
Systemic and Airway Epigenetic Disruptions Are Associated with Health Status in COPD
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Differential Effects of Inhaled Symbicort and Advair on Lung Microbiota (DISARM) Study Cohort
2.2. DNA Methylation Profiling
2.3. Differential Methylation Analysis
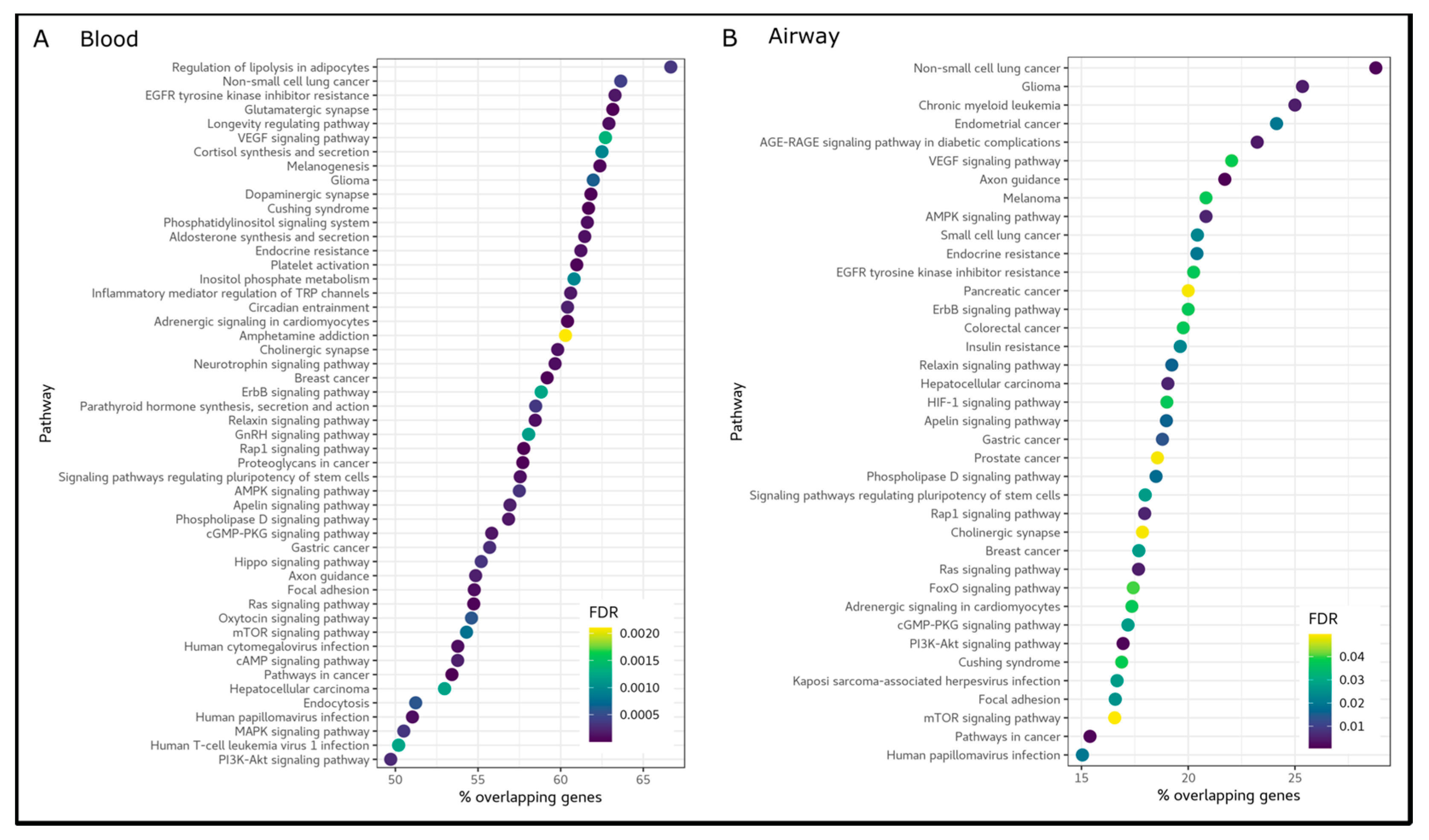
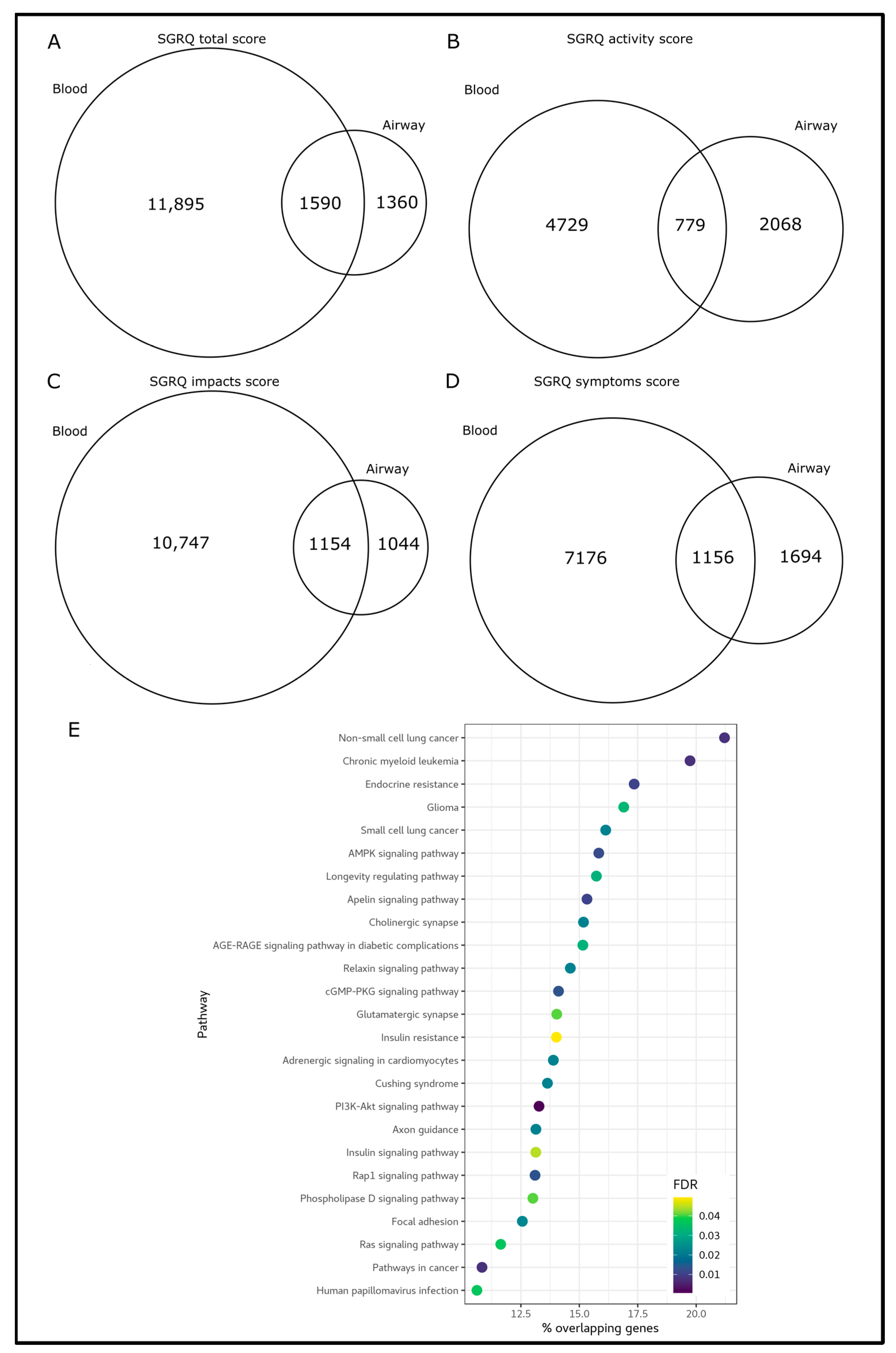
2.4. Pathway Enrichment Analysis
3. Results
3.1. Study Cohort Overview
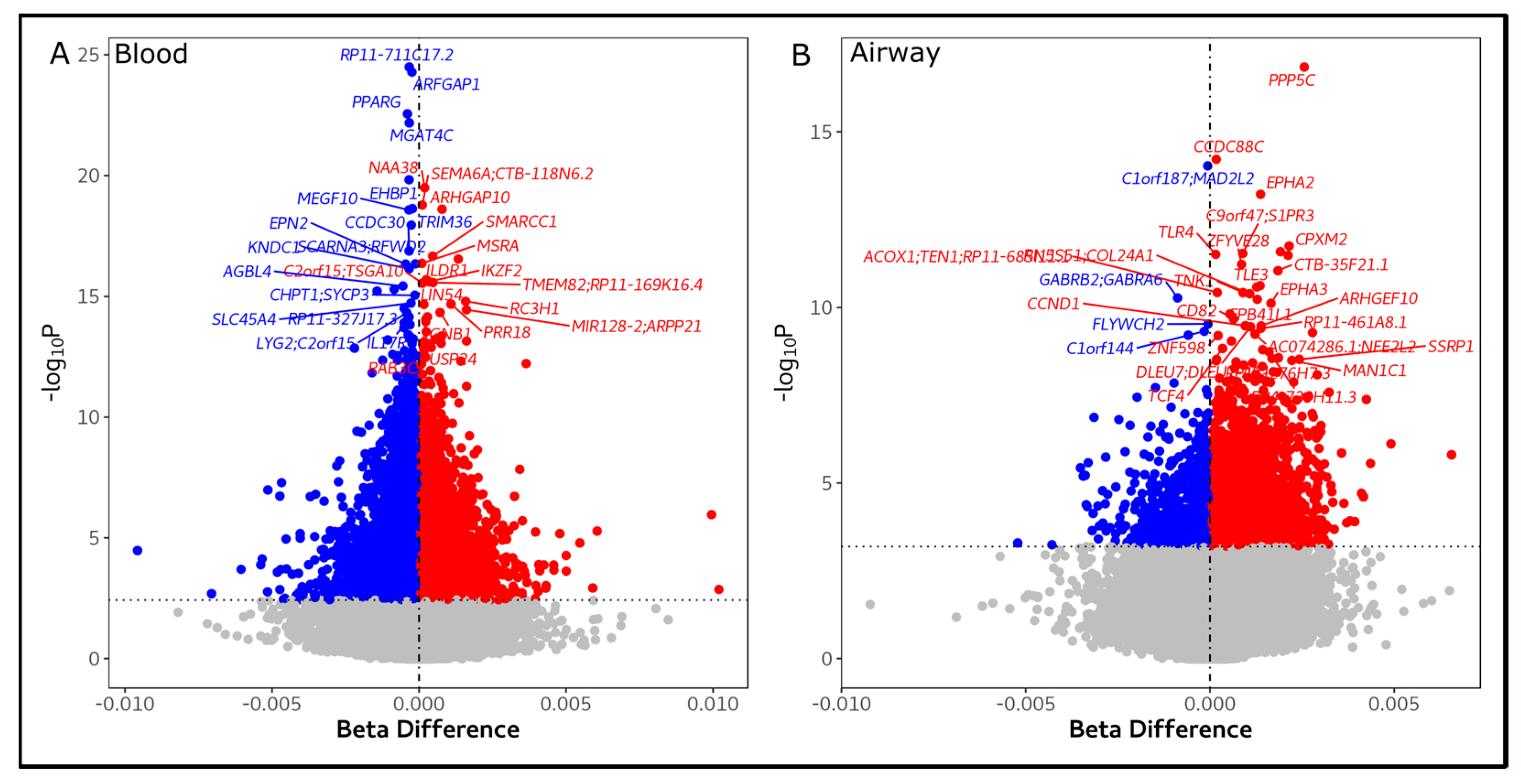
3.2. Blood and Airway Epigenetic Disruptions Are Associated with SGRQ Scores
3.3. A Systemic Epigenetic Signature of Health Status in COPD
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.K.; Raja, A.; Brown, B.D. Chronic Obstructive Pulmonary Disease; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar]
- Stolz, D.; Mkorombindo, T.; Schumann, D.M.; Agusti, A.; Ash, S.Y.; Bafadhel, M.; Bai, C.; Chalmers, J.D.; Criner, G.J.; Dharmage, S.C.; et al. Towards the Elimination of Chronic Obstructive Pulmonary Disease: A Lancet Commission. Lancet 2022, 400, 921–972. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Sin, D.D. GETting to Know the Many Causes and Faces of COPD. Lancet Respir. Med. 2022, 10, 426–428. [Google Scholar] [CrossRef] [PubMed]
- Hernández Cordero, A.I.; Yang, C.X.; Yang, J.; Horvath, S.; Shaipanich, T.; MacIsaac, J.; Lin, D.T.; Kobor, M.S.; Guillemi, S.; Harris, M.; et al. Airway Aging and Methylation Disruptions in HIV-Associated Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2022, 206, 150–160. [Google Scholar] [CrossRef]
- Müllerova, H.; Gelhorn, H.; Wilson, H.; Benson, V.S.; Karlsson, N.; Menjoge, S.; Rennard, S.I.; Tabberer, M.; Tal-Singer, R.; Merrill, D.; et al. St George’s Respiratory Questionnaire Score Predicts Outcomes in Patients with COPD: Analysis of Individual Patient Data in the COPD Biomarkers Qualification Consortium Database. Chronic Obstr. Pulm. Dis. 2017, 4, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Weatherall, M.; Marsh, S.; Shirtcliffe, P.; Williams, M.; Travers, J.; Beasley, R. Quality of Life Measured by the St George’s Respiratory Questionnaire and Spirometry. Eur. Respir. J. 2009, 33, 1025–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.W. St. George’s Respiratory Questionnaire: MCID. COPD 2005, 2, 75–79. [Google Scholar] [CrossRef]
- Yip, W.; Li, X.; Koelwyn, G.J.; Milne, S.; Leitao Filho, F.S.; Yang, C.X.; Hernández Cordero, A.I.; Yang, J.; Yang, C.W.T.; Shaipanich, T.; et al. Inhaled Corticosteroids Selectively Alter the Microbiome and Host Transcriptome in the Small Airways of Patients with Chronic Obstructive Pulmonary Disease. Biomedicines 2022, 10, 1110. [Google Scholar] [CrossRef]
- Leitao Filho, F.S.; Takiguchi, H.; Akata, K.; Ra, S.W.; Moon, J.-Y.; Kim, H.K.; Cho, Y.; Yamasaki, K.; Milne, S.; Yang, J.; et al. Effects of Inhaled Corticosteroid/Long-Acting Β2-Agonist Combination on the Airway Microbiome of Patients with Chronic Obstructive Pulmonary Disease: A Randomized Controlled Clinical Trial (DISARM). Am. J. Respir. Crit. Care Med. 2021, 204, 1143–1152. [Google Scholar] [CrossRef]
- Milne, S.; Li, X.; Yang, C.X.; Leitao Filho, F.S.; Hernández Cordero, A.I.; Yang, C.W.T.; Shaipanich, T.; van Eeden, S.F.; Leung, J.M.; Lam, S.; et al. Inhaled Corticosteroids Downregulate SARS-CoV-2-Related Genes in COPD: Results from a Randomised Controlled Trial. Eur. Respir. J. 2021, 58, 2100130. [Google Scholar] [CrossRef]
- Ho, C.G.; Milne, S.; Li, X.; Yang, C.X.; Leitao Filho, F.S.; Cheung, C.Y.; Yang, J.S.W.; Hernández Cordero, A.I.; Yang, C.W.T.; Shaipanich, T.; et al. Airway Eosinophilia on Bronchoalveolar Lavage and the Risk of Exacerbations in COPD. Biomedicines 2022, 10, 1412. [Google Scholar] [CrossRef]
- Graham, B.L.; Steenbruggen, I.; Miller, M.R.; Barjaktarevic, I.Z.; Cooper, B.G.; Hall, G.L.; Hallstrand, T.S.; Kaminsky, D.A.; McCarthy, K.; McCormack, M.C.; et al. Standardization of Spirometry 2019 Update. An Official American Thoracic Society and European Respiratory Society Technical Statement. Am. J. Respir. Crit. Care Med. 2019, 200, e70–e88. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.X.; Schon, E.; Obeidat, M.; Kobor, M.S.; McEwen, L.; MacIsaac, J.; Lin, D.; Novak, R.M.; Hudson, F.; Klinker, H.; et al. Occurrence of Accelerated Epigenetic Aging and Methylation Disruptions in Human Immunodeficiency Virus Infection Before Antiretroviral Therapy. J. Infect. Dis. 2021, 223, 1681–1689. [Google Scholar] [CrossRef] [PubMed]
- Hernandez Cordero, A.I.; Yang, C.X.; Obeidat, M.; Yang, J.; MacIsaac, J.; McEwen, L.; Lin, D.; Kobor, M.; Novak, R.; Hudson, F.; et al. DNA Methylation Is Associated with Airflow Obstruction in Patients Living with HIV. Thorax 2021, 76, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Hernandez Cordero, A.I.; Yang, C.X.; Milne, S.; Li, X.; Hollander, Z.; Chen, V.; Ng, R.; Tebbutt, S.J.; Leung, J.M.; Sin, D.D. Epigenetic Blood Biomarkers of Ageing and Mortality in COPD. Eur. Respir. J. 2021, 58, 2101890. [Google Scholar] [CrossRef]
- Hernandez Cordero, A.I.; Yang, C.X.; Li, X.; Milne, S.; Chen, V.; Hollander, Z.; Ng, R.; Criner, G.; Woodruff, P.; Lazarus, S.; et al. Epigenetic Marker of Telomeric Age Is Associated with Exacerbations and Hospitalizations in Chronic Obstructive Pulmonary Disease. Respir. Res. 2021, 22, 316. [Google Scholar] [CrossRef]
- Triche, T.J.; Weisenberger, D.J.; Van Den Berg, D.; Laird, P.W.; Siegmund, K.D. Low-Level Processing of Illumina Infinium DNA Methylation BeadArrays. Nucleic Acids Res. 2013, 41, e90. [Google Scholar] [CrossRef] [Green Version]
- Teschendorff, A.E.; Marabita, F.; Lechner, M.; Bartlett, T.; Tegner, J.; Gomez-Cabrero, D.; Beck, S. A Beta-Mixture Quantile Normalization Method for Correcting Probe Design Bias in Illumina Infinium 450 k DNA Methylation Data. Bioinformatics 2013, 29, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting Batch Effects in Microarray Expression Data Using Empirical Bayes Methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef]
- Rahmani, E.; Shenhav, L.; Schweiger, R.; Yousefi, P.; Huen, K.; Eskenazi, B.; Eng, C.; Huntsman, S.; Hu, D.; Galanter, J.; et al. Genome-Wide Methylation Data Mirror Ancestry Information. Epigenetics Chromatin 2017, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA Methylation Arrays as Surrogate Measures of Cell Mixture Distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S. In Statistics and Computing, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95457-8. [Google Scholar]
- Lee, M.K.; Hong, Y.; Kim, S.-Y.; Kim, W.J.; London, S.J. Epigenome-Wide Association Study of Chronic Obstructive Pulmonary Disease and Lung Function in Koreans. Epigenomics 2017, 9, 971–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, T.J.; Buckley, M.J.; Statham, A.L.; Pidsley, R.; Samaras, K.; Lord, R.V.; Clark, S.J.; Molloy, P.L. De Novo Identification of Differentially Methylated Regions in the Human Genome. Epigenetics Chromatin 2015, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Agusti, A.; Anzueto, A.; Barnes, P.J.; Bourbeau, J.; Celli, B.R.; Criner, G.J.; Frith, P.; Halpin, D.M.G.; Han, M.; et al. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease: The GOLD Science Committee Report 2019. Eur. Respir. J. 2019, 53, 1900164. [Google Scholar] [CrossRef] [PubMed]
- Sin, D.D.; Anthonisen, N.R.; Soriano, J.B.; Agusti, A.G. Mortality in COPD: Role of Comorbidities. Eur. Respir. J. 2006, 28, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Benayoun, L.; Letuve, S.; Druilhe, A.; Boczkowski, J.; Dombret, M.C.; Mechighel, P.; Megret, J.; Leseche, G.; Aubier, M.; Pretolani, M. Regulation of Peroxisome Proliferator-Activated Receptor Gamma Expression in Human Asthmatic Airways: Relationship with Proliferation, Apoptosis, and Airway Remodeling. Am. J. Respir. Crit. Care Med. 2001, 164, 1487–1494. [Google Scholar] [CrossRef]
- Wang, A.C.; Dai, X.; Luu, B.; Conrad, D.J. Peroxisome Proliferator-Activated Receptor-Gamma Regulates Airway Epithelial Cell Activation. Am. J. Respir. Cell Mol. Biol. 2001, 24, 688–693. [Google Scholar] [CrossRef]
- Calnek, D.S.; Mazzella, L.; Roser, S.; Roman, J.; Hart, C.M. Peroxisome Proliferator-Activated Receptor Gamma Ligands Increase Release of Nitric Oxide from Endothelial Cells. Arterioscler Thromb. Vasc. Biol. 2003, 23, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Patel, H.J.; Belvisi, M.G.; Bishop-Bailey, D.; Yacoub, M.H.; Mitchell, J.A. Activation of Peroxisome Proliferator-Activated Receptors in Human Airway Smooth Muscle Cells Has a Superior Anti-Inflammatory Profile to Corticosteroids: Relevance for Chronic Obstructive Pulmonary Disease Therapy. J. Immunol. 2003, 170, 2663–2669. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.T.; Lakshmi, S.P.; Kleinhenz, J.M.; Sutliff, R.L.; Hart, C.M.; Reddy, R.C. Endothelial Cell Peroxisome Proliferator-Activated Receptor γ Reduces Endotoxemic Pulmonary Inflammation and Injury. J. Immunol. 2012, 189, 5411–5420. [Google Scholar] [CrossRef] [Green Version]
- Chinetti, G.; Griglio, S.; Antonucci, M.; Torra, I.P.; Delerive, P.; Majd, Z.; Fruchart, J.C.; Chapman, J.; Najib, J.; Staels, B. Activation of Proliferator-Activated Receptors Alpha and Gamma Induces Apoptosis of Human Monocyte-Derived Macrophages. J. Biol. Chem. 1998, 273, 25573–25580. [Google Scholar] [CrossRef]
- Belvisi, M.G.; Hele, D.J. Peroxisome Proliferator-Activated Receptors as Novel Targets in Lung Disease. Chest 2008, 134, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; de Heer, H.J.; Soullié, T.; Angeli, V.; Trottein, F.; Hoogsteden, H.C.; Lambrecht, B.N. Activation of Peroxisome Proliferator-Activated Receptor-Gamma in Dendritic Cells Inhibits the Development of Eosinophilic Airway Inflammation in a Mouse Model of Asthma. Am. J. Pathol. 2004, 164, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Michael, L.F.; Lazar, M.A.; Mendelson, C.R. Peroxisome Proliferator-Activated Receptor Gamma1 Expression Is Induced during Cyclic Adenosine Monophosphate-Stimulated Differentiation of Alveolar Type II Pneumonocytes. Endocrinology 1997, 138, 3695–3703. [Google Scholar] [CrossRef] [PubMed]
- Solleti, S.K.; Simon, D.M.; Srisuma, S.; Arikan, M.C.; Bhattacharya, S.; Rangasamy, T.; Bijli, K.M.; Rahman, A.; Crossno, J.T.; Shapiro, S.D.; et al. Airway Epithelial Cell PPARγ Modulates Cigarette Smoke-Induced Chemokine Expression and Emphysema Susceptibility in Mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L293–L304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, S.F.P.; Connett, J.E.; Anthonisen, N.R.; Wise, R.A.; Tashkin, D.P.; Sin, D.D. C-Reactive Protein and Mortality in Mild to Moderate Chronic Obstructive Pulmonary Disease. Thorax 2006, 61, 849–853. [Google Scholar] [CrossRef] [Green Version]
- Hogg, J.C.; Chu, F.; Utokaparch, S.; Woods, R.; Elliott, W.M.; Buzatu, L.; Cherniack, R.M.; Rogers, R.M.; Sciurba, F.C.; Coxson, H.O.; et al. The Nature of Small-Airway Obstruction in Chronic Obstructive Pulmonary Disease. N. Engl. J. Med. 2004, 350, 2645–2653. [Google Scholar] [CrossRef]
- Kaur, G.; Batra, S. Regulation of DNA Methylation Signatures on NF-ΚB and STAT3 Pathway Genes and TET Activity in Cigarette Smoke Extract-Challenged Cells/COPD Exacerbation Model in Vitro. Cell Biol. Toxicol. 2020, 36, 459–480. [Google Scholar] [CrossRef]
- Papadopoli, D.; Boulay, K.; Kazak, L.; Pollak, M.; Mallette, F.A.; Topisirovic, I.; Hulea, L. MTOR as a Central Regulator of Lifespan and Aging. F1000Research 2019, 8, 998. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, Y.; Hsieh, T.; Wang, C.; Cheng, K.; Wang, L.; Lin, T.; Cheung, C.H.A.; Wu, C.; Chiang, H. Aging-induced Akt Activation Involves in Aging-related Pathologies and Aβ-induced Toxicity. Aging Cell 2019, 18, e12989. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Wasswa-Kintu, S.; Gan, W.Q.; Man, S.F.P.; Pare, P.D.; Sin, D.D. Relationship between Reduced Forced Expiratory Volume in One Second and the Risk of Lung Cancer: A Systematic Review and Meta-Analysis. Thorax 2005, 60, 570–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinta, S.J.; Andersen, J.K. Dopaminergic Neurons. Int. J. Biochem. Cell Biol. 2005, 37, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Aubier, M.; Murciano, D.; Menu, Y.; Boczkowski, J.; Mal, H.; Pariente, R. Dopamine Effects on Diaphragmatic Strength during Acute Respiratory Failure in Chronic Obstructive Pulmonary Disease. Ann. Intern. Med. 1989, 110, 17–23. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Tracey, K.J. The Cholinergic Anti-Inflammatory Pathway. Brain Behav. Immun. 2005, 19, 493–499. [Google Scholar] [CrossRef]
- Maurer, S.V.; Williams, C.L. The Cholinergic System Modulates Memory and Hippocampal Plasticity via Its Interactions with Non-Neuronal Cells. Front. Immunol. 2017, 8, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, A.E.; Irizarry, R.A. Accounting for Cellular Heterogeneity Is Critical in Epigenome-Wide Association Studies. Genome Biol. 2014, 15, R31. [Google Scholar] [CrossRef] [PubMed]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and Allergy: From Basic Mechanisms to Clinical Applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| DISARM Study Cohort | |
|---|---|
| n | 64 |
| Age, years | 64 ± 8 |
| Female, % | 17 |
| BMI, kg/m2 | 24.58 (21.09–29.35) |
| Smoking status | |
| Current, % | 45 |
| Former, % | 55 |
| Pack per years | 48.00 (33.00–59.50) |
| SGRQ total score | 44.06 (33.08–54.90) |
| SGRQ activity score | 65.89 (48.52–74.08) |
| SGRQ impacts score | 28.45 (18.10–39.68) |
| SGRQ symptoms score | 56.20 (39.30–70.63) |
| FEV1% of predicted | 55.00 (43.65–67.25) |
| FVC% of predicted | 83.35 (71.90–92.25) |
| FEV1/FVC, percent | 52.64 (42.93–60.09) |
| SGRQ Score | Tissue | Probe | Beta Difference | p | FDR | Chr | Relation to Island | Position in Relation to Gene | Gene Symbol |
|---|---|---|---|---|---|---|---|---|---|
| Total | Blood | cg00542760 | −0.0002 | 5.22 × 10−25 | 2.06 × 10−19 | 20 | Island | 5’UTR; 1stExon; TSS1500; 3’UTR | ARFGAP1 |
| cg02344187 | −0.0003 | 3.21 × 10−25 | 2.06 × 10−19 | 12 | Open Sea | 5’UTR | RP11-711C17.2 | ||
| cg25911248 | −0.0004 | 2.78 × 10−23 | 7.32 × 10−18 | 3 | Open Sea | 3’UTR | PPARG | ||
| cg00151915 | −0.0003 | 6.59 × 10−23 | 1.30 × 10−17 | 12 | Open Sea | 5’UTR | MGAT4C | ||
| cg02213440 | −0.0003 | 1.49 × 10−20 | 2.35 × 10−15 | 7 | Open Sea | ||||
| Airway | cg16929656 | 0.0026 | 1.44 × 10−17 | 1.13 × 10−11 | 19 | Open Sea | 3’UTR | PPP5C | |
| cg05245430 | 0.0002 | 6.08 × 10−15 | 2.40 × 10−09 | 14 | Island | 3’UTR; 1stExon | CCDC88C | ||
| cg21153875 | −0.0001 | 9.40 × 10−15 | 2.47 × 10−09 | 1 | Island | TSS200 | C1orf187; MAD2L2 | ||
| cg15346134 | 0.0014 | 6.01 × 10−14 | 1.18 × 10−08 | 1 | Open Sea | TSS200 | EPHA2 | ||
| cg15550234 | 0.0021 | 1.78 × 10−12 | 2.81 × 10−07 | 10 | Open Sea | 3’UTR; 5’UTR | CPXM2 | ||
| Activity | Blood | cg09711814 | −0.0001 | 4.35 × 10−22 | 3.43 × 10−16 | 7 | Open Sea | ||
| cg24639069 | −0.0002 | 1.76 × 10−18 | 6.94 × 10−13 | 1 | Open Sea | TSS1500; 5’UTR | CCDC30 | ||
| cg10677105 | −0.0005 | 6.97 × 10−15 | 1.83 × 10−09 | 5 | Open Sea | 5’UTR; 3’UTR | DOCK2 | ||
| cg11893552 | 0.0015 | 1.86 × 10−14 | 3.66 × 10−09 | 6 | Open Sea | ||||
| cg00371195 | −0.0001 | 3.40 × 10−14 | 4.47 × 10−09 | 11 | Open Sea | TSS1500 | F2 | ||
| Airway | cg00278597 | 0.0002 | 4.35 × 10−12 | 1.71 × 10−06 | 8 | Open Sea | TSS1500 | RP11-1057B8.2 | |
| cg09397653 | 0.0009 | 2.60 × 10−12 | 1.71 × 10−06 | 15 | Open Sea | TSS1500 | ITGA11 | ||
| cg27547307 | 0.0005 | 1.11 × 10−11 | 2.91 × 10−06 | 17 | Open Sea | TSS1500; 5’UTR | CYTH1 | ||
| cg00413620 | 0.0011 | 1.74 × 10−11 | 3.43 × 10−06 | 1 | Open Sea | ||||
| cg04926227 | 0.0011 | 4.02 × 10−11 | 6.33 × 10−06 | 8 | Open Sea | TSS1500; 3’UTR; 5’UTR | RP11-463D19.1; STAU2 | ||
| Impact | Blood | cg23444468 | 0.0004 | 6.72 × 10−30 | 5.29 × 10−24 | 3 | Island | TSS1500; 5’UTR | ERC2 |
| cg13886298 | −0.0002 | 1.45 × 10−24 | 5.70 × 10−19 | 5 | Open Sea | TSS1500; 3’UTR | AC004041.2; RAD50 | ||
| cg15751204 | 0.0008 | 5.45 × 10−22 | 1.43 × 10−16 | 3 | Open Sea | ||||
| cg00851837 | −0.0003 | 4.31 × 10−20 | 8.48 × 10−15 | 14 | Open Sea | TSS200 | AP4S1 | ||
| cg15534855 | 0.0005 | 1.24 × 10−19 | 1.95 × 10−14 | 18 | Island | ||||
| Airway | cg08738303 | 0.0005 | 3.69 × 10−13 | 2.91 × 10−07 | 18 | Open Sea | |||
| cg01585096 | 0.0012 | 3.64 × 10−12 | 7.16 × 10−07 | 19 | Open Sea | TSS200; 3’UTR | KRTDAP | ||
| cg03053018 | 0.0030 | 2.11 × 10−12 | 7.16 × 10−07 | 7 | Open Sea | ||||
| cg20447038 | 0.0018 | 3.22 × 10−12 | 7.16 × 10−07 | 6 | Open Sea | ||||
| cg02065293 | 0.0011 | 1.08 × 10−11 | 1.46 × 10−06 | 2 | Open Sea | ||||
| Symptom | Blood | cg06894541 | 0.0003 | 3.10 × 10−19 | 2.44 × 10−13 | 2 | Open Sea | ||
| cg25670076 | 0.0021 | 4.09 × 10−18 | 1.61 × 10−12 | 6 | Open Sea | 5’UTR; 3’UTR | BACH2 | ||
| cg11743078 | −0.0004 | 1.87 × 10−15 | 4.92 × 10−10 | 1 | Open Sea | ||||
| cg02415617 | −0.0004 | 2.08 × 10−14 | 3.25 × 10−09 | 9 | South Shore | 1stExon; 3’UTR; 5’UTR | WNK2 | ||
| cg04028140 | 0.0007 | 2.48 × 10−14 | 3.25 × 10−09 | 11 | Open Sea | ||||
| Airway | cg07380540 | −0.0010 | 6.04 × 10−26 | 4.76 × 10−20 | 1 | South Shelf | |||
| cg10789584 | 0.0005 | 2.76 × 10−17 | 1.09 × 10−11 | 11 | Open Sea | 5’UTR | CD82 | ||
| cg18910215 | −0.0007 | 5.49 × 10−14 | 1.44 × 10−08 | 9 | Open Sea | 5’UTR | MAPKAP1 | ||
| cg20708037 | 0.0018 | 9.37 × 10−14 | 1.85 × 10−08 | 1 | Open Sea | ||||
| cg21088488 | 0.0008 | 4.81 × 10−12 | 7.58 × 10−07 | 7 | South Shore | 3’UTR; TSS1500 | DBNL; PGAM2 |
| SGRQ Score | Tissue | Chr | Start | End | # CpGs | Min FDR | Gene Symbols |
|---|---|---|---|---|---|---|---|
| Total | Blood | 3 | 47,823,638 | 47,825,578 | 7 | 4.71 × 10−27 | SMARCC1 |
| 12 | 124,246,976 | 124,248,926 | 5 | 8.50 × 10−24 | DNAH10 | ||
| 20 | 61,917,085 | 61,918,367 | 5 | 9.22 × 10−23 | ARFGAP1, MIR4326 | ||
| 4 | 148,653,624 | 148,654,701 | 5 | 7.03 × 10−20 | ARHGAP10 | ||
| 5 | 126,779,737 | 126,780,974 | 4 | 1.47 × 10−19 | MEGF10 | ||
| Airway | 18 | 56,296,094 | 56,296,607 | 10 | 2.90 × 10−22 | ALPK2, RPL9P31 | |
| 9 | 91,604,473 | 91,605,695 | 7 | 5.88 × 10−18 | C9orf47, S1PR3 | ||
| 19 | 46,894,811 | 46,895,714 | 3 | 1.41 × 10−16 | AC007193.8 | ||
| 12 | 11,698,534 | 11,699,363 | 5 | 3.03 × 10−14 | RP11-434C1.1, RNU7-60P | ||
| 1 | 120,173,989 | 120,175,029 | 7 | 3.03 × 10−14 | |||
| Activity | Blood | 11 | 2,019,436 | 2,021,103 | 32 | 9.13 × 10−19 | H19 |
| 6 | 168,045,268 | 168,046,457 | 6 | 4.18 × 10−18 | |||
| 3 | 30,936,070 | 30,936,955 | 11 | 4.98 × 10−14 | GADL1 | ||
| 17 | 699,291 | 700,672 | 4 | 5.21 × 10−13 | |||
| 7 | 94,285,270 | 94,287,242 | 60 | 1.11 × 10−12 | SGCE, PEG10 | ||
| Airway | 18 | 56,296,094 | 56,296,607 | 10 | 5.98 × 10−23 | ALPK2, RPL9P31 | |
| 11 | 86,085,026 | 86,086,489 | 12 | 1.24 × 10−13 | CCDC81 | ||
| 19 | 29,217,858 | 29,218,774 | 7 | 1.31 × 10−11 | AC005307.3 | ||
| 18 | 3,411,487 | 3,412,713 | 11 | 1.37 × 10−11 | TGIF1 | ||
| 7 | 157,866,683 | 157,868,361 | 13 | 2.33 × 10−11 | |||
| Impact | Blood | 6 | 42,927,199 | 42,928,920 | 31 | 6.89 × 10−33 | GNMT |
| 3 | 56,501,352 | 56,502,814 | 12 | 1.26 × 10−30 | ERC2 | ||
| 18 | 74,960,629 | 74,963,364 | 35 | 6.35 × 10−27 | GALR1 | ||
| 10 | 134,598,316 | 134,601,851 | 37 | 4.30 × 10−24 | NKX6-2, RP11-288G11.3 | ||
| 3 | 47,823,674 | 47,825,578 | 6 | 1.03 × 10−23 | SMARCC1 | ||
| Airway | 19 | 35,981,224 | 35,982,442 | 10 | 1.20 × 10−19 | KRTDAP | |
| 17 | 75,470,567 | 75,472,168 | 12 | 2.22 × 10−15 | SEPT9, RP11-75C10.9 | ||
| 21 | 43,315,518 | 43,316,705 | 6 | 9.74 × 10−14 | |||
| 12 | 11,698,367 | 11,699,363 | 6 | 2.63 × 10−13 | RP11-434C1.1, RNU7-60P | ||
| 3 | 113,160,071 | 113,161,177 | 14 | 1.70 × 10−12 | WDR52 | ||
| Symptom | Blood | 6 | 32,807,895 | 32,811,521 | 30 | 3.27 × 10−24 | PSMB8, TAP2, PSMB9, TAPSAR1 |
| 5 | 78,364,769 | 78,366,302 | 14 | 2.24 × 10−22 | DMGDH, BHMT2 | ||
| 6 | 30,850,207 | 30,852,354 | 24 | 2.24 × 10−22 | DDR1 | ||
| 2 | 110,969,641 | 110,970,909 | 8 | 4.86 × 10−22 | LINC00116 | ||
| 2 | 98,329,337 | 98,330,493 | 10 | 1.96 × 10−21 | ZAP70 | ||
| Airway | 6 | 33,244,976 | 33,246,895 | 44 | 1.04 × 10−22 | B3GALT4, WDR46, RPS18 | |
| 12 | 63,025,490 | 63,026,424 | 7 | 2.02 × 10−21 | |||
| 9 | 139,425,582 | 139,427,171 | 5 | 5.63 × 10−20 | |||
| 17 | 46,655,164 | 46,656,572 | 20 | 6.39 × 10−17 | HOXB4, MIR10A, HOXB3 | ||
| 2 | 161,992,157 | 161,993,364 | 6 | 1.19 × 10−16 | TANK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez Cordero, A.I.; Li, X.; Yang, C.X.; Yang, J.; MacIsaac, J.L.; Dever, K.; Kobor, M.S.; Milne, S.; van Eeden, S.F.; Shaipanich, T.; et al. Systemic and Airway Epigenetic Disruptions Are Associated with Health Status in COPD. Biomedicines 2023, 11, 134. https://doi.org/10.3390/biomedicines11010134
Hernandez Cordero AI, Li X, Yang CX, Yang J, MacIsaac JL, Dever K, Kobor MS, Milne S, van Eeden SF, Shaipanich T, et al. Systemic and Airway Epigenetic Disruptions Are Associated with Health Status in COPD. Biomedicines. 2023; 11(1):134. https://doi.org/10.3390/biomedicines11010134
Chicago/Turabian StyleHernandez Cordero, Ana I., Xuan Li, Chen Xi Yang, Julia Yang, Julia L. MacIsaac, Kristy Dever, Michael S. Kobor, Stephen Milne, Stephan F. van Eeden, Tawimas Shaipanich, and et al. 2023. "Systemic and Airway Epigenetic Disruptions Are Associated with Health Status in COPD" Biomedicines 11, no. 1: 134. https://doi.org/10.3390/biomedicines11010134