
Analysis of Purine Metabolism to Elucidate the Pathogenesis of Acute Kidney Injury in Renal Hypouricemia

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
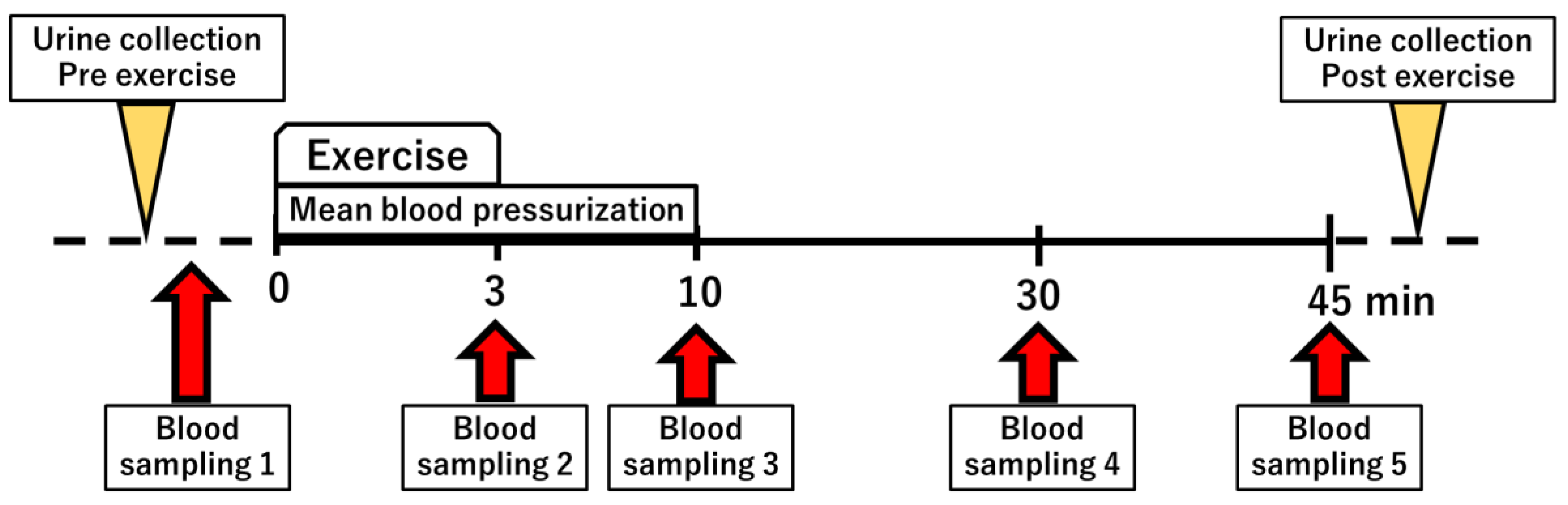
2.1. Semi-Ischemic Forearm Exercise Test
2.2. Analysis of Purine Metabolites
2.3. Evaluation of Oxidative Stress Due to Exercise Load
2.4. Adenine Phosphoribosyltransferase and Hypoxanthine Phosphoribosyl Transferase Activity
3. Results
3.1. Patient Background
- Case 6. A 35-year-old man (hereditary type 1 XU): compound heterozygote of two nonsense variants: one base pair deletion in exon 8 (p. P214QfsX4) and one nonsense variant in exon 23 of the XOR gene (p. R825X) [21].
- Case 7. A 45-year-old man (type 1 RHUC): compound heterozygote of c.1096G>C (p. G366R) in exon 7 and c.1430G>A (p. R477H) in exon 9 of URAT1. Variants p.G366R and p.R477H have low urate transport activity, and their combination synergistically loosens the urate transport activity [22].
- Case 8. A 49-year-old woman (type 1 RHUC): compound heterozygote of c.1245_1253del (delGGCAGGGCT, p.L415_G417del) in exon 7 c.1400C>T (p. T467M) in exon 9 of SLC22A12. The urate transport of both variants was significantly decreased in comparison with the wt (p.L415_G417delP < 0.01, p. T467M p < 0.05) [23].
- Case 9. A 62-year-old woman (type 1 RHUC): The subject was the older sister of Case 8 and had the same genotype [23].
- Case 10. A 58-year-old woman (type 2 RHUC) was heterozygous for the variant c.215G>A (p.G72D) in exon 2 of URAT1. The urate transport of the variant was significantly decreased compared to that of wt (p < 0.05) [24].
- Case 11. A 39-year-old woman (type 1 RHUC): compound heterozygote of c.774G>A (p. W258X;rs121907892) in exon 4 and c.269G>A (p. R90H;rs121907896) in exon 1 of URAT1. Both the variants lost the urate transport activity [25].
- Case 12. A 42-year-old man (type 1 RHUC): compound heterozygote of c.774G>A (p. W258X;rs121907892) in exon 4 and c.269G>A (p. R90H;rs121907896) in exon 1 of SLC22A12. Both variants lost the urate transport activity [25].
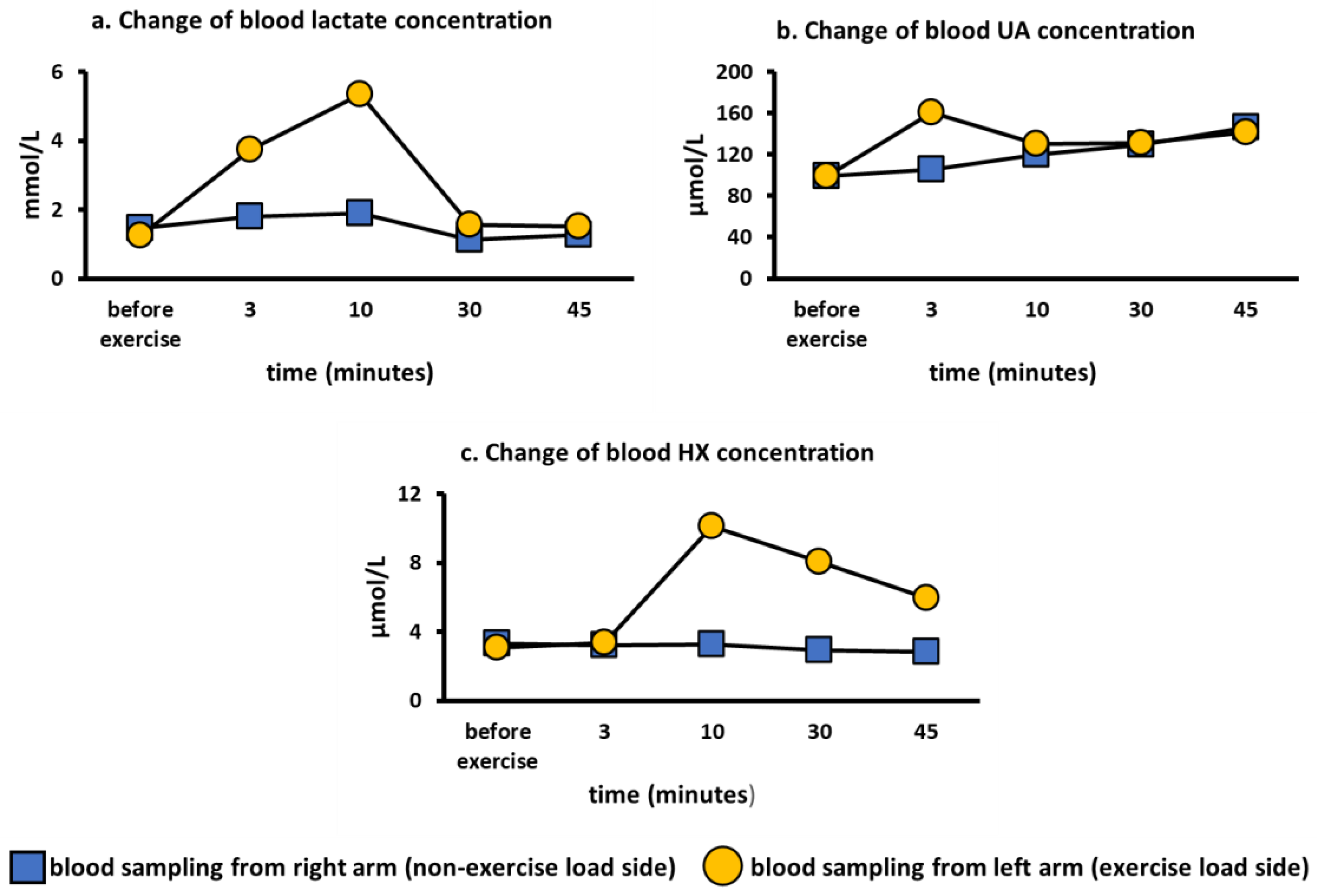
3.2. Comparison of Lactate and Purine Metabolites in the Exercise Loading Arm and Non-Loading Arm
3.3. Comparison of Blood and Urine Analytes in Healthy Subjects and Hypouricemic Subjects
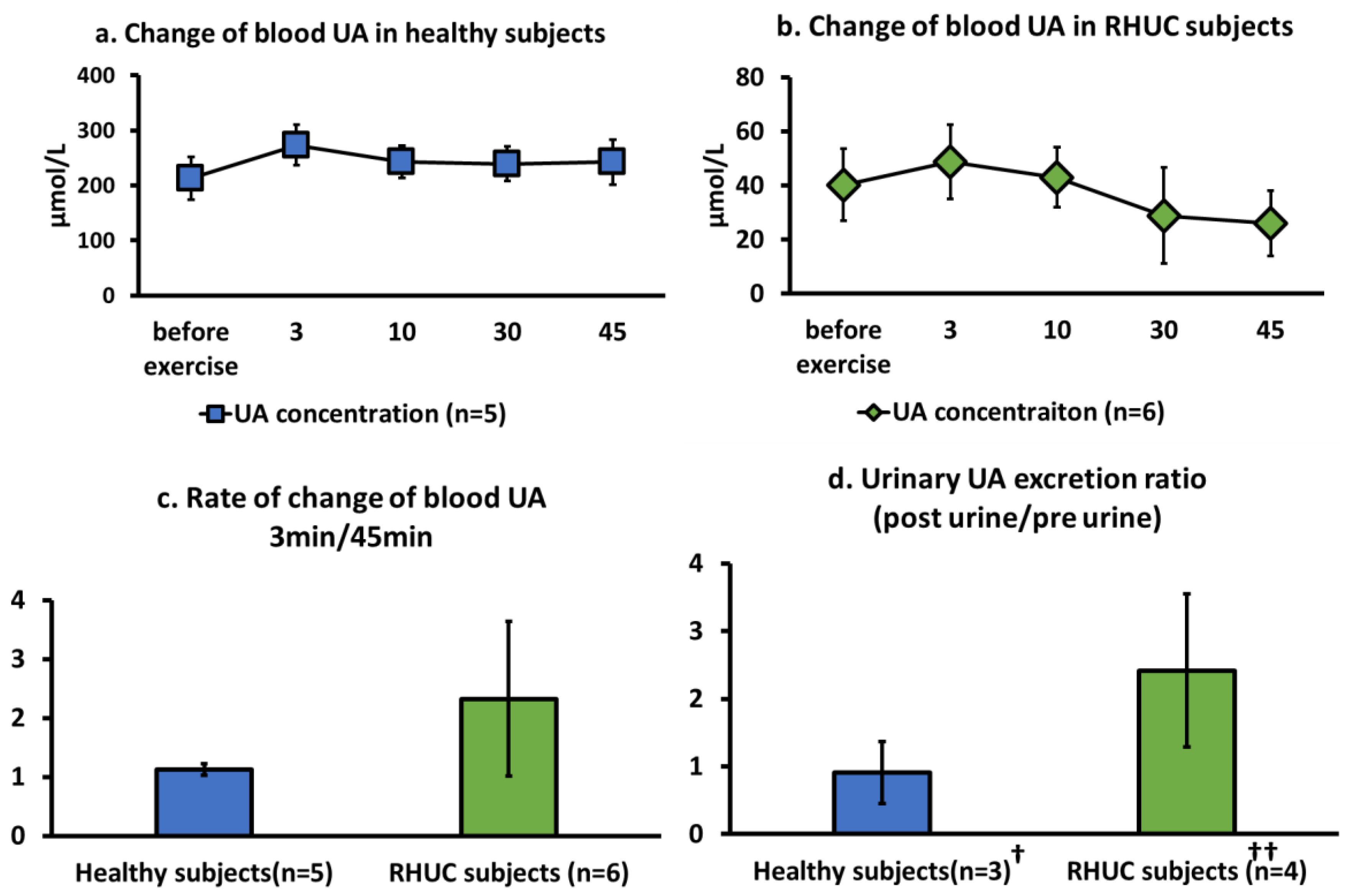
3.3.1. Change of Blood UA Concentration
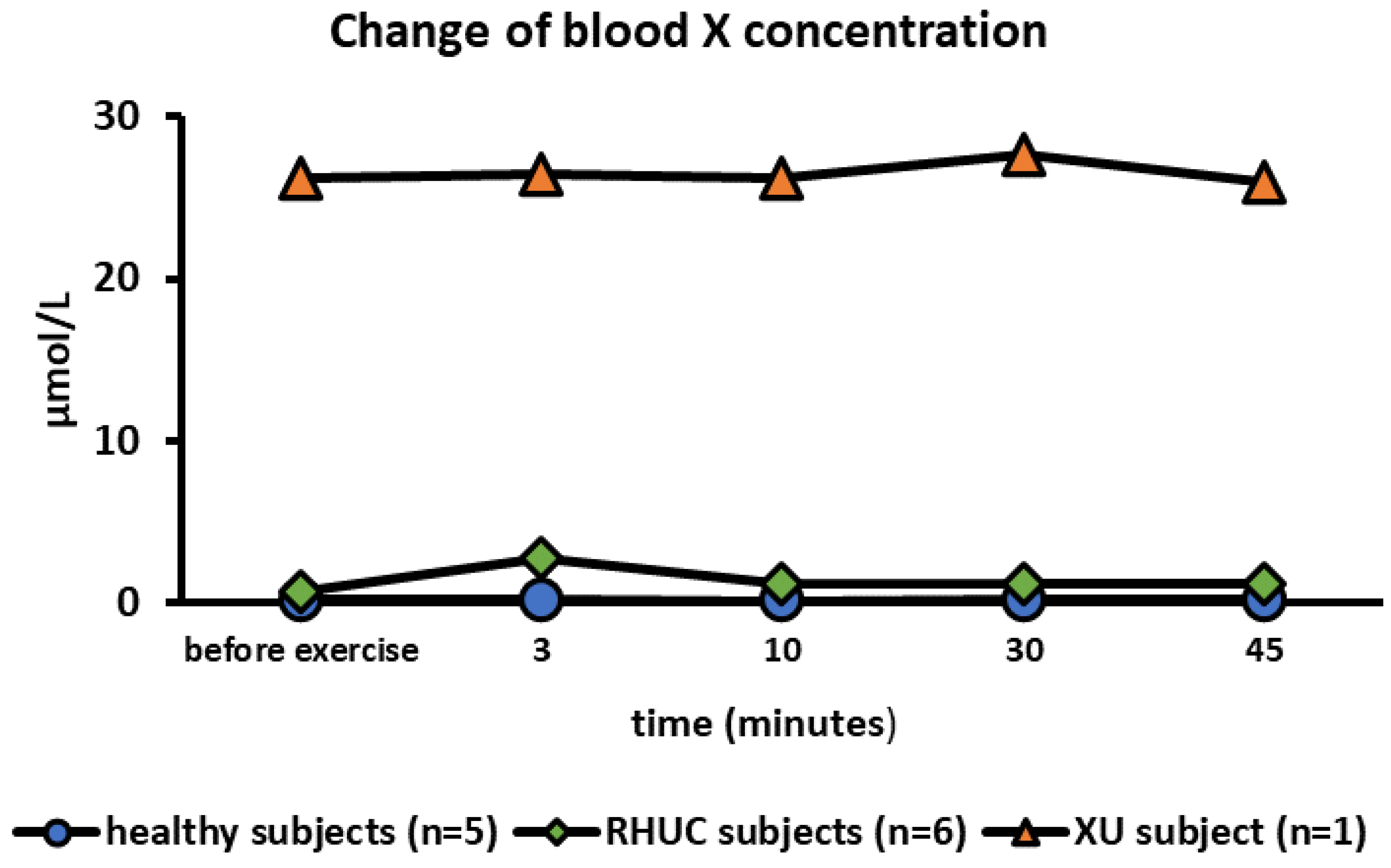
3.3.2. Change of Blood Xanthine Concentration
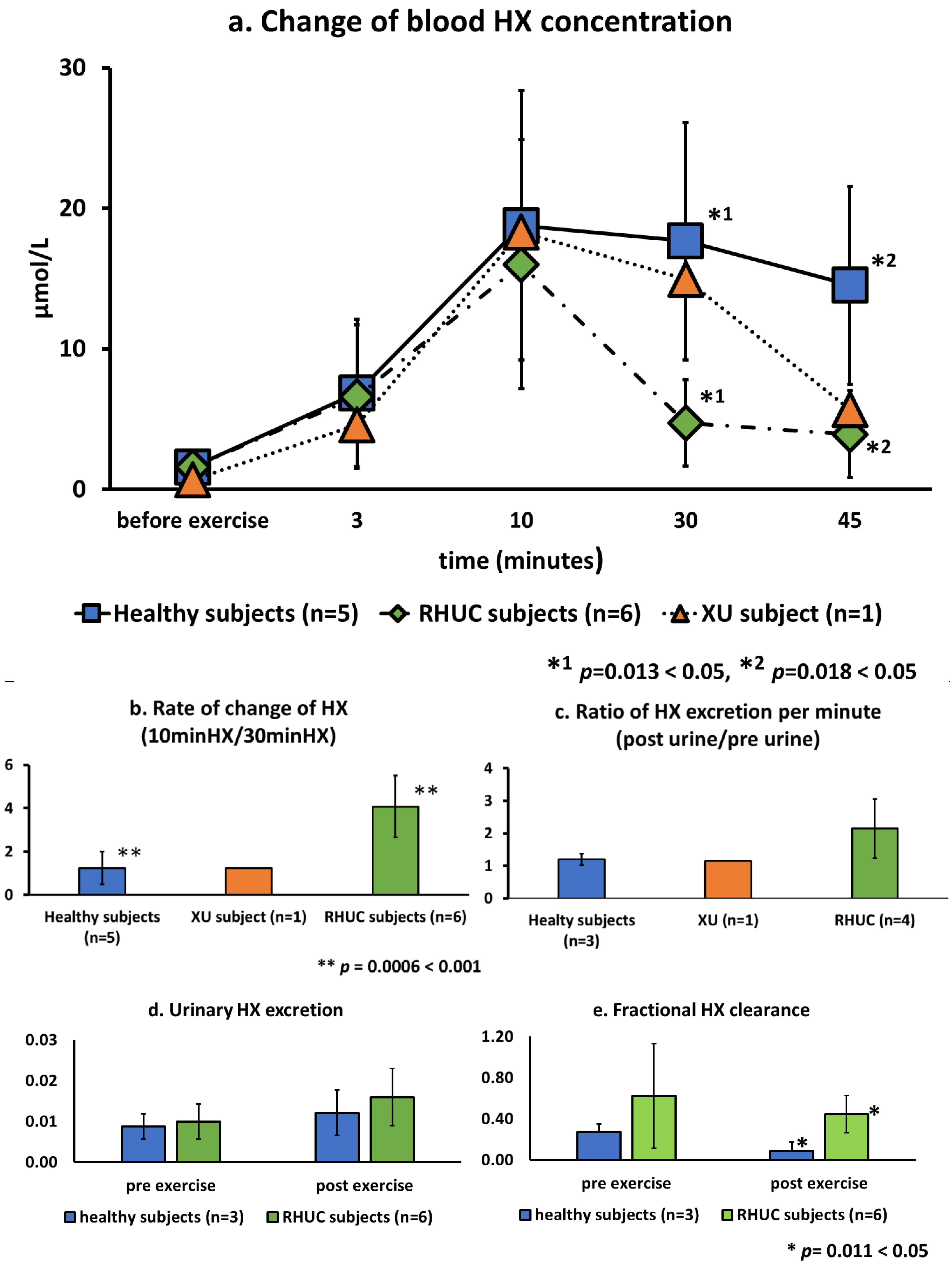
3.3.3. Change of Blood Hypoxanthine Concentration
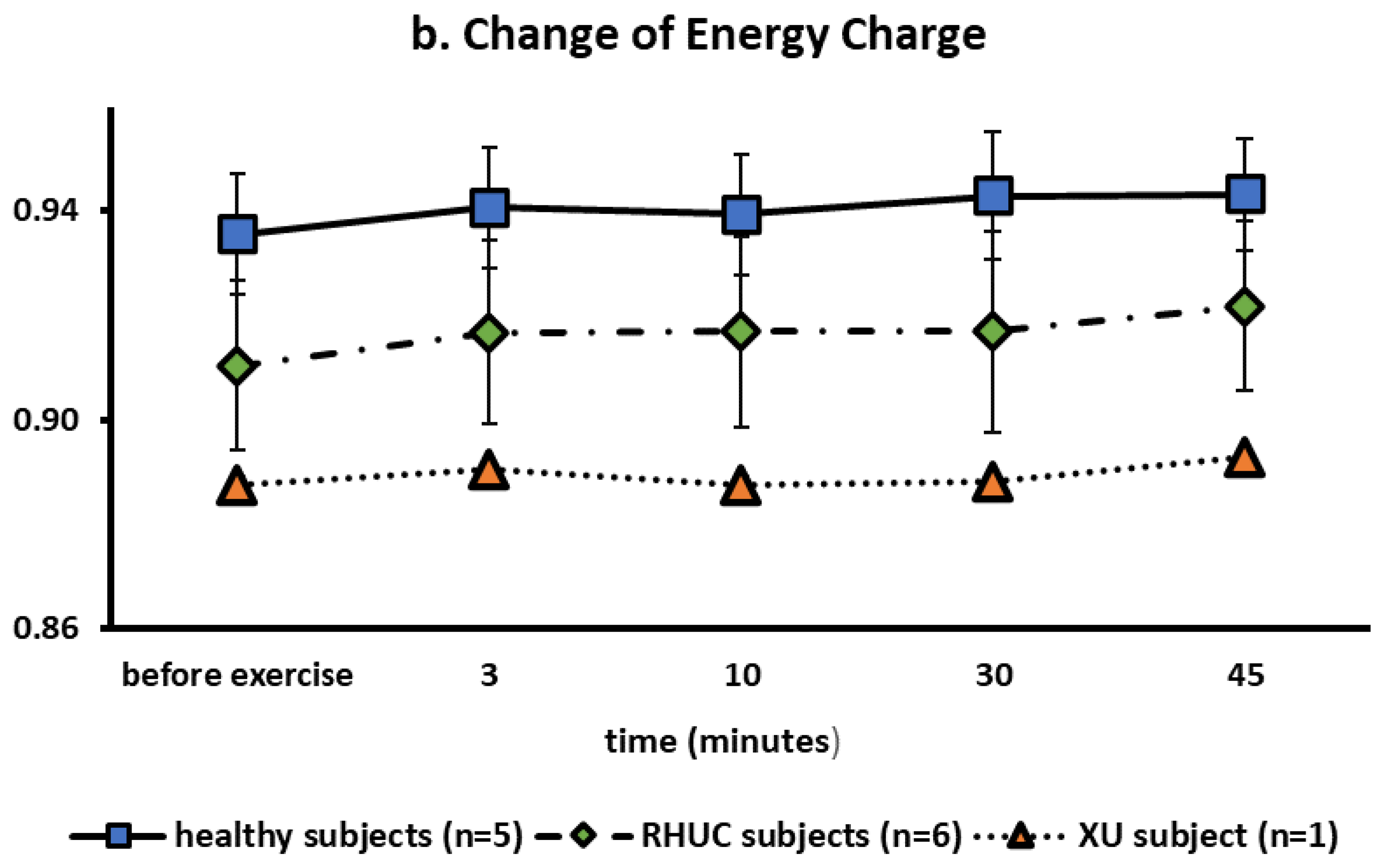
3.3.4. Change of Blood ATP Concentration
3.3.5. APRT and HPRT Activity in RBC
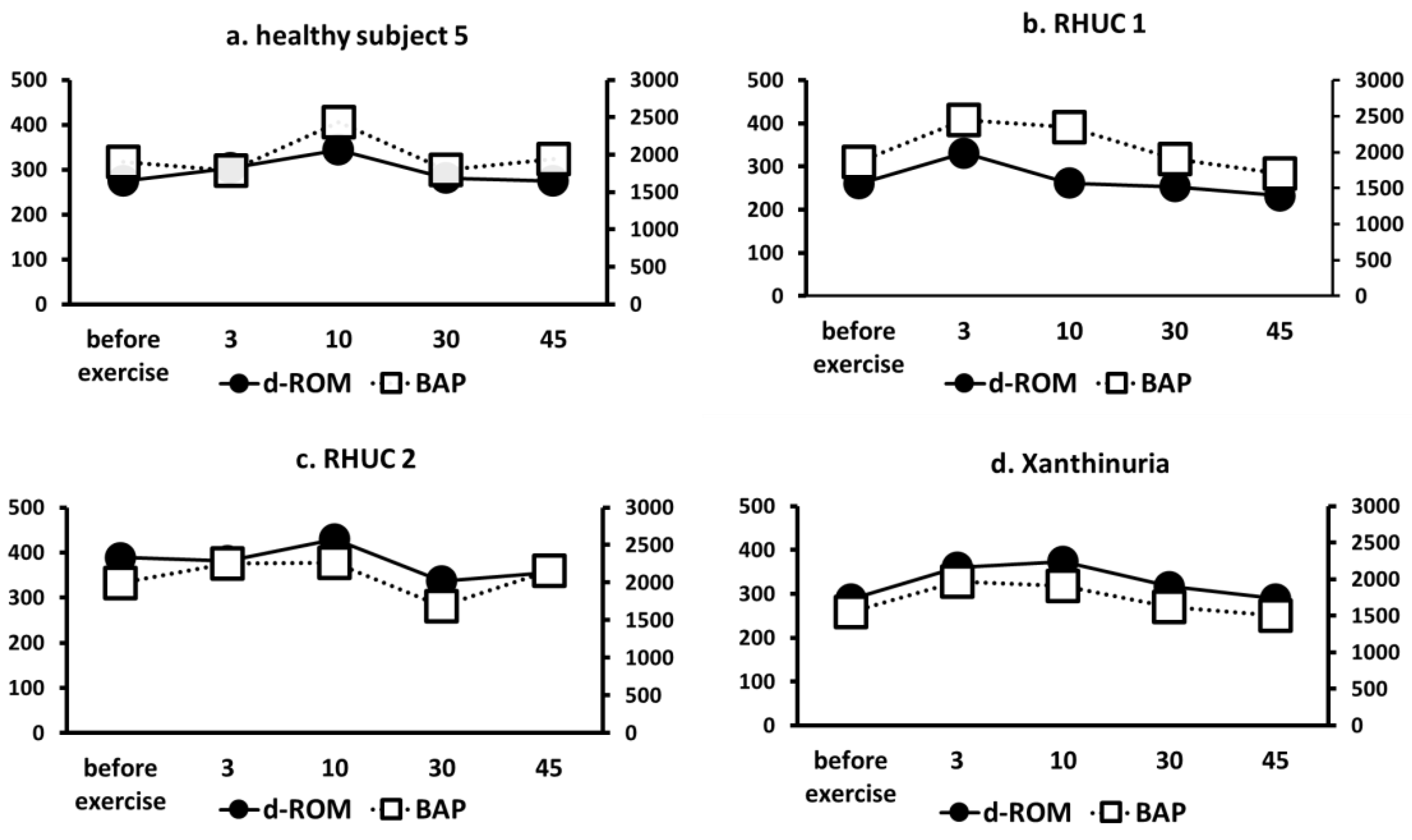
3.3.6. Changes in ROS Markers (d-ROM) and Antioxidant Capacity Markers (BAP)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramsdell, C.M.; Kelley, W.N. The Clinical Significance of Hypouricemia. Ann. Intern. Med. 1973, 78, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Pineda, C.; Soto-Fajardo, C.; Mendoza, J.; Gutiérrez, J.; Sandoval, H. Hypouricemia: What the practicing rheumatologist should know about this condition. Clin. Rheumatol. 2020, 39, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Shi, Y.; Zhuang, S.; Liu, N. Recent advances on uric acid transporters. Oncotarget 2017, 8, 100852–100862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, A.; Matsuo, H.; Ohtahara, A.; Ogino, K.; Hakoda, M.; Hamada, T.; Hosoyamada, M.; Yamaguchi, S.; Hisatome, I.; Ichida, K.; et al. Clinical practice guideline for renal hypouricemia (1st edition). Hum. Cell 2019, 32, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, Y.; Nakayama, A.; Shimizu, S.; Toyoda, Y.; Nishida, Y.; Hishida, A.; Katsuura-Kamano, S.; Shibuya, K.; Tamura, T.; Kawaguchi, M.; et al. A Proposal for Practical Diagnosis of Renal Hypouricemia: Evidenced from Genetic Studies of Nonfunctional Variants of URAT1/SLC22A12 among 30,685 Japanese Individuals. Biomedicines 2021, 9, 1012. [Google Scholar] [CrossRef]
- Ichida, K.; Hosoyamada, M.; Kamatani, N.; Kamitsuji, S.; Hisatome, I.; Shibasaki, T.; Hosoya, T. Age and origin of the G774A mutation in SLC22A12 causing renal hypouricemia in Japanese. Clin. Genet. 2008, 74, 243–251. [Google Scholar] [CrossRef]
- Claverie-Martin, F.; Trujillo-Suarez, J.; Gonzalez-Acosta, H.; Aparicio, C.; Roldan, M.L.J.; Stiburkova, B.; Ichida, K.; Martín-Gomez, M.A.; Goñi, M.H.; Hidalgo-Barquero, M.C.; et al. URAT1 and GLUT9 mutations in Spanish patients with renal hypouricemia. Clin. Chim. Acta 2018, 481, 83–89. [Google Scholar] [CrossRef]
- Stiburkova, B.; Bohatá, J.; Pavelcová, K.; Tasic, V.; Plaseska-Karanfilska, D.; Cho, S.-K.; Potočnaková, L.; Šaligová, J. Renal Hypouricemia 1: Rare Disorder as Common Disease in Eastern Slovakia Roma Population. Biomedicines 2021, 9, 1607. [Google Scholar] [CrossRef]
- Stiburkova, B.; Gabrikova, D.; Čepek, P.; Šimek, P.; Kristian, P.; Cordoba-Lanus, E.; Claverie-Martin, F. Prevalence of URAT1 allelic variants in the Roma population. Nucleosides Nucleotides Nucleic Acids 2016, 35, 529–535. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Liu, S.; Liang, X.; Song, Y.; Feng, L.; Zhong, L.; Guo, X. Idiopathic renal hypouricemia: A case report and literature review. Mol. Med. Rep. 2019, 20, 5118–5124. [Google Scholar] [CrossRef] [Green Version]
- Ames, B.N.; Cathcart, R.; Schwiers, E.; Hochstein, P. Uric acid provides an antioxidant defense in humans against oxidant- and radical-caused aging and cancer: A hypothesis. Proc. Natl. Acad. Sci. USA 1981, 78, 6858–6862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Becker, B.; Hupkens, E.; Dewachter, L.; Coremans, C.; Delporte, C.; van Antwerpen, P.; Franck, T.; Boudjeltia, K.Z.; Cullus, P.; van de Borne, P. Acute effects of hypouricemia on endothelium, oxidative stress, and arterial stiffness: A randomized, double-blind, crossover study. Physiol. Rep. 2021, 9, e15018. [Google Scholar] [CrossRef] [PubMed]
- Simmouds, H.A. Hereditary Xanthinuria. Available online: https://www.orpha.net/data/patho/GB/uk-XDH.pdf (accessed on 31 May 2022).
- Ichida, K.; Amaya, Y.; Kamatani, N.; Nishino, T.; Hosoya, T.; Sakai, O. Identification of two mutations in human xanthine dehydrogenase gene responsible for classical type I xanthinuria. J. Clin. Investig. 1997, 99, 2391–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebesta, I.; Miyamoto, D.; Stiburkova, B.; Blahova, S.; Sato, N.; Nagata, K.; Okamoto, K.; Tsuruoka, S.; Ichida, K. Modified forearm ischemic test in hypouricemic patients. Nucleosides Nucleotides Nucleic Acids 2020, 39, 1432–1439. [Google Scholar] [CrossRef]
- Tani, T.; Okamoto, K.; Fujiwara, M.; Katayama, A.; Tsuruoka, S. Metabolomics analysis elucidates unique influences on purine/pyrimidine metabolism by xanthine oxidoreductase inhibitors in a rat model of renal ischemia-reperfusion injury. Mol. Med. 2019, 25, 40. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Terawaki, H.; Nakayama, M.; Iwabuchi, M.; Sato, T.; Ito, S. Reduction of serum antioxidative capacity during hemodialysis. Clin. Exp. Nephrol. 2007, 11, 218–224. [Google Scholar] [CrossRef]
- Iamele, L.; Fiocchi, R.; Vernocchi, A. Evaluation of an automated spectrophotometric assay for reactive oxygen metabolites in serum. Clin. Chem. Lab. Med. 2002, 40, 673–676. [Google Scholar] [CrossRef]
- Sakuma, R.; Nishina, T.; Kitamura, M.; Yamanaka, H.; Kamatani, N.; Nishioka, K. Screening for adenine and hypoxanthine phosphoribosyltransferase deficiencies in human erythrocytes by high-performance liquid chromatography. Clin. Chim. Acta 1987, 170, 281–289. [Google Scholar] [CrossRef]
- Dudzinska, W.; Suska, M.; Lubkowska, A.; Jakubowska, K.; Olszewska, M.; Safranow, K.; Chlubek, D. Comparison of human erythrocyte purine nucleotide metabolism and blood purine and pyrimidine degradation product concentrations before and after acute exercise in trained and sedentary subjects. J. Physiol. Sci. 2018, 68, 293–305. [Google Scholar] [CrossRef]
- Stiburkova, B.; Krijt, J.; Vyletal, P.; Bartl, J.; Gerhatova, E.; Korinek, M.; Sebesta, I. Novel mutations in xanthine dehydrogenase/oxidase cause severe hypouricemia: Biochemical and molecular genetic analysis in two Czech families with xanthinuria type I. Clin. Chim. Acta 2012, 413, 93–99. [Google Scholar] [CrossRef]
- Mancikova, A.; Krylov, V.; Hurba, O.; Šebesta, I.; Nakamura, M.; Ichida, K.; Stiburkova, B. Functional analysis of novel allelic variants in URAT1 and GLUT9 causing renal hypouricemia type 1 and 2. Clin. Exp. Nephrol. 2015, 20, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Stiburkova, B.; Šebesta, I.; Ichida, K.; Nakamura, M.; Hůlková, H.; Krylov, V.; Kryspinova, L.; Jahnová, H. Novel allelic variants and evidence for a prevalent mutation in URAT1 causing renal hypouricemia: Biochemical, genetics and functional analysis. Eur. J. Hum. Genet. 2013, 21, 1067–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiburkova, B.; Stekrova, J.; Nakamura, M.; Ichida, K. Hereditary Renal Hypouricemia Type 1 and Autosomal Dominant Polycystic Kidney Disease. Am. J. Med Sci. 2015, 350, 268–271. [Google Scholar] [CrossRef]
- Ichida, K.; Hosoyamada, M.; Hisatome, I.; Enomoto, A.; Hikita, M.; Endou, H.; Hosoya, T. Clinical and Molecular Analysis of Patients with Renal Hypouricemia in Japan-Influence of URAT1 Gene on Urinary Urate Excretion. J. Am. Soc. Nephrol. 2004, 15, 164–173. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hisatome, I.; Kinugawa, T.; Tanaka, H.; Tomikura, Y.; Ando, F.; Matsuura, T.; Igawa, G.; Matsubara, K.; Yamamoto, Y.; et al. Excessive purine degradation during semi-ischemic forearm test in patients with diabetes mellitus. Intern. Med. 2003, 42, 788–792. [Google Scholar] [CrossRef] [Green Version]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The Human Serum Metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanďár, R.; Štramová, X.; Drábková, P.; Křenková, J. A Monitoring of Allantoin, Uric Acid, and Malondialdehyde Levels in Plasma and Erythrocytes After Ten Minutes of Running Activity. Physiol. Res. 2014, 63, 753–762. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kario, K.; Suda, M.; Moriwaki, Y.; Takahashi, S.; Higashino, K. A Case of Xanthinuria: A Study on the Metabolism of Pyrazinamide and Allopurinol. Jpn. J. Med. 1991, 30, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Dudzinska, W.; Hlynczak, A.J.; Skotnicka, E.; Suska, M. The purine metabolism of human erythrocytes. Biochemistry 2006, 71, 467–475. [Google Scholar] [CrossRef]
- Casali, E.; Berni, P.; Spisni, A.; Baricchi, R.; Pertinhez, T.A. Hypoxanthine: A new paradigm to interpret the origin of transfusion toxicity. Blood Transfus. 2015, 14, 555–556. [Google Scholar] [CrossRef]
- Atkinson, D.E.; Walton, G.M. Adenosine Triphosphate Conservation in Metabolic Regulation. J. Biol. Chem. 1967, 242, 3239–3241. [Google Scholar] [CrossRef]
- Dudzinska, W.; Lubkowska, A.; Dolegowska, B.; Safranow, K.; Jakubowska, K. Adenine, guanine and pyridine nucleotides in blood during physical exercise and restitution in healthy subjects. Eur. J. Appl. Physiol. 2010, 110, 1155–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, J.; Inoue, K.; Maeda, J.; Yasujima, T.; Ohta, K.; Kanai, Y.; Takada, T.; Matsuo, H.; Yuasa, H. Functional identification of SLC43A3 as an equilibrative nucleobase transporter involved in purine salvage in mammals. Sci. Rep. 2015, 5, 15057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, H.; Amezawa, N.; Kawakatsu, Y.; Tamai, I. Renal Reabsorptive Transport of Uric Acid Precursor Xanthine by URAT1 and GLUT9. Biol. Pharm. Bull. 2020, 43, 1792–1798. [Google Scholar] [CrossRef]
- Aomura, D.; Sonoda, K.; Harada, M.; Hashimoto, K.; Kamijo, Y. A Case of Acute Kidney Injury in a Patient with Renal Hypouricemia without Intense Exercise. Case Rep. Nephrol. Dial. 2020, 10, 26–34. [Google Scholar] [CrossRef]
- Erley, C.M.M.; Hirschberg, R.R.; Hoefer, W.; Schaefer, K. Acute renal failure due to uric acid nephropathy in a patient with renal hypouricemia. Klin. Wochenschr. 1989, 67, 308–312. [Google Scholar] [CrossRef]
- Kaneko, K.; Taniguchi, N.; Tanabe, Y.; Nakano, T.; Hasui, M.; Nozu, K. Oxidative imbalance in idiopathic renal hypouricemia. Pediatr. Nephrol. 2009, 24, 869–871. [Google Scholar] [CrossRef]
- Bhasin, B.; Stiburkova, B.; De Castro-Pretelt, M.; Beck, N.; Bodurtha, J.N.; Atta, M.G. Hereditary renal hypouricemia: A new role for allopurinol? Am. J. Med. 2014, 127, e3–e4. [Google Scholar] [CrossRef]
- Yeun, J.Y.; Hasbargen, J.A. Renal hypouricemia: Prevention of exercise-induced acute renal failure and a review of the literature. Am. J. Kidney Dis. 1995, 25, 937–946. [Google Scholar] [CrossRef]
- Stout, J.T.; Caskey, C.T. HPRT: Gene structure, expression, and mutation. Annu. Rev. Genet. 1985, 19, 127–148. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case No. | Age/Sex | Diagnosis | Race | BMI | SBP/DBP/MAP | Notation on the Table and Figure |
|---|---|---|---|---|---|---|
| 1 | 23/F | Healthy subject | Japanese | 18.7 | 92/52/65.3 | healthy subject 1 |
| 2 | 23/M | Healthy subject | Japanese | 21.2 | 94/68/76.7 | healthy subject 2 |
| 3 | 55/M | Healthy subject | Japanese | 30.6 | 128/82/96.7 | healthy subject 3 |
| 4 | 24/M | Healthy subject | Japanese | 20.3 | 108/72/84 | healthy subject 4 |
| 5 | 54/M | Healthy subject | Japanese | 22.7 | 118/78/91 | healthy subject 5 |
| 6 | 35/M | Xanthinuria | Czech | 25.3 | 130/80/96 | XU |
| 7 | 45/M | Renal hypouricemia | Czech | 26.3 | 133/80/97 | RHUC 1 |
| 8 | 49/F | Renal hypouricemia | Czech | 46.8 | 125/90/101 | RHUC 2 |
| 9 | 62/F | Renal hypouricemia | Czech | 18 | 127/84/98 | RHUC 3 |
| 10 | 58/F | Renal hypouricemia | Czech | 21.2 | 120/87/98 | RHUC 4 |
| 11 | 39/F | Renal hypouricemia | Japanese | 19.1 | 102/72/82 | RHUC 5 |
| 12 | 42/M | Renal hypouricemia | Japanese | 23.4 | 104/72/83 | RHUC 6 |
| APRT Activity (Unit: μmol/min/g Hb) | HPRT Activity (Unit: μmol/min/g Hb) | |
|---|---|---|
| healthy subject 5 | 0.46 | 2.34 |
| XU | 0.67 | 2.23 |
| RHUC 1 | 0.69 | 2.16 |
| RHUC 2 | 0.65 | 2.09 |
| RHUC 6 | 0.51 | 2.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyamoto, D.; Sato, N.; Nagata, K.; Sakai, Y.; Sugihara, H.; Ohashi, Y.; Stiburkova, B.; Sebesta, I.; Ichida, K.; Okamoto, K. Analysis of Purine Metabolism to Elucidate the Pathogenesis of Acute Kidney Injury in Renal Hypouricemia. Biomedicines 2022, 10, 1584. https://doi.org/10.3390/biomedicines10071584
Miyamoto D, Sato N, Nagata K, Sakai Y, Sugihara H, Ohashi Y, Stiburkova B, Sebesta I, Ichida K, Okamoto K. Analysis of Purine Metabolism to Elucidate the Pathogenesis of Acute Kidney Injury in Renal Hypouricemia. Biomedicines. 2022; 10(7):1584. https://doi.org/10.3390/biomedicines10071584
Chicago/Turabian StyleMiyamoto, Daisuke, Nana Sato, Koji Nagata, Yukinao Sakai, Hitoshi Sugihara, Yuki Ohashi, Blanka Stiburkova, Ivan Sebesta, Kimiyoshi Ichida, and Ken Okamoto. 2022. "Analysis of Purine Metabolism to Elucidate the Pathogenesis of Acute Kidney Injury in Renal Hypouricemia" Biomedicines 10, no. 7: 1584. https://doi.org/10.3390/biomedicines10071584