
Trifluoperazine and Its Analog Suppressed the Tumorigenicity of Non-Small Cell Lung Cancer Cell; Applicability of Antipsychotic Drugs to Lung Cancer Treatment

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microarray Data Processing
2.3. Curation and Scoring of Functional Gene Set
2.4. Survival Analysis
2.5. Cell Culture
2.6. Cell Proliferation Assay
2.7. Colony Formation Assay
2.8. Soft Agar Colony Formation Assay
2.9. Wound Healing Assay
2.10. Western Blot Analysis
2.11. Apoptosis and DNA Fragmentation Assay
2.12. Measurement of Changes in Intracellular Ca2+ Concentration
2.13. Tumor Xenograft
2.14. Tissue Preparation and Histological Analysis
2.15. Statistical Analysis
3. Results
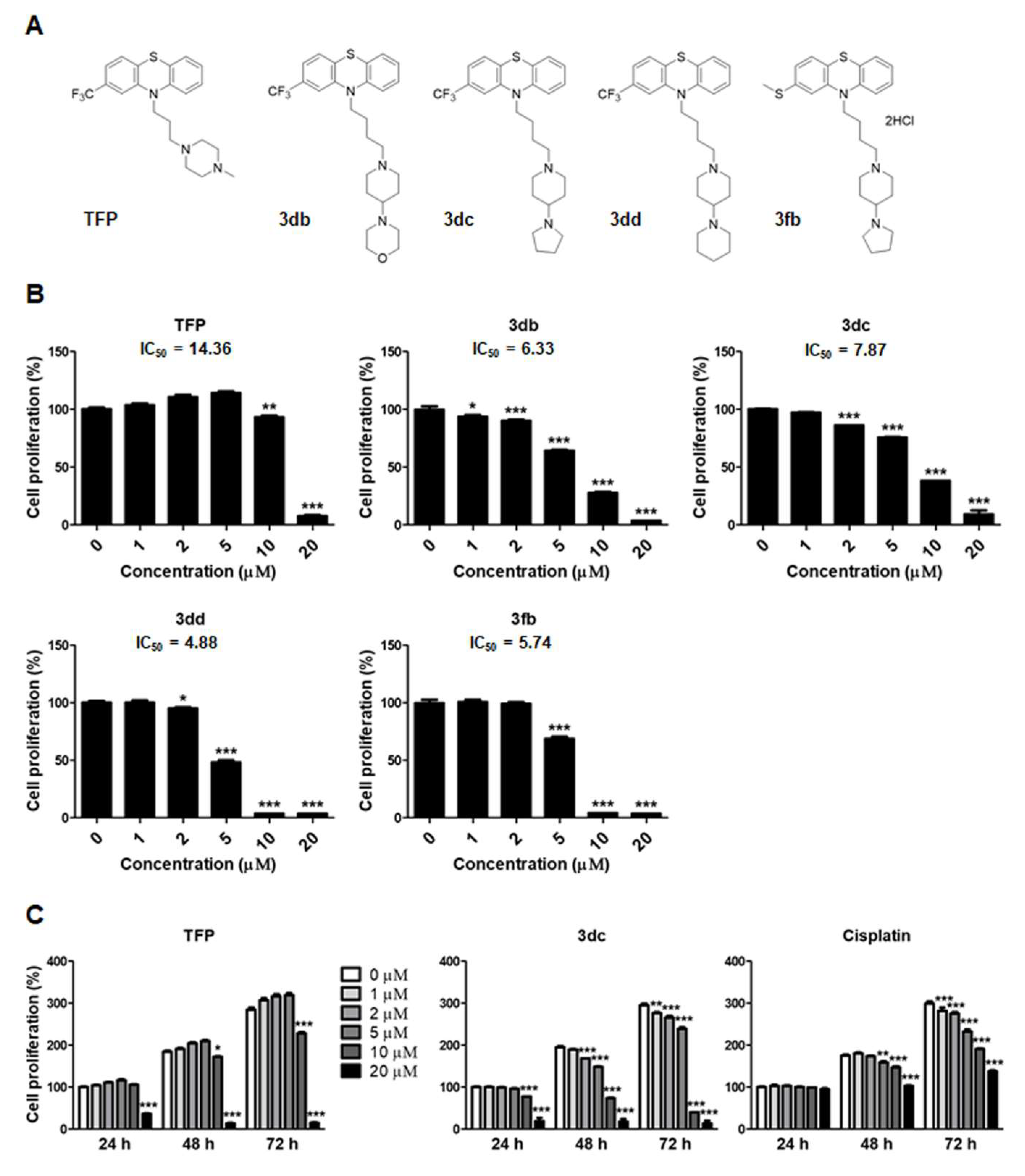
3.1. TFP and TFP Analogs Inhibit the Proliferation of A549 Cells
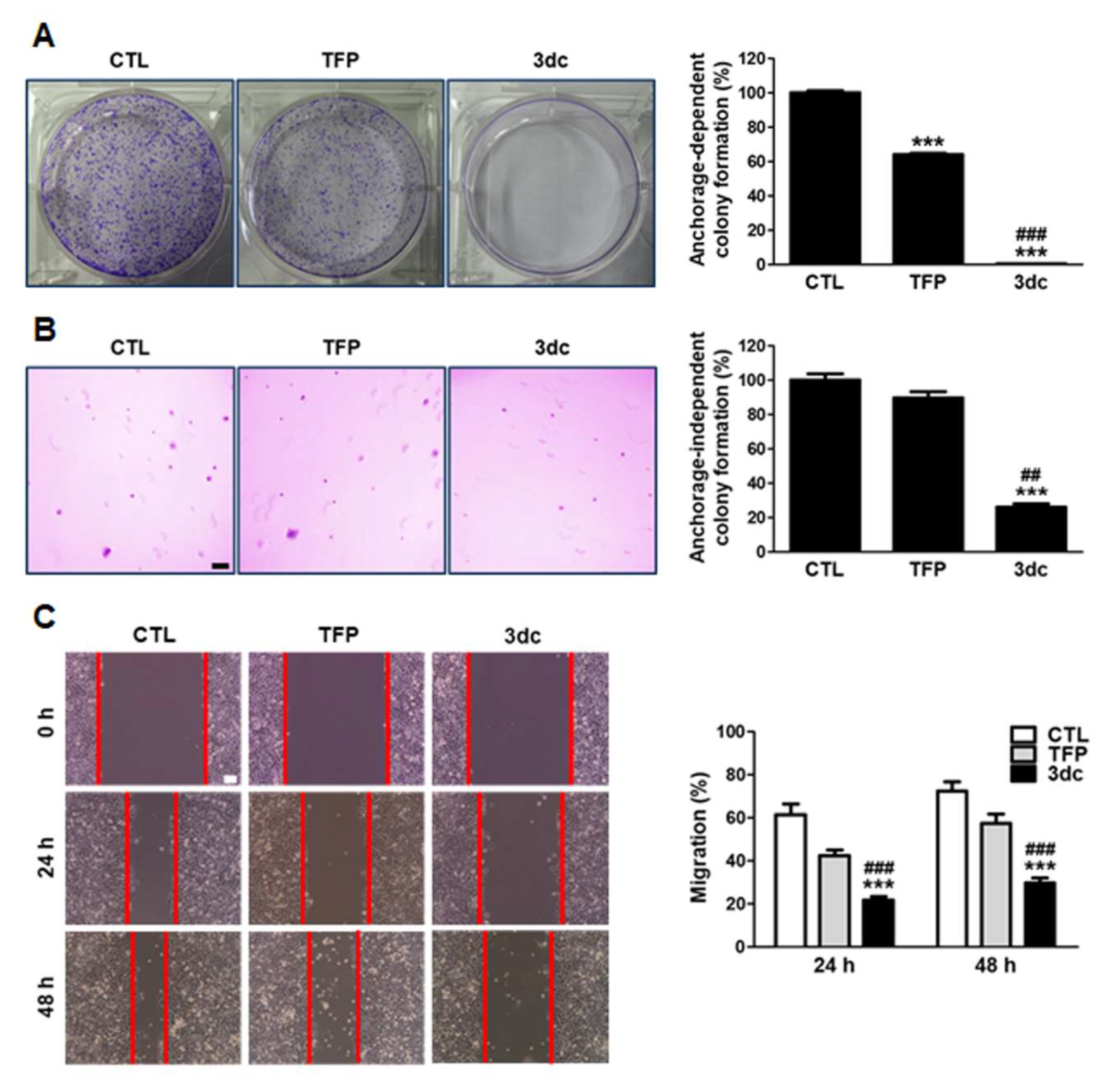
3.2. TFP and 3dc Suppress the Anchorage-Dependent and Anchorage-Independent Growth and Migration of A549 Cells
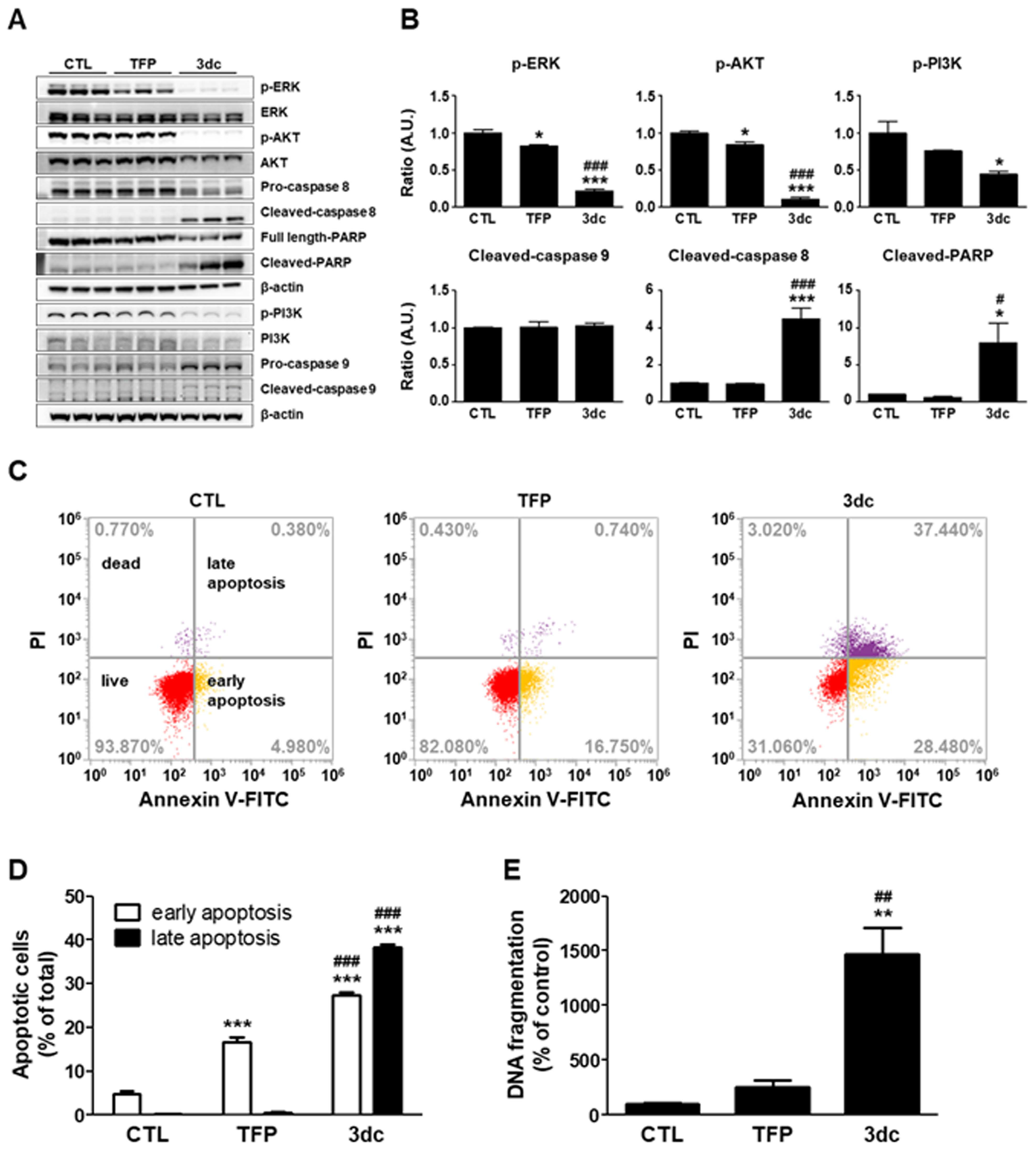
3.3. 3dc Alters the Expression of Factors Related to Apoptosis and Survival
3.4. TFP and 3dc Induce A549 Cell Apoptosis and DNA Fragmentation
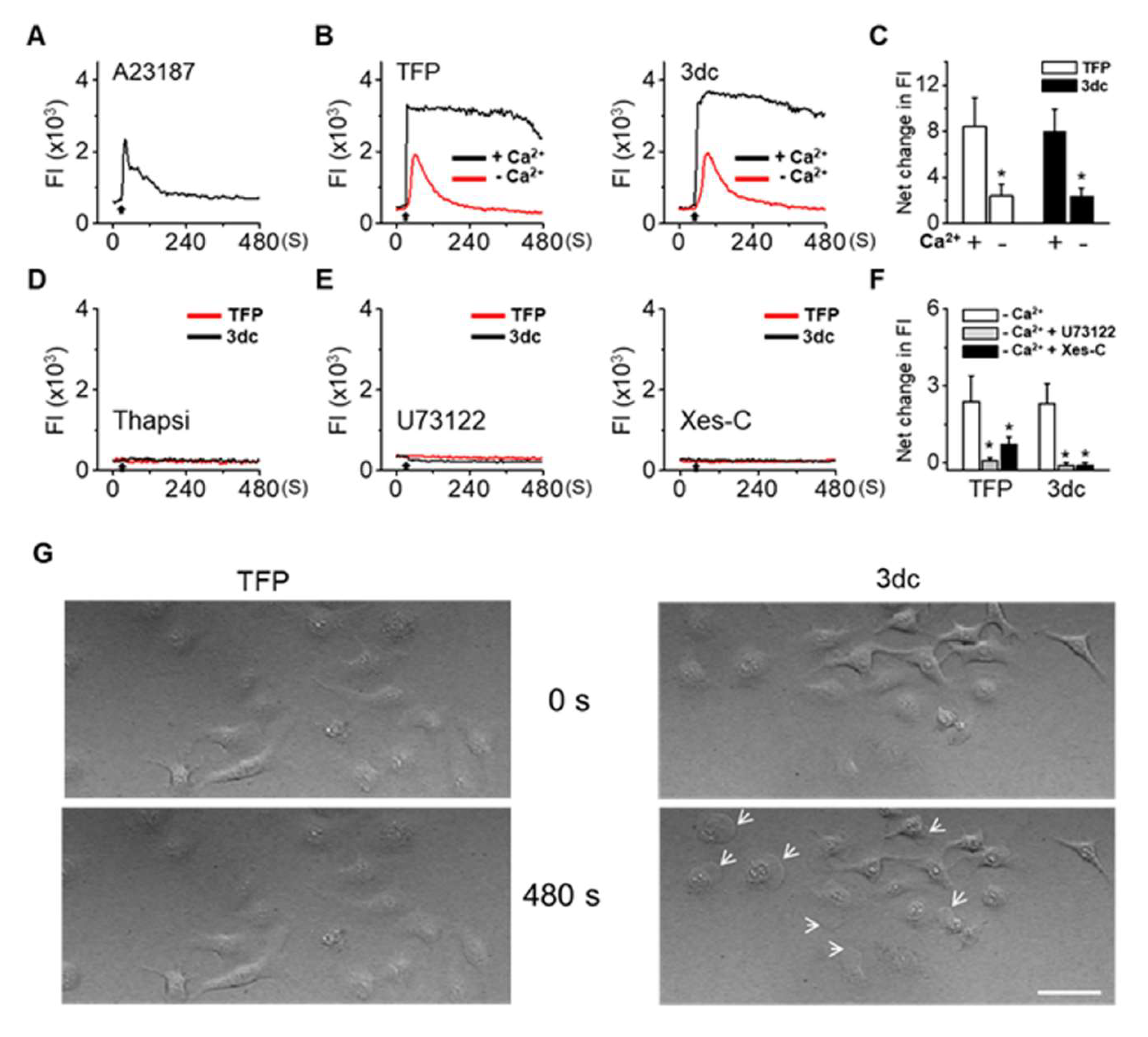
3.5. TFP and 3dc Treatment Increased Intracellular Ca2+ Levels
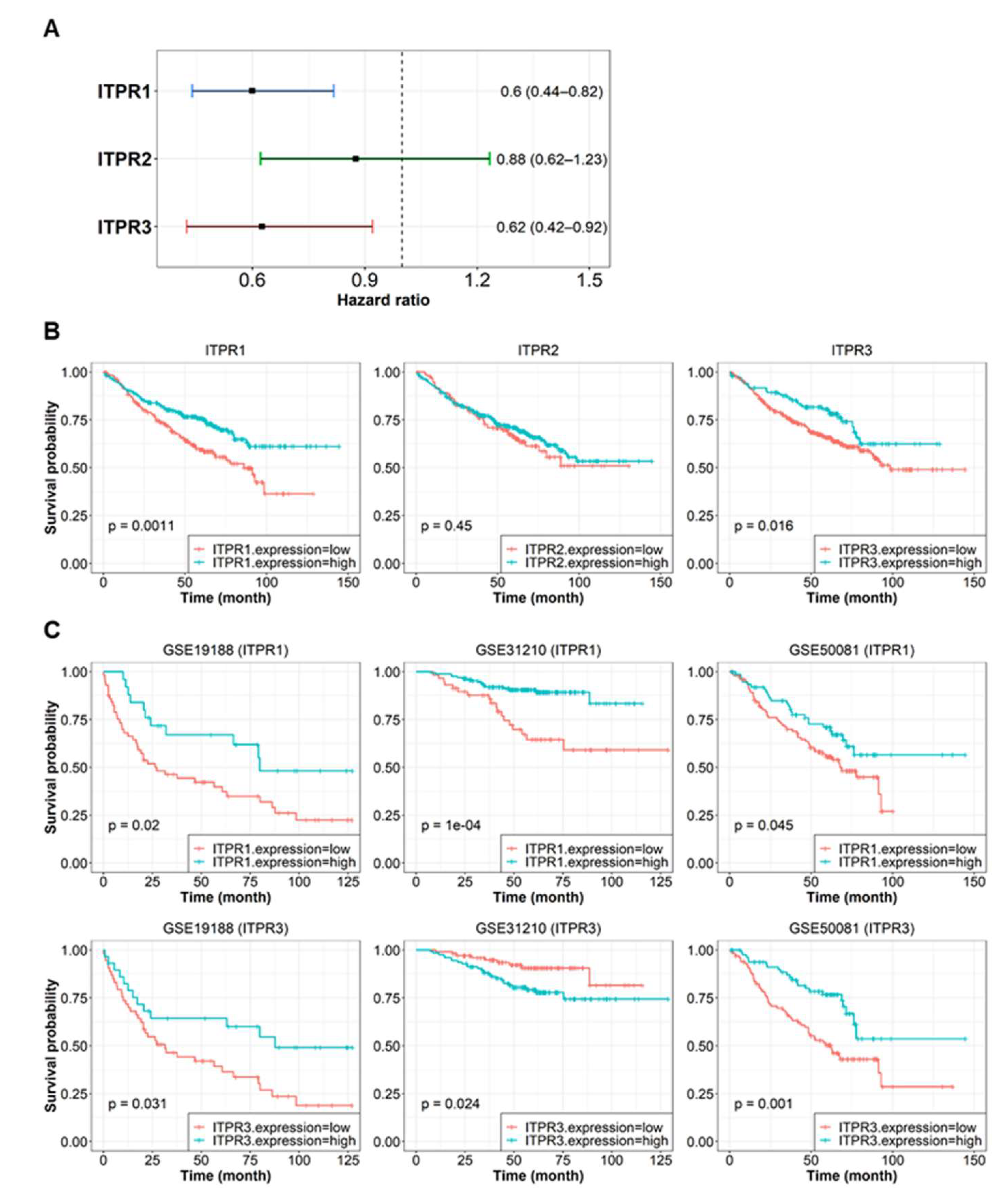
3.6. ITPR Expression and Associated Pathway Activity Are Related to Patient Survival Probability
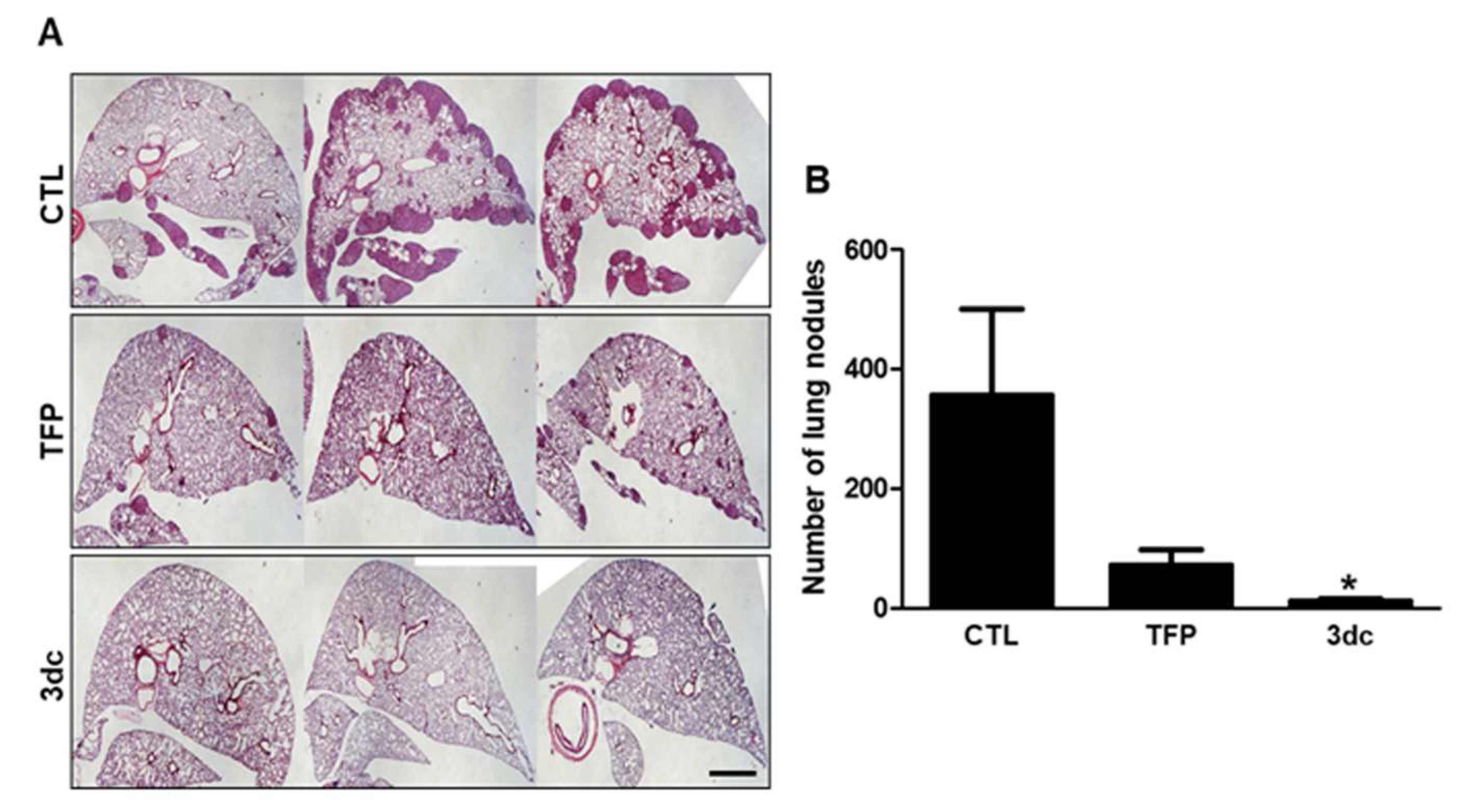
3.7. Inhibitory Effects of TFP and 3dc on Metastasis in A549 Cell-Derived Xenograft Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bittner, N.; Ostoros, G.; Géczi, L. New Treatment Options for Lung Adenocarcinoma-in View of Molecular Background. Pathol. Oncol. Res. 2014, 20, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Pfister, D.G.; Johnson, D.H.; Azzoli, C.G.; Sause, W.; Smith, T.J.; Baker, J.S.; Olak, J.; Stover, D.; Strawn, J.R.; Turrisi, A.T.; et al. American Society of Clinical Oncology treatment of unresec non-small-cell lung cancer guideline: Update. J. Clin. Oncol. 2004, 22, 330–353. [Google Scholar]
- Nordenberg, J.; Fenig, E.; Landau, M.; Weizman, R.; Weizman, A. Effect of psychotropic drugs on cell proliferation and differentiation. Biochem. Pharmacol. 1999, 58, 1229–1236. [Google Scholar] [CrossRef]
- Monrtensen, B. The occurrence of cancer in first admitted schizophrenic patients. Schizophr. Res. 1994, 12, 185–194. [Google Scholar] [CrossRef]
- Grinshpoon, A.; Barchana, M.; Ponizovsky, A.; Nahon, D.; Tal, O.; Weizman, A.; Levav, I. Cancer in schizophrenia: Is the risk higher or lower? Schizophr. Res. 2005, 73, 333–341. [Google Scholar] [CrossRef]
- Gulbinat, W.; Dupont, A.; Jablensky, A.; Jensen, O.M.; Marsella, A.; Nakane, Y.; Sartorius, N. Cancer incidence of schizophrenic patients. Results of record linkage studies in three countries. Br. J. Psychiatry 1992, 161 (Suppl. S18), 75–83. [Google Scholar] [CrossRef]
- Barak, Y.; Achiron, A.; Mandel, M.; Mirecki, I.; Aizenberg, D. Reduced cancer incidence among patients with schizophrenia. Cancer 2005, 104, 2817–2821. [Google Scholar] [CrossRef]
- Dalton, S.O.; Mellemkjaer, L.; Thomassen, L.; Mortensen, P.B.; Johansen, C. Risk for cancer in a cohort of patients hospitalized for schizophrenia in Denmark, 1969–1993. Schizophr. Res. 2005, 75, 315–324. [Google Scholar] [CrossRef]
- Hippisley-Cox, J.; Vinogradova, Y.; Coupland, C.; Parker, C. Risk of malignancy in patients with schizophrenia or bipolar disorder: Nested case-control study. Arch. Gen. Psychiatry 2007, 64, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- Chou, F.H.; Tsai, K.Y.; Su, C.Y.; Lee, C.C. The incidence and relative risk factors for developing cancer among patients with schizophrenia: A nine-year follow-up study. Schizophr. Res. 2011, 129, 97–103. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lane, H.Y.; Chen, T.T.; Wu, Y.H.; Wu, C.Y.; Wu, V.Y. Inverse association between cancer risks and age in schizophrenic patients: A 12-year nationwide cohort study. Cancer Sci. 2013, 104, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Sundquist, K.; Ning, Y.; Kendler, K.S.; Sundquist, J.; Chen, X. Incidence of cancer in patients with schizophrenia and their first-degree relatives: A population-based study in Sweden. Schizophr. Bull. 2013, 39, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.P.; Limburg, H.; Walters, K.; Petersen, I.; King, M.; Green, J.; Watson, J.; Nazareth, I. Relative incidence of common cancers in people with severe mental illness. Cohort study in the United Kingdom THIN primary care database. Schizophr. Res. 2013, 143, 44–49. [Google Scholar] [CrossRef]
- Synder, S.H. Stereoselective features of catecholamine dispostion and their behavioral implications. J. Psychiatr. Res. 1974, 11, 39–49. [Google Scholar]
- Osborn, M.; Weber, K. Damage of cellular functions by trifluoperazine, a calmodulin-specific durg. Exp. Cell Res. 1980, 34, 27–31. [Google Scholar]
- Schleuning, M.; Brumme, V.; Wilmanns, W. Growth inhibition of human leukemic cell lines by the phenothiazine derivate fluphenazine. Anticancer Res. 1993, 13, 599–602. [Google Scholar]
- Silver, M.A.; Yang, Z.W.; Ganguli, R.; Nimgaonkar, V.L. An inhibitory effect of psychoactive drugs on a human neuroblastoma cell line. Biol. Psychiatry 1994, 35, 824–826. [Google Scholar] [CrossRef]
- Kang, U.G.; Kim, M.J.; Suh, P.G.; Ryu, S.H.; Park, J.B.; Kim, J.H.; Kim, Y.S.; Lee, Y.H. Inhibition of trifluoperazine-induced DNA fragmentation by cyclic AMP mediated signaling. Mol. Cells 1999, 31, 596–602. [Google Scholar]
- Sullivan, G.F.; Garcia-Welch, A.; White, E.; Lutzker, S.; Hait, W.N. Augmentation of apoptosis by the combination of bleomycin with trifluoperazine in the presence of mutation p53. J. Exp. Ther. Oncol. 2002, 2, 19–26. [Google Scholar] [CrossRef]
- Smego, R.A., Jr.; Durack, D.T. The neuroleptic malignant syndrome. Arch. Intern. Med. 1982, 142, 1183–1185. [Google Scholar] [CrossRef]
- Kang, S.; Lee, J.M.; Jeon, B.; Elkamhawy, A.; Paik, S.; Hong, J.; Oh, S.J.; Paek, S.H.; Lee, C.J.; Hassan, A.H.E.; et al. Repositioning of the antipsychotic trifluoperazine: Synthesis, biological evaluation and in silico study of trifluoperazine analogs as anti-glioblastoma agents. Eur. J. Med. Chem. 2018, 151, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Aerts, J.; den Hamer, B.; van Ijcken, W.; den Bakker, M.; Riegman, P.; van der Leest, C.; van der Spek, P.; Foekens, J.A.; Hoogsteden, H.C.; et al. Gene expression-based classification of non-small cell lung carcinomas and survival prediction. PLoS ONE 2010, 5, e10312. [Google Scholar] [CrossRef] [PubMed]
- Okayama, H.; Kohno, T.; Ishii, Y.; Shimada, Y.; Shiraishi, K.; Iwakawa, R.; Furuta, K.; Tsuta, K.; Shibata, T.; Yamamoto, S.; et al. Identification of Genes Upregulated in ALK-positive and EGFR/KRAS/ALK-negative Lung Adenocarcinomas. Cancer Res. 2012, 72, 100–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Der, S.D.; Sykes, J.; Pintilie, M.; Zhu, C.Q.; Strumpf, D.; Liu, N.; Jurisica, I.; Shepherd, F.A.; Tsao, M.S. Validation of a Histology-Independent Prognostic Gene Signature for Early-Stage, Non-Small-Cell Lung Cancer Including Stage IA Patients. J. Thorac. Oncol. 2014, 9, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets--Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, Normalization, and Summaries of High Density Oligonucleotide Array Probe Level Data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, D. BioCarta. Biotech Softw. Internet Rep. 2001, 2, 117–120. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, C.F.; Anthony, K.; Krupa, S.; Buchoff, J.; Day, M.; Hannay, T.; Buetow, K.H. PID: The Pathway Interaction Database. Nucleic Acids Res. 2009, 37, D674–D679. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F.; McKay, S.; et al. The Reactome pathway Knowledgebase. Nucleic Acids Res. 2016, 44, D481–D487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.K.; Yoon, S.Y.; Hur, C.G.; Yang, H.Y.; Choe, C.; Kim, E.J.; Joo, J.S.; Kang, K.R.; Park, J.Y.; Hong, S.G.; et al. Acetylcholine rescues two-cell block through activation of IP3 receptors and Ca2+/calmodulin-dependent kinase II in an ICR mouse strain. Pflugers Arch. 2009, 458, 1125–1136. [Google Scholar] [CrossRef]
- Tian, L.; Shen, D.; Li, X.; Shan, X.; Wang, X.; Yan, Q.; Liu, J. Ginsenoside Rg3 inhibits epithelial-mesenchymal transition (EMT) and invasion of lung cancer by down-regulating FUT4. Oncotarget 2015, 7, 1619–1632. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, H.; Pan, T.; Li, L.; Li, J.; Yang, H. STIM1 silencing inhibits the migration and invasion of A549 cells. Mol. Med. Rep. 2017, 16, 3283–3289. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Sun, L.; Li, J.; Yu, H.; Lin, H.; Yu, T.; Zhao, F.; Zhu, M.; Liu, L.; Geng, Q.; et al. RNA-binding protein KHSRP promotes tumor growth and metastasis in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2019, 38, 478. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiang, S.; Qi, J.; Wei, Y.; Zhang, M.; Yao, J.; Zhang, T.; Meng, M.; Wang, X.; Zhou, Q. Correction of the tumor suppressor Salvador homolog-1 deficiency in tumors by lycorine as a new strategy in lung cancer therapy. Cell Death Dis. 2020, 11, 387. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/index.html (accessed on 22 March 2022).
- Davis, S.; Meltzer, P.S. GEOquery: A bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 2007, 14, 1846–1847. [Google Scholar] [CrossRef] [Green Version]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affyߞAnalysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using ‘ggplot2’, R package version 0.4.6; 2019. Available online: https://CRAN.R-project.org/package=survminer (accessed on 22 March 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Gordon, M.; Lumley, T. Forestplot: Advanced Forest Plot Using ‘Grid’ Graphics, R package version 1.9.; Umeå University: Umeå, Sweden, 2019.
- Razi, S.S.; Rehmani, S.; Li, X.; Park, K.; Schwartz, G.S.; Latif, M.J.; Bhora, F.Y. Antitumor activity of paclitaxel is significantly enhanced by a novel proapoptotic agent in non-small cell lung cancer. J. Surg. Res. 2015, 194, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.; Nguewa, P.A.; Parrondo, R.; Perez-Stable, C.; Manrique, I.; Redrado, M.; Catena, R.; Collantes, M.; Peñuelas, I.; Díaz-González, J.A.; et al. Antitumor and antiangiogenic effect of the dual EGFR and HER-2 tyrosine kinase inhibitor lapatinib in a lung cancer model. BMC Cancer 2010, 10, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.Y.; Wu, L.J.; Wu, Y.Q.; Lu, G.H.; Jiang, Z.Y.; Zhan, J.W.; Jie, Y.; Zhou, J.Y. Molecular mechanism of trifluoperazine induces apoptosis in human A549 lung adenocarcinoma cell lines. Mol. Med. Rep. 2009, 2, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Ding, Y. Potential antitumor mechanisms of phenothiazine drugs. Sci. China Life Sci. 2013, 56, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Hong, J.; Lee, J.M.; Moon, H.E.; Jeon, B.; Choi, J.; Yoon, N.A.; Paek, S.H.; Roh, E.J.; Lee, C.J.; et al. Trifluoperazine, a Well-Known Antipsychotic, Inhibits Glioblastoma Invasion by Binding to Calmodulin and Disinhibiting Calcium Release Channel IP3R. Mol. Cancer Ther. 2017, 16, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Yde, C.W.; Clausen, M.P.; Bennetzen, M.V.; Lykkesfeldt, A.E.; Mouritsen, O.G.; Guerra, B. The antipsychotic drug chlorpromazine enhances the cytotoxic effect of tamoxifen in tamoxifen-sensitive and tamoxifen-resistant human breast cancer cells. Anticancer Drugs 2009, 20, 723–735. [Google Scholar] [CrossRef]
- Lee, M.S.; Johansen, L.; Zhang, Y.; Wilson, A.; Keegan, M.; Avery, W.; Elliott, P.; Borisy, A.A.; Keith, C.T. The novel combination of chlorpromazine and pentamidine exerts synergistic antiproliferative effects through dual mitotic action. Cancer Res. 2007, 67, 11359–11367. [Google Scholar] [CrossRef] [Green Version]
- Wiklund, E.D.; Catts, V.S.; Catts, S.V.; Ng, T.F.; Whitaker, N.J.; Brown, A.J.; Lutze-Mann, L.H. Cytotoxic effects of antipsychotic drugs implicate cholesterol homeostasis as a novel chemotherapeutic target. Int. J. Cancer 2010, 126, 28–40. [Google Scholar] [CrossRef]
- Eriksson, A.; Yachnin, J.; Lewensohn, R.; Nilsson, A. DNA-dependent protein kinase is inhibited by trifluoperazine. Biochem. Biophys. Res. Commun. 2001, 283, 726–731. [Google Scholar] [CrossRef]
- Liang, W.; Yang, C.; Qi, J.; Peng, H.; Duan, J.; Liu, H.; Zheng, D. Modulating multidrug resistance through inhibiting of protein kinase C activity by phenothiazines. Chin. Sci. Bull. 1998, 43, 1179–1183. [Google Scholar] [CrossRef]
- Zhelev, Z.; Ohba, H.; Bakalova, R.; Hadjimitova, V.; Ishikawa, M.; Shinohara, Y.; Baba, Y. Phenothiazines suppress proliferation and induce apoptosis in cultured leukemic cells without any influence on the viability of normal lymphocytes. Phenothiazines and leukemia. Cancer Chemother. Pharmacol. 2004, 53, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Macgregor, A.; Attal, J.; Larue, A.; Brittner, M.; Ducasse, D.; Capdevielle, D. Antipsychotic drugs: Pro-cancer or anti-cancer? A systematic review. Med. Hypotheses 2012, 79, 38–42. [Google Scholar] [CrossRef]
- Reardon, G.T.; Rifkin, A.; Schwartz, A.; Myerson, A.; Siris, S.G. Changing patterns of neuroleptic dosage over a decade. Am. J. Psychiatry 1989, 146, 726–729. [Google Scholar] [PubMed]
- Shin, S.Y.; Choi, B.H.; Ko, J.; Kim, S.H.; Kim, Y.S.; Lee, Y.H. Clozapine, a neuroleptic agent, inhibits Akt by counteracting Ca2+/Calmodulin in PTEN-negative U-87MG human glioblastoma cells. Cell Signal. 2006, 18, 1876–1886. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E.; Altschuler, E.L. Consideration of use of phenothiazines in particular trifluorperazine for epidermal growth factor receptor associated cancers. Med. Hypotheses 2007, 69, 1074–1075. [Google Scholar] [CrossRef] [PubMed]
- Cuddapah, V.A.; Sontheimer, H. Molecular interaction and functional regulation of ClC-3 by Ca2+/calmodulin-dependent protein kinase II (CaMKII) in human malignant glioma. J. Biol. Chem. 2010, 285, 11188–11196. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Huang, S.M.; Chen, C.C.; Tsai, C.F.; Yeh, W.L.; Chou, S.J.; Hsieh, W.T.; Lu, D.Y. Ghrelin induces cell migration through GHS-R, CaMKII, AMPK, and NF-κB signaling pathway in glioma cells. J. Cell. Biochem. 2011, 112, 2931–2941. [Google Scholar] [CrossRef]
- Fedida-Metula, S.; Feldman, B.; Koshelev, V.; Levin-Gromiko, U.; Voronov, E.; Fishman, D. Lipid rafts couple store-operated Ca2+ entry to constitutive activation of PKB/Akt in a Ca2+/calmodulin-, Src- and PP2A-mediated pathway and promote melanoma tumor growth. Carcinogenesis 2012, 33, 740–750. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, M.J.; Ceña, V.; de Pablo, Y.; Llovera, M.; Comella, J.X.; Soler, R.M. Glial cell line-derived neurotrophic factor increases intracellular calcium concentration. Role of calcium/calmodulin in the activation of the phosphatidylinositol 3-kinase pathway. J. Biol. Chem. 2004, 279, 6132–6142. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Valencia, C.A.; Liu, R. Ca2+/calmodulin directly interacts with the pleckstrin homology domain of AKT1. J. Biol. Chem. 2007, 282, 25131–25140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.T.; Wu, A.T.; Chang, P.M.; Chen, K.Y.; Yang, C.N.; Yang, S.C.; Ho, C.C.; Chen, C.C.; Kuo, Y.L.; Lee, P.Y.; et al. Trifluoperazine, an antipsychotic agent, inhibits cancer stem cell growth and overcomes drug resistance of lung cancer. Am. J. Respir. Crit. Care Med. 2012, 186, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, Q.; He, J.; Lu, W. Regulation of calcium signaling in lung cancer. J. Thorac. Dis. 2010, 2, 52–56. [Google Scholar] [PubMed]
- Bergner, A.; Kellner, J.; Tufman, A.; Huber, R.M. Endoplasmic reticulum Ca2+-homeostasis is altered in small and non-small cell lung cancer cell lines. J. Exp. Clin. Cancer Res. 2009, 28, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, S.; Tokumitsu, H.; Soderling, T.R. Calcium promotes cell survival through CaM-K kinase activation of the protein-kinase-B pathway. Nature 1998, 396, 584–587. [Google Scholar] [CrossRef]
- Li, J.; Yen, C.; Liaw, D.; Podsypanina, K.; Bose, S.; Wang, S.I.; Puc, J.; Miliaresis, C.; Rodgers, L.; McCombie, R.; et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science 1997, 75, 1943–1947. [Google Scholar] [CrossRef]
- Steck, P.A.; Pershouse, M.A.; Jasser, S.A.; Yung, W.K.; Lin, H.; Ligon, A.H.; Langford, L.A.; Baumgard, M.L.; Hattier, T.; Davis, T.; et al. Identification of a candidate tumour suppressor gene, MMAC1, at chromosome 10q23.3 that is mutated in multiple advanced cancers. Nat. Genet. 1997, 15, 356–362. [Google Scholar] [CrossRef]
- Li, D.M.; Sun, H. PTEN/MMAC1/TEP1 suppresses the tumorigenicity and induces G1 cell cycle arrest in human glioblastoma cells. Proc. Natl. Acad. Sci. USA 1998, 95, 15406–15411. [Google Scholar] [CrossRef] [Green Version]
- Cantley, L.C.; Neel, B.G. New insights into tumor suppression: PTEN suppresses tumor formation by restraining the phosphoinositide 3-kinase/AKT pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 4240–4245. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, J.J.; Huang, X.Y. Orai1 and STIM1 are critical for breast tumor cell migration and metastasis. Cancer Cell 2009, 15, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Monet, M.; Lehen’kyi, V.; Gackiere, F.; Firlej, V.; Vandenberghe, M.; Roudbaraki, M.; Gkika, D.; Pourtier, A.; Bidaux, G.; Slomianny, C.; et al. Role of cationic channel TRPV2 in promoting prostate cancer migration and progression to androgen resistance. Cancer Res. 2010, 70, 1225–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seales, E.C.; Micoli, K.J.; McDonald, J.M. Calmodulin is a critical regulator of osteoclastic differentiation, function, and survival. J. Cell. Biochem. 2006, 97, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ahn, E.Y.; McKenna, M.A.; Yeo, H.; McDonald, J.M. Fas binding to calmodulin regulates apoptosis in osteoclasts. J. Biol. Chem. 2005, 280, 29964–29970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Feng, X.; McDonald, J.M. The role of calmodulin in the regulation of osteoclastogenesis. Endocrinology 2003, 144, 4536–4543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Li, S.; Shi, Q.; Yan, R.; Liu, G.; Dai, K. Calmodulin antagonists induce platelet apoptosis. Thromb. Res. 2010, 125, 340–350. [Google Scholar] [CrossRef]
- Chen, M.H.; Lin, K.J.; Yang, W.L.; Kao, Y.W.; Chen, T.W.; Chao, S.C.; Chang, P.M.; Liu, C.Y.; Tzeng, C.H.; Chao, Y.; et al. Gene expression-based chemical genomics identifies heat-shock protein 90 inhibitors as potential therapeutic drugs in cholangiocarcinoma. Cancer 2013, 119, 293–303. [Google Scholar] [CrossRef]
- Sangodkar, J.; Dhawan, N.S.; Melville, H.; Singh, V.J.; Yuan, E.; Rana, H.; Izadmehr, S.; Farrington, C.; Mazhar, S.; Katz, S.; et al. Targeting the FOXO1/KLF6 axis regulates EGFR signaling and treatment response. J. Clin. Investig. 2012, 122, 2637–2651. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.H.; Yang, W.L.; Lin, K.T.; Liu, C.H.; Liu, Y.W.; Huang, K.W.; Chang, P.M.; Lai, J.M.; Hsu, C.N.; Chao, K.M.; et al. Gene expression-based chemical genomics identifies potential therapeutic drugs in hepatocellular carcinoma. PLoS ONE 2011, 6, e27186. [Google Scholar] [CrossRef] [Green Version]
- Brosius, S.N.; Turk, A.N.; Byer, S.J.; Longo, J.F.; Kappes, J.C.; Roth, K.A.; Carroll, S.L. Combinatorial therapy with tamoxifen and trifluoperazine effectively inhibits malignant peripheral nerve sheath tumor growth by targeting complementary signaling cascades. J. Neuropathol. Exp. Neurol. 2014, 73, 1078–1090. [Google Scholar] [CrossRef] [Green Version]
- Marques, L.O.; Lima, M.S.; Soares, B.G. Trifluoperazine for schizophrenia. Cochrane Database Syst. Rev. 2004, 2004, CD003545. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Gene Set | Log-Rank p-Value | HR | 95% CI |
|---|---|---|---|
| SIG_PIP3_SIGNALING_IN_B_LYMPHOCYTES | 2.30 × 10−5 | 0.517 | 0.38–0.71 |
| REACTOME_EFFECTS_OF_PIP2_HYDROLYSIS | 7.13 × 10−4 | 0.588 | 0.43–0.8 |
| REACTOME_DAG_AND_IP3_SIGNALING | 2.63 × 10−3 | 0.624 | 0.46–0.85 |
| REACTOME_ION_HOMEOSTASIS | 7.99 × 10−3 | 0.638 | 0.46–0.89 |
| GO_LIGAND_GATED_CALCIUM_CHANNEL_ACTIVITY | 0.0127 | 0.672 | 0.49–0.92 |
| REACTOME_ANTIGEN_ACTIVATES_ B_CELL_RECEPTOR_BCR_LEADING_TO_ GENERATION_OF_SECOND_MESSENGERS | 0.0145 | 0.667 | 0.48–0.92 |
| GO_REGULATION_OF_CARDIAC_CONDUCTION | 0.0149 | 0.669 | 0.48–0.93 |
| KEGG_PHOSPHATIDYLINOSITOL_SIGNALING_SYSTEM | 0.0226 | 0.639 | 0.43–0.94 |
| REACTOME_REGULATION_OF_INSULIN_SECRETION | 0.0257 | 0.690 | 0.5–0.96 |
| REACTOME_G_PROTEIN_MEDIATED_EVENTS | 0.0372 | 0.719 | 0.53–0.98 |
| GO_CALCIUM_ION_IMPORT_INTO_CYTOSOL | 0.0391 | 0.664 | 0.45–0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.Y.; Park, H.; Yoo, H.; Kim, E.-J.; Jeon, B.; Lee, J.D.; Kang, D.; Lee, C.J.; Paek, S.H.; Roh, E.J.; et al. Trifluoperazine and Its Analog Suppressed the Tumorigenicity of Non-Small Cell Lung Cancer Cell; Applicability of Antipsychotic Drugs to Lung Cancer Treatment. Biomedicines 2022, 10, 1046. https://doi.org/10.3390/biomedicines10051046
Jeong JY, Park H, Yoo H, Kim E-J, Jeon B, Lee JD, Kang D, Lee CJ, Paek SH, Roh EJ, et al. Trifluoperazine and Its Analog Suppressed the Tumorigenicity of Non-Small Cell Lung Cancer Cell; Applicability of Antipsychotic Drugs to Lung Cancer Treatment. Biomedicines. 2022; 10(5):1046. https://doi.org/10.3390/biomedicines10051046
Chicago/Turabian StyleJeong, Joo Yeon, Haangik Park, Hong Yoo, Eun-Jin Kim, Borami Jeon, Jong Deog Lee, Dawon Kang, Changjoon Justin Lee, Sun Ha Paek, Eun Joo Roh, and et al. 2022. "Trifluoperazine and Its Analog Suppressed the Tumorigenicity of Non-Small Cell Lung Cancer Cell; Applicability of Antipsychotic Drugs to Lung Cancer Treatment" Biomedicines 10, no. 5: 1046. https://doi.org/10.3390/biomedicines10051046