
Gender-Specific Interactions in a Visual Object Recognition Task in Persons with Opioid Use Disorder
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Ethical Considerations
2.3. Inclusion Criteria
2.4. EEG Procedures
2.5. Preparation for Evoked Potential Recording
2.6. Object Recognition Task
2.7. Wavelet Analysis
2.8. Statistical Analyses
3. Results
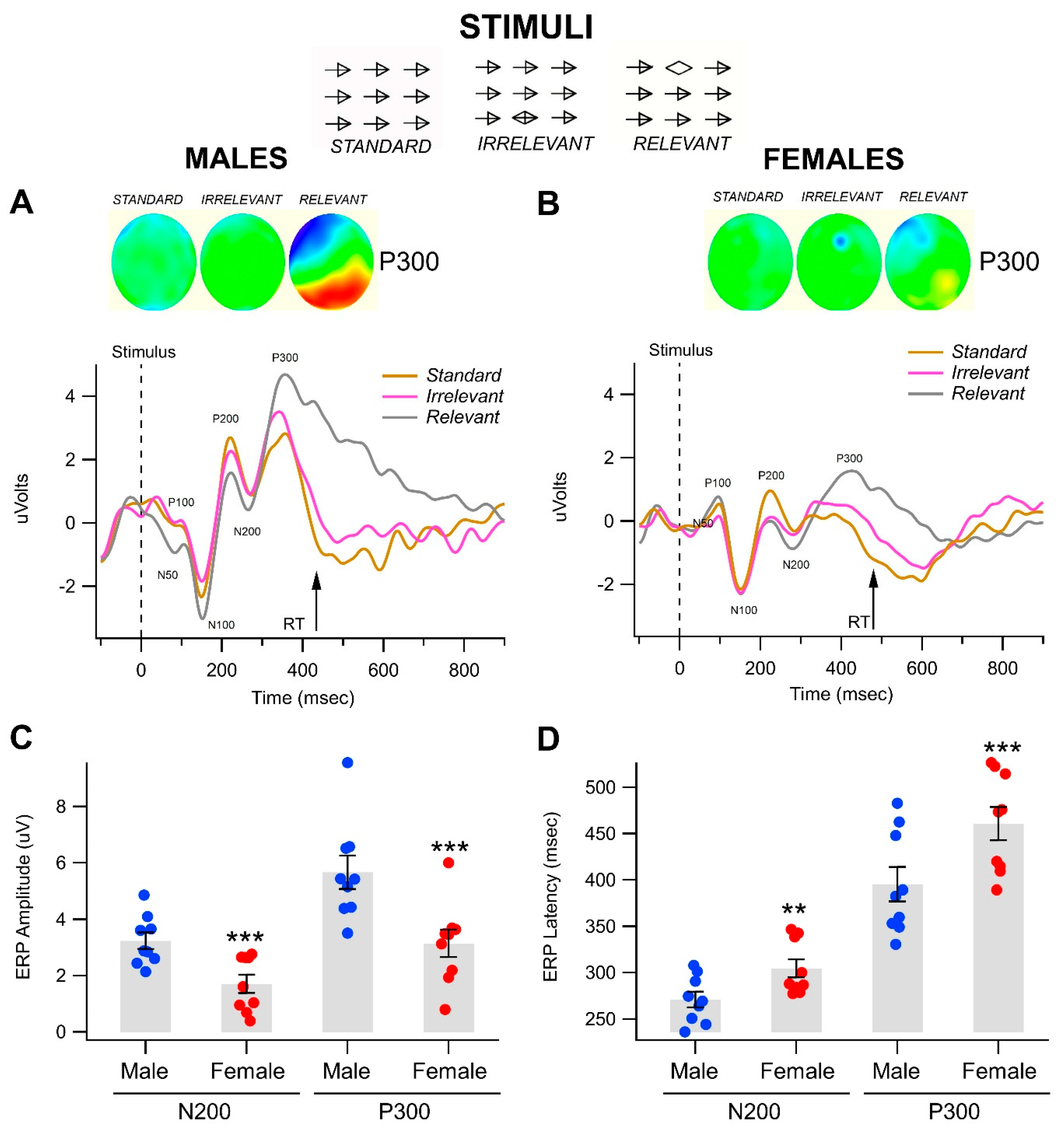
3.1. Gender-Related Differences in Event-Related Potentials in the Visual Object Recognition Task
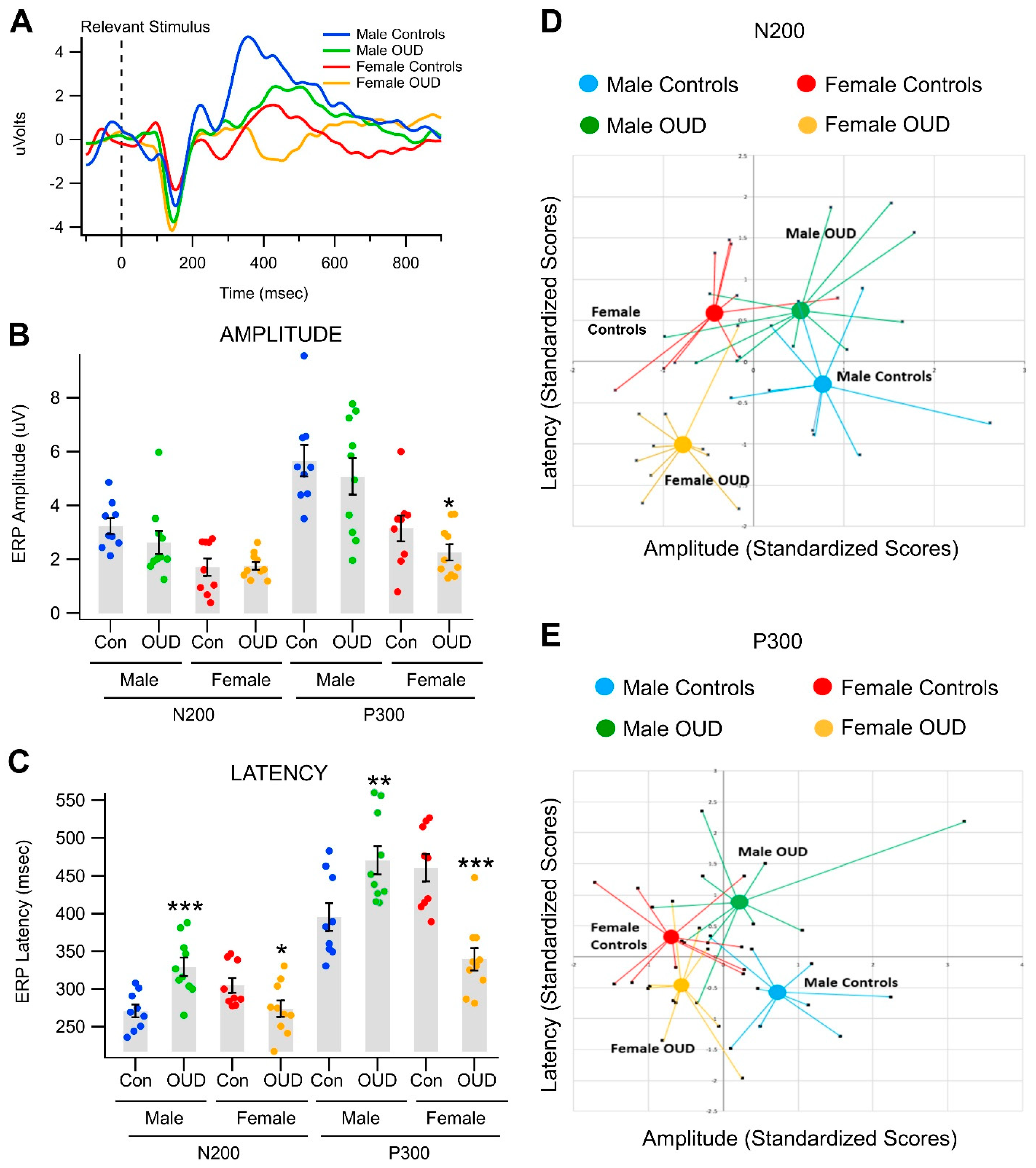
3.2. Effects of Opioid Use Disorder on Event-Related Potentials in a Visual Object Recognition Task
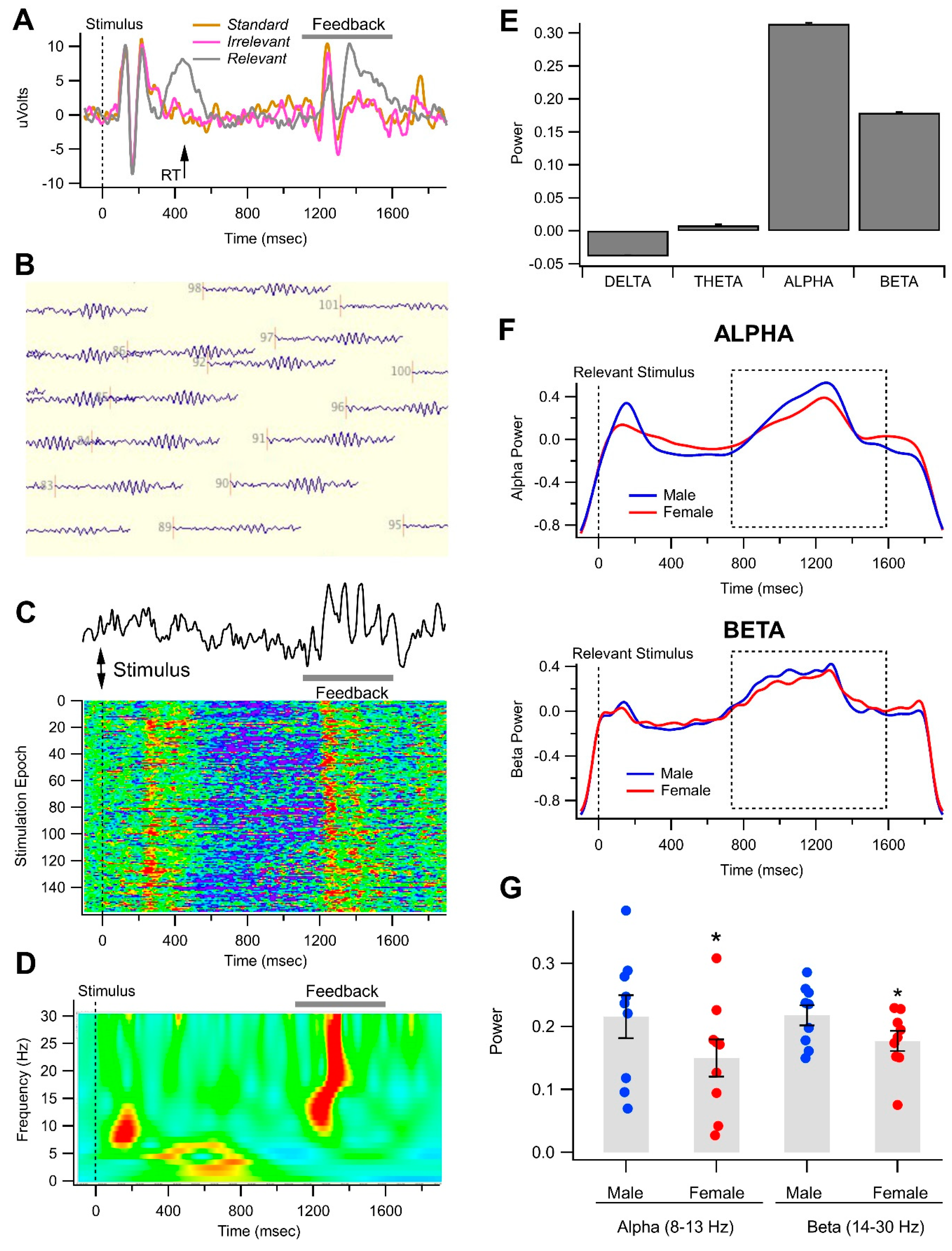
3.3. Gender-Related Differences in Alpha and Beta Oscillations in the Visual Object Recognition Task
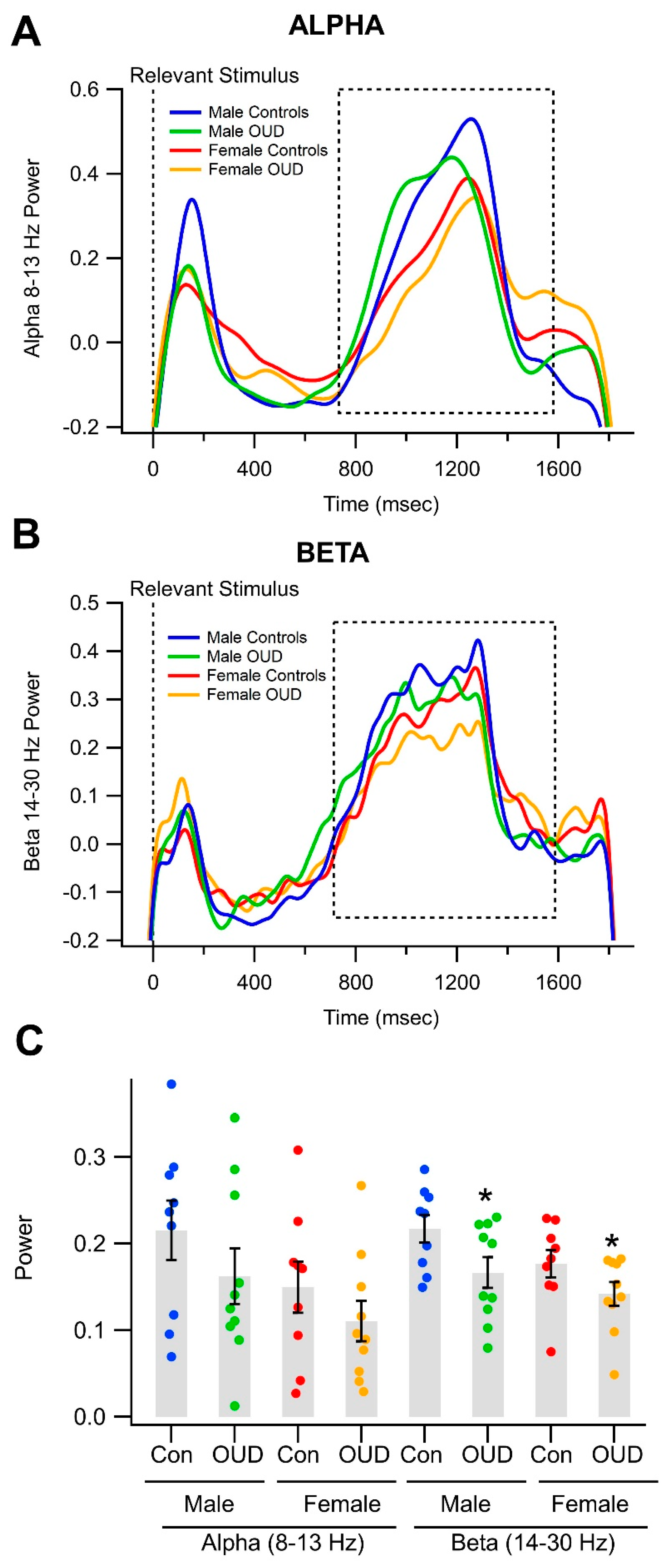
3.4. Effects of Opioid Use Disorder on Alpha/Beta Oscillations in a Visual Object Recognition Task
4. Discussion
4.1. Event-Related Potential (ERP) Studies
The P300 and N200 as Good Measures of OUD
4.2. Potential Limitations
4.3. Correlation of ERPs with Wavelets
4.4. Feedback Message Oscillation Studies
4.5. Future Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association (APA). Diagnostic and Statistical Manual of Mental Disorders—Fourth Edition (DSM-IV); American Psychiatric Association: Washington, DC, USA, 1994; p. 886. [Google Scholar]
- American Psychiatric Association (APA). DSM-5 Development: Substance Related Disorders. Available online: http://www.dsm5.org/ProposedRevisions/Pages/Substance-RelatedDisorders.aspx (accessed on 20 February 2012).
- American Psychiatric Association (APA). Diagnostic and Statistical Manual of Mental Disorders—Fifth Edition (DSM-5); American Psychiatric Publishing: Arlington, VA, USA, 2013; p. 947. [Google Scholar]
- Grant, J.E.; Chamberlain, S.R. Expanding the definition of addiction: DSM-5 vs. ICD-11. CNS Spectr. 2016, 21, 300–303. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.P. Recent developments in the pharmacotherapy of substance abuse. J. Consult. Clin. Psychol. 1996, 64, 677–686. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association (APA). What Is Addiction? Available online: https://www.psychiatry.org/patients-families/addiction/what-is-addiction (accessed on 12 November 2020).
- Koob, G.F. Neurobiological substrates for the dark side of compulsivity in addiction. Neuropharmacology 2009, 56 (Suppl. S1), 18–31. [Google Scholar] [CrossRef] [PubMed]
- Dobs, Y.E.; Ali, M.M. The epigenetic modulation of alcohol/ethanol and cannabis exposure/co-exposure during different stages. Open Biol. 2019, 9, 180115. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, N.W.; Koob, G.F. Neurobiology of alcohol dependence: Focus on motivational mechanisms. Alcohol Res. Health 2008, 31, 185–195. [Google Scholar] [PubMed]
- Polter, A.M.; Kauer, J.A. Stress and VTA synapses: Implications for addiction and depression. Eur. J. Neurosci. 2014, 39, 1179–1188. [Google Scholar] [CrossRef]
- DiFranza, J.R. Hooked from the first cigarette. Sci. Am. 2008, 298, 82–87. [Google Scholar] [CrossRef]
- United Nations Office on Drugs and Crime (UNODC). World Drug Report 2020; Sales No. E.20.XI.6; United Nations: New York, NY, USA, 20 June 2020; p. 52.
- Bartram, A.; Eliott, J.; Crabb, S. ‘Why can’t I just not drink?’ A qualitative study of adults’ social experiences of stopping or reducing alcohol consumption. Drug Alcohol Rev. 2017, 36, 449–455. [Google Scholar] [CrossRef]
- Hoffman, J.; Froemke, S. (Eds.) Addiction: Why Can’t They Just Stop? 1st ed.; Rodale Books: Emmaus, PA, USA, 2007; p. 256. [Google Scholar]
- Substance Abuse and Mental Health Services Administration (SAMHSA). Key Substance Use and Mental Health Indicators in the United States: Results from the 2019 National Survey on Drug Use and Health; Center for Behavioral Health Statistics and Quality, Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2020.
- The Council of Economic Advisors: Executive Office of the President of the United States 2019. The Underestimated Cost of the Opioid Crisis. Available online: https://crisisnextdoor.trumpwhitehouse.archives.gov/ (accessed on 20 July 2023).
- Substance Abuse and Mental Health Services Administration (SAMHSA). Behavioral Health Treatment and Services. Available online: https://www.samhsa.gov/find-help/treatment#opioid (accessed on 14 November 2020).
- U.S. Department of Health & Human Services. Fact Sheet: SAMHSA 42 CFR Part 2 Revised Rule; U.S. Department of Health & Human Services: Washington, DC, USA, 2020.
- Volkow, N.D.; Blanco, C. The changing opioid crisis: Development, challenges and opportunities. Mol. Psychiatry 2020, 26, 218–233. [Google Scholar] [CrossRef]
- Wilson, N.; Kariisa, M.; Seth, P.; Smith, H., IV; Davis, N.L. Drug and opioid-involved overdose deaths—United States, 2017–2018. Morb. Mortal. Wkly. Rep. 2020, 69, 290–297. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Parker, T.S. The Opioid Epidemic: A Geography in Two Phases ERR-287; U.S. Department of Agriculture, Economic Research Service: Washington, DC, USA, 2021; p. 48.
- National Institute on Drug Abuse (NIDA). Drug Overdose Death Rates. In Trends & Statistics; National Institute on Drug Abuse (NIDA): North Bethesda, MD, USA, 2023. [Google Scholar]
- National Institute on Drug Abuse (NIDA). Fentanyl DrugFacts. 1 June 2021. Available online: https://nida.nih.gov/publications/drugfacts/fentanyl (accessed on 21 July 2023).
- Lee, S.K. Sex as an important biological variable in biomedical research. BMB Rep. 2018, 51, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Hilderbrand, E.R.; Lasek, A.W. Studying sex differences in animal models of addiction: An emphasis on alcohol-related behaviors. ACS Chem. Neurosci. 2018, 9, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Halpern, D.F.; Benbow, C.P.; Geary, D.C.; Gur, R.C.; Hyde, J.S.; Gernsbacher, M.A. The science of sex differences in science and mathematics. Psychol. Sci. Public Interest 2007, 8, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.B.; Grella, C.E. Gender differences among older heroin users. J. Women Aging 2009, 21, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Back, S.E.; Payne, R.L.; Wahlquist, A.H.; Carter, R.E.; Stroud, Z.; Haynes, L.; Hillhouse, M.; Brady, K.T.; Ling, W. Comparative profiles of men and women with opioid dependence: Results from a national multisite effectiveness trial. Am. J. Drug Alcohol Abuse 2011, 37, 313–323. [Google Scholar] [CrossRef]
- Green, T.C.; Grimes Serrano, J.M.; Licari, A.; Budman, S.H.; Butler, S.F. Women who abuse prescription opioids: Findings from the Addiction Severity Index-Multimedia Version Connect prescription opioid database. Drug Alcohol Depend. 2009, 103, 65–73. [Google Scholar] [CrossRef]
- Ait-Daoud, N.; Blevins, D.; Khanna, S.; Sharma, S.; Holstege, C.P.; Amin, P. Women and addiction: An update. Med. Clin. N. Am. 2019, 103, 699–711. [Google Scholar] [CrossRef]
- Barbosa-Leiker, C.; McPherson, S.; Layton, M.E.; Burduli, E.; Roll, J.M.; Ling, W. Sex differences in opioid use and medical issues during buprenorphine/naloxone treatment. Am. J. Drug Alcohol Abuse 2018, 44, 488–496. [Google Scholar] [CrossRef]
- Barker, J.M.; Taylor, J.R. Sex differences in incentive motivation and the relationship to the development and maintenance of alcohol use disorders. Physiol. Behav. 2019, 203, 91–99. [Google Scholar] [CrossRef]
- Becker, J.B. Sex differences in addiction. Dialogues Clin. Neurosci. 2016, 18, 395–402. [Google Scholar] [CrossRef]
- Becker, J.B.; Chartoff, E. Sex differences in neural mechanisms mediating reward and addiction. Neuropsychopharmacology 2019, 44, 166–183. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.B.; McClellan, M.L.; Reed, B.G. Sex differences, gender and addiction. J. Neurosci. Res. 2017, 95, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Bisagno, V.; Cadet, J.L. Stress, sex, and addiction: Potential roles of corticotropin-releasing factor, oxytocin, and arginine-vasopressin. Behav. Pharmacol. 2014, 25, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Bobzean, S.A.; DeNobrega, A.K.; Perrotti, L.I. Sex differences in the neurobiology of drug addiction. Exp. Neurol. 2014, 259, 64–74. [Google Scholar] [CrossRef]
- Carroll, M.E.; Lynch, W.J. How to study sex differences in addiction using animal models. Addict. Biol. 2016, 21, 1007–1029. [Google Scholar] [CrossRef]
- Chowdhury, T.G.; Wallin-Miller, K.G.; Rear, A.A.; Park, J.; Diaz, V.; Simon, N.W.; Moghaddam, B. Sex differences in reward- and punishment-guided actions. Cogn. Affect. Behav. Neurosci. 2019, 19, 1404–1417. [Google Scholar] [CrossRef]
- Dahan, A.; Kest, B.; Waxman, A.R.; Sarton, E. Sex-specific responses to opiates: Animal and human studies. Anesth. Analg. 2008, 107, 83–95. [Google Scholar] [CrossRef]
- Fattore, L.; Melis, M. Sex differences in impulsive and compulsive behaviors: A focus on drug addiction. Addict. Biol. 2016, 21, 1043–1051. [Google Scholar] [CrossRef]
- Huhn, A.S.; Berry, M.S.; Dunn, K.E. Review: Sex-based differences in treatment outcomes for persons with opioid use disorder. Am. J. Addict. 2019, 28, 246–261. [Google Scholar] [CrossRef]
- Kaplovitch, E.; Gomes, T.; Camacho, X.; Dhalla, I.A.; Mamdani, M.M.; Juurlink, D.N. Sex differences in dose escalation and overdose death during chronic opioid therapy: A population-based cohort study. PLoS ONE 2015, 10, e0134550. [Google Scholar] [CrossRef]
- Kokane, S.S.; Perrotti, L.I. Sex differences and the role of estradiol in mesolimbic reward circuits and vulnerability to cocaine and opiate addiction. Front. Behav. Neurosci. 2020, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, J.; McEwen, B.S. Sex in the brain: Hormones and sex differences. Dialogues Clin. Neurosci. 2016, 18, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Orsini, C.A.; Setlow, B. Sex differences in animal models of decision making. J. Neurosci. Res. 2017, 95, 260–269. [Google Scholar] [CrossRef]
- Shimamoto, A. Social defeat stress, sex, and addiction-kike behaviors. Int. Rev. Neurobiol. 2018, 140, 271–313. [Google Scholar] [CrossRef] [PubMed]
- Songtachalert, T.; Roomruangwong, C.; Carvalho, A.F.; Bourin, M.; Maes, M. Anxiety disorders: Sex differences in serotonin and tryptophan metabolism. Curr. Top. Med. Chem. 2018, 18, 1704–1715. [Google Scholar] [CrossRef]
- Zachry, J.E.; Johnson, A.R.; Calipari, E.S. Sex differences in value-based decision making underlie substance use disorders in females. Alcohol Alcohol. 2019, 54, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhao, M.; Zhou, C.; Li, R. Sex differences in drug addiction and response to exercise intervention: From human to animal studies. Front. Neuroendocrinol. 2016, 40, 24–41. [Google Scholar] [CrossRef]
- Ballenger, J.C.; Post, R.M. Kindling as a model for alcohol withdrawal syndromes. Br. J. Psychiatry 1978, 133, 1–14. [Google Scholar] [CrossRef]
- Bauer, L.O. Predicting relapse to alcohol and drug abuse via quantitative electroencephalography. Neuropsychopharmacology 2001, 25, 332–340. [Google Scholar] [CrossRef]
- Begleiter, H.; Porjesz, B. Genetics of human brain oscillations. Int. J. Psychophysiol. 2006, 60, 162–171. [Google Scholar] [CrossRef]
- Bigler, E.D.; Fleming, D.E. Pharmacological suppression of photically evoked after-discharges in rats: Incremental dose, hippocampal EEG and behavioral activity correlates. Psychopharmacologia 1976, 46, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, N.A.; Bauer, L.O.; Houston, R.J. Recent EEG and ERP findings in substance abusers. Clin. EEG Neurosci. 2009, 40, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Hensel, M.; Wolter, S.; Kox, W.J. EEG controlled rapid opioid withdrawal under general anaesthesia. Br. J. Anaesth. 2000, 84, 236–238. [Google Scholar] [CrossRef]
- Herning, R.I.; Glover, B.J.; Koeppl, B.; Phillips, R.L.; London, E.D. Cocaine-induced increases in EEG alpha and beta activity: Evidence for reduced cortical processing. Neuropsychopharmacology 1994, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Howe, R.C.; Hegge, F.W.; Phillips, J.L. Acute heroin abstinence in man: II. Alterations in rapid eye movement (REM) sleep. Drug Alcohol Depend. 1980, 6, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Howe, R.C.; Hegge, F.W.; Phillips, J.L. Acute heroin abstinence in man: I. Changes in behavior and sleep. Drug Alcohol Depend. 1980, 5, 341–356. [Google Scholar] [CrossRef]
- Jurado-Barba, R.; Sion, A.; Martínez-Maldonado, A.; Domínguez-Centeno, I.; Prieto-Montalvo, J.; Navarrete, F.; García-Gutierrez, M.S.; Manzanares, J.; Rubio, G. Neuropsychophysiological measures of alcohol dependence: Can we use EEG in the clinical assessment? Front. Psychiatry 2020, 11, 676. [Google Scholar] [CrossRef]
- Kamarajan, C.; Porjesz, B.; Jones, K.; Chorlian, D.; Padmanabhapillai, A.; Rangaswamy, M.; Stimus, A.; Begleiter, H. Event-related oscillations in offspring of alcoholics: Neurocognitive disinhibition as a risk for alcoholism. Biol. Psychiatry 2006, 59, 625–634. [Google Scholar] [CrossRef]
- Logar, C.; Grabmair, W.; Reinbacher, N.; Ladurner, G. EEG changes in the withdrawal phase of tranquilizer or drug abuse. EEG EMG Z Elektroenzephalogr Elektromyogr Verwandte Geb 1986, 17, 37–40. [Google Scholar]
- Lukas, S.E.; Mendelson, J.H.; Kouri, E.; Bolduc, M.; Amass, L. Ethanol-induced alterations in EEG alpha activity and apparent source of the auditory P300 evoked response potential. Alcohol 1990, 7, 471–477. [Google Scholar] [CrossRef]
- Prichep, L.S.; Alper, K.; Kowalik, S.C.; Rosenthal, M. Neurometric QEEG studies of crack cocaine dependence and treatment outcome. J. Addict. Dis. 1996, 15, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Prichep, L.S.; Kowalik, S.C.; Alper, K.; de Jesus, C. Quantitative EEG characteristics of children exposed in utero to cocaine. Clin. Electroencephalogr. 1995, 26, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Rangaswamy, M.; Jones, K.A.; Porjesz, B.; Chorlian, D.B.; Padmanabhapillai, A.; Kamarajan, C.; Kuperman, S.; Rohrbaugh, J.; O’Connor, S.J.; Bauer, L.O.; et al. Delta and theta oscillations as risk markers in adolescent offspring of alcoholics. Int. J. Psychophysiol. 2007, 63, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Roohi-Azizi, M.; Azimi, L.; Heysieattalab, S.; Aamidfar, M. Changes of the brain’s bioelectrical activity in cognition, consciousness, and some mental disorders. Med. J. Islam. Repub. Iran 2017, 31, 53. [Google Scholar] [CrossRef]
- Urbano, F.J.; Bisagno, V.; Garcia-Rill, E. Arousal and drug abuse. Behav. Brain Res. 2017, 333, 276–281. [Google Scholar] [CrossRef]
- Volavka, J.; Zaks, A.; Roubicek, J.; Fink, M. Acute EEG effects of heroin and naloxone. Electroencephalogr. Clin. Neurophysiol. 1971, 30, 165. [Google Scholar]
- Wang, Y.K.; Jung, T.P.; Lin, C.T. Theta and alpha oscillations in attentional interaction during distracted driving. Front. Behav. Neurosci. 2018, 12, 3. [Google Scholar] [CrossRef]
- Yener, G.G.; Başar, E. Brain oscillations as biomarkers in neuropsychiatric disorders: Following an interactive panel discussion and synopsis. Suppl. Clin. Neurophysiol. 2013, 62, 343–363. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, H.; Hu, B.; Wu, H.; Liu, Q. Abstinent heroin addicts tend to take risks: ERP and source localization. Front. Neurosci. 2017, 11, 681. [Google Scholar] [CrossRef]
- Höller, Y. Quantitative EEG in Cognitive Neuroscience. Brain Sci. 2021, 11, 517. [Google Scholar] [CrossRef]
- Amass, L.; Lukas, S.E.; Weiss, R.D.; Mendelson, J. Evaluation of cognitive skills in ethanol- and cocaine-dependent patients during detoxification using P300 evoked response potentials (ERPs). NIDA Res. Monogr. 1989, 95, 353–354. [Google Scholar]
- Bahramali, H.; Gordon, E.; Li, W.M.; Rennie, C.; Wright, J.; Meares, R. Fast and slow reaction times and associated ERPs in patients with schizophrenia and controls. Int. J. Neurosci. 1998, 95, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.R.; McLarnon, M.E.; Iacono, W.G. P300 amplitude, externalizing psychopathology, and earlier- versus later-onset substance-use disorder. J. Abnorm. Psychol. 2007, 116, 565–577. [Google Scholar] [CrossRef]
- Elkashef, A.; Vocci, F. Biological markers of cocaine addiction: Implications for medications development. Addict. Biol. 2003, 8, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Emmerson, R.Y.; Dustman, R.E.; Shearer, D.E.; Chamberlin, H.M. EEG, visually evoked and event related potentials in young abstinent alcoholics. Alcohol 1987, 4, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Iacono, W.G.; Carlson, S.R.; Malone, S.M. Identifying a multivariate endophenotype for substance use disorders using psychophysiological measures. Int. J. Psychophysiol. 2000, 38, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Iacono, W.G.; Malone, S.M.; McGue, M. Substance use disorders, externalizing psychopathology, and P300 event-related potential amplitude. Int. J. Psychophysiol. 2003, 48, 147–178. [Google Scholar] [CrossRef]
- Lubman, D.I.; Yucel, M.; Kettle, J.W.; Scaffidi, A.; Mackenzie, T.; Simmons, J.G.; Allen, N.B. Responsiveness to drug cues and natural rewards in opiate addiction: Associations with later heroin use. Arch. Gen. Psychiatry 2009, 66, 205–212. [Google Scholar] [CrossRef]
- Marques-Teixeira, J.E.; Barbosa, M.F. Emotional states and informational brain processing in drug addicts free of drugs: An ERPs study. Int. J. Psychiatry Clin. Pract. 2005, 9, 213–220. [Google Scholar] [CrossRef]
- Polich, J. Clinical application of the P300 event-related brain potential. Phys. Med. Rehabil. Clin. N. Am. 2004, 15, 133–161. [Google Scholar] [CrossRef]
- Motlagh, F.; Ibrahim, F.; Rashid, R.; Seghatoleslam, T.; Habil, H. Investigation of brain electrophysiological properties among heroin addicts: Quantitative EEG and event-related potentials. J. Neurosci. Res. 2017, 95, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Bauer, L.O. CNS recovery from cocaine, cocaine and alcohol, or opioid dependence: A P300 study. Clin. Neurophysiol. 2001, 112, 1508–1515. [Google Scholar] [CrossRef]
- Steffensen, S.C.; Ohran, A.J.; Shipp, D.N.; Hales, K.; Stobbs, S.H.; Fleming, D.E. Gender-selective effects of the P300 and N400 components of the visual evoked potential. Vis. Res. 2008, 48, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Hedden, T.; Gabrieli, J.D. Insights into the ageing mind: A view from cognitive neuroscience. Nat. Rev. Neurosci. 2004, 5, 87–96. [Google Scholar] [CrossRef]
- Faresjö, T.; Faresjö, A. To match or not to match in epidemiological studies--same outcome but less power. Int. J. Environ. Res. Public Health 2010, 7, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Király, A.; Szabó, N.; Tóth, E.; Csete, G.; Faragó, P.; Kocsis, K.; Must, A.; Vécsei, L.; Kincses, Z.T. Male brain ages faster: The age and gender dependence of subcortical volumes. Brain Imaging Behav. 2016, 10, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Lortie, C.L.; Thibeault, M.; Guitton, M.J.; Tremblay, P. Effects of age on the amplitude, frequency and perceived quality of voice. Age 2015, 37, 117. [Google Scholar] [CrossRef]
- Saygili, E.; Kluttig, R.; Rana, O.R.; Saygili, E.; Gemein, C.; Zink, M.D.; Rackauskas, G.; Weis, J.; Schwinger, R.H.; Marx, N.; et al. Age-related regional differences in cardiac nerve growth factor expression. Age 2012, 34, 659–667. [Google Scholar] [CrossRef]
- Sella, O.; Jones, R.D.; Huckabee, M.L. Age and gender effects on submental motor-evoked potentials. Age 2014, 36, 9735. [Google Scholar] [CrossRef]
- Steffensen, S.C.; Taylor, S.R.; Horton, M.L.; Barber, E.N.; Lyle, L.T.; Stobbs, S.H.; Allison, D.W. Cocaine disinhibits dopamine neurons in the ventral tegmental area via use-dependent blockade of GABA neuron voltage-sensitive sodium channels. Eur. J. Neurosci. 2008, 28, 2028–2040. [Google Scholar] [CrossRef]
- Steffensen, S.C.; Walton, C.H.; Hansen, D.M.; Yorgason, J.T.; Gallegos, R.A.; Criado, J.R. Contingent and non-contingent effects of low-dose ethanol on GABA neuron activity in the ventral tegmental area. Pharmacol. Biochem. Behav. 2009, 92, 68–75. [Google Scholar] [CrossRef] [PubMed]
- CITI Program. The Collaborative Institutional Training Initiative (CITI Program). Available online: https://about.citiprogram.org/en/module/research-and-hipaa-privacy-protections/ (accessed on 8 December 2020).
- Jarvis, M.; Williams, J.; Hurford, M.; Lindsay, D.; Lincoln, P.; Giles, L.; Luongo, P.; Safarian, T. Appropriate use of drug testing in clinical addiction medicine. J. Addict. Med. 2017, 11, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Jasper, H. Progress and problems in brain research. J. Mt. Sinai Hosp. N. Y. 1958, 25, 244–253. [Google Scholar] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 6 June 2020).
- SAS Institute Inc. SAS/STAT® 9.2 User’s Guide; SAS Institute Inc.: Hongkong, China, 2008. [Google Scholar]
- Chiappa, K.A. Evoked Potentials in Clinical Medicine, 3rd ed.; Lippincott Williams & Wilkins: New York, NY, USA, 1997; p. 720. [Google Scholar]
- Luck, S.J. An Introduction to the Event-Related Potential Technique, 2nd ed.; A Bradford Book; The MIT Press: Cambridge, MA, USA, 2014; p. 406. [Google Scholar]
- Kouri, E.M.; Lukas, S.E.; Mendelson, J.H. P300 assessment of opiate and cocaine users: Effects of detoxification and buprenorphine treatment. Biol. Psychiatry 1996, 40, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Kopell, B.S.; Roth, W.T.; Tinklenberg, J.R. Time course effects of marijuana and ethanol on event-related potentials. Psychopharmacology 1978, 56, 15–20. [Google Scholar] [CrossRef]
- Begleiter, H.; Porjesz, B.; Tenner, M. Neuroradiological and neurophysiological evidence of brain deficits in chronic alcoholics. Acta Psychiatr. Scand. Suppl. 1980, 286, 3–13. [Google Scholar] [CrossRef]
- Pfefferbaum, A.; Horvath, T.B.; Roth, W.T.; Kopell, B.S. Event-related potential changes in chronic alcoholics. Electroencephalogr. Clin. Neurophysiol. 1979, 47, 637–647. [Google Scholar] [CrossRef]
- Pfefferbaum, A.; Horvath, T.B.; Roth, W.T.; Clifford, S.T.; Kopell, B.S. Acute and chronic effects of ethanol on event-related potentials. Adv. Exp. Med. Biol. 1980, 126, 625–639. [Google Scholar] [CrossRef]
- Herning, R.I.; Jones, R.T.; Hooker, W.D.; Tulunay, F.C. Information processing components of the auditory event related potential are reduced by cocaine. Psychopharmacology 1985, 87, 178–185. [Google Scholar] [CrossRef]
- Bauer, L.O.; Hesselbrock, V.M. P300 decrements in teenagers with conduct problems: Implications for substance abuse risk and brain development. Biol. Psychiatry 1999, 46, 263–272. [Google Scholar] [CrossRef]
- Bauer, L.O.; Hesselbrock, V.M. Signal in the noise: Altered brain activation among adolescent alcohol users detected via the analysis of intra-individual variability(1). Psychopharmacology 2022, 239, 3595–3604. [Google Scholar] [CrossRef] [PubMed]
- Iacono, W.G.; Carlson, S.R.; Malone, S.M.; McGue, M. P3 event-related potential amplitude and the risk for disinhibitory disorders in adolescent boys. Arch. Gen. Psychiatry 2002, 59, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Porjesz, B.; Rangaswamy, M. Neurophysiological endophenotypes, CNS disinhibition, and risk for alcohol dependence and related disorders. ScientificWorldJournal 2007, 7, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Ritter, W.; Ford, J.M.; Gaillard, A.W.; Harter, M.R.; Kutas, M.; Näätänen, R.; Polich, J.; Renault, B.; Rohrbaugh, J. Cognition and event-related potentials. I. The relation of negative potentials and cognitive processes. Ann. N. Y. Acad. Sci. 1984, 425, 24–38. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kamarajan, C.; Tang, Y.; Chorlian, D.B.; Roopesh, B.N.; Manz, N.; Stimus, A.; Rangaswamy, M.; Porjesz, B. Neurocognitive deficits in male alcoholics: An ERP/sLORETA analysis of the N2 component in an equal probability Go/NoGo task. Biol. Psychol. 2012, 89, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Olbrich, H.M.; Maes, H.; Gann, H.; Hagenbuch, F.; Feige, B. Auditory and visual event-related potentials in alcoholics: Abnormalities of components and brain electrical field. Eur. Arch. Psychiatry Clin. Neurosci. 2000, 250, 215–220. [Google Scholar] [CrossRef]
- Bauer, L.O.; Hesselbrock, V.M. Brain maturation and subtypes of conduct disorder: Interactive effects on p300 amplitude and topography in male adolescents. J. Am. Acad. Child Adolesc. Psychiatry 2003, 42, 106–115. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kamarajan, C.; Manz, N.; Chorlian, D.B.; Stimus, A.; Porjesz, B. Delta, theta, and alpha event-related oscillations in alcoholics during Go/NoGo task: Neurocognitive deficits in execution, inhibition, and attention processing. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 65, 158–171. [Google Scholar] [CrossRef]
- Nash, M. The Effect of Menstrual Phases on Visual Attention and Event-Related Potentials. Master’s Thesis, Brigham Young University, Provo, UT, USA, 2009. [Google Scholar]
- Wolf, M.M. Association between Academic Performance and Electro-Cortical Processing of Cognitive Stimuli in College Students. Master’s Thesis, Brigham Young University, Provo, UT, USA, 2011. [Google Scholar]
- Matthews, S.; Dwyer, R.; Snoek, A. Stigma and Self-Stigma in Addiction. J. Bioeth. Inq. 2017, 14, 275–286. [Google Scholar] [CrossRef]
- Nyblade, L.; Stockton, M.A.; Giger, K.; Bond, V.; Ekstrand, M.L.; Lean, R.M.; Mitchell, E.M.H.; Nelson, R.E.; Sapag, J.C.; Siraprapasiri, T.; et al. Stigma in health facilities: Why it matters and how we can change it. BMC Med. 2019, 17, 25. [Google Scholar] [CrossRef]
- Smookler, H.H.; Buckley, J.P. Effect of drugs on animals exposed to chronic environmental stress. Fed. Proc. 1970, 29, 1980–1984. [Google Scholar] [PubMed]
- Gonzalez, G.; Oliveto, A.; Kosten, T.R. Combating opiate dependence: A comparison among the available pharmacological options. Expert Opin. Pharmacother. 2004, 5, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.D.; McKenzie, M.; Shield, D.C.; Wolf, F.A.; Key, R.G.; Poshkus, M.; Clarke, J. Linkage with methadone treatment upon release from incarceration: A promising opportunity. J. Addict. Dis. 2005, 24, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Coviello, D.M.; Alterman, A.I.; Cacciola, J.S.; Rutherford, M.J.; Zanis, D.A. The role of family history in addiction severity and treatment response. J. Subst. Abuse Treat. 2004, 26, 303–313. [Google Scholar] [CrossRef]
- Sewards, T.V.; Sewards, M.A. Alpha-band oscillations in visual cortex: Part of the neural correlate of visual awareness? Int. J. Psychophysiol. 1999, 32, 35–45. [Google Scholar] [CrossRef]
- Ro, T. Alpha oscillations and feedback processing in visual cortex for conscious perception. J. Cogn. Neurosci. 2019, 31, 948–960. [Google Scholar] [CrossRef]
- Romei, V.; Brodbeck, V.; Michel, C.; Amedi, A.; Pascual-Leone, A.; Thut, G. Spontaneous fluctuations in posterior alpha-band EEG activity reflect variability in excitability of human visual areas. Cereb. Cortex 2008, 18, 2010–2018. [Google Scholar] [CrossRef]
- Başar, E. Brain oscillations in neuropsychiatric disease. Dialogues Clin. Neurosci. 2013, 15, 291–300. [Google Scholar] [CrossRef]
- van Diepen, R.M.; Miller, L.M.; Mazaheri, A.; Geng, J.J. The role of alpha activity in spatial and feature-based attention. eNeuro 2016, 3, ENEURO.0204-16.2016. [Google Scholar] [CrossRef]
- Herring, J.D.; Thut, G.; Jensen, O.; Bergmann, T.O. Attention modulates TMS-locked alpha oscillations in the visual cortex. J. Neurosci. 2015, 35, 14435–14447. [Google Scholar] [CrossRef]
- Miyauchi, E.; Kawasaki, M. Electroencephalogram oscillations support the involvement of task-unrelated thoughts in the mechanism of boredom: A pilot study. Neurosci. Lett. 2018, 677, 72–77. [Google Scholar] [CrossRef]
- Foxe, J.J.; Simpson, G.V.; Ahlfors, S.P. Parieto-occipital approximately 10 Hz activity reflects anticipatory state of visual attention mechanisms. Neuroreport 1998, 9, 3929–3933. [Google Scholar] [CrossRef]
- Mathewson, K.E.; Gratton, G.; Fabiani, M.; Beck, D.M.; Ro, T. To see or not to see: Prestimulus alpha phase predicts visual awareness. J. Neurosci. 2009, 29, 2725–2732. [Google Scholar] [CrossRef]
- Newson, J.J.; Thiagarajan, T.C. EEG frequency bands in psychiatric disorders: A review of resting state studies. Front. Hum. Neurosci. 2018, 12, 521. [Google Scholar] [CrossRef]
- Cona, G.; Chiossi, F.; Di Tomasso, S.; Pellegrino, G.; Piccione, F.; Bisiacchi, P.; Arcara, G. Theta and alpha oscillations as signatures of internal and external attention to delayed intentions: A magnetoencephalography (MEG) study. Neuroimage 2020, 205, 116295. [Google Scholar] [CrossRef]
- Deiber, M.P.; Hasler, R.; Colin, J.; Dayer, A.; Aubry, J.M.; Baggio, S.; Perroud, N.; Ros, T. Linking alpha oscillations, attention and inhibitory control in adult ADHD with EEG neurofeedback. Neuroimage Clin. 2020, 25, 102145. [Google Scholar] [CrossRef]
- Fingelkurts, A.; Kahkonen, S.; Fingelkurts, A.; Kivisaari, R.; Borisov, S.; Puuskari, V.; Jokela, O.; Autti, T. Reorganization of the composition of brain oscillations and their temporal characteristics during opioid withdrawal. J. Psychopharmacol. 2008, 22, 270–284. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kivisaari, R.; Autti, T.; Borisov, S.; Puuskari, V.; Jokela, O.; Kahkonen, S. Increased local and decreased remote functional connectivity at EEG alpha and beta frequency bands in opioid-dependent patients. Psychopharmacology 2006, 188, 42–52. [Google Scholar] [CrossRef]
- Forschack, N.; Nierhaus, T.; Müller, M.M.; Villringer, A. Alpha-band brain oscillations shape the processing of perceptible as well as imperceptible somatosensory stimuli during selective attention. J. Neurosci. 2017, 37, 6983–6994. [Google Scholar] [CrossRef]
- Foster, J.J.; Awh, E. The role of alpha oscillations in spatial attention: Limited evidence for a suppression account. Curr. Opin. Psychol. 2019, 29, 34–40. [Google Scholar] [CrossRef]
- Gardner, E.L. Addiction and brain reward and antireward pathways. Adv. Psychosom. Med. 2011, 30, 22–60. [Google Scholar] [CrossRef]
- Ikkai, A.; Dandekar, S.; Curtis, C.E. Lateralization in alpha-band oscillations predicts the locus and spatial distribution of attention. PLoS ONE 2016, 11, e0154796. [Google Scholar] [CrossRef]
- Koob, G.F. A role for brain stress systems in addiction. Neuron 2008, 59, 11–34. [Google Scholar] [CrossRef]
- Magosso, E.; De Crescenzio, F.; Ricci, G.; Piastra, S.; Ursino, M. EEG alpha power is modulated by attentional changes during cognitive tasks and virtual reality immersion. Comput. Intell. Neurosci. 2019, 2019, 7051079. [Google Scholar] [CrossRef]
- Mangun, G.R. Neural mechanisms of visual selective attention. Psychophysiology 1995, 32, 4–18. [Google Scholar] [CrossRef]
- Mo, J.; Liu, Y.; Huang, H.; Ding, M. Coupling between visual alpha oscillations and default mode activity. Neuroimage 2013, 68, 112–118. [Google Scholar] [CrossRef]
- Motlagh, F.; Ibrahim, F.; Menke, J.M.; Rashid, R.; Seghatoleslam, T.; Habil, H. Neuroelectrophysiological approaches in heroin addiction research: A review of literatures. J. Neurosci. Res. 2016, 94, 297–309. [Google Scholar] [CrossRef]
- Samaha, J.; Postle, B.R. The speed of alpha-band oscillations predicts the temporal resolution of visual perception. Curr. Biol. 2015, 25, 2985–2990. [Google Scholar] [CrossRef]
- Samaha, J.; Sprague, T.C.; Postle, B.R. Decoding and Reconstructing the Focus of Spatial Attention from the Topography of Alpha-band Oscillations. J. Cogn. Neurosci. 2016, 28, 1090–1097. [Google Scholar] [CrossRef]
- White, F.J.; Kalivas, P.W. Neuroadaptations involved in amphetamine and cocaine addiction. Drug Alcohol Depend. 1998, 51, 141–153. [Google Scholar] [CrossRef]
- Wise, R.A.; Hoffman, D.C. Localization of drug reward mechanisms by intracranial injections. Synapse 1992, 10, 247–263. [Google Scholar] [CrossRef]
- Buckley, D.I.; Calvert, J.F.; Lapidus, J.A.; Morris, C.D. Chronic opioid therapy and preventive services in rural primary care: An Oregon rural practice-based research network study. Ann. Fam. Med. 2010, 8, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Chou, R.; Ballantyne, J.C.; Fanciullo, G.J.; Fine, P.G.; Miaskowski, C. Research gaps on use of opioids for chronic noncancer pain: Findings from a review of the evidence for an American Pain Society and American Academy of Pain Medicine clinical practice guideline. J. Pain 2009, 10, 147–159. [Google Scholar] [CrossRef]
- Fishbein, D.H.; Krupitsky, E.; Flannery, B.A.; Langevin, D.J.; Bobashev, G.; Verbitskaya, E.; Augustine, C.B.; Bolla, K.I.; Zvartau, E.; Schech, B.; et al. Neurocognitive characterizations of Russian heroin addicts without a significant history of other drug use. Drug Alcohol Depend. 2007, 90, 25–38. [Google Scholar] [CrossRef]
- Webster, L.R.; Webster, R.M. Predicting aberrant behaviors in opioid-treated patients: Preliminary validation of the Opioid Risk Tool. Pain Med. 2005, 6, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Pascoli, V.; Turiault, M.; Luscher, C. Reversal of cocaine-evoked synaptic potentiation resets drug-induced adaptive behaviour. Nature 2012, 481, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.E. Neuroscience: Behavioural effects of cocaine reversed. Nature 2012, 481, 36–37. [Google Scholar] [CrossRef]
- Chauvet, C.; Lardeux, V.; Jaber, M.; Solinas, M. Brain regions associated with the reversal of cocaine conditioned place preference by environmental enrichment. Neuroscience 2011, 184, 88–96. [Google Scholar] [CrossRef]
- Gazzaniga, M.S. Who’s in Charge?: Free Will and the Science of the Brain; Ecco: New York, NY, USA, 2011; p. 272. [Google Scholar]
- Petrie, J. Visual Attention, Color Processing and Physiological Measure Differences in Males and Females with Substance Abuse and Opiate Addiction. Ph.D. Thesis, Brigham Young University, Provo, UT, USA, 2012. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Univariate Test | ||||||
| Gender Effects | OUD Effects within Gender | |||||
| (Fixed) | (Random) | |||||
| Variable | F(1, 34) | p | η2 | F(2, 34) | p | η2 |
| P300 | ||||||
| Amplitude | 25.16 | <0.0001 *** | 0.41 | 1 | 0.3788 | 0.033 |
| Latency | 3.52 | 0.0692 | 0.049 | 16.63 | <0.0001 *** | 0.462 |
| N200 | ||||||
| Amplitude | 14.5 | 0.0006 *** | 0.289 | 0.94 | 0.4018 | 0.037 |
| Latency | 1.06 | 0.3105 | 0.019 | 9.84 | 0.0004 * | 0.356 |
| Wavelet Power | ||||||
| Alpha | 3.83 | 0.0585 | 0.095 | 1.21 | 0.3111 | 0.06 |
| Beta | 4.22 | 0.0477 *** | 0.071 | 3.76 | 0.0333 * | 0.165 |
| B. Multivariate Tests | ||||||
| Gender Effects (Fixed) | Opioid Effects within Gender ‡ (Random) | |||||
| † Variable and Test | Value | p | Value | p | ||
| P300 | ||||||
| Wilks’ lambda | 0.573515 | 0.0001 *** | 0.469855 | <0.0001 *** | ||
| Pillai’s trace | 0.426485 | 0.0001 *** | 0.554621 | 0.0002 *** | ||
| Hotelling-Lawley Trace | 0.743632 | 0.0001 *** | 1.076223 | <0.0001 *** | ||
| Roy’s Greatest Root | 0.743632 | 0.0001 *** | 1.025422 | <0.0001 *** | ||
| N200 | ||||||
| Wilks’ lambda | 0.69395 | 0.0024 *** | 0.598011 | 0.0018 *** | ||
| Pillai’s trace | 0.30605 | 0.0024 *** | 0.405014 | 0.0036 *** | ||
| Hotelling-Lawley Trace | 0.441026 | 0.0024 *** | 0.667152 | 0.0014 *** | ||
| Roy’s Greatest Root | 0.441026 | 0.0024 *** | 0.659482 | 0.0002 *** | ||
| Wavelet Power | ||||||
| Wilks’ lambda | 0.845376 | 0.0626 | 0.806848 | 0.1263 | ||
| Pillai’s trace | 0.154624 | 0.0626 | 0.193169 | 0.1355 | ||
| Hotelling-Lawley Trace | 0.182905 | 0.0626 | 0.239371 | 0.1206 | ||
| Roy’s Greatest Root | 0.182905 | 0.0626 | 0.239286 | 0.0261 | ||
| Variables | Means (95% CI) | Z-Score Means (95% CI) |
|---|---|---|
| P300 Amplitude | ||
| Female | ||
| OUD | 2.257 (1.670, 2.844) | −0.358 (−0.698, −0.018) |
| Control | 3.148 (2.201, 4.095) | 0.158 (−0.390, 0.706) |
| Male | ||
| OUD | 5.082 (3.749, 6.415) | 1.279 (0.507, 2.051) |
| Control | 5.665 (4.514, 6.816) | 1.616 (0.949, 2.283) |
| P300 Latency | ||
| Female | ||
| OUD | 339.5 (309.8, 369.1) | −0.897 (−1.348, −0.446) |
| Control | 460.7 (425.7, 495.8) | 0.945 (0.413, 1.477) |
| Male | ||
| OUD | 470.4 (434.4, 506.4) | 1.092 (0.545, 1.639) |
| Control | 395.4 (359.2, 431.6) | −0.047 (−0.596, 0.502) |
| N200 Amplitude | ||
| Female | ||
| OUD | 1.750 (1.459, 2.041) | −0.518 (−0.858, −0.178) |
| Control | 1.710 (1.075, 2.345) | −0.553 (−1.101, −0.004) |
| Male | ||
| OUD | 2.620 (1.785, 3.455) | 0.144 (−0.628, 0.917) |
| Control | 3.240 (2.664, 3.816) | 0.606 (−0.061, 1.273) |
| N200 Latency | ||
| Female | ||
| OUD | 274.0 (252.7, 295.3) | −0.660 (−1.110, −0.209) |
| Control | 304.7 (285.6, 323.8) | 0.124 (−0.408, 0.656) |
| Male | ||
| OUD | 329.5 (305.1, 353.8) | 0.755 (0.208, 1.302) |
| Control | 271.0 (254.5, 287.5) | −0.737 (−1.286, −0.189) |
| Alpha Wavelet Power | ||
| Female | ||
| OUD | 0.1107 (0.0651, 0.1563) | −0.795 (−1.278, −0.312) |
| Control | 0.1498 (0.0918, 0.2078) | −0.381 (−0.995, 0.233) |
| Male | ||
| OUD | 0.1624 (0.1994, 0.2255) | −0.248 (−0.916, 0.420) |
| Control | 0.2155 (0.1483, 0.2826) | 0.314 (−0.397, 1.025) |
| Beta Wavelet Power | ||
| Female | ||
| OUD | 0.1421 (0.1154, 0.1687) | −0.219 (−0.748, 0.310) |
| Control | 0.1769 (0.1458, 0.2080) | 0.472 (−0.146, 1.089) |
| Male | ||
| OUD | 0.1667 (0.1320, 0.2013) | 0.269 (−0.418, 0.957) |
| Control | 0.2175 (0.1863, 0.2487) | 1.227 (0.659, 1.896) |
| N200 Amplitude | P300 Amplitude | N200 Latency | P300 Latency | Alpha | Beta | |
|---|---|---|---|---|---|---|
| N200 amplitude | 1 | |||||
| P300 amplitude | 0.58 | 1 | ||||
| N200 latency | 0.04 | 0.04 | 1 | |||
| P300 latency | 0.15 | 0.37 | 0.54 | 1 | ||
| alpha | 0.03 | 0.01 | 0.27 | 0.03 | 1 | |
| beta | 0.15 | −0.03 | −0.17 | −0.04 | 0.43 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrie, J.; Kowallis, L.R.; Kamhout, S.; Bills, K.B.; Adams, D.; Fleming, D.E.; Brown, B.L.; Steffensen, S.C. Gender-Specific Interactions in a Visual Object Recognition Task in Persons with Opioid Use Disorder. Biomedicines 2023, 11, 2460. https://doi.org/10.3390/biomedicines11092460
Petrie J, Kowallis LR, Kamhout S, Bills KB, Adams D, Fleming DE, Brown BL, Steffensen SC. Gender-Specific Interactions in a Visual Object Recognition Task in Persons with Opioid Use Disorder. Biomedicines. 2023; 11(9):2460. https://doi.org/10.3390/biomedicines11092460
Chicago/Turabian StylePetrie, JoAnn, Logan R. Kowallis, Sarah Kamhout, Kyle B. Bills, Daniel Adams, Donovan E. Fleming, Bruce L. Brown, and Scott C. Steffensen. 2023. "Gender-Specific Interactions in a Visual Object Recognition Task in Persons with Opioid Use Disorder" Biomedicines 11, no. 9: 2460. https://doi.org/10.3390/biomedicines11092460