J.; Achreja, A.; Bernard, V.; Moss, T.; Marini, J.C.; Tudawe, T.; Seviour, E.G.; Lucas, F.A.S.; et al. Tumor microenvironment derived exosomes pleiotropically modulate cancer cell metabolism. eLife 2016, 5, 10250. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.; Tao, L.-Y.; Yuan, C.; Zhang, L.; Xiu, D. Serum Derived Exosomes From Pancreatic Cancer Patients Promoted Metastasis: An iTRAQ-Based Proteomic Analysis. OncoTargets Ther. 2019, 12, 9329–9339. [Google Scholar]
- Shen, T.; Huang, Z.; Shi, C.; Pu, X.; Xu, X.; Wu, Z.; Ding, G.; Cao, L. Pancreatic cancer-derived exosomes induce apoptosis of T lymphocytes through the p38 MAPK-mediated endoplasmic reticulum stress. FASEB J. 2020, 34, 8442–8458. [Google Scholar] [CrossRef]
- Javeed, N.; Gustafson, M.P.; Dutta, S.K.; Lin, Y.; Bamlet, W.R.; Oberg, A.L.; Petersen, G.M.; Chari, S.T.; Dietz, A.B.; Mukhopadhyay, D. Immunosuppressive cd14(+)hla-dr(lo/neg) monocytes are elevated in pancreatic cancer and “primed” by tumor-derived exosomes. Oncoimmunology 2017, 6, e1252013. [Google Scholar] [CrossRef] [Green Version]
- Chiba, M.; Kubota, S.; Sato, K.; Monzen, S. Exosomes released from pancreatic cancer cells enhance angiogenic activities via dynamin-dependent endocytosis in endothelial cells in vitro. Sci. Rep. 2018, 8, 11972. [Google Scholar] [CrossRef] [Green Version]
- Catalano, M.; O’Driscoll, L.R. Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. J. Extracell. Vesicles 2019, 9, 1703244. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wu, X.; Xia, M.; Wu, F.; Ding, J.; Jiao, Y.; Zhan, Q.; An, F. Upregulated exosomic miR-23b-3p plays regulatory roles in the progression of pancreatic cancer. Oncol. Rep. 2017, 38, 2182–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Tao, Y.; Wang, X.; Jiang, P.; Li, J.; Peng, M.; Zhang, X.; Chen, K.; Liu, H.; Zhen, P.; et al. Tumor-Secreted Exosomal miR-222 Promotes Tumor Progression via Regulating P27 Expression and Re-Localization in Pancreatic Cancer. Cell. Physiol. Biochem. 2018, 51, 610–629. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Kim, H.J.; Chun, Y.-J.; Kim, M.Y. Ceramide Produces Apoptosis Through Induction of p27kip1 by Protein Phosphatase 2A-dependent Akt Dephosphorylation in PC-3 Prostate Cancer Cells. J. Toxicol. Environ. Heal. Part A 2010, 73, 1465–1476. [Google Scholar] [CrossRef]
- Cristóbal, I.; González-Alonso, P.; Daoud, L.; Solano, E.; Torrejón, B.; Manso, R.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Activation of the Tumor Suppressor PP2A Emerges as a Potential Therapeutic Strategy for Treating Prostate Cancer. Mar. Drugs 2015, 13, 3276–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikamori, M.; Yamada, D.; Eguchi, H.; Hasegawa, S.; Kishimoto, T.; Tomimaru, Y.; Asaoka, T.; Noda, T.; Wada, H.; Kawamoto, K.; et al. MicroRNA-155 Controls Exosome Synthesis and Promotes Gemcitabine Resistance in Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2017, 7, 42339. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.K.; Khan, M.A.; Bhardwaj, A.; Srivastava, S.K.; Zubair, H.; Patton, M.C.; Singh, S.; Khushman, M.; Singh, A.P. Exosomes confer chemoresistance to pancreatic cancer cells by promoting ROS detoxification and miR-155-mediated suppression of key gemcitabine-metabolising enzyme, DCK. Br. J. Cancer 2017, 116, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Shahbazi, J.; Lock, R.; Liu, T. Tumor Protein 53-Induced Nuclear Protein 1 Enhances p53 Function and Represses Tumorigenesis. Front. Genet. 2013, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Amrutkar, M.; Gladhaug, I.P. Pancreatic Cancer Chemoresistance to Gemcitabine. Cancers 2017, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Saiki, Y.; Yoshino, Y.; Fujimura, H.; Manabe, T.; Kudo, Y.; Shimada, M.; Mano, N.; Nakano, T.; Lee, Y.; Shimizu, S.; et al. DCK is frequently inactivated in acquired gemcitabine-resistant human cancer cells. Biochem. Biophys. Res. Commun. 2012, 421, 98–104. [Google Scholar] [CrossRef]
- Jiang, M.-J.; Chen, Y.-Y.; Dai, J.-J.; Gu, D.-N.; Mei, Z.; Liu, F.-R.; Huang, Q.; Tian, L. Dying tumor cell-derived exosomal miR-194-5p potentiates survival and repopulation of tumor repopulating cells upon radiotherapy in pancreatic cancer. Mol. Cancer 2020, 19, 68. [Google Scholar] [CrossRef]
- Yuan, X.; Yuan, Y.; He, Z.; Li, D.; Zeng, B.; Ni, Q.; Yang, M.Y.; Yang, D. The Regulatory Functions of Circular RNAs in Digestive System Cancers. Cancers 2020, 12, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Shi, Y.; Zhang, Y.; Sun, J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. OncoTargets Ther. 2017, 10, 5783–5794. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Cai, H.; Zhang, Y.; Liu, S.; Duan, Y.-F.; Sun, N.; Chen, X.; He, X. circZMYM2 Competed Endogenously with miR-335-5p to Regulate JMJD2C in Pancreatic Cancer. Cell. Physiol. Biochem. 2018, 51, 2224–2236. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yanfang, W.; Li, J.; Jiang, P.; Peng, T.; Chen, K.; Zhao, X.; Zhang, Y.; Zhen, P.; Zhu, J.; et al. Tumor-Released exosomal circular rna pde8a promotes invasive growth via the mir-338/macc1/met pathway in pancreatic cancer. Cancer Lett. 2018, 432, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.A.; Moeng, S.; Sim, S.; Kuh, H.-J.; Choi, S.Y.; Park, J.-K. MicroRNA-Based Combinatorial Cancer Therapy: Effects of MicroRNAs on the Efficacy of Anti-Cancer Therapies. Cells 2019, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Boil. 2017, 18, 206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wu, D.-D.; Sang, X.-B.; Wang, L.-L.; Zong, Z.-H.; Sun, K.-X.; Liu, B.-L.; Zhao, Y. The lncRNA HULC functions as an oncogene by targeting ATG7 and ITGB1 in epithelial ovarian carcinoma. Cell Death Dis. 2017, 8, e3118. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Wei, M.-H.; Lu, J.-G.; Bi, C.-Y. Long non-coding RNA HULC interacts with miR-613 to regulate colon cancer growth and metastasis through targeting RTKN. Biomed. Pharmacother. 2019, 109, 2035–2042. [Google Scholar] [CrossRef]
- Takahashi, K.; Ota, Y.; Kogure, T.; Suzuki, Y.; Iwamoto, H.; Yamakita, K.; Kitano, Y.; Fujii, S.; Haneda, M.; Patel, T.; et al. Circulating extracellular vesicle-encapsulated hulc is a potential biomarker for human pancreatic cancer. Cancer Sci. 2020, 111, 98–111. [Google Scholar] [CrossRef] [Green Version]
- Vaddi, P.K.; Stamnes, M.A.; Cao, H.; Chen, S. Elimination of sox2/oct4-associated prostate cancer stem cells blocks tumor development and enhances therapeutic response. Cancers 2019, 11, 1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.; Chen, Z.; He, S.; Gong, Y.; He, A.; Li, Y.; Zhang, L.; Zhang, X.; Fang, D.; Li, X.; et al. Long non-coding RNA SOX2OT promotes the stemness phenotype of bladder cancer cells by modulating SOX2. Mol. Cancer 2020, 19, 25. [Google Scholar]
- Li, Z.; Jiang, P.; Li, J.; Peng, M.; Zhao, X.; Zhang, X.; Chen, K.; Zhang, Y.; Liu, H.; Gan, L.; et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma. Oncogene 2018, 37, 3822–3838. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, J.; Wang, C.; Chi, Y.; Wei, Q.; Fu, Z.; Lian, C.; Huang, Q.; Liao, C.; Yang, Z.; et al. LncRNA SOX2OT promotes temozolomide resistance by elevating SOX2 expression via ALKBH5-mediated epigenetic regulation in glioblastoma. Cell Death Dis. 2020, 11, 384. [Google Scholar] [CrossRef]
- Guo, P.; Zhu, Z.; Sun, Z.; Wang, Z.; Zheng, X.; Xu, H. Expression of Legumain Correlates with Prognosis and Metastasis in Gastric Carcinoma. PLoS ONE 2013, 8, e73090. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, Y.; Li, H.; Li, Q.; Yu, Y.; Xu, X.; Xu, B.; Liu, T. Asparaginyl endopeptidase promotes the invasion and metastasis of gastric cancer through modulating epithelial-to-mesenchymal transition and analysis of their phosphorylation signaling pathways. Oncotarget 2016, 7, 34356–34370. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Z.; Zhang, G.; Zhu, Q.; Zeng, H.; Wang, T.; Gao, F.; Qi, Z.; Zhang, J.; Wang, R. Overexpression of asparaginyl endopeptidase is significant for esophageal carcinoma metastasis and predicts poor patient prognosis. Oncol. Lett. 2017, 15, 1229–1235. [Google Scholar] [CrossRef]
- Yan, Q.; Yuan, W.-B.; Sun, X.; Zhang, M.-J.; Cen, F.; Zhou, S.-Y.; Wu, W.-B.; Xu, Y.; Tong, L.-H.; Ma, Z.-H. Asparaginyl endopeptidase enhances pancreatic ductal adenocarcinoma cell invasion in an exosome-dependent manner and correlates with poor prognosis. Int. J. Oncol. 2018, 52, 1651–1660. [Google Scholar] [CrossRef] [Green Version]
- Edgington-Mitchell, L.E.; Wartmann, T.; Fleming, A.K.; Gocheva, V.; Van Der Linden, W.A.; Withana, N.P.; Verdoes, M.; Aurelio, L.; Edgington-Mitchell, D.; Lieu, T.; et al. Legumain is activated in macrophages during pancreatitis. Am. J. Physiol. Liver Physiol. 2016, 311. [Google Scholar] [CrossRef] [Green Version]
- Mahadevan, D.; Von Hoff, D.D. Tumor-Stroma interactions in pancreatic ductal adenocarcinoma. Mol. Cancer Ther. 2007, 6, 1186–1197. [Google Scholar] [CrossRef] [Green Version]
- Belvedere, R.; Bizzarro, V.; Popolo, A.; Piaz, F.D.; Vasaturo, M.; Picardi, P.; Parente, L.; Petrella, A. Role of intracellular and extracellular annexin A1 in migration and invasion of human pancreatic carcinoma cells. BMC Cancer 2014, 14, 961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bena, S.; Brancaleone, V.; Wang, J.M.; Perretti, M.; Flower, R.J. Annexin a1 interaction with the fpr2/alx receptor: Identification of distinct domains and downstream associated signaling. J. Biol. Chem. 2012, 287, 24690–24697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessolano, E.; Belvedere, R.; Bizzarro, V.; Franco, P.; De Marco, I.; Porta, A.; Tosco, A.; Parente, L.; Perretti, M.; Petrella, A. Annexin A1 May Induce Pancreatic Cancer Progression as a Key Player of Extracellular Vesicles Effects as Evidenced in the In Vitro MIA PaCa-2 Model System. Int. J. Mol. Sci. 2018, 19, 3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.-H.; Yong, H.-M.; Zhuang, Q.-X.; Zhang, X.-P.; Hou, P.-F.; Chen, Y.-S.; Zhu, M.-H.; Bai, J. Reduced expression of annexin A1 promotes gemcitabine and 5-fluorouracil drug resistance of human pancreatic cancer. Investig. New Drugs 2019, 38, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Martini, G.; Cardone, C.; Vitiello, P.P.; Belli, V.; Napolitano, S.; Troiani, T.; Ciardiello, D.; Della Corte, C.M.; Morgillo, F.; Matrone, N.; et al. EPHA2 Is a Predictive Biomarker of Resistance and a Potential Therapeutic Target for Improving Antiepidermal Growth Factor Receptor Therapy in Colorectal Cancer. Mol. Cancer Ther. 2019, 18, 845–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, K.R.; Wang, S.; Tan, L.; Hastings, A.K.; Song, W.; Lovly, C.M.; Meador, C.B.; Ye, F.; Lu, P.; Balko, J.M.; et al. EPHA2 Blockade Overcomes Acquired Resistance to EGFR Kinase Inhibitors in Lung Cancer. Cancer Res. 2016, 76, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Miao, B.; Ji, Z.; Tan, L.; Taylor, M.; Zhang, J.; Choi, H.G.; Frederick, D.T.; Kumar, R.; Wargo, J.A.; Flaherty, K.T.; et al. EPHA2 is a mediator of vemurafenib resistance and a novel therapeutic target in melanoma. Cancer Discov. 2014, 5, 274–287. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Wei, Q.; Koay, E.J.; Liu, Y.; Ning, B.; Bernard, P.W.; Zhang, N.; Han, H.; Katz, M.H.; Zhao, Z.; et al. Chemoresistance Transmission via Exosome-Mediated EphA2 Transfer in Pancreatic Cancer. Theranostics 2018, 8, 5986–5994. [Google Scholar] [CrossRef]
- Markosyan, N.; Li, J.; Sun, Y.H.; Richman, L.P.; Lin, J.H.; Yan, F.; Quinones, L.; Sela, Y.; Yamazoe, T.; Gordon, N.; et al. Tumor cell-intrinsic EPHA2 suppresses anti-tumor immunity by regulating PTGS2 (COX-2). J. Clin. Investig. 2019, 130, 3594–3609. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Wang, D.; Yuan, Z.; Xue, X.; Zhang, Y.; An, Y.; Chen, J.; Tu, M.; Lu, Z.; Wei, J.; et al. Persistent activation of pancreatic stellate cells creates a microenvironment favorable for the malignant behavior of pancreatic ductal adenocarcinoma. Int. J. Cancer 2012, 132, 993–1003. [Google Scholar] [CrossRef]
- Xu, Z.; Vonlaufen, A.; Phillips, P.A.; Fiala-Beer, E.; Zhang, X.; Yang, L.; Biankin, A.V.; Goldstein, D.; Pirola, R.C.; Wilson, J.S.; et al. Role of Pancreatic Stellate Cells in Pancreatic Cancer Metastasis. Am. J. Pathol. 2010, 177, 2585–2596. [Google Scholar] [CrossRef]
- Zhang, Y.-F.; Zhou, Y.-Z.; Zhang, B.; Huang, S.-F.; Li, P.-P.; He, X.-M.; Cao, G.-D.; Kang, M.-X.; Dong, X.; Wu, Y. Pancreatic cancer-derived exosomes promoted pancreatic stellate cells recruitment by pancreatic cancer. J. Cancer 2019, 10, 4397–4407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The lin28/let-7 pathway in cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.M.; Großhans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F. RAS Is Regulated by the let-7 MicroRNA Family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, I.-C.; Ustiyan, V.; Zhang, Y.; Cai, Y.; Kalin, T.V.; Kalinichenko, V.V. Abstract B18: FOXM1 transcription factor is required for the initiation of lung tumorigenesis by oncogenic KrasG12D. Models RAS Driv. Cancers 2014, 12, 5391–5396. [Google Scholar]
- Yu, G.; Zhou, A.; Xue, J.; Huang, C.; Zhang, X.; Kang, S.-H.; Chiu, W.-T.; Tan, C.; Xie, K.; Wang, J.; et al. FoxM1 promotes breast tumorigenesis by activating PDGF-A and forming a positive feedback loop with the PDGF/AKT signaling pathway. Oncotarget 2015, 6, 11281–11294. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhang, Y.; Yang, J.; Cui, X.; Zhou, Z.; Zhan, H.; Ding, K.; Tian, X.; Yang, Z.; Fung, K.A.; et al. Zip4 increases expression of transcription factor zeb1 to promote integrin alpha3beta1 signaling and inhibit expression of the gemcitabine transporter ent1 in pancreatic cancer cells. Gastroenterology 2020, 158, 679–692, e671. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Cui, X.; Chen, Y.; Zhu, V.F.; Hagan, J.P.; Wang, H.; Yu, X.-J.; Hodges, S.E.; Fang, J.; et al. A novel epigenetic CREB-miR-373 axis mediates ZIP4-induced pancreatic cancer growth. EMBO Mol. Med. 2013, 5, 1322–1334. [Google Scholar] [CrossRef]
- Jin, H.; Liu, P.; Wu, Y.; Meng, X.; Wu, M.; Han, J.; Tan, X. Exosomal zinc transporter ZIP4 promotes cancer growth and is a novel diagnostic biomarker for pancreatic cancer. Cancer Sci. 2018, 109, 2946–2956. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Huang, H.; Feng, M.; Yang, G.; Zheng, S.; You, L.; Zheng, L.; Hu, Y.; Zhang, T.-P.; Zhao, Y. MiR-10a-5p targets TFAP2C to promote gemcitabine resistance in pancreatic ductal adenocarcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 76. [Google Scholar] [CrossRef]
- Yu, T.; Liu, L.; Li, J.; Yan, M.; Lin, H.; Liu, Y.; Chu, D.; Tu, H.; Gu, A.; Yao, M. MiRNA-10a is upregulated in NSCLC and may promote cancer by targeting PTEN. Oncotarget 2015, 6, 30239–30250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, M.-J.; Wu, F.-X.; Li, P.; Liu, M.; Li, X.; Tang, H. MicroRNA-10a targets CHL1 and promotes cell growth, migration and invasion in human cervical cancer cells. Cancer Lett. 2012, 324, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Bloomston, M.; Frankel, W.L.; Petrocca, F.; Volinia, S.; Alder, H.; Hagan, J.P.; Liu, C.-G.; Bhatt, D.; Taccioli, C.; Croce, C.M. MicroRNA Expression Patterns to Differentiate Pancreatic Adenocarcinoma From Normal Pancreas and Chronic Pancreatitis. JAMA 2007, 297, 1901–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Li, L.; Wang, G.; Deng, X.; Li, Z.; Kong, X. VDR signaling inhibits cancer-associated-fibroblasts’ release of exosomal miR-10a-5p and limits their supportive effects on pancreatic cancer cells. Gut 2018, 68, 950–951. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M.H.; Yu, R.T.; Engle, D.D.; Ding, N.; Atkins, A.R.; Tiriac, H.; Collisson, E.A.; Connor, F.; Van Dyke, T.; Kozlov, S.; et al. Vitamin D receptor-mediated stromal reprogramming suppresses pancreatitis and enhances pancreatic cancer therapy. Cell 2014, 159, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Suresh, R.; Banerjee, S.; Bao, B.; Xu, Z.; Wilson, J.; A Philip, P.; Apte, M.; Sarkar, F. Contribution of microRNAs in understanding the pancreatic tumor microenvironment involving cancer associated stellate and fibroblast cells. Am. J. Cancer Res. 2015, 5, 1251–1264. [Google Scholar]
- Takikawa, T.; Masamune, A.; Yoshida, N.; Hamada, S.; Kogure, T.; Shimosegawa, T. Exosomes Derived From Pancreatic Stellate Cells. Pancreas 2017, 46, 19–27. [Google Scholar] [CrossRef]
- Fang, Y.; Zhou, W.; Rong, Y.; Kuang, T.; Xu, X.; Wu, W.; Wang, D.; Lou, W. Exosomal mirna-106b from cancer-associated fibroblast promotes gemcitabine resistance in pancreatic cancer. Exp. Cell Res. 2019, 383, 111543. [Google Scholar] [CrossRef]
- Jiao, D.; Yan, Y.; Shui, S.; Wu, G.; Ren, J.; Wang, Y.; Han, X. miR-106b regulates the 5-fluorouracil resistance by targeting Zbtb7a in cholangiocarcinoma. Oncotarget 2017, 8, 52913–52922. [Google Scholar] [CrossRef] [Green Version]
- Plate, M.; Li, T.; Wang, Y.; Mo, X.; Zhang, Y.; Ma, D.; Han, W. Identification and characterization of cmtm4, a novel gene with inhibitory effects on hela cell growth through inducing g2/m phase accumulation. Mol. Cells 2010, 29, 355–361. [Google Scholar] [CrossRef]
- Xue, H.; Li, T.; Wang, P.; Mo, X.; Zhang, H.; Ding, S.; Ma, D.; Lv, W.; Zhang, J.; Han, W. CMTM4 inhibits cell proliferation and migration via AKT, ERK1/2, and STAT3 pathway in colorectal cancer. Acta Biochim. et Biophys. Sin. 2019, 51, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Cheng, Y.; Wang, P.; Wang, W.; Hu, F.; Mo, X.; Lv, H.; Xu, T.; Han, W. CMTM4 is frequently downregulated and functions as a tumour suppressor in clear cell renal cell carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Guo, H.; Wang, Q.; Chen, K.; Marko, K.; Tian, X.; Yang, Y.-M. Pancreatic stellate cells derived exosomal miR-5703 promotes pancreatic cancer by downregulating CMTM4 and activating PI3K/Akt pathway. Cancer Lett. 2020, 490, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Tomono, T.; Yano, K.; Ogihara, T. Snail-Induced Epithelial-to-Mesenchymal Transition Enhances P-gp-Mediated Multidrug Resistance in HCC827 Cells. J. Pharm. Sci. 2017, 106, 2642–2649. [Google Scholar] [CrossRef] [Green Version]
- Richards, K.E.; Zeleniak, A.E.; Fishel, M.L.; Wu, J.; Littlepage, L.E.; Hill, R. Cancer-associated fibroblast exosomes regulate survival and proliferation of pancreatic cancer cells. Oncogene 2016, 36, 1770–1778. [Google Scholar] [CrossRef] [Green Version]
- Binenbaum, Y.; Fridman, E.; A Yaari, Z.; Milman, N.; Schroeder, A.; Ben David, G.; Shlomi, T.; Gil, Z. Transfer of miRNA in Macrophage-Derived Exosomes Induces Drug Resistance in Pancreatic Adenocarcinoma. Cancer Res. 2018, 78, 5287–5299. [Google Scholar] [CrossRef] [Green Version]
- Gordon, K.J.; Dong, M.; Chislock, E.M.; Fields, T.A.; Blobe, G.C. Loss of type iii transforming growth factor beta receptor expression increases motility and invasiveness associated with epithelial to mesenchymal transition during pancreatic cancer progression. Carcinogenesis 2008, 29, 252–262. [Google Scholar] [CrossRef]
- Fang, W.-Y.; Kuo, Y.-Z.; Chang, J.-Y.; Hsiao, J.-R.; Kao, H.-Y.; Tsai, S.-T.; Wu, L.-W. The Tumor Suppressor TGFBR3 Blocks Lymph Node Metastasis in Head and Neck Cancer. Cancers 2020, 12, 1375. [Google Scholar] [CrossRef]
- Mythreye, K.; Blobe, G.C. The type iii tgf-beta receptor regulates epithelial and cancer cell migration through beta-arrestin2-mediated activation of cdc42. Proc. Natl. Acad. Sci. USA 2009, 106, 8221–8226. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Ma, T.; Huang, B.; Lin, L.; Zhou, Y.; Yan, J.; Zou, Y.; Chen, S. Macrophage-Derived exosomal microrna-501-3p promotes progression of pancreatic ductal adenocarcinoma through the tgfbr3-mediated tgf-beta signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 310. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Zhou, Y.; Ma, T.; Chen, S.; Shi, N.; Zou, Y.; Hou, B.; Zhang, C. Down-regulated lncRNA SBF2-AS1 in M2 macrophage-derived exosomes elevates miR-122-5p to restrict XIAP, thereby limiting pancreatic cancer development. J. Cell. Mol. Med. 2020, 24, 5028–5038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.; Zhang, Y.; Xu, Z.; Jin, M. MicroRNA-122-5p inhibits cell proliferation, migration and invasion by targeting CCNG1 in pancreatic ductal adenocarcinoma. Cancer Cell Int. 2020, 20, 98. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.-Q.; Zhu, Y.-D.; Xie, G.-Q.; Zhang, K.; Sheng, J.; Zhu, Z.-F.; Ning, Z.-Y.; Chen, H.; Chen, Z.; Meng, Z.-Q.; et al. Long non-coding SBF2-AS1 acting as a competing endogenous RNA to sponge microRNA-142-3p to participate in gemcitabine resistance in pancreatic cancer via upregulating TWF1. Aging 2019, 11, 8860–8878. [Google Scholar] [CrossRef]
- Barbato, L.; Bocchetti, M.; Di Biase, A.; Regad, T. Cancer Stem Cells and Targeting Strategies. Cells 2019, 8, 926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Sun, H.; Provaznik, J.; Hackert, T.; Zöller, M. Pancreatic cancer-initiating cell exosome message transfer into noncancer-initiating cells: The importance of CD44v6 in reprogramming. J. Exp. Clin. Cancer Res. 2019, 38, 132. [Google Scholar] [CrossRef] [Green Version]
- Ullmann, P.; Qureshi-Baig, K.; Rodriguez, F.; Ginolhac, A.; Nonnenmacher, Y.; Ternes, D.; Weiler, J.; Gäbler, K.; Bahlawane, C.; Hiller, K.; et al. Hypoxia-responsive miR-210 promotes self-renewal capacity of colon tumor-initiating cells by repressing ISCU and by inducing lactate production. Oncotarget 2016, 7, 65454–65470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Yang, Z.; Zhu, Q.; Wu, Y.; Sun, K.; Alahdal, M.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. Up-regulation of miR-210 induced by a hypoxic microenvironment promotes breast cancer stem cell metastasis, proliferation, and self-renewal by targeting E-cadherin. FASEB J. 2018, 32, 6965–6981. [Google Scholar] [CrossRef]
- Yang, Z.; Zhao, N.; Cui, J.; Wu, H.; Xiong, J.; Peng, T. Exosomes derived from cancer stem cells of gemcitabine-resistant pancreatic cancer cells enhance drug resistance by delivering miR-210. Cell. Oncol. 2019, 43, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Urbich, C.; Kaluza, D.; Frömel, T.; Knau, A.; Bennewitz, K.; Boon, R.A.; Bonauer, A.; Doebele, C.; Boeckel, J.-N.; Hergenreider, E.; et al. MicroRNA-27a/b controls endothelial cell repulsion and angiogenesis by targeting semaphorin 6A. Blood 2012, 119, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Tong, J.-L.; Zhang, C.-P.; Xiao, Q.; Lin, X.-L.; Xiao, X.-Y. miR-27a induced by colon cancer cells in HLECs promotes lymphangiogenesis by targeting SMAD4. PLoS ONE 2017, 12, e0186718. [Google Scholar] [CrossRef] [Green Version]
- Shang, D.; Xie, C.; Hu, J.; Tan, J.; Yuan, Y.; Liu, Z.; Yang, Z. Pancreatic cancer cell–derived exosomal microRNA-27a promotes angiogenesis of human microvascular endothelial cells in pancreatic cancer via BTG2. J. Cell. Mol. Med. 2020, 24, 588–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Ryu, J.; Ryu, M.S.; Lim, S.; Han, K.O.; Lim, I.K.; Han, K.H. C-Reactive protein induces g2/m phase cell cycle arrest and apoptosis in monocytes through the upregulation of b-cell translocation gene 2 expression. FEBS Lett. 2014, 588, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Morel, A.-P.; Sentis, S.; Bianchin, C.; Le Romancer, M.; Jonard, L.; Rostan, M.-C.; Rimokh, R.; Corbo, L. BTG2 antiproliferative protein interacts with the human CCR4 complex existing in vivo in three cell-cycle-regulated forms. J. Cell Sci. 2003, 116, 2929–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, B.; Zhang, Z.; Wang, G. BTG2: A rising star of tumor suppressors (Review). Int. J. Oncol. 2014, 46, 459–464. [Google Scholar] [CrossRef]
- Tichet, M.; Prod’Homme, V.; Fenouille, N.; Ambrosetti, D.; Mallavialle, A.; Cerezo, M.; Ohanna, M.; Audebert, S.; Rocchi, S.; Giacchero, D.; et al. Tumour-derived SPARC drives vascular permeability and extravasation through endothelial VCAM1 signalling to promote metastasis. Nat. Commun. 2015, 6, 6993. [Google Scholar] [CrossRef] [Green Version]
- Monaghan-Benson, E.; Burridge, K. The Regulation of Vascular Endothelial Growth Factor-induced Microvascular Permeability Requires Rac and Reactive Oxygen Species. J. Boil. Chem. 2009, 284, 25602–25611. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, Z.; Jiang, P.; Peng, M.; Zhang, X.; Chen, K.; Liu, H.; Bi, H.; Liu, X.; Li, X. Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis. J. Exp. Clin. Cancer Res. 2018, 37, 177. [Google Scholar] [CrossRef] [Green Version]
- Pronk, M.C.; Van Bezu, J.S.M.; Amerongen, G.P.V.N.; Van Hinsbergh, V.W.M.; Hordijk, P.L. RhoA, RhoB and RhoC differentially regulate endothelial barrier function. Small GTPases 2017, 10, 466–484. [Google Scholar] [CrossRef] [Green Version]
- Pin, A.-L.; Houle, F.; Fournier, P.; Guillonneau, M.; Paquet, É.R.; Simard, M.J.; Royal, I.; Huot, J. Annexin-1-mediated Endothelial Cell Migration and Angiogenesis Are Regulated by Vascular Endothelial Growth Factor (VEGF)-induced Inhibition of miR-196a Expression *. J. Boil. Chem. 2012, 287, 30541–30551. [Google Scholar] [CrossRef] [Green Version]
- Côté, M.C.; Lavoie, J.R.; Houle, F.; Poirier, A.; Rousseau, S.; Huot, J. Regulation of Vascular Endothelial Growth Factor-induced Endothelial Cell Migration by LIM Kinase 1-mediated Phosphorylation of Annexin 1. J. Boil. Chem. 2010, 285, 8013–8021. [Google Scholar] [CrossRef] [Green Version]
- Rademaker, G.; Costanza, B.; Anania, S.; Agirman, F.; Maloujahmoum, N.; Di Valentin, E.; Goval, J.J.; Bellahcène, A.; Castronovo, V.; Peulen, O.; et al. Myoferlin Contributes to the Metastatic Phenotype of Pancreatic Cancer Cells by Enhancing Their Migratory Capacity through the Control of Oxidative Phosphorylation. Cancers 2019, 11, 853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rademaker, G.; Hennequière, V.; Brohée, L.; Nokin, M.-J.; Lovinfosse, P.; Durieux, F.; Gofflot, S.; Bellier, J.; Costanza, B.; Herfs, M.; et al. Myoferlin controls mitochondrial structure and activity in pancreatic ductal adenocarcinoma, and affects tumor aggressiveness. Oncogene 2018, 37, 4398–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernatchez, P.N.; Acevedo, L.; Fernández-Hernando, C.; Murata, T.; Chalouni, C.; Kim, J.; Erdjument-Bromage, H.; Shah, V.; Gratton, J.-P.; McNally, E.M.; et al. Myoferlin Regulates Vascular Endothelial Growth Factor Receptor-2 Stability and Function. J. Boil. Chem. 2007, 282, 30745–30753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blomme, A.; Fahmy, K.; Peulen, O.; Costanza, B.; Fontaine, M.; Struman, I.; Baiwir, D.; De Pauw, E.; Thiry, M.; Bellahcène, A.; et al. Myoferlin is a novel exosomal protein and functional regulator of cancer-derived exosomes. Oncotarget 2016, 7, 83669–83683. [Google Scholar] [CrossRef] [PubMed]
- Date, K.; Ettelaie, C.; Maraveyas, A. Tissue factor-bearing microparticles and inflammation: A potential mechanism for the development of venous thromboembolism in cancer. J. Thromb. Haemost. 2017, 15, 2289–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, Q.; Zhao, H.; Ma, L.; Meng, T.; Qian, J.; Jin, R.; Shen, J.; Yu, K. Pathological expression of tissue factor confers promising antitumor response to a novel therapeutic antibody SC1 in triple negative breast cancer and pancreatic adenocarcinoma. Oncotarget 2017, 8, 59086–59102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arce, M.; Pinto, M.P.; Galleguillos, M.; Muñoz, C.; Lange, S.; Ramirez, C.; Erices, R.; Gonzalez, P.; Velasquez, E.V.; Tempio, F.; et al. Coagulation Factor Xa Promotes Solid Tumor Growth, Experimental Metastasis and Endothelial Cell Activation. Cancers 2019, 11, 1103. [Google Scholar] [CrossRef] [Green Version]
- Soh, U.J.; Trejo, J. Activated protein c promotes protease-activated receptor-1 cytoprotective signaling through beta-arrestin and dishevelled-2 scaffolds. Proc. Natl. Acad. Sci. USA 2011, 108, E1372–E1380. [Google Scholar] [CrossRef] [Green Version]
- Che, S.P.Y.; Park, J.Y.; Stokol, T. Tissue Factor-Expressing Tumor-Derived Extracellular Vesicles Activate Quiescent Endothelial Cells via Protease-Activated Receptor-1. Front. Oncol. 2017, 7, 261. [Google Scholar] [CrossRef] [Green Version]
- Pang, W.; Su, J.; Wang, Y.; Feng, H.; Dai, X.; Yuan, Y.; Chen, X.; Yao, W. Pancreatic cancer-secreted miR-155 implicates in the conversion from normal fibroblasts to cancer-associated fibroblasts. Cancer Sci. 2015, 106, 1362–1369. [Google Scholar] [CrossRef]
- Vu, T.L.; Peng, B.; Zhang, D.X.; Ma, V.; Mathey-Andrews, C.A.; Lam, C.K.; Kiomourtzis, T.; Jin, J.; McReynolds, L.; Huang, L.; et al. Tumor-Secreted extracellular vesicles promote the activation of cancer-associated fibroblasts via the transfer of microRNA-125b. J. Extracell. Vesicles 2019, 8, 1599680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, J.P.; Timpson, P.; Karim, S.A.; Ridgway, R.A.; Athineos, D.; Doyle, B.; Jamieson, N.B.; Oien, K.A.; Lowy, A.M.; Brunton, V.G.; et al. Mutant p53 drives metastasis and overcomes growth arrest/senescence in pancreatic cancer. Proceeding Natl. Academy Sci. USA 2009, 107, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorini, C.; Cordani, M.; Padroni, C.; Blandino, G.; Di Agostino, S.; Donadelli, M. Mutant p53 stimulates chemoresistance of pancreatic adenocarcinoma cells to gemcitabine. Biochim. Biophys. Acta (BBA) Bioenerg. 2015, 1853, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wörmann, S.M.; Song, L.; Ai, J.; Diakopoulos, K.N.; Kurkowski, M.U.; Görgülü, K.; Ruess, D.A.; Campbell, A.; Doglioni, C.; Jodrell, D.; et al. Loss of P53 Function Activates JAK2–STAT3 Signaling to Promote Pancreatic Tumor Growth, Stroma Modification, and Gemcitabine Resistance in Mice and Is Associated With Patient Survival. Gastroenterology 2016, 151, 180–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novo, D.; Heath, N.; Mitchell, L.; Caligiuri, G.; Macfarlane, A.; Reijmer, D.; Charlton, L.; Knight, J.R.P.; Calka, M.; McGhee, E.; et al. Mutant p53s generate pro-invasive niches by influencing exosome podocalyxin levels. Nat. Commun. 2018, 9, 5069. [Google Scholar] [CrossRef] [Green Version]
- Bachem, M.G.; Schünemann, M.; Ramadani, M.; Siech, M.; Beger, H.; Buck, A.; Zhou, S.; Schmid-Kotsas, A.; Adler, G. Pancreatic carcinoma cells induce fibrosis by stimulating proliferation and matrix synthesis of stellate cells. Gastroenterology 2005, 128, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Habisch, H.; Zhou, S.; Siech, M.; Bachem, M.G. Interaction of Stellate Cells with Pancreatic Carcinoma Cells. Cancers 2010, 2, 1661–1682. [Google Scholar] [CrossRef] [Green Version]
- Masamune, A.; Yoshida, N.; Hamada, S.; Takikawa, T.; Nabeshima, T.; Shimosegawa, T. Exosomes derived from pancreatic cancer cells induce activation and profibrogenic activities in pancreatic stellate cells. Biochem. Biophys. Res. Commun. 2018, 495, 71–77. [Google Scholar] [CrossRef]
- Li, A.; Yu, J.; Kim, H.; Wolfgang, C.L.; Canto, M.I.; Hruban, R.H.; Goggins, M. MicroRNA array analysis finds elevated serum miR-1290 accurately distinguishes patients with low-stage pancreatic cancer from healthy and disease controls. Clin. Cancer Res. 2013, 19, 3600–3610. [Google Scholar] [CrossRef] [Green Version]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nature 2015, 17, 816–826. [Google Scholar] [CrossRef]
- Yang, S.; He, P.; Wang, J.; Schetter, A.; Tang, W.; Funamizu, N.; Yanaga, K.; Uwagawa, T.; Satoskar, A.R.; Gaedcke, J.; et al. A Novel MIF Signaling Pathway Drives the Malignant Character of Pancreatic Cancer by Targeting NR3C2. Cancer Res. 2016, 76, 3838–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patton, M.C.; Zubair, H.; Khan, M.A.; Singh, S.; Singh, A.P. Hypoxia alters the release and size distribution of extracellular vesicles in pancreatic cancer cells to support their adaptive survival. J. Cell. Biochem. 2019, 121, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.K.; Tewari, B.N.; Kanchan, R.K.; Baghel, K.S.; Nautiyal, N.; Shrivastava, R.; Kaur, H.; Bhatt, M.L.B.; Bhadauria, S. Macrophages are recruited to hypoxic tumor areas and acquire a Pro-Angiogenic M2-Polarized phenotype via hypoxic cancer cell derived cytokines Oncostatin M and Eotaxin. Oncotarget 2014, 5, 5350–5368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henze, A.-T.; Mazzone, M. The impact of hypoxia on tumor-associated macrophages. J. Clin. Investig. 2016, 126, 3672–3679. [Google Scholar] [CrossRef]
- Wang, X.; Luo, G.; Zhang, K.; Cao, J.; Huang, C.; Jiang, T.; Liu, B.; Su, L.; Qiu, Z. Hypoxic tumor-derived exosomal mir-301a mediates m2 macrophage polarization via pten/pi3kgamma to promote pancreatic cancer metastasis. Cancer. Res. 2018, 78, 4586–4598. [Google Scholar] [CrossRef] [Green Version]
- Quan, C.; Sun, J.; Lin, Z.; Jin, T.; Dong, B.; Meng, Z.; Chen, L.; Jin, T. Ezrin promotes pancreatic cancer cell proliferation and invasion through activating the Akt/mTOR pathway and inducing YAP translocation. Cancer Manag. Res. 2019, 11, 6553–6566. [Google Scholar] [CrossRef] [Green Version]
- Penchev, V.R.; Chang, Y.-T.; Begum, A.; Ewachiw, T.; Gocke, C.; Li, J.; McMillan, R.H.; Wang, Q.; Anders, R.; Marchionni, L.; et al. Ezrin Promotes Stem Cell Properties in Pancreatic Ductal Adenocarcinoma. Mol. Cancer Res. 2019, 17, 929–936. [Google Scholar] [CrossRef]
- Chang, Y.T.; Peng, H.Y.; Hu, C.M.; Huang, S.C.; Tien, S.C.; Jeng, Y.M. Pancreatic cancer-derived small extracellular vesical ezrin regulates macrophage polarization and promotes metastasis. Am. J. Cancer Res. 2020, 10, 12–37. [Google Scholar]
- A Prior, I.; Lewis, K.; Mattos, C. A comprehensive survey of Ras mutations in cancer. Cancer Res. 2012, 72, 2457–2467. [Google Scholar] [CrossRef] [Green Version]
- Dai, E.; Han, L.; Liu, J.; Xie, Y.; Kroemer, G.; Klionsky, D.J.; Zeh, H.J.; Kang, R.; Wang, J.; Tang, D. Autophagy-Dependent ferroptosis drives tumor-associated macrophage polarization via release and uptake of oncogenic KRAS protein. Autophagy 2020, 15. [Google Scholar] [CrossRef]
- Fang, H.; Ang, B.; Xu, X.; Huang, X.; Wu, Y.; Sun, Y.; Wang, W.; Li, N.; Cao, X.; Wan, T. TLR4 is essential for dendritic cell activation and anti-tumor T-cell response enhancement by DAMPs released from chemically stressed cancer cells. Cell. Mol. Immunol. 2013, 11, 150–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffell, B.; Chang-Strachan, D.; Chan, V.; Rosenbusch, A.; Ho, C.M.; Pryer, N.; Daniel, D.; Hwang, E.S.; Rugo, H.S.; Coussens, L.M. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell 2014, 26, 623–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Chen, J.; Zhou, L.; Chen, W.; Ding, G.; Cao, L. Pancreatic cancer derived exosomes regulate the expression of TLR4 in dendritic cells via miR-203. Cell. Immunol. 2014, 292, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Zhou, L.; Qian, Y.; Fu, M.; Chen, J.; Chen, J.; Xiang, J.; Wu, Z.; Jiang, G.; Cao, L. Pancreatic cancer-derived exosomes transfer miRNAs to dendritic cells and inhibit RFXAP expression via miR-212-3p. Oncotarget 2015, 6, 29877–29888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villard, J.; Lisowska-Grospierre, B.; Elsen, P.V.D.; Fischer, A.; Reith, W.; Mach, B. Mutation of RFXAP, a Regulator of MHC Class II Genes, in Primary MHC Class II Deficiency. N. Engl. J. Med. 1997, 337, 748–753. [Google Scholar] [CrossRef]
- Eden, E.R.; Sanchez-Heras, E.; Tsapara, A.; Sobota, A.; Levine, T.P.; Futter, C. Annexin A1 Tethers Membrane Contact Sites that Mediate ER to Endosome Cholesterol Transport. Dev. Cell 2016, 37, 473–483. [Google Scholar] [CrossRef] [Green Version]
- White, I.J.; Bailey, L.M.; Aghakhani, M.R.; E Moss, S.; Futter, C. EGF stimulates annexin 1-dependent inward vesiculation in a multivesicular endosome subpopulation. EMBO J. 2005, 25, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Babiychuk, E.B.; Monastyrskaya, K.; Draeger, A. Fluorescent Annexin A1 Reveals Dynamics of Ceramide Platforms in Living Cells. Traffic 2008, 9, 1757–1775. [Google Scholar] [CrossRef]
- Muders, M.H.; Dutta, S.K.; Wang, L.; Lau, J.S.; Bhattacharya, R.; Smyrk, T.C.; Chari, S.; Datta, K.; Mukhopadhyay, D. Expression and Regulatory Role of GAIP-Interacting Protein GIPC in Pancreatic Adenocarcinoma. Cancer Res. 2006, 66, 10264–10268. [Google Scholar] [CrossRef] [Green Version]
- Muders, M.H.; Vohra, P.K.; Dutta, S.K.; Wang, E.; Ikeda, Y.; Wang, L.; Udugamasooriya, D.G.; Memic, A.; Rupasinghe, C.N.; Rupashinghe, C.N.; et al. Targeting GIPC/synectin in pancreatic cancer inhibits tumor growth. Clin. Cancer Res. 2009, 15, 4095–4103. [Google Scholar] [CrossRef] [Green Version]
- Borchardt, H.; Schulz, A.; Datta, K.; Muders, M.H.; Aigner, A. Silencing of Neuropilins and GIPC1 in pancreatic ductal adenocarcinoma exerts multiple cellular and molecular antitumor effects. Sci. Rep. 2019, 9, 15471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Pal, K.; Sharma, A.K.; Dutta, S.K.; Lau, J.S.; Yan, I.K.; Wang, E.; Elkhanany, A.; Alkharfy, K.M.; Sanyal, A.; et al. GAIP Interacting Protein C-Terminus Regulates Autophagy and Exosome Biogenesis of Pancreatic Cancer through Metabolic Pathways. PLoS ONE 2014, 9, e114409. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, A.; Kadam, S.M.; Liu, L.; Kelly, L.E.; Rapp, C.M.; Chen, Y.; Sahu, R.P. Gemcitabine Induces Microvesicle Particle Release in a Platelet-Activating Factor-Receptor-Dependent Manner via Modulation of the MAPK Pathway in Pancreatic Cancer Cells. Int. J. Mol. Sci. 2018, 20, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Featherby, S.; Madkhali, Y.; Maraveyas, A.; Ettelaie, C. Apixaban Suppresses the Release of TF-Positive Microvesicles and Restrains Cancer Cell Proliferation through Directly Inhibiting TF-fVIIa Activity. Thromb. Haemost. 2019, 119, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Carmo, N.; Krumeich, S.; Fanget, I.; Raposo, G.; Savina, A.; Moita, C.F.; Schauer, K.; Hume, A.N.; Freitas, R.P.; et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nature 2009, 12, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ni, Q.; Wang, X.; Zhu, H.; Wang, Z.; Huang, J. High expression of RAB27A and TP53 in pancreatic cancer predicts poor survival. Med. Oncol. 2014, 32, 372. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Q.; Wang, X.; Zhu, H.; Zhang, S.; Wang, W.; Wang, Z.; Huang, J. Correlation Between RAB27B and p53 Expression and Overall Survival in Pancreatic Cancer. Pancreas 2016, 45, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jin, Q.; Huang, F.; Tang, Z.; Huang, J. Effects of Rab27A and Rab27B on Invasion, Proliferation, Apoptosis, and Chemoresistance in Human Pancreatic Cancer Cells. Pancreas 2017, 46, 1173–1179. [Google Scholar] [CrossRef]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brugger, B.; Simons, M. Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Guo, B.B.; Bellingham, S.A.; Hill, A.F. The Neutral Sphingomyelinase Pathway Regulates Packaging of the Prion Protein into Exosomes. J. Boil. Chem. 2014, 290, 3455–3467. [Google Scholar] [CrossRef] [Green Version]
- McAndrews, K.M.; Kalluri, R. Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer 2019, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Vuckovic, S.; VanDyke, K.; Rickards, D.A.; Winter, P.M.; Brown, S.; Mitchell, T.; Liu, J.; Lu, J.; Askenase, P.W.; Yuriev, E.; et al. The cationic small molecule GW4869 is cytotoxic to high phosphatidylserine-expressing myeloma cells. Br. J. Haematol. 2017, 177, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Luberto, C.; Hassler, D.F.; Signorelli, P.; Okamoto, Y.; Sawai, H.; Boros, E.; Hazen-Martin, D.J.; Obeid, L.M.; Hannun, Y.A.; Smith, G.K. Inhibition of Tumor Necrosis Factor-induced Cell Death in MCF7 by a Novel Inhibitor of Neutral Sphingomyelinase. J. Boil. Chem. 2002, 277, 41128–41139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Li, Y.; Sang, Y.; Yu, B.; Lv, D.; Zhang, W.; Feng, H. Lncrna pvt1 regulates triple-negative breast cancer through klf5/beta-catenin signaling. Oncogene 2018, 37, 4723–4734. [Google Scholar] [CrossRef]
- Chen, J.; Yu, Y.; Li, H.; Hu, Q.; Chen, X.; He, Y.; Xue, C.; Ren, F.; Ren, Z.; Li, J.; et al. Long non-coding RNA pvt1 promotes tumor progression by regulating the mir-143/hk2 axis in gallbladder cancer. Mol. Cancer 2019, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Du, P.; Cui, P.; Qin, Y.; Hu, C.; Wu, J.; Zhou, Z.; Zhang, W.; Qin, L.; Huang, G. Lncrna pvt1 promotes angiogenesis via activating the stat3/vegfa axis in gastric cancer. Oncogene 2018, 37, 4094–4109. [Google Scholar] [CrossRef]
- Du, P.; Hu, C.; Qin, Y.; Zhao, J.; Patel, R.; Fu, Y.; Zhu, M.; Zhang, W.; Huang, G. LncRNA PVT1 Mediates Antiapoptosis and 5-Fluorouracil Resistance via Increasing Bcl2 Expression in Gastric Cancer. J. Oncol. 2019, 2019, 9325407. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Chen, W.; Peng, J.; Li, Y.; Zhuang, Y.; Zhu, Z.; Shao, C.-K.; Yang, W.; Yao, H.-R.; Zhang, S. LncRNA PVT1 triggers Cyto-protective autophagy and promotes pancreatic ductal adenocarcinoma development via the miR-20a-5p/ULK1 Axis. Mol. Cancer 2018, 17, 98. [Google Scholar] [CrossRef]
- Zhao, L.; Kong, H.; Sun, H.; Chen, Z.; Chen, B.; Zhou, M. Lncrna-Pvt1 promotes pancreatic cancer cells proliferation and migration through acting as a molecular sponge to regulate mir-448. J. Cell Phys. 2018, 233, 4044–4055. [Google Scholar] [CrossRef]
- Sun, C.; Wang, P.; Dong, W.; Liu, H.; Sun, J.; Zhao, L. LncRNA PVT1 promotes exosome secretion through YKT6, RAB7, and VAMP3 in pancreatic cancer. Aging 2020, 12, 10427–10440. [Google Scholar] [CrossRef]
- VanLandingham, P.A.; Ceresa, B.P. Rab7 Regulates Late Endocytic Trafficking Downstream of Multivesicular Body Biogenesis and Cargo Sequestration. J. Boil. Chem. 2009, 284, 12110–12124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, D.E.; Chia, J.; Jayawardena, K.; Antrobus, R.; Bard, F.; Peden, A.A. Vamp3/syb and ykt6 are required for the fusion of constitutive secretory carriers with the plasma membrane. PLoS Genet. 2017, 13, e1006698. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Kim, H.; Lal, M.; McGee, L.; Johnson, A.; Moustafa, A.A.; Jones, J.C.; Mondal, D.; Ferrer, M.; Abdel-Mageed, A.B. Manumycin a suppresses exosome biogenesis and secretion via targeted inhibition of ras/raf/erk1/2 signaling and hnrnp h1 in castration-resistant prostate cancer cells. Cancer Lett. 2017, 408, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Kim, H.; McGee, L.; Johnson, A.E.; Talwar, S.; Marugan, J.; Southall, N.; Hu, X.; Lal, M.; Mondal, D.; et al. High-Throughput screening identified selective inhibitors of exosome biogenesis and secretion: A drug repurposing strategy for advanced cancer. Sci. Rep. 2018, 8, 8161. [Google Scholar] [CrossRef] [Green Version]
- Kosgodage, U.S.; Trindade, R.P.; Thompson, P.R.; Inal, J.M.; Lange, S. Chloramidine/Bisindolylmaleimide-I-Mediated Inhibition of Exosome and Microvesicle Release and Enhanced Efficacy of Cancer Chemotherapy. Int. J. Mol. Sci. 2017, 18, 1007. [Google Scholar] [CrossRef]
- Kainuma, O.; Asano, T.; Hasegawa, M.; Kenmochi, T.; Nakagohri, T.; Tokoro, Y.; Isono, K. Inhibition of Growth and Invasive Activity of Human Pancreatic Cancer Cells by a Farnesyltransferase Inhibitor, Manumycin. Pancreas 1997, 15, 379–383. [Google Scholar] [CrossRef]
- Ding, N.; Cui, X.-X.; Gao, Z.; Huang, H.; Wei, X.; Du, Z.; Lin, Y.; Shih, W.J.; Rabson, A.B.; Conney, A.H.; et al. A triple combination of atorvastatin, celecoxib and tipifarnib strongly inhibits pancreatic cancer cells and xenograft pancreatic tumors. Int. J. Oncol. 2014, 44, 2139–2145. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Miyasaka, Y.; Ohuchida, K.; Okumura, T.; Zheng, B.; Torata, N.; Fujita, H.; Nabae, T.; Manabe, T.; Shimamoto, M.; et al. Calpain inhibitor calpeptin suppresses pancreatic cancer by disrupting cancer-stromal interactions in a mouse xenograft model. Cancer Sci. 2016, 107, 1443–1452. [Google Scholar] [CrossRef]
- Mu, G.; Ding, Q.; Li, H.; Zhang, L.; Zhang, L.; He, K.; Wu, L.; Deng, Y.; Yang, D.; Wu, L.; et al. Gastrin stimulates pancreatic cancer cell directional migration by activating the galpha12/13-rhoa-rock signaling pathway. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef]
- Rojas, A.; Araya, P.; Romero, J.; Delgado-López, F.; Gonzalez, I.; Añazco, C.; Perez-Castro, R. Skewed Signaling through the Receptor for Advanced Glycation End-Products Alters the Proinflammatory Profile of Tumor-Associated Macrophages. Cancer Microenviron. 2018, 11, 97–105. [Google Scholar] [CrossRef]
- Heo, K.; Lee, S. TSPAN8 as a Novel Emerging Therapeutic Target in Cancer for Monoclonal Antibody Therapy. Biomology 2020, 10, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khushman, M.; Patel, G.K.; Laurini, J.A.; Bhardwaj, A.; Roveda, K.; Donnell, R.; Sherling, K.; Case, B.; Frankel, A.E.; Pai, S.; et al. Exosomal markers (CD63 and CD9) expression and their prognostic significance using immunohistochemistry in patients with pancreatic ductal adenocarcinoma. J. Gastrointest. Oncol. 2019, 10, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Von Au, A.; Schnölzer, M.; Hackert, T.; Zöller, M. CD44v6-Competent tumor exosomes promote motility, invasion and cancer-initiating cell marker expression in pancreatic and colorectal cancer cells. Oncotarget 2016, 7, 55409–55436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, K.M.; Cao, H.; Chen, J.; Qiang, L.; Krueger, E.W.; Johnson, K.M.; Bamlet, W.R.; Zhang, L.; McNiven, M.A.; Razidlo, G.L. Dynamin 2 interacts with α-actinin 4 to drive tumor cell invasion. Mol. Boil. Cell 2020, 31, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Razidlo, G.L.; Wang, Y.; Chen, J.; Krueger, E.W.; Billadeau, D.D.; McNiven, M.A. Dynamin 2 Potentiates Invasive Migration of Pancreatic Tumor Cells through Stabilization of the Rac1 GEF Vav1. Dev. Cell 2013, 24, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Chiba, M.; Kubota, S.; Sakai, A.; Monzen, S. Cell-To-Cell communication via extracellular vesicles among human pancreatic cancer cells derived from the same patient. Mol. Med. Rep. 2018, 18, 3989–3996. [Google Scholar] [CrossRef] [Green Version]
- Koivusalo, M.; Welch, C.; Hayashi, H.; Scott, C.C.; Kim, M.; Alexander, T.; Touret, N.; Hahn, K.M.; Grinstein, S. Amiloride inhibits macropinocytosis by lowering submembranous pH and preventing Rac1 and Cdc42 signaling. J. Cell Boil. 2010, 188, 547–563. [Google Scholar] [CrossRef] [Green Version]
- Commisso, C.; Davidson, S.M.; Soydaner-Azeloglu, R.G.; Parker, S.J.; Kamphorst, J.J.; Hackett, S.; Grabocka, E.; Nofal, M.; Drebin, J.A.; Thompson, C.B.; et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 2013, 497, 633–637. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhang, J.; Zhang, L.; Zhu, Z.; Fan, J.; Chen, L.; Zhuang, L.; Luo, J.; Chen, H.; Liu, L.; et al. MicroRNA 23b Regulates Autophagy Associated With Radioresistance of Pancreatic Cancer Cells. Gastroenterology 2013, 145, 1133–1143. [Google Scholar] [CrossRef]
- Ceppi, M.; Pereira, P.M.; Dunand-Sauthier, I.; Barras, E.; Reith, W.; Santos, M.A.S.; Pierre, P. MicroRNA-155 modulates the interleukin-1 signaling pathway in activated human monocyte-derived dendritic cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2735–2740. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Liang, C.; Hua, J.; Zhang, B.; Liu, J.; Zhang, Y.; Wei, M.; Yu, X.; Xu, J.; Shi, S. A mir-146a-5p/traf6/nf-kb p65 axis regulates pancreatic cancer chemoresistance: Functional validation and clinical significance. Theranostics 2020, 10, 3967–3979. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Okada, R.; Nagao, K.; Kawamata, Y.; Hanyu, A.; Yoshimoto, S.; Takasugi, M.; Watanabe, S.; Kanemaki, M.T.; Obuse, C.; et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat. Commun. 2017, 8, 15287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan-Chari, V.; Kohan, H.G.; Asimakopoulos, A.G.; Sudha, T.; Sell, S.; Kannan, K.; Boroujerdi, M.; Davis, P.J.; Mousa, S. Microvesicle removal of anticancer drugs contributes to drug resistance in human pancreatic cancer cells. Oncotarget 2016, 7, 50365–50379. [Google Scholar] [CrossRef] [PubMed]
- Giallombardo, M.; Taverna, S.; Alessandro, R.; Hong, D.; Rolfo, C. Exosome-Mediated drug resistance in cancer: The near future is here. Ther. Adv. Med Oncol. 2016, 8, 320–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosgodage, U.S.; Mould, R.; Henley, A.B.; Nunn, A.V.; Guy, G.W.; Thomas, E.L.; Inal, J.M.; Bell, J.D.; Lange, S. Cannabidiol (CBD) Is a Novel Inhibitor for Exosome and Microvesicle (EMV) Release in Cancer. Front. Pharmacol. 2018, 9, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salimi, L.; Akbari, A.; Jabbari, N.; Mojarad, B.; Vahhabi, A.; Szafert, S.; Kalashani, S.A.; Soraya, H.; Nawaz, M.; Rezaie, J. Synergies in exosomes and autophagy pathways for cellular homeostasis and metastasis of tumor cells. Cell Biosci. 2020, 10, 1–18. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moeng, S.; Son, S.W.; Lee, J.S.; Lee, H.Y.; Kim, T.H.; Choi, S.Y.; Kuh, H.J.; Park, J.K. Extracellular Vesicles (EVs) and Pancreatic Cancer: From the Role of EVs to the Interference with EV-Mediated Reciprocal Communication. Biomedicines 2020, 8, 267. https://doi.org/10.3390/biomedicines8080267
Moeng S, Son SW, Lee JS, Lee HY, Kim TH, Choi SY, Kuh HJ, Park JK. Extracellular Vesicles (EVs) and Pancreatic Cancer: From the Role of EVs to the Interference with EV-Mediated Reciprocal Communication. Biomedicines. 2020; 8(8):267. https://doi.org/10.3390/biomedicines8080267
Chicago/Turabian StyleMoeng, Sokviseth, Seung Wan Son, Jong Sun Lee, Han Yeoung Lee, Tae Hee Kim, Soo Young Choi, Hyo Jeong Kuh, and Jong Kook Park. 2020. "Extracellular Vesicles (EVs) and Pancreatic Cancer: From the Role of EVs to the Interference with EV-Mediated Reciprocal Communication" Biomedicines 8, no. 8: 267. https://doi.org/10.3390/biomedicines8080267