

Esaxerenone Protects against Diabetic Cardiomyopathy via Inhibition of the Chemokine and PI3K-Akt Signaling Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
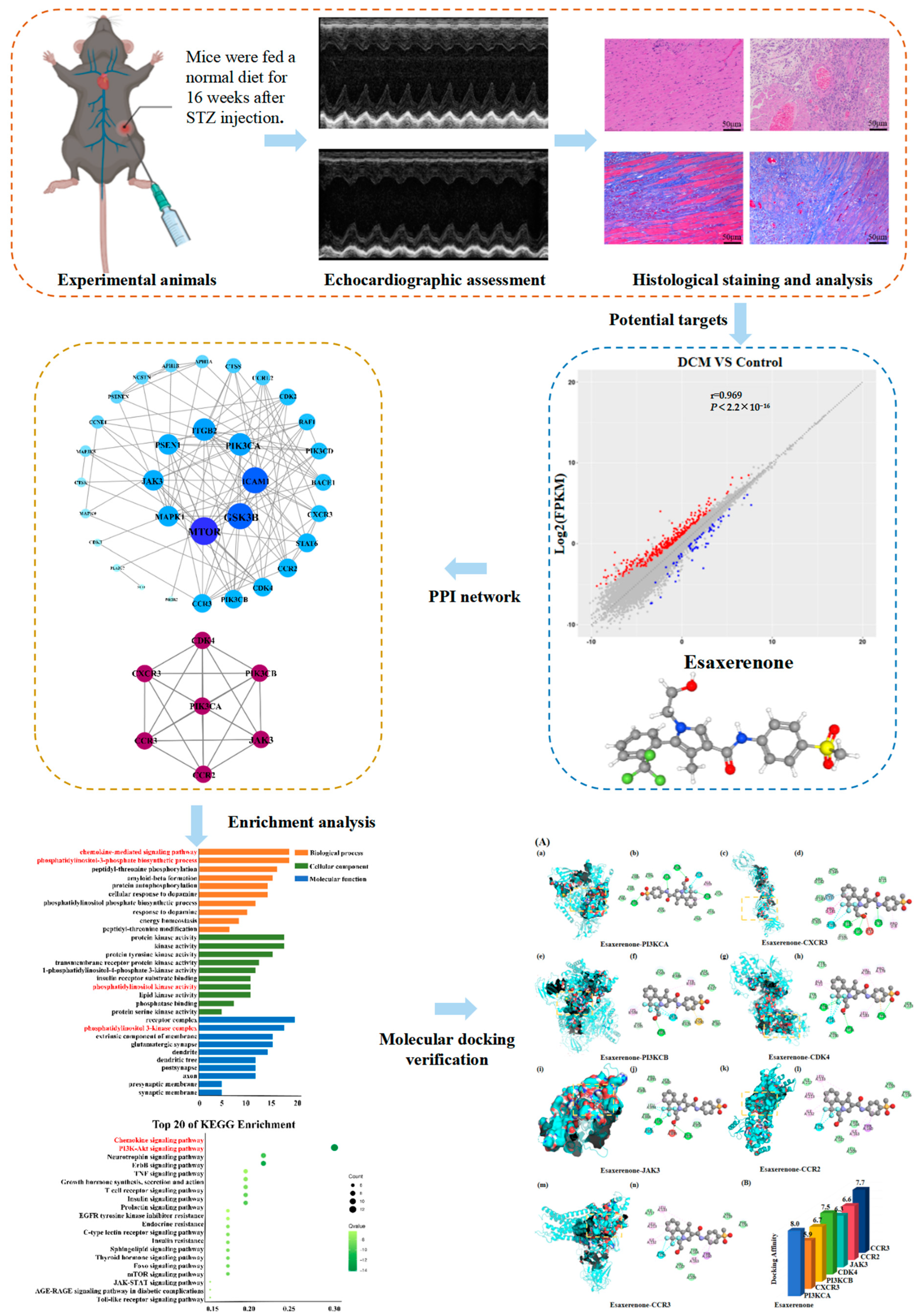
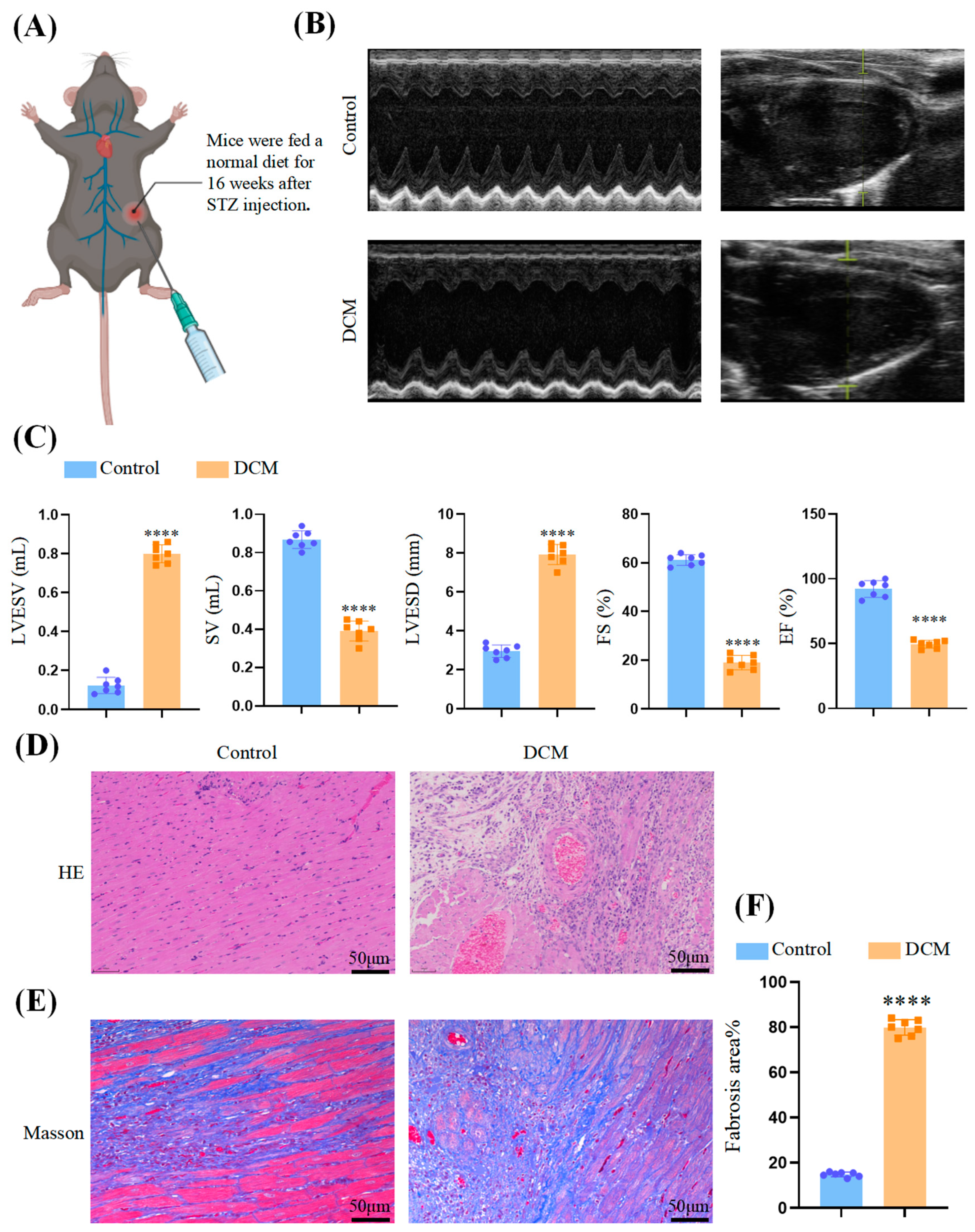
2.1. Experimental Animals
2.2. Echocardiographic Assessment
2.3. Histological Staining
2.4. Data Analysis
2.5. Network Pharmacology
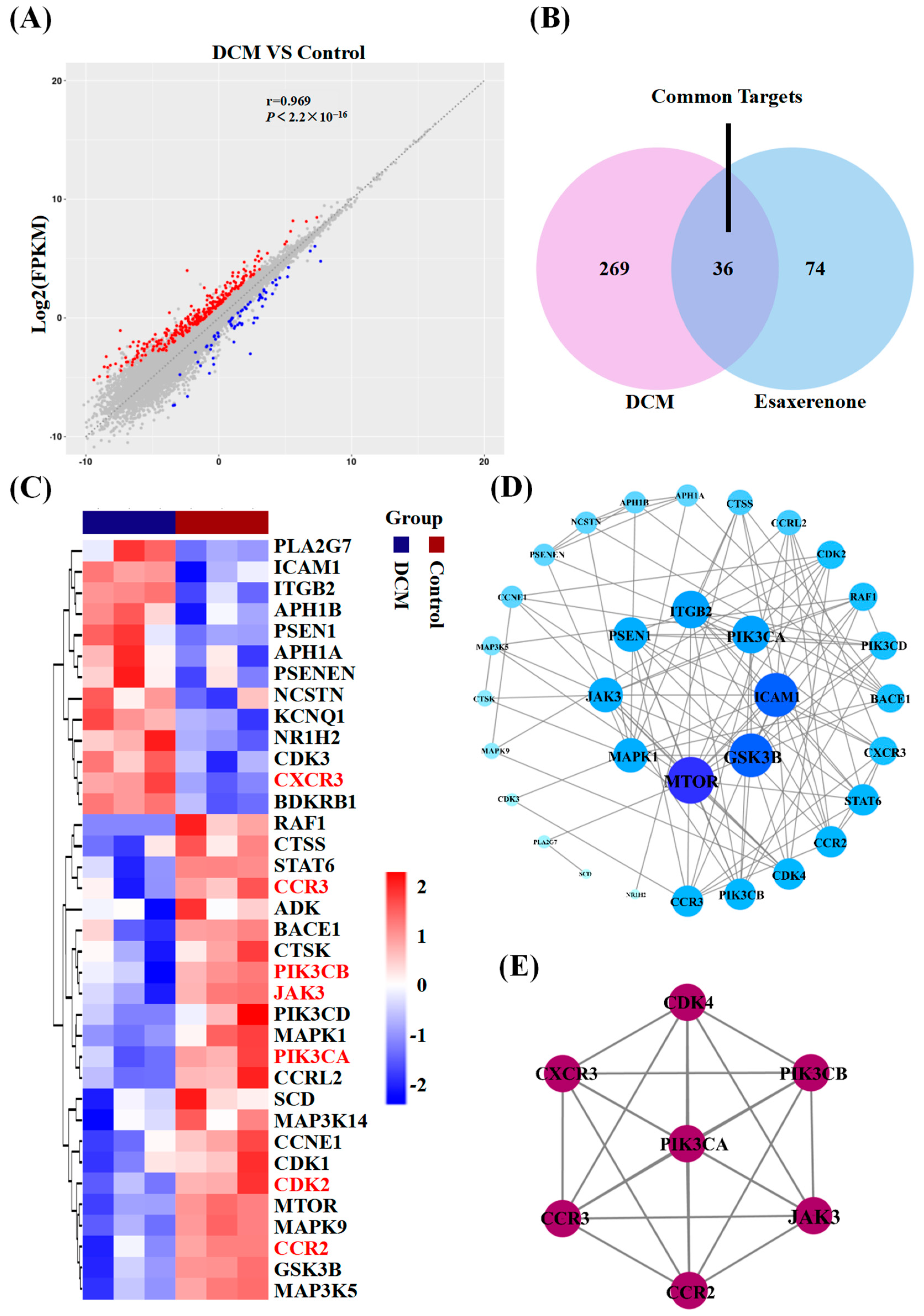
2.5.1. Potential Targets of Esaxerenone
2.5.2. Construction of PPI Network
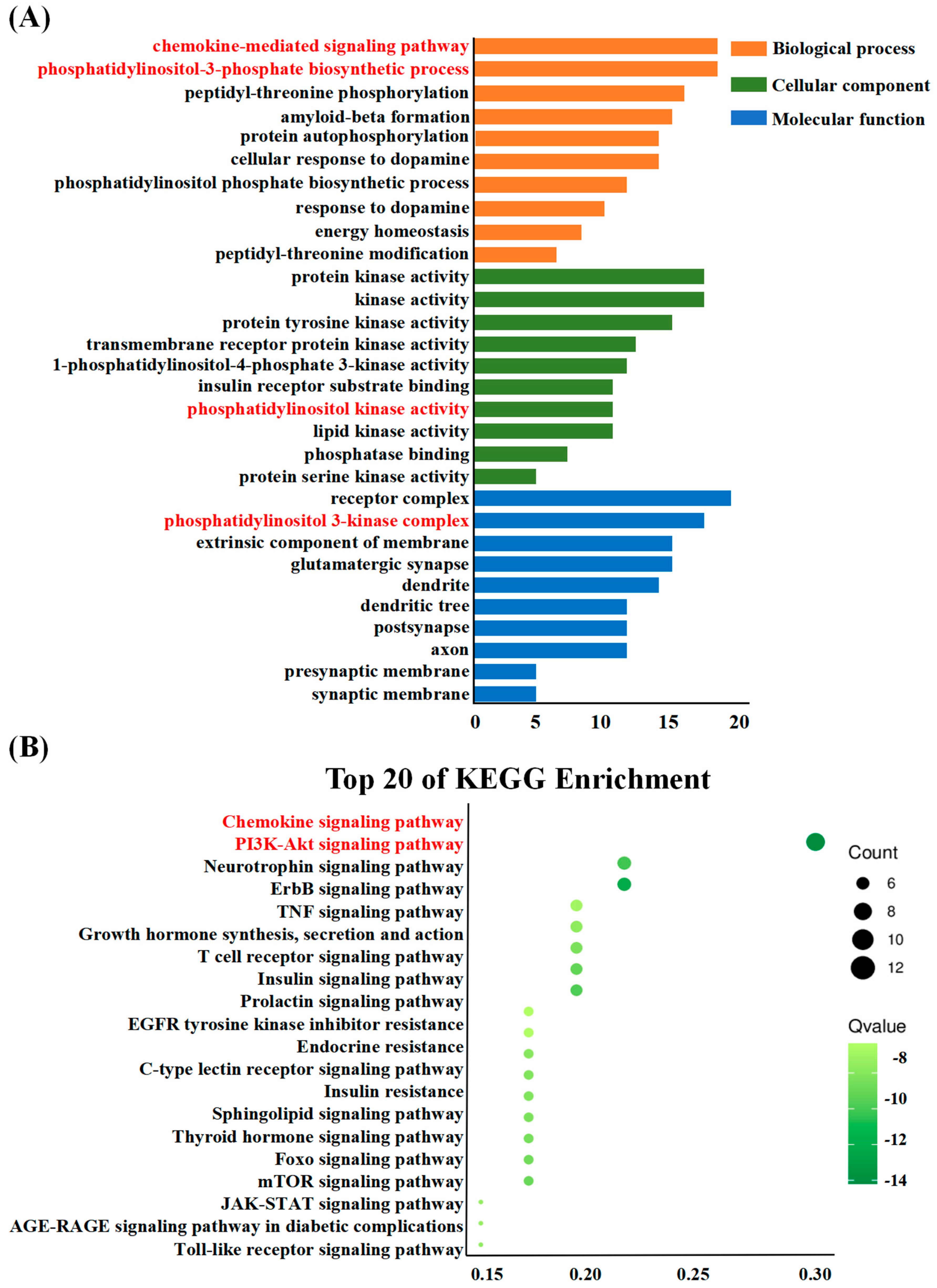
2.5.3. Enrichment Analysis
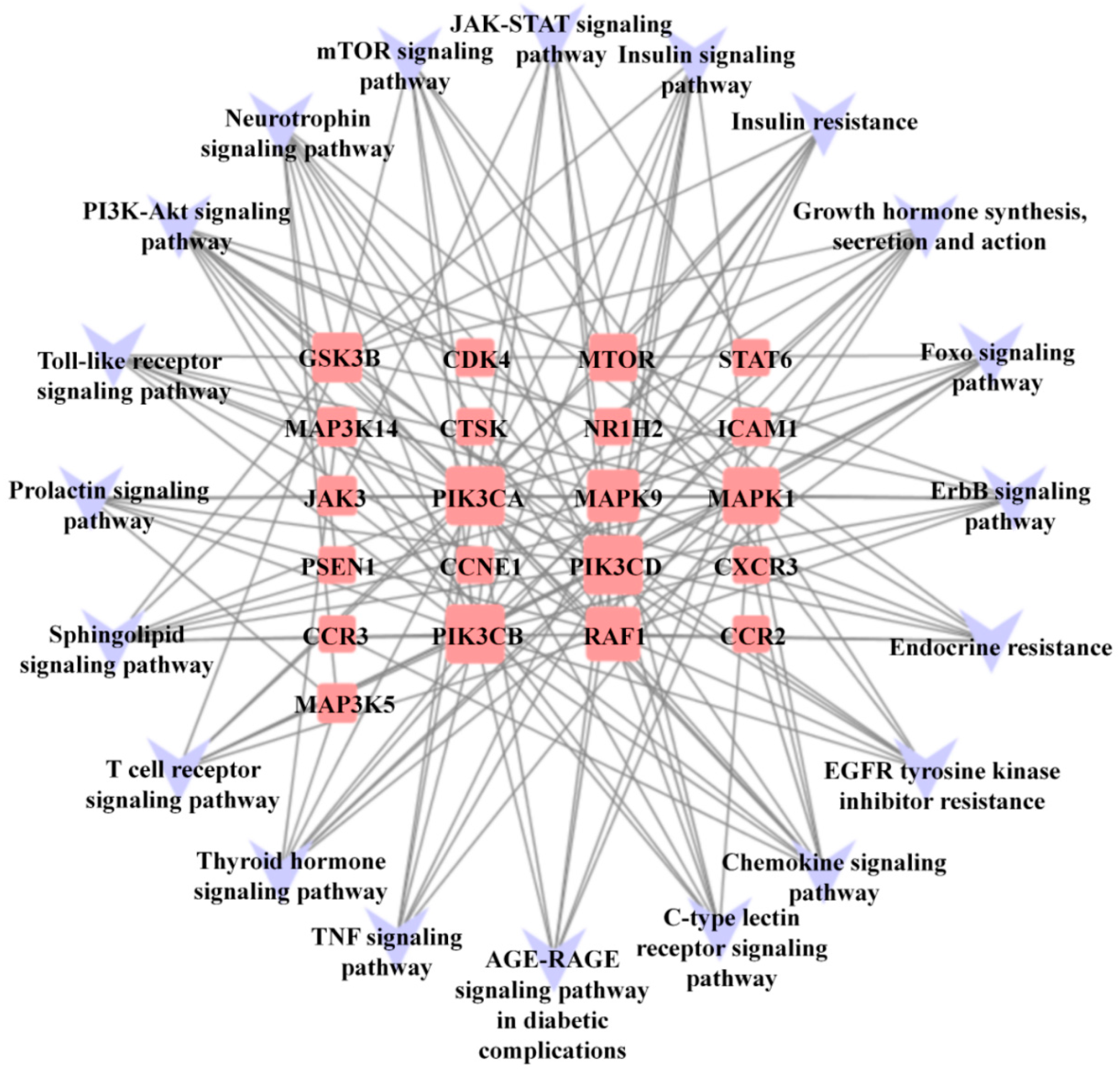
2.5.4. Construction of Target-Pathway Network
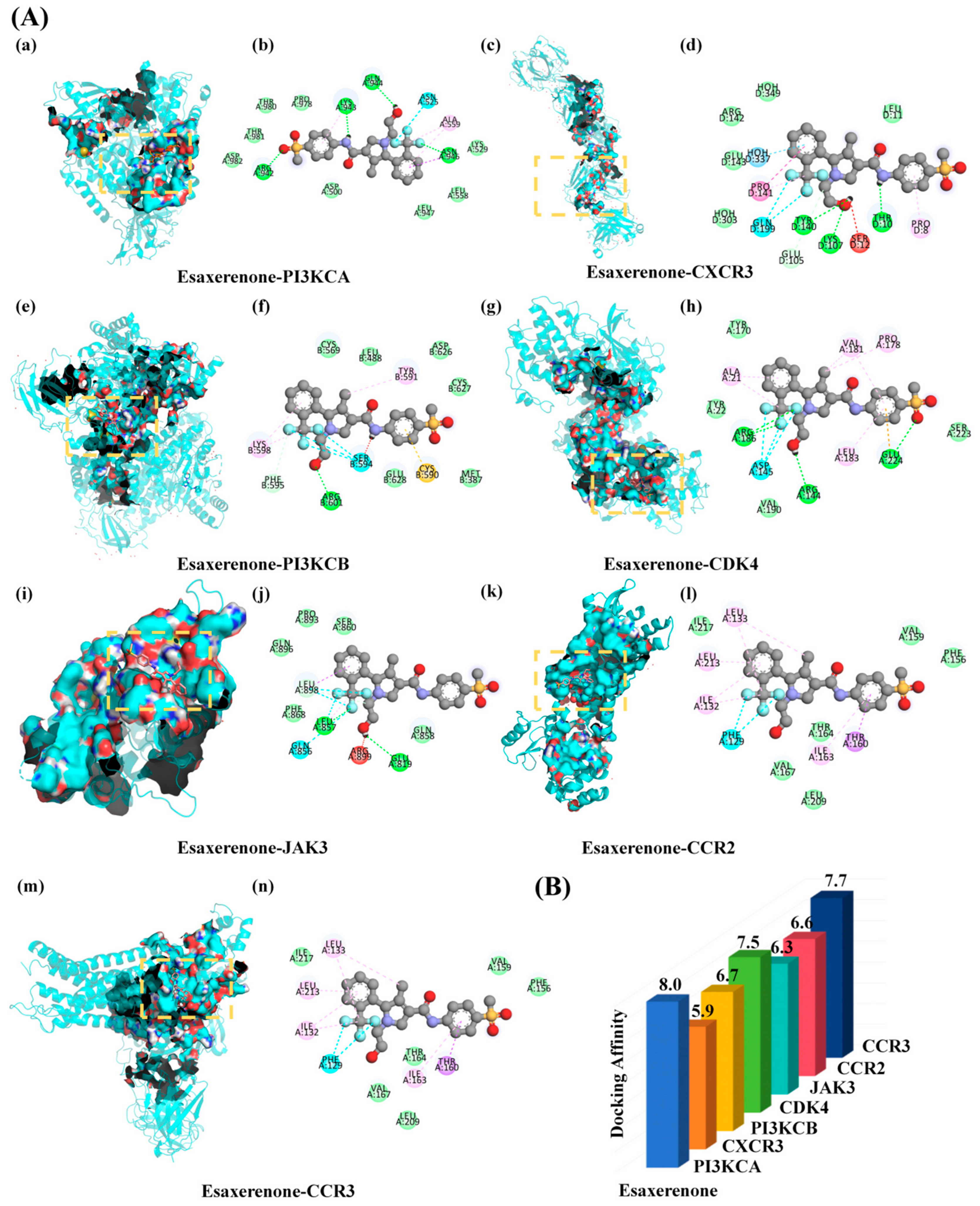
2.5.5. Molecular Docking Verification
3. Results
3.1. Myocardial Injury in DCM Mice
3.2. Candidate Targets of Esaxerenone and DCM
3.3. PPI Network
3.4. Enrichment Analysis
3.5. Compound-Pathway Network
3.6. Molecular Docking Result Analysis
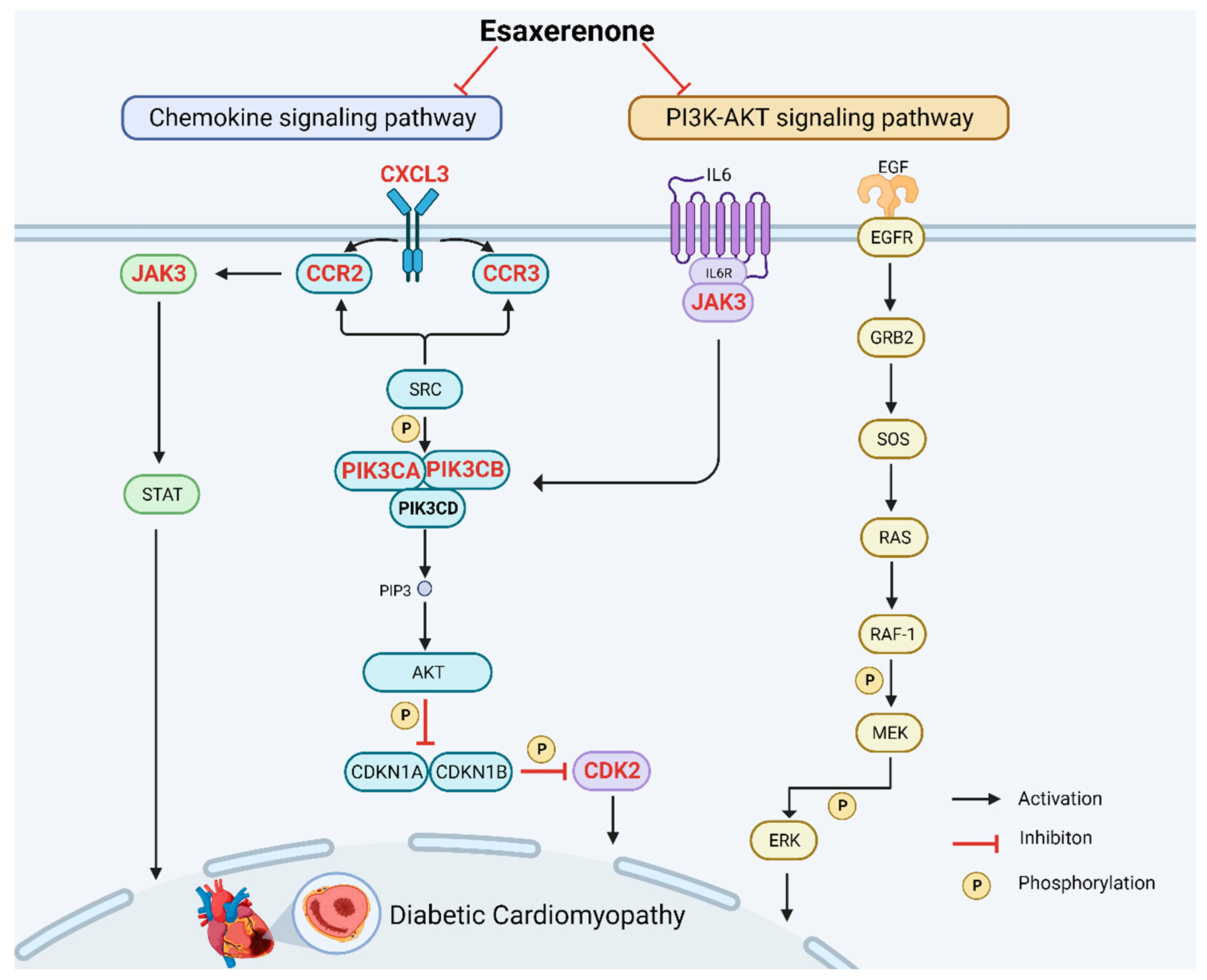
4. Discussion
Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Jia, G.; DeMarco, V.G.; Sowers, J.R. Insulin resistance and hyperinsulinaemia in diabetic cardiomyopathy. Nat. Rev. Endocrinol. 2016, 12, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Dillmann, W.H. Diabetic Cardiomyopathy. Circ. Res. 2019, 124, 1160–1162. [Google Scholar] [CrossRef] [PubMed]
- Pitt, B.; Remme, W.; Zannad, F.; Neaton, J.; Martinez, F.; Roniker, B.; Bittman, R.; Hurley, S.; Kleiman, J.; Gatlin, M. Eplerenone, a selective aldosterone blocker, in patients with left ventricular dysfunction after myocardial infarction. N. Engl. J. Med. 2003, 348, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Ubukata, O.; Homma, T.; Asoh, Y.; Honzumi, M.; Hayashi, N.; Saito, K.; Tsuruoka, H.; Aoki, K.; Hanzawa, H. Crystal structure of the mineralocorticoid receptor ligand-binding domain in complex with a potent and selective nonsteroidal blocker, esaxerenone (CS-3150). FEBS Lett. 2020, 594, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Munkhjargal, U.; Fukuda, D.; Ganbaatar, B.; Suto, K.; Matsuura, T.; Ise, T.; Kusunose, K.; Yamaguchi, K.; Yagi, S.; Yamada, H.; et al. A Selective Mineralocorticoid Receptor Blocker, Esaxerenone, Attenuates Vascular Dysfunction in Diabetic C57BL/6 Mice. J. Atheroscler. Thromb. 2023, 30, 326–334. [Google Scholar] [CrossRef]
- Arai, K.; Tsuruoka, H.; Homma, T. CS-3150, a novel non-steroidal mineralocorticoid receptor antagonist, prevents hypertension and cardiorenal injury in Dahl salt-sensitive hypertensive rats. Eur. J. Pharmacol. 2015, 769, 266–273. [Google Scholar] [CrossRef]
- Iwahana, T.; Saito, Y.; Okada, S.; Kato, H.; Ono, R.; Kobayashi, Y. Safety and efficacy of esaxerenone in Japanese hypertensive patients with heart failure with reduced ejection fraction: A retrospective study. PLoS ONE 2021, 16, e0259485. [Google Scholar] [CrossRef]
- Gao, X.; Chang, J.; Chang, Y.; Fan, L.; Liu, Z.; Zhang, C.; Shimosawa, T.; Yang, F.; Xu, Q. Esaxerenone Inhibits Renal Angiogenesis and Endothelial-Mesenchymal Transition via the VEGFA and TGF-β1 Pathways in Aldosterone-Infused Mice. Int. J. Mol. Sci. 2023, 24, 11766. [Google Scholar] [CrossRef]
- Ito, S.; Itoh, H.; Rakugi, H.; Okuda, Y.; Yoshimura, M.; Yamakawa, S. Double-Blind Randomized Phase 3 Study Comparing Esaxerenone (CS-3150) and Eplerenone in Patients With Essential Hypertension (ESAX-HTN Study). Hypertension 2020, 75, 51–58. [Google Scholar] [CrossRef]
- Ito, S.; Itoh, H.; Rakugi, H.; Okuda, Y.; Yamakawa, S. Efficacy and safety of esaxerenone (CS-3150) for the treatment of essential hypertension: A phase 2 randomized, placebo-controlled, double-blind study. J. Hum. Hypertens. 2019, 33, 542–551. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, 357–364. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, 506–515. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, 1102–1109. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Goodsell, D.S.; Olson, A.J. Automated docking of substrates to proteins by simulated annealing. Proteins. 1990, 8, 195–202. [Google Scholar] [CrossRef]
- Liu, H.; Jin, Y.; Ding, H. MDBuilder: A PyMOL plugin for the preparation of molecular dynamics simulations. Brief Bioinform. 2023, 24, bbad057. [Google Scholar] [CrossRef]
- Marfella, R.; D’Onofrio, N.; Mansueto, G.; Grimaldi, V.; Trotta, M.C.; Sardu, C.; Sasso, F.C.; Scisciola, L.; Amarelli, C.; Esposito, S.; et al. Glycated ACE2 reduces anti-remodeling effects of renin-angiotensin system inhibition in human diabetic hearts. Cardiovasc. Diabetol. 2022, 21, 146. [Google Scholar] [CrossRef]
- Marfella, R.; D’Onofrio, N.; Trotta, M.C.; Sardu, C.; Scisciola, L.; Amarelli, C.; Balestrieri, M.L.; Grimaldi, V.; Mansueto, G.; Esposito, S.; et al. Sodium/glucose cotransporter 2 (SGLT2) inhibitors improve cardiac function by reducing JunD expression in human diabetic hearts. Metabolism 2022, 127, 154936. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Kumar, V.; Gupta, S.; Bermeo-Blanco, O.; Stratton, M.S.; Gumina, R.J.; Bansal, S.S. Estrogen Receptor-β Agonists Modulate T-Lymphocyte Activation and Ameliorate Left Ventricular Remodeling During Chronic Heart Failure. Circ. Heart Fail. 2022, 15, e008997. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Zhang, H.; Xu, T.; Hao, J.; Chen, X.; Sun, S.; Yang, J.; Sun, J.; Lin, H.; Guo, H. Identification and verification of immune-related biomarkers and immune infiltration in diabetic heart failure. Front. Cardiovasc. Med. 2022, 9, 931066. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, C.S.; Li, Z.; Wang, X.; Jin, Z.Q. Depletion of T lymphocytes ameliorates cardiac fibrosis in streptozotocin-induced diabetic cardiomyopathy. Int. Immunopharmacol. 2016, 39, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Gupta, S.; Kumar, V.; Gumina, R.J.; Bansal, S.S. Estrogenic bias in T-Lymphocyte biology: Implications for cardiovascular disease. Pharmacol. Res. 2021, 170, 105606. [Google Scholar] [CrossRef] [PubMed]
- De Blasio, M.J.; Huynh, K.; Qin, C.; Rosli, S.; Kiriazis, H.; Ayer, A.; Cemerlang, N.; Stocker, R.; Du, X.J.; McMullen, J.R.; et al. Therapeutic targeting of oxidative stress with coenzyme Q10 counteracts exaggerated diabetic cardiomyopathy in a mouse model of diabetes with diminished PI3K(p110α) signaling. Free Radic. Biol. Med. 2015, 87, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Duan, J.; Liao, J.; Wang, Y.; Xiao, X.; Li, L.; Liu, Y.; Gu, H.; Yang, P.; Fu, D.; et al. LncRNA H19 inhibits ER stress induced apoptosis and improves diabetic cardiomyopathy by regulating PI3K/AKT/mTOR axis. Aging 2022, 14, 6809–6828. [Google Scholar] [CrossRef]
- Wu, X.; Yang, X.; Xiong, Y.; Li, R.; Ito, T.; Ahmed, T.A.; Karoulia, Z.; Adamopoulos, C.; Wang, H.; Wang, L.; et al. Distinct CDK6 complexes determine tumor cell response to CDK4/6 inhibitors and degraders. Nat. Cancer 2021, 2, 429–443. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Wang, Y.; Liu, Q. Palbociclib improves cardiac dysfunction in diabetic cardiomyopathy by regulating Rb phosphorylation. Am. J. Transl. Res. 2019, 11, 3481–3489. [Google Scholar]
- Quero, L.; Tiaden, A.N.; Hanser, E.; Roux, J.; Laski, A.; Hall, J.; Kyburz, D. miR-221-3p Drives the Shift of M2-Macrophages to a Pro-Inflammatory Function by Suppressing JAK3/STAT3 Activation. Front. Immunol. 2019, 10, 3087. [Google Scholar] [CrossRef]
- Thomis, D.C.; Berg, L.J. The role of Jak3 in lymphoid development, activation, and signaling. Curr. Opin. Immunol. 1997, 9, 541–547. [Google Scholar] [CrossRef]
- Al-Rasheed, N.M.; Al-Rasheed, N.M.; Hasan, I.H.; Al-Amin, M.A.; Al-Ajmi, H.N.; Mahmoud, A.M. Sitagliptin attenuates cardiomyopathy by modulating the JAK/STAT signaling pathway in experimental diabetic rats. Drug Des. Devel Ther. 2016, 10, 2095–2107. [Google Scholar] [CrossRef]
- Pease, J.E.; Williams, T.J. Tipping the balance: A biased nanobody antagonist of CCR3 with potential for the treatment of eosinophilic inflammation. J. Allergy Clin. Immunol. 2019, 143, 552–553. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, K.; Chen, F.; Liu, Q.; Ni, J.; Cao, W.; Hua, Y.; He, F.; Liu, Z.; Li, L.; et al. Role of the CCL2-CCR2 axis in cardiovascular disease: Pathogenesis and clinical implications. Front. Immunol. 2022, 13, 975367. [Google Scholar] [CrossRef]
- Van Sweringen, H.L.; Sakai, N.; Tevar, A.D.; Burns, J.M.; Edwards, M.J.; Lentsch, A.B. CXC chemokine signaling in the liver: Impact on repair and regeneration. Hepatology 2011, 54, 1445–1453. [Google Scholar] [CrossRef]
- Kong, D.; Huang, S.; Miao, X.; Li, J.; Wu, Z.; Shi, Y.; Liu, H.; Jiang, Y.; Yu, X.; Xie, M.; et al. The dynamic cellular landscape of grafts with acute rejection after heart transplantation. J. Heart Lung Transplant. 2023, 42, 160–172. [Google Scholar] [CrossRef]
- Saxena, A.; Bujak, M.; Frunza, O.; Dobaczewski, M.; Gonzalez-Quesada, C.; Lu, B.; Gerard, C.; Frangogiannis, N.G. CXCR3-independent actions of the CXC chemokine CXCL10 in the infarcted myocardium and in isolated cardiac fibroblasts are mediated through proteoglycans. Cardiovasc. Res. 2014, 103, 217–227. [Google Scholar] [CrossRef]
- Ramírez, E.; Klett-Mingo, M.; Ares-Carrasco, S.; Picatoste, B.; Ferrarini, A.; Rupérez, F.J.; Caro-Vadillo, A.; Barbas, C.; Egido, J.; Tuñón, J.; et al. Eplerenone attenuated cardiac steatosis, apoptosis and diastolic dysfunction in experimental type-II diabetes. Cardiovasc. Diabetol. 2013, 12, 172. [Google Scholar] [CrossRef]
- Filippatos, G.; Anker, S.D.; Agarwal, R.; Pitt, B.; Ruilope, L.M.; Rossing, P.; Kolkhof, P.; Schloemer, P.; Tornus, I.; Joseph, A.; et al. Finerenone and Cardiovascular Outcomes in Patients With Chronic Kidney Disease and Type 2 Diabetes. Circulation 2021, 143, 540–552. [Google Scholar] [CrossRef]
- Bakris, G.L.; Agarwal, R.; Anker, S.D.; Pitt, B.; Ruilope, L.M.; Rossing, P.; Kolkhof, P.; Nowack, C.; Schloemer, P.; Joseph, A.; et al. Effect of Finerenone on Chronic Kidney Disease Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2020, 383, 2219–2229. [Google Scholar] [CrossRef]
- Keane, M.P.; Strieter, R.M. Chemokine signaling in inflammation. Crit. Care Med. 2000, 28, N13–N26. [Google Scholar] [CrossRef]
- Wang, J.; Knaut, H. Chemokine signaling in development and disease. Development 2014, 141, 4199–4205. [Google Scholar] [CrossRef]
- Li, M.; Murabito, A.; Ghigo, A.; Hirsch, E. PI3Ks in Diabetic Cardiomyopathy. J. Cardiovasc. Pharmacol. 2017, 70, 422–429. [Google Scholar] [CrossRef]
- McMullen, J.R.; Amirahmadi, F.; Woodcock, E.A.; Schinke-Braun, M.; Bouwman, R.D.; Hewitt, K.A.; Mollica, J.P.; Zhang, L.; Zhang, Y.; Shioi, T.; et al. Protective effects of exercise and phosphoinositide 3-kinase(p110alpha) signaling in dilated and hypertrophic cardiomyopathy. Proc. Natl. Acad. Sci. USA 2007, 104, 612–617. [Google Scholar] [CrossRef]
- Ruilope, L.M.; Agarwal, R.; Anker, S.D.; Bakris, G.L.; Filippatos, G.; Nowack, C.; Kolkhof, P.; Joseph, A.; Mentenich, N.; Pitt, B. Design and Baseline Characteristics of the Finerenone in Reducing Cardiovascular Mortality and Morbidity in Diabetic Kidney Disease Trial. Am. J. Nephrol. 2019, 50, 345–356. [Google Scholar] [CrossRef]
- Frustaci, A.; Letizia, C.; Chimenti, C.; Verardo, R.; Alfarano, M.; Scialla, R.; Bagnato, G.; Miraldi, F.; Sansone, L.; Russo, M.A. Myocardial Aldosterone Receptor and Aquaporin 1 Up-Regulation Is Associated with Cardiomyocyte Remodeling in Human Heart Failure. J. Clin. Med. 2021, 10, 4854. [Google Scholar] [CrossRef]
- Frustaci, A.; Letizia, C.; Verardo, R.; Grande, C.; Francone, M.; Sansone, L.; Russo, M.A.; Chimenti, C. Primary aldosteronism-associated cardiomyopathy: Clinical-pathologic impact of aldosterone normalization. Int. J. Cardiol. 2019, 292, 141–147. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhang, H.; Zheng, W.; Yan, Z.; Yang, J.; Li, S.; Huang, W. Esaxerenone Protects against Diabetic Cardiomyopathy via Inhibition of the Chemokine and PI3K-Akt Signaling Pathway. Biomedicines 2023, 11, 3319. https://doi.org/10.3390/biomedicines11123319
Li Z, Zhang H, Zheng W, Yan Z, Yang J, Li S, Huang W. Esaxerenone Protects against Diabetic Cardiomyopathy via Inhibition of the Chemokine and PI3K-Akt Signaling Pathway. Biomedicines. 2023; 11(12):3319. https://doi.org/10.3390/biomedicines11123319
Chicago/Turabian StyleLi, Ziyue, Huihui Zhang, Weihan Zheng, Zi Yan, Jiaxin Yang, Shiyu Li, and Wenhua Huang. 2023. "Esaxerenone Protects against Diabetic Cardiomyopathy via Inhibition of the Chemokine and PI3K-Akt Signaling Pathway" Biomedicines 11, no. 12: 3319. https://doi.org/10.3390/biomedicines11123319