The Role of GPR120 Receptor in Essential Fatty Acids Metabolism in Schizophrenia

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Blood Collection
2.3. Fatty Acids Assay
2.4. GPR120 Assay
2.5. Dietary Assessment
2.6. Sociodemographic and Clinical Data
2.7. The Severity of Schizophrenia Symptoms Assessment
2.8. Statistical Analysis
3. Results
3.1. Study Participant Characteristics
3.2. Nutritional Status and Metabolism of PUFAs
3.3. Effect of Nutritional Status on PUFA Metabolism
3.4. Effect of Demographic and Clinical Variables on PUFAs Nutritional Status
3.5. GPR120 Serum Concentration Variability
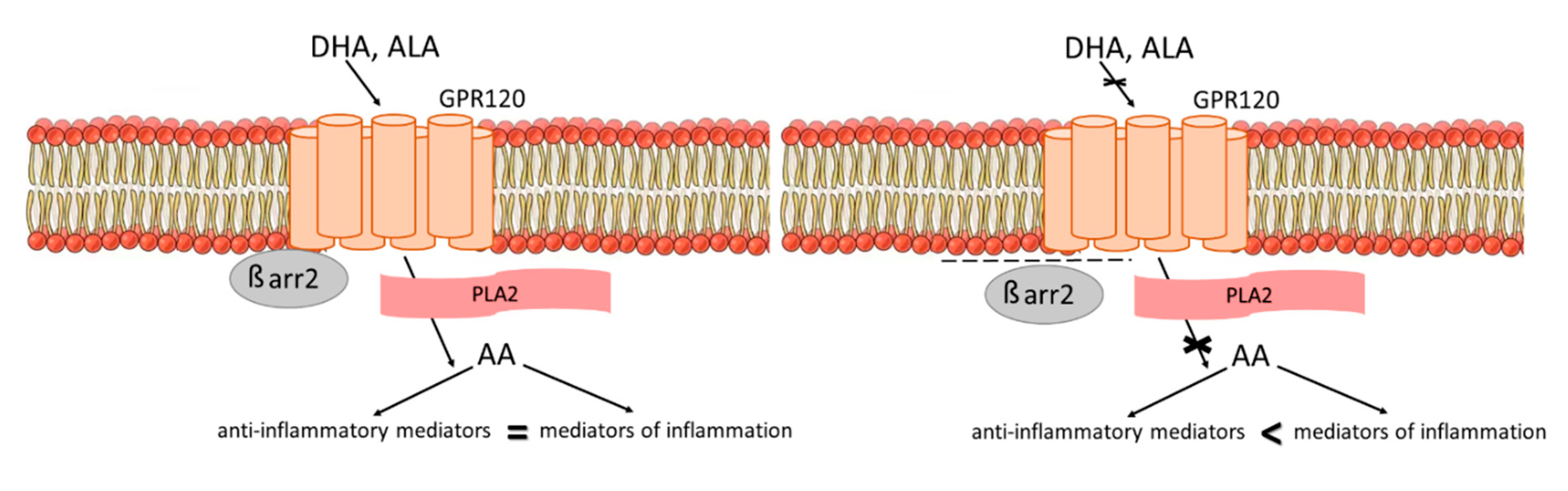
4. Discussion
5. Conclusions
6. Advantages and Limitations
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PUFAs | Polyunsaturated fatty acids |
| FAs | Fatty acids |
| DHA | Docosahexaenoic acid |
| DPA | Docosapentaenoic acid |
| EPA | Eicosapentaenoic acid |
| AA | Arachidonic acid |
| SZ | Schizophrenia |
| AMN | Aqueous methyl nicotinate |
| PLA2 | Phospholipase A2 |
| GPR120 | G-coupled receptor responsive to fatty acids |
| LA | Linolenic acid |
| ALA | α-Linolenic acid |
| HC | Healthy control |
| BMI | Body mass index |
| PANSS | Positive and Negative Symptom Scale |
| RDA | Recommended daily intake |
| SREBP1 | Sterol regulatory element-binding protein type 1 |
| DSM-5 | The Diagnostic and Statistical Manual of Mental Disorders |
| GS/MS | Gas chromatography–mass spectrometry |
References
- Clari, R.; McNamara, R.K.; Szeszko, P.R. Omega-3 Polyunsaturated Fatty Acids and Antioxidants for the Treatment of Schizophrenia: A Role for Magnetic Resonance Imaging. In Neuroimaging in Schizophrenia; Kubicki, M., Shenton, M.E., Eds.; Springer: Cham, Switzerland, 2020; pp. 367–383. [Google Scholar]
- McNamara, R.K.; Almeida, D.M. Omega-3 Polyunsaturated Fatty Acid Deficiency and Progressive Neuropathology in Psychiatric Disorders: A Review of Translational Evidence and Candidate Mechanisms. Harv. Rev. Psychiatry 2019, 27, 94–107. [Google Scholar] [CrossRef]
- Bozzatello, P.; Rocca, P.; Mantelli, E.; Bellino, S. Polyunsaturated Fatty Acids: What is Their Role in Treatment of Psychiatric Disorders? Int. J. Mol. Sci. 2019, 20, 5257. [Google Scholar] [CrossRef] [Green Version]
- Van der Kemp, W.J.M.; Klomp, D.W.J.; Kahn, R.S.; Luijten, P.R.; Hulshoff, H.E.P. A meta-analysis of the polyunsaturated fatty acid composition of erythrocyte membranes in schizophrenia. Schizophr. Res. 2012, 141, 153–161. [Google Scholar] [CrossRef]
- De Almeida, V.; Alexandrino, G.L.; Aquino, A.; Gomes, A.F.; Murgu, M.; Dobrowolny, H.; Guest, P.C.; Steiner, J.; Martins-de-Souza, D. Changes in the blood plasma lipidome associated with effective or poor response to atypical antipsychotic treatments in schizophrenia patients. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 101, 109945. [Google Scholar] [CrossRef]
- Messamore, E. The niacin response biomarker as a schizophrenia endophenotype: A status update. Prostaglandins Leukot. Essent. Fatty Acids 2018, 136, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Messamore, E.; Yao, J.K. Phospholipid, arachidonate and eicosanoid signaling in schizophrenia. OCL 2016, 23, D112. [Google Scholar] [CrossRef] [Green Version]
- Mouchlis, V.D.; Dennis, E.A. Phospholipase A2 catalysis and lipid mediator lipidomics. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2019, 1864, 766–771. [Google Scholar] [CrossRef]
- Tallima, H.; El Ridi, R. Arachidonic acid: Physiological roles and potential health benefits—A review. J. Adv. Res. 2018, 11, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Alvarez-Curto, E.; Hudson, B.D.; Prihandoko, R.; Tobin, A.B. FFA4/GPR120: Pharmacology and Therapeutic Opportunities. Trends Pharmacol. Sci. 2017, 38, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Im, D.-S. FFA4 (GPR120) as a fatty acid sensor involved in appetite control, insulin sensitivity and inflammation regulation. Mol. Asp. Med. 2018, 64, 92–108. [Google Scholar] [CrossRef]
- Karakuła-Juchnowicz, H.; Róg, J.; Juchnowicz, D.; Morylowska-Topolska, J. GPR120: Mechanism of action, role and potential for medical applications. Postepy Hig. Med. Dosw. 2017, 71, 942–953. [Google Scholar] [CrossRef]
- Dragano, N.R.V.; Solon, C.; Ramalho, A.F.; de Moura, R.F.; Razolli, D.S.; Christiansen, E.; Azevedo, C.; Ulven, T.; Velloso, L.A. Polyunsaturated fatty acid receptors, GPR40 and GPR120, are expressed in the hypothalamus and control energy homeostasis and inflammation. J. Neuroinflammation 2017, 14, 91. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Pub: Washington, DC, USA, 2013. [Google Scholar]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191. [Google Scholar] [CrossRef] [Green Version]
- Dams, S.; Holasek, S.; Tsiountsioura, M.; Edelsbrunner, M.; Dietz, P.; Koefeler, H.; Malliga, D.-E.; Gürbüz, A.; Meier-Allard, N.; Poncza, B.; et al. Effects of a plant-based fatty acid supplement and a powdered fruit, vegetable and berry juice concentrate on omega-3-indices and serum micronutrient concentrations in healthy subjects. Int. J. Food Sci. Nutr. 2020, 1–12. [Google Scholar] [CrossRef]
- Dung, N.T.; Van Chi, P. A Survey of Membrane Proteins in Human Serum. Proteom. Insights 2012, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Kunachowicz, H.; Nadolna, I.; Przygoda, B.; Iwanow, K. Tables of Composition and Nutritional Value; Food and Nutrition Institute: Warsaw, Poland, 2005; pp. 145–149. [Google Scholar]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Multiple Significance Tests: The Bonferroni Method|The BMJ. Available online: https://www.bmj.com/content/310/6973/170.short (accessed on 14 July 2020).
- Leucht, S.; Samara, M.; Heres, S.; Davis, J.M. Dose Equivalents for Antipsychotic Drugs: The DDD Method. Schizophr. Bull. 2016, 42, S90–S94. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, M.; Rychlik, E.; Stoś, K.; Wierzejska, R.; Wojtasik, A.; Charzewska, J.; Mojska, H.; Szponar, L.; Sajór, I.; Kłosiewicz-Latoszek, L.; et al. Normy żywienia dla populacji Polski; Instytut Żywności i Żywienia: Warszawa, Poland, 2017. [Google Scholar]
- Horrobin, D.F.; Glen, A.I.M.; Vaddadi, K. The membrane hypothesis of schizophrenia. Schizophr. Res. 1994, 13, 195–207. [Google Scholar] [CrossRef]
- The Procurement, Storage, and Quality Assurance of Frozen Blood and Tissue Biospecimens in Pathology, Biorepository, and Biobank Settings. Abstract—Europe PMC. Available online: https://europepmc.org/article/PMC/3982909 (accessed on 17 July 2020).
- B7-H4 Is a Novel Membrane-Bound Protein and a Candidate Serum and Tissue Biomarker for Ovarian Cancer|Cancer Research. Available online: https://cancerres.aacrjournals.org/content/66/3/1570.short (accessed on 17 July 2020).
- Gozal, D.; Kheirandish-Gozal, L.; Carreras, A.; Khalyfa, A.; Peris, E. Obstructive sleep apnea and obesity are associated with reduced GPR120 plasma levels in children. Sleep Med. 2013, 14, e142. [Google Scholar] [CrossRef] [Green Version]
- Horrobin, D.F. Fatty Acids, Phospholipids, and Schizophrenia. In Handbook of Essential Fatty Acid Biology; Yehuda, S., Mostofsky, D.I., Eds.; Humana Press: Totowa, NJ, USA, 1997; pp. 245–256. [Google Scholar]
- The Fish Oil Ingredient, Docosahexaenoic Acid, Activates Cytosolic Phospholipase A2 via GPR120 Receptor to Produce Prostaglandin E2 and Plays an Anti-Inflammatory Role in Macrophages. Immunology 2014, 143, 81–95. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/imm.12296 (accessed on 23 April 2020). [CrossRef]
- Li, X.; Ballantyne, L.L.; Che, X.; Mewburn, J.D.; Kang, J.X.; Barkley, R.M.; Murphy, R.C.; Yu, Y.; Funk, C.D. Endogenously Generated Omega-3 Fatty Acids Attenuate Vascular Inflammation and Neointimal Hyperplasia by Interaction With Free Fatty Acid Receptor 4 in Mice. J. Am. Heart Assoc. 2015, 4, e001856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khandaker, G. Causal Associations between Inflammation, Cardiometablic Markers and Schizophrenia: The known unknowns. Int. J. Epidemiol. 2019, 48. [Google Scholar] [CrossRef] [PubMed]
- Omega-3 Fatty Acids in Cause, Prevention and Management of Violence in Schizophrenia: Conceptualization and Application|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S1359178918302064?token=0848FEFBEFB483C5BCDA97AA2A003B03AA91EE74DFB0C03D62CB53B0889C696177E1BF7FE8CBB5BBE28CC13B39F4C801 (accessed on 23 April 2020).
- Tang, W.; Wang, Y.; Xu, F.; Fan, W.; Zhang, Y.; Fan, K.; Wang, W.; Zhang, Y.; Zhang, C. Omega-3 fatty acids ameliorate cognitive dysfunction in schizophrenia patients with metabolic syndrome. Brain Behav. Immun. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sterol Regulatory Element Binding Protein 1 Couples Mechanical Cues and Lipid Metabolism|Nature Communications. Available online: https://www.nature.com/articles/s41467-019-09152-7 (accessed on 23 April 2020).
- Dipasquale, S.; Pariante, C.M.; Dazzan, P.; Aguglia, E.; McGuire, P.; Mondelli, V. The dietary pattern of patients with schizophrenia: A systematic review. J. Psychiatr. Res. 2013, 47, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, S.B.; Ward, P.B.; Samaras, K.; Firth, J.; Stubbs, B.; Tripodi, E.; Burrows, T.L. Dietary intake of people with severe mental illness: Systematic review and meta-analysis. Br. J. Psychiatry 2019, 214, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Sun, L.; Zhao, A.; Hu, X.; Qing, Y.; Jiang, J.; Yang, C.; Xu, T.; Wang, P.; Liu, J.; et al. Serum fatty acid patterns in patients with schizophrenia: A targeted metabonomics study. Transl. Psychiatry 2017, 7, e1176. [Google Scholar] [CrossRef]
- Martins, J.G. EPA but Not DHA Appears to Be Responsible for the Efficacy of Omega-3 Long Chain Polyunsaturated Fatty Acid Supplementation in Depression: Evidence from a Meta-Analysis of Randomized Controlled Trials. J. Am. Coll. Nutr. 2009, 28, 525–542. [Google Scholar] [CrossRef]
- Peet, M.; Brind, J.; Ramchand, C.N.; Shah, S.; Vankar, G.K. Two double-blind placebo-controlled pilot studies of eicosapentaenoic acid in the treatment of schizophrenia. Schizophr. Res. 2001, 49, 243–251. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Messamore, E.; McNamara, R.K. Detection and treatment of omega-3 fatty acid deficiency in psychiatric practice: Rationale and implementation. Lipids Health Dis. 2016, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Thompson, F.E.; Subar, A.F. Dietary Assessment Methodology. In Nutrition in the Prevention and Treatment of Disease; Elsevier: Amsterdam, The Netherlands, 2017; pp. 5–48. [Google Scholar]
- Metherel, A.H.; Stark, K.D. The stability of blood fatty acids during storage and potential mechanisms of degradation: A review. Prostaglandins Leukot. Essent. Fatty Acids 2016, 104, 33–43. [Google Scholar] [CrossRef]
- Van Der Burg, K.P.; Cribb, L.; Firth, J.; Karmacoska, D.; Sarris, J. Nutrient and genetic biomarkers of nutraceutical treatment response in mood and psychotic disorders: A systematic review. Nutr. Neurosci. 2019. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Clinical Data | Schizophrenia (n = 40) | Healthy Controls (n = 40) | SZ vs. HC | ||
|---|---|---|---|---|---|
| Mean (Median) | SD | Mean (Median) | SD | ||
| Age | 31 (30) | 7.32 | 29 (27) | 7.93 | NS |
| BMI (kg/m2) | 26.6 (26.6) | 5.12 | 24.6 (24.2) | 4.39 | NS |
| Duration of illness (months) | 90 (78) | 83.43 | NA | NA | NA |
| Number of hospitalization | 2.7 (2) | 2.25 | NA | NA | NA |
| Olanzapine equivalents | 18.12 (15) | 13.93 | NA | NA | NA |
| PANSS total | 55.35 (54) | 26.71 | NA | NA | NA |
| Schizophrenia (n = 40) | Healthy Controls (n = 40) | SZ vs. HC | |||
|---|---|---|---|---|---|
| Mean (Median) | SD | Mean (Median) | SD | ||
| Serum Measurement | |||||
| ALA (mcg/mL) | 1.61 (1.16) | 1.74 | 0.91 (0.67) | 0.93 | NS |
| EPA (mcg/mL) | 2.03 (1.37) | 2.45 | 1.45 (1.38) | 0.89 | NS |
| DHA (mcg/mL) | 6.88 (6.81) | 2.26 | 6.80 (6.41) | 1.91 | NS |
| AA (mcg/mL) | 19.63 (19.48) | 4.10 | 19.92 (19.36) | 3.75 | NS |
| PUFAs (mcg/mL) | 29.80 (28.61) | 7.62 | 28.57 (27.19) | 5.76 | NS |
| Omega-3 (mcg/mL) | 10.18 (10.05) | 4.50 | 8.65 (8.34) | 3.27 | NS |
| Omega-3/6 ratio 1 | 0.52 (0.50) | 0.20 | 0.44 (0.45) | 0.18 | NS |
| GPR120 (ng/mL) | 2.41 (1.24) | 2.72 | 2.00 (1.02) | 2.32 | NS |
| Dietary Assessment | |||||
| Fat (g) | 81.11 (78.63) | 34.65 | 85.62 (80.26) | 39.04 | NS |
| PUFAs (g) | 14.01 (10.69) | 8.20 | 12.33 (10.84) | 6.22 | NS |
| Omega-3 (g) | 2.33 (1.42) | 2.78 | 1.87 (1.66) | 1.04 | NS |
| Omega-6 (g) | 11.67 (9.35) | 7.25 | 10.45 (9.40) | 5.56 | NS |
| 18:2 LA (g) | 11.55 (9.23) | 7.24 | 10.34 (9.35) | 5.51 | NS |
| 20:5 EPA (mg) | 35.45 (0) | 187.63 | 6.21 (0) | 17.54 | NS |
| 22:6 DHA (mg) | 52.72 (10) | 177.18 | 28.65 (10) | 71.07 | NS |
| GPR120 | ALA | EPA | DHA | AA | Omega-3 | Omega-3/6 Ratio 1 |
|---|---|---|---|---|---|---|
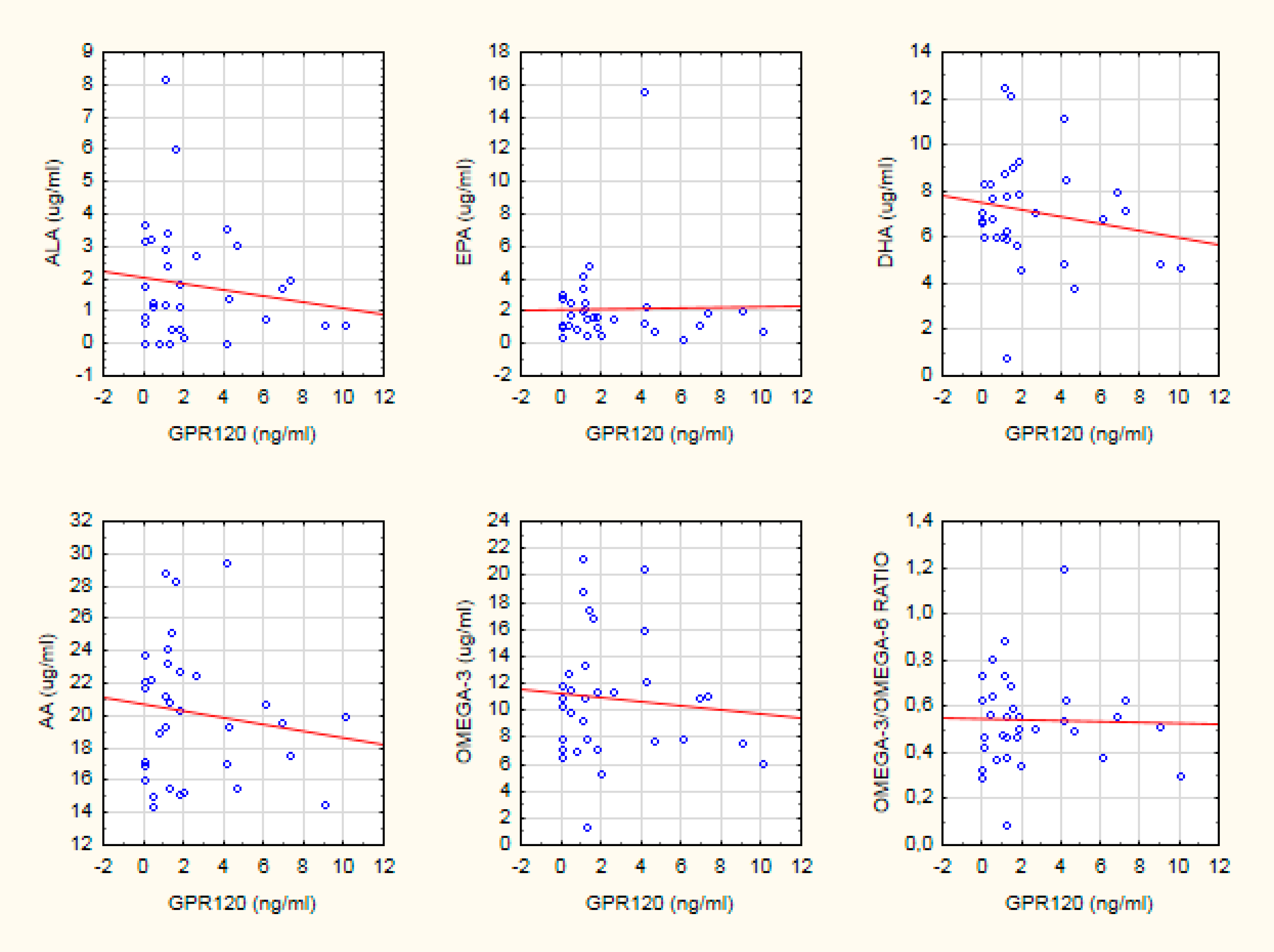
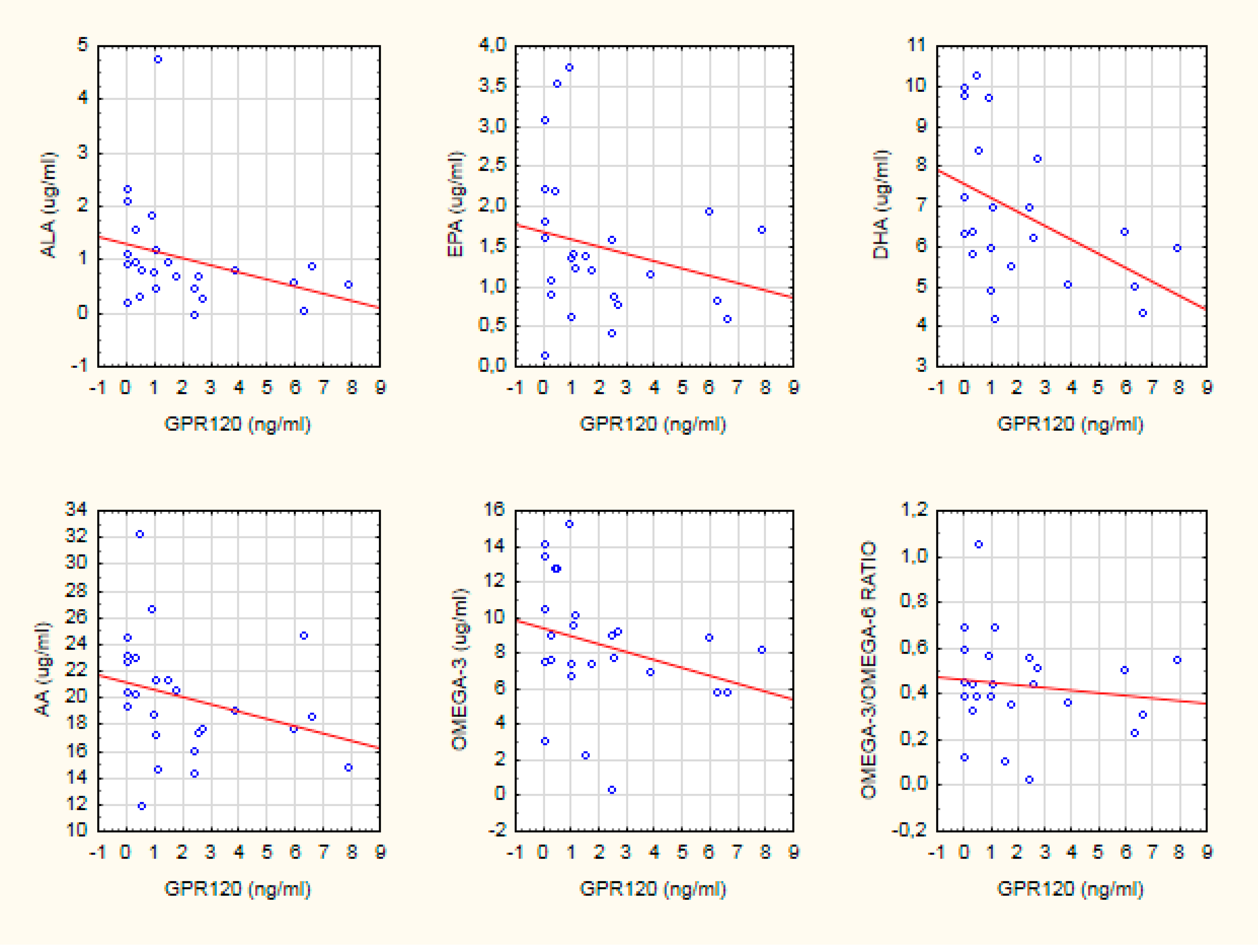
| Healthy controls | −0.46 * | NS | −0.54 * | −0.44 * | −0.41 * | NS |
| Schizophrenia | NS | NS | NS | NS | NS | NS |
| Blood | ALA | EPA | DHA | AA | Omega-3/6 Ratio 1 | GPR120 | |
|---|---|---|---|---|---|---|---|
| Diet | |||||||
| SZ group | |||||||
| 20:5 EPA | 0.46 * | NS | 0.46 * | 0.44 * | 0.35 * | NS | |
| HC group | |||||||
| PUFAs | NS | NS | 0.40 * | NS | NS | NS | |
| Omega-6 | NS | NS | 0.39 * | NS | NS | NS | |
| 18:2 LA | NS | NS | 0.40 * | NS | NS | NS | |
| 22:6 DHA | −0.34 * | NS | NS | NS | NS | NS | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rog, J.; Błażewicz, A.; Juchnowicz, D.; Ludwiczuk, A.; Stelmach, E.; Kozioł, M.; Karakula, M.; Niziński, P.; Karakula-Juchnowicz, H. The Role of GPR120 Receptor in Essential Fatty Acids Metabolism in Schizophrenia. Biomedicines 2020, 8, 243. https://doi.org/10.3390/biomedicines8080243
Rog J, Błażewicz A, Juchnowicz D, Ludwiczuk A, Stelmach E, Kozioł M, Karakula M, Niziński P, Karakula-Juchnowicz H. The Role of GPR120 Receptor in Essential Fatty Acids Metabolism in Schizophrenia. Biomedicines. 2020; 8(8):243. https://doi.org/10.3390/biomedicines8080243
Chicago/Turabian StyleRog, Joanna, Anna Błażewicz, Dariusz Juchnowicz, Agnieszka Ludwiczuk, Ewa Stelmach, Małgorzata Kozioł, Michal Karakula, Przemysław Niziński, and Hanna Karakula-Juchnowicz. 2020. "The Role of GPR120 Receptor in Essential Fatty Acids Metabolism in Schizophrenia" Biomedicines 8, no. 8: 243. https://doi.org/10.3390/biomedicines8080243