
Placental Models for Evaluation of Nanocarriers as Drug Delivery Systems for Pregnancy Associated Disorders

 , ,
, ,  , and
, and
Abstract
:1. Introduction
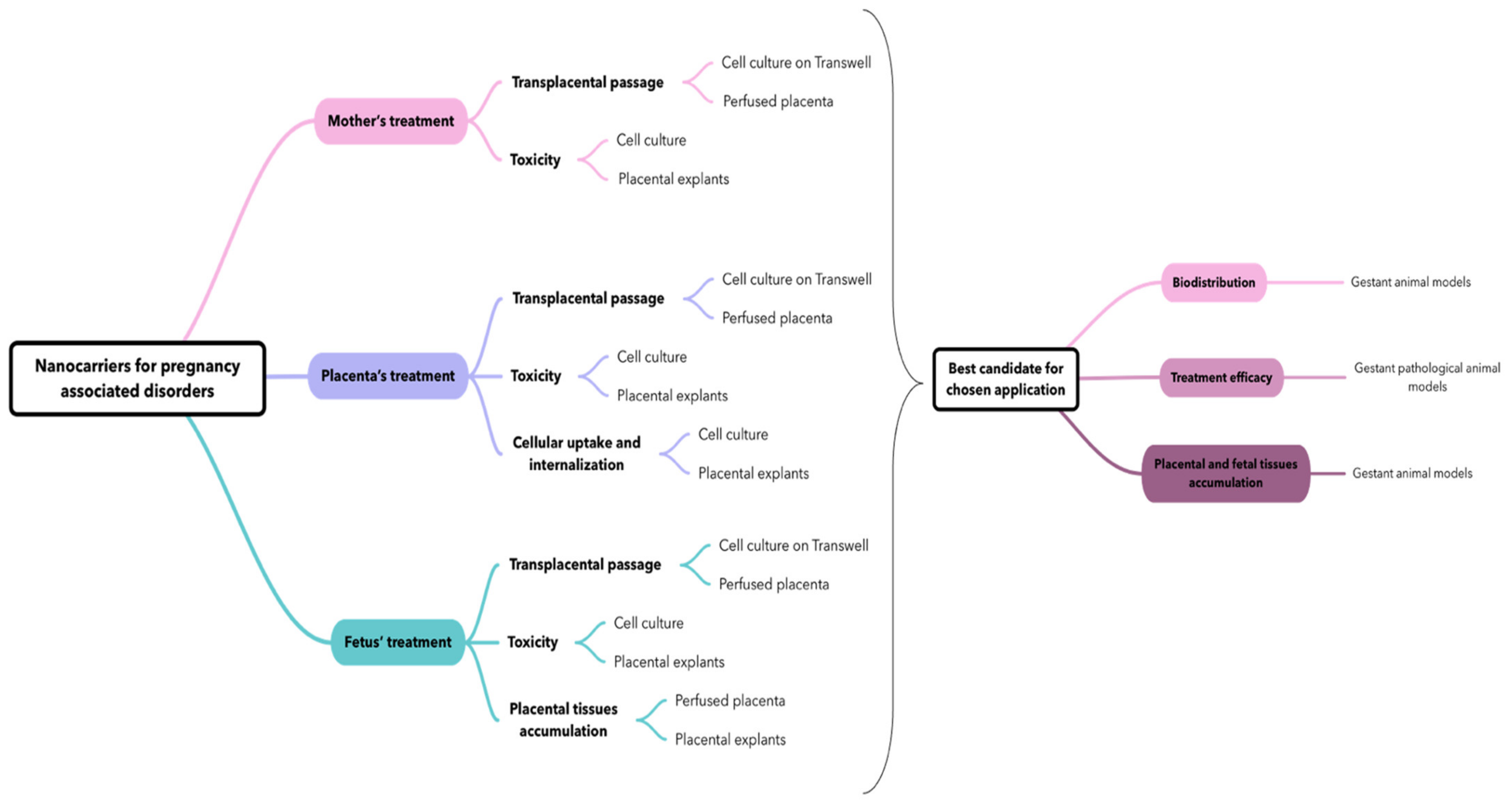
2. Nanocarriers for Pregnancy-Associated Disorders
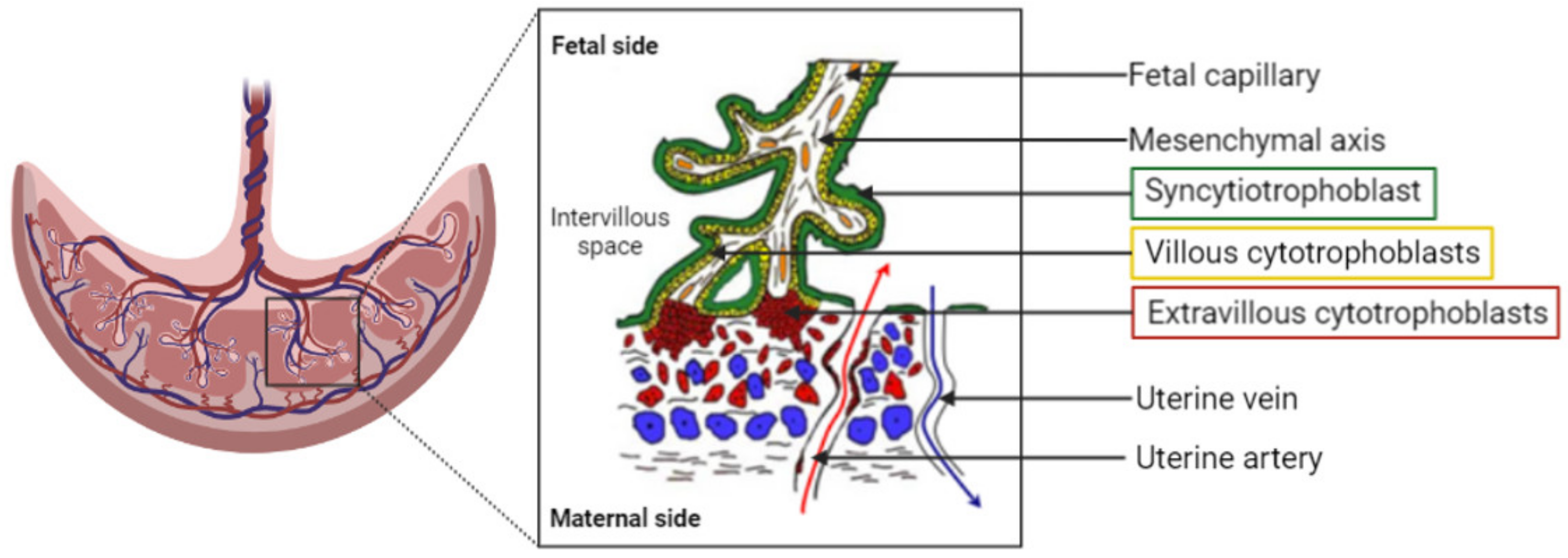
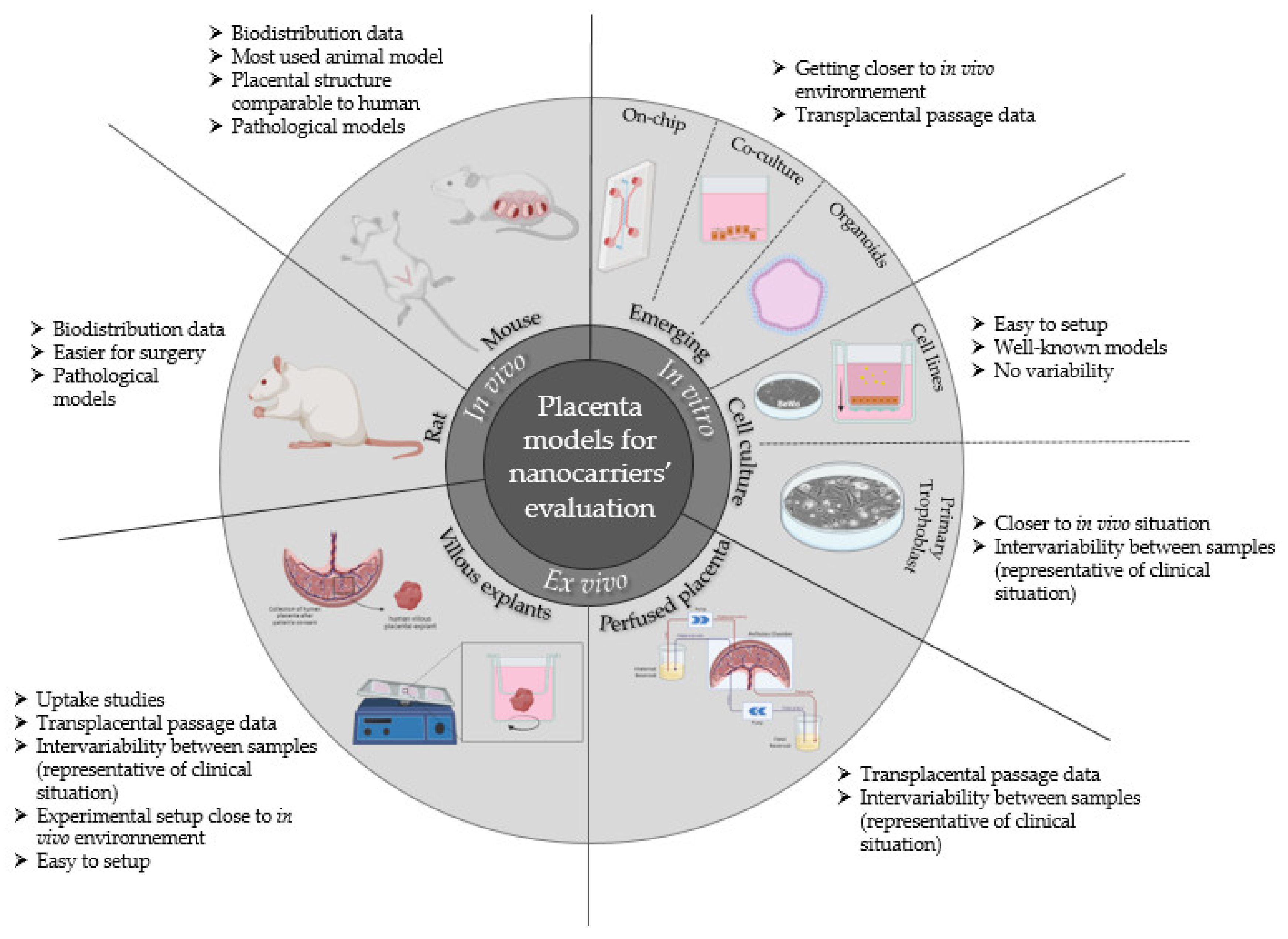
3. Which Placental Evaluation Models to Evaluate Nanocarriers
- In vivo, using living animal models;
- In vitro, using one or several cell types in culture;
- Ex vivo, using mostly organs or parts of organs outside of a living organism.
3.1. In Vivo Models
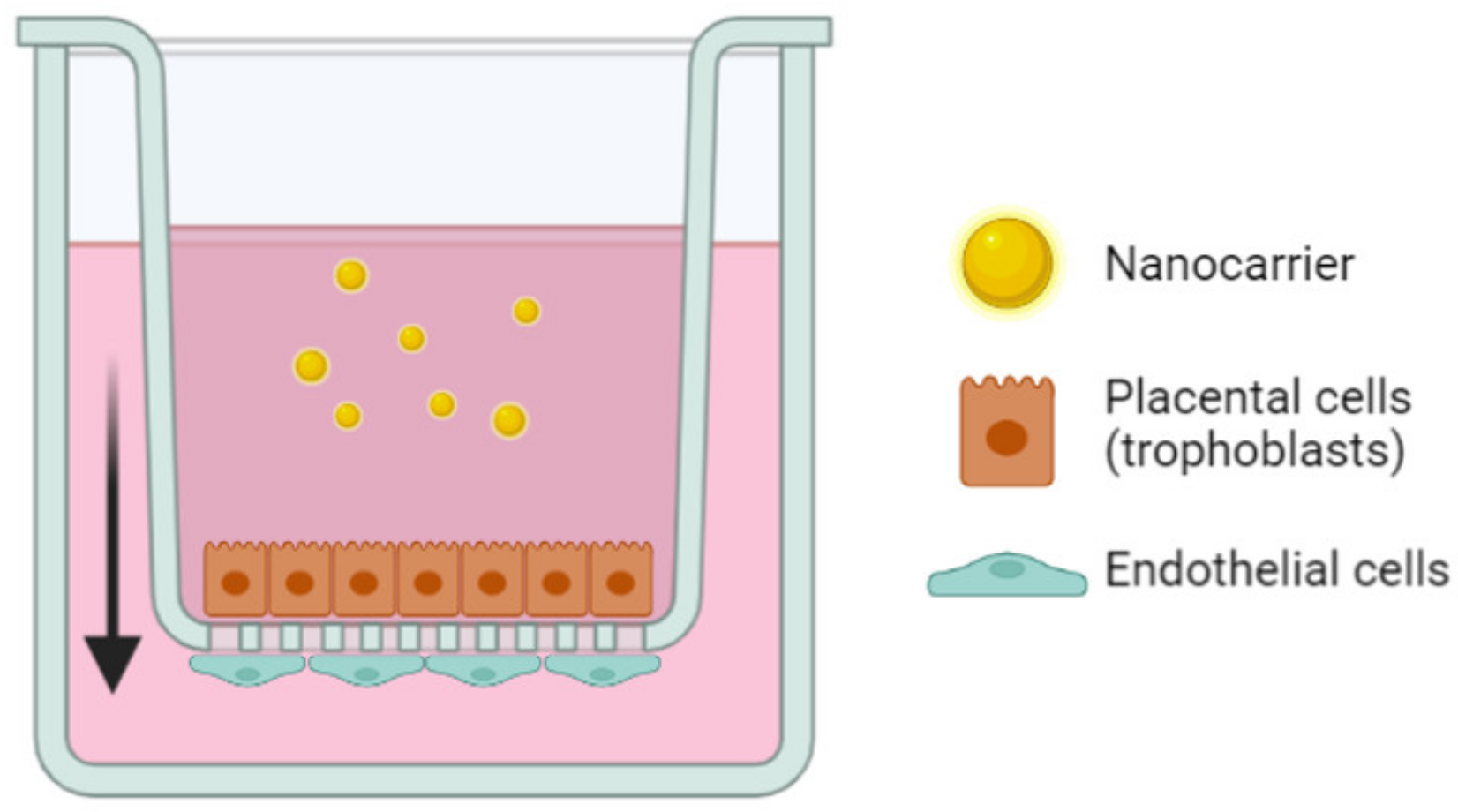
3.2. In Vitro Models
3.2.1. Cell Culture
BeWo Cell Line
JEG-3 Cell Line
HTR8/SVneo
Comparative Nanoparticle Translocation Study between BeWo, JEG-3, JAR, and ACH-3P Cell Lines
Primary Human Trophoblast Cell Culture
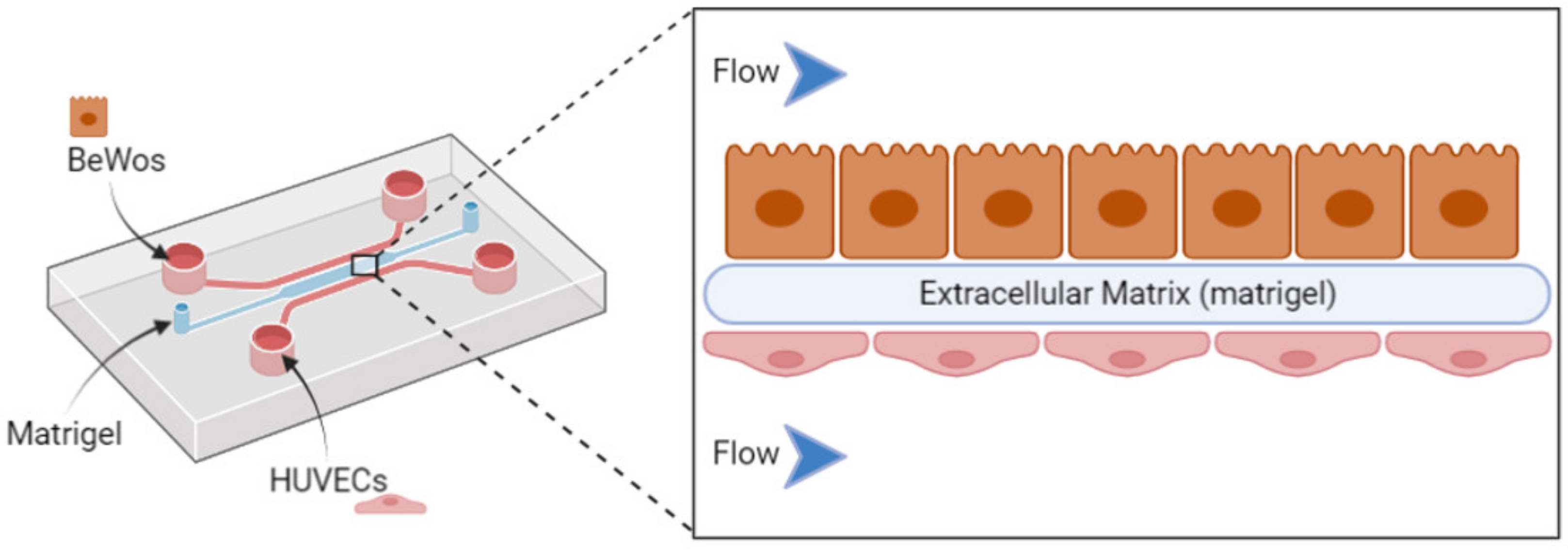
3.2.2. Emerging In Vitro Models
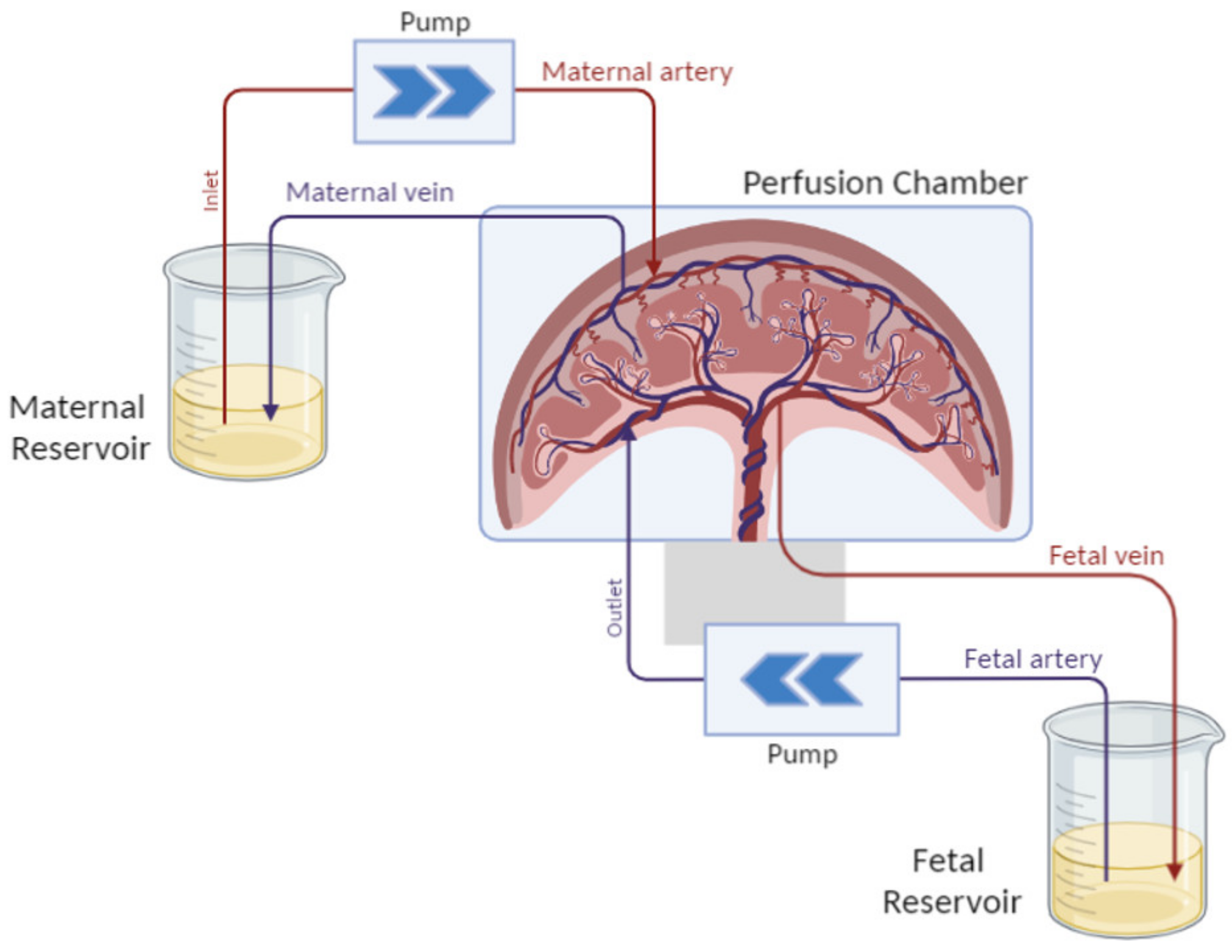
3.3. Ex Vivo Models
3.3.1. Perfused Human Placenta
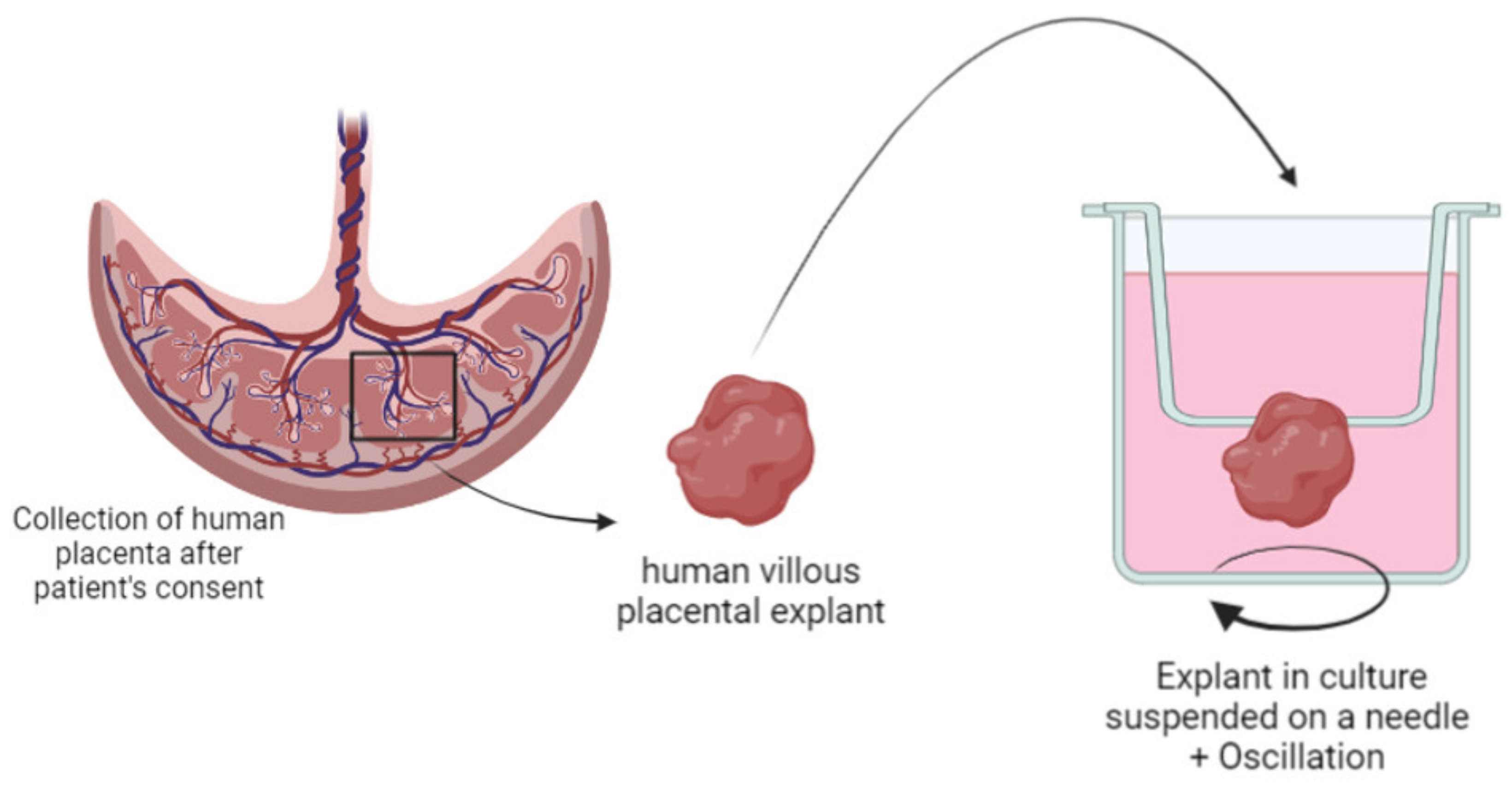
3.3.2. Human Placental Explants
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Model | Nanoparticles/Nanocarriers Evaluated | Aim of the Study | Reference |
|---|---|---|---|
| Pregnant mice | Quantum dots coated with silica or PEG | Evaluation of quantum dot transfer across the placenta | [40] |
| Silica NPs and titanium dioxide NPs (70 and 35 nm) | Evaluation of cytotoxicity of inorganic nanoparticles during pregnancy | [36] | |
| Gold NPs with surface modifications (ferritin, PEG, and citrate) | Effect of gestational age and surface modification of gold NPs on materno-fetal transfer | [32] | |
| Quantum dots | Evaluation of quantum dot placental crossing and influence on erythrocytes | [41] | |
| Gold NPs (20 and 50 nm) | Evaluation of endocytosis mechanisms at the placental barrier of gold NPs | [33] | |
| Fluorescent-labelled carboxylate-modified polystyrene NPs (20, 40, 100, 200, and 500 nm) | Evaluation of NPs crossing mouse placenta | [43] | |
| Fluorescent-labelled carboxylate of PEG-modified polystyrene NPs (50–90 nm) | Evaluation of tissue and cell-type specific compartmentalization of polystyrene NPs | [34] | |
| Silver NPs (18–20 nm) | Pregnancy outcomes after inhalation of silver NPs | [155] | |
| Amorphous carboxylate or amino-modified silica NPs (25, 50, and 115 nm) | Influence of pregnancy stages on toxic effects of silica NPs | [38] | |
| Mesoporous silica NPs | Real-time evaluation of toxicity of silica NPs during fetal development | [39] | |
| Titanium dioxide NPs | Evaluation of cytotoxicity of nanoparticles during pregnancy | [156] | |
| Fluorescently labelled liposome loaded with indomethacin (150–200 nm) | Liposomes used to prevent indomethacin passage to fetus | [44] | |
| Tumor-homing peptide CGKRK and iRGD-coated liposomes loaded with carboxyfluorescein or insulin growth factor 2 (IGF-II) | Evaluation of placenta-targeting liposomes and drug delivery | [49] | |
| Liposomes coated with oxytocin receptor antagonist loaded with indomethacin | Evaluation of uterus-targeting liposomes in preterm labor management | [45] | |
| Liposomal gadolinium NPs | Evaluation of transplacental passage of liposomal Gd | [47] | |
| CSA-binding peptide-conjugated NPs loaded with indocyanine green or methotrexate | Placenta-specific drug delivery with trophoblast-targeted NPs | [107] | |
| Pregnant mouse model of intrauterine inflammation | Gold NPs (3, 13, and 32 nm) | Evaluation of transfer of gold NPs in intrauterine inflammation context | [51] |
| Pregnant rats | Pegylated gold NPs | Evaluation of PEGylated gold NP placental transfer | [31] |
| Magnetic core mesoporous silica NPs (60 nm) | Evaluation of cytotoxicity, genotoxicity, transplacental transfer, and tissue disposition of silica NPs | [37] | |
| Radio-labelled gold NPs (1.4, 18, and 80 nm) | Influence of the size on the placental translocation of gold NPs | [30] | |
| RGD-modified cationic liposomes complexed with siRNA | Development of preeclamspia-like mouse model with siRNA transfer | [54] | |
| Preeclamspia pregnant rat model (injection of TNF-α) | Poly-amidoamine (PAMAM) NPs loaded with siRNA to knock down sFlt1 | Treatment of preeclampsia by administration of PAMAM NPs for siRNA delivery | [53] |
| Experimental Model | Nanoparticles/Nanocarriers Evaluated | Aim of the Study | Reference |
|---|---|---|---|
| BeWo | Alpha-Fe2O3 NPs of 15, 50, and 78 nm | Evaluation of potential toxicity of iron NPs on placental integrity | [92] |
| Silica NPs | Evaluation of fetal transfer of silica NPs | [91] | |
| Dexamethasone-loaded polymeric NPs | Development of a treatment for fetal congenital adrenal hyperplasia | [96] | |
| Digoxin-loaded pegylated polymeric NPs | Development of a treatment for fetal arrythmia | [95] | |
| Oxacarbazepine-loaded polymeric NPs | Permeability studies of an antiepileptic drug across an in vitro model of human placental trophoblasts | [157] | |
| Liposomal doxorubicin | Uptake and toxicity of doxorubicin free and loaded into liposomes for therapies for pregnant women | [93] | |
| Clonazepam-loaded micelle-like NPs | Cellular uptake and cytotoxicity studies on placental and brain endothelial cells | [94] | |
| BeWo Transwell® | Fluorescent polystyrene NPs (50 and 100 nm) | Penetration and transcellular transport across BeWo monolayer on Transwell® of 3 µm pore size) | [97] |
| Iron dioxide and silica NPs (8, 25, and 50 nm) | Comparison between toxicity, uptake, and transport of several inorganic nanoparticles by placental cells | [71] | |
| Positively and negatively charged polystyrene NPs | Translocation studies of NPs across BeWo cell layer compared to amoxicillin, a reference compound | [98] | |
| Fluorescent pullulan acetate NPs (200–300 nm) | Study of endocytotic mechanisms of NPS translocation across BeWo cells | [99] | |
| JEG-3 | NPs of Fe3O4-dextran-anti-β-human chorionic gonadotropin (HCG) carrying heparanase (Hpa) antisense oligodeoxynucleotide (ASODN) | Evaluation of the inhibitory effect of NPs on JEG-3 invasion and proliferation ability | [106] |
| Polymeric NPs functionalized with CSA-binding peptide derived from the VAR2CSA protein | Evaluation of specific targeting of the placental cells by functionalized NPs and delivery of doxorubicin | [107] | |
| HTR-8/SVNeo | Magnetic NPs coated with specific leucocyte antigens | Isolation of extra-villous cytotrophoblasts in order to retrieve the fetal antigen in a non-invasive way for prenatal diagnosis | [114] |
| PAMAM NPs loaded with siRNA | Synthesis and evaluation of siRNA NPs against sFlt-1 to treat preeclampsia | [53] | |
| Primary cytotrophoblast | Carboxyfluorescein-encapsulated neutral, anionic, and cationic liposomes | Evaluation of the placental uptake of the liposomes and carboxyfluoresceine | [146] |
| Dendritic polyglycerol NPs (5 nm) | Evaluation of the placental uptake of NPs and trophoblast integrity and endocrine function | [100] | |
| Rhodamine-labelled siRNA complexed with liposomes | Evaluation of internalization of siRNA delivered using liposomal formulation in placental cells | [119] | |
| Co-culture | Polystyrene NPs | Evaluation of NP retention in trophoblastic and endothelial cells and translocation across the cellular layers | [90] |
| Cationic, anionic, and neutral magnetic NPs | Study of the uptake and fate of magnetic NPs through a blood–placenta barrier model | [126] | |
| Placenta-on-a-chip | Titanium dioxide NPs | Evaluation of the impact of environmental NPs on placental cells | [136] |
| 3D co-culture | Gold NPs | Placental uptake of gold NPs | [131] |
| Experimental Model | Nanoparticles/Nanocarriers Evaluated | Aim of the Study | Reference |
|---|---|---|---|
| Dually perfused human placenta | Gold NPs (10–30 nm) | Study of the kinetics of gold NP passage through the human placentas | [144] |
| Silica NPs | Study of the kinetic of silica NP passage through the human placenta | [91] | |
| Neutral, cationic, and anionic liposomes loaded with carboxyfluoresceine | Evaluation of the charge influence of the liposome surface on the delivery kinetics through the materno-fetal interface of a small hydrophilic molecule | [146] | |
| Fluorescently labelled PAMAM dendrimers (16 kDa) | Study of the potential transfer of NPs across the human placenta | [145] | |
| Cationic small unilamellar liposomes with sterylamine of lecithin | Evaluation of the influence of the lipid composition in liposomes on the delivery kinetics through the materno-fetal interface of warfarin | [147] | |
| PEGylated fluorescent liposomes loaded with carboxyfluoresceine | Evaluation of fluorescent PEGylated liposomes and fluorescent compound passage kinetics across the human placenta | [151] | |
| Term placental explants | EnGeneIC Delivery Vehicles loaded with doxorubicin targeting the EGFR at the outer cell layer of the placenta | NPs targeting the placenta to treat ectopic pregnancies | [153] |
| PEGylated fluorescent liposomes loaded with carboxyfluoresceine (100 nm) | Evaluation of fluorescent PEGylated liposomes and fluorescent compound uptake by placental cells | [151] | |
| Fluorescent liposomes loaded with fluorescent siRNA (150 nm) | Evaluation of fluorescent PEGylated liposomes and fluorescent compound uptake by placental cells | [152] | |
| First-trimester placental explants | CNKGLRNK-decorated liposomes loaded with a vasodilator SE175 | NPs targeting the uterine vessels to deliver a vasodilator | [46] |
| Fluorescently labelled dendritic polyglycerol NPs (5 nm) | Assessment of NP uptake by placental cells | [99] |
References
- Scaffidi, J.; Mol, B.W.; Keelan, J.A. The pregnant women as a drug orphan: A global survey of registered clinical trials of pharmacological interventions in pregnancy. BJOG Int. J. Obstet. Gynaecol. 2017, 124, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, M. Transplacental transport of nanomaterials. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2009, 1, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Borgelt, L.M.; Hart, F.M.; Bainbridge, J.L. Epilepsy during pregnancy: Focus on management strategies. Int. J. Women’s Health 2016, 8, 505–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phipps, E.A.; Thadhani, R.; Benzing, T.; Karumanchi, S.A. Pre-eclampsia: Pathogenesis, novel diagnostics and therapies. Nat. Rev. Nephrol. 2019, 15, 275–289. [Google Scholar] [CrossRef]
- Lupattelli, A.; Spigset, O.; Twigg, M.J.; Zagorodnikova, K.; Mårdby, A.C.; Moretti, M.E.; Drozd, M.; Panchaud, A.; Hämeen-Anttila, K.; Rieutord, A.; et al. Medication use in pregnancy: A cross-sectional, multinational web-based study. BMJ Open 2014, 4, e004365. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.D. Drug delivery during pregnancy: How can nanomedicine be used? Ther. Deliv. 2017, 8, 1023–1025. [Google Scholar] [CrossRef]
- Chambers, C.D.; Polifka, J.E.; Friedman, J.M. Drug safety in pregnant women and their babies: Ignorance not bliss. Clin. Pharmacol. Ther. 2008, 83, 181–183. [Google Scholar] [CrossRef]
- Vargesson, N. Thalidomide-induced teratogenesis: History and mechanisms. Birth Defects Res. Part C Embryo Today Rev. 2015, 105, 140–156. [Google Scholar] [CrossRef] [Green Version]
- Sheffield, J.S.; Siegel, D.; Mirochnick, M.; Heine, R.P.; Nguyen, C.; Bergman, K.L.; Savic, R.M.; Long, J.; Dooley, K.E.; Nesin, M. Designing drug trials: Considerations for pregnant women. Clin. Infect. Dis. 2014, 59, S437–S444. [Google Scholar] [CrossRef]
- Blehar, M.C.; Spong, C.; Grady, C.; Goldkind, S.F.; Sahin, L.; Clayton, J.A. Enrolling pregnant women: Issues in clinical research. Womens. Health Issues 2013, 23, e39–e45. [Google Scholar] [CrossRef] [Green Version]
- Chamundeeswari, M.; Jeslin, J.; Verma, M.L. Nanocarriers for drug delivery applications. Environ. Chem. Lett. 2019, 17, 849–865. [Google Scholar] [CrossRef]
- Costantine, M.M. Physiologic and pharmacokinetic changes in pregnancy. Front. Pharmacol. 2014, 5, 65. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Tan, E.L. Alterations in physiology and anatomy during pregnancy. Best Pract. Res. Clin. Obstet. Gynaecol. 2013, 27, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Hodge, L.S.; Tracy, T.S. Alterations in drug disposition during pregnancy. Expert Opin. Drug Metab. Toxicol. 2007, 3, 557–571. [Google Scholar] [CrossRef]
- Feghali, M.; Venkataramanan, R.; Caritis, S. Pharmacokinetics of drugs in pregnancy. Semin. Perinatol. 2015, 39, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.A.; Gilboa, S.M.; Werler, M.M.; Kelley, K.E.; Louik, C.; Hernández-Díaz, S. Medication use during pregnancy, with particular focus on prescription drugs: 1976–2008. Am. J. Obstet. Gynecol. 2011, 205, 51.e1–51.e8. [Google Scholar] [CrossRef] [Green Version]
- Shields, K.E.; Lyerly, A.D. Exclusion of pregnant women from industry-sponsored clinical trials. Obstet. Gynecol. 2013, 122, 1077–1081. [Google Scholar] [CrossRef]
- Wilczewska, A.Z.; Niemirowicz, K.; Markiewicz, K.H.; Car, H. Nanoparticles as drug delivery systems. Pharmacol. Rep. 2012, 64, 1020–1037. [Google Scholar] [CrossRef]
- Suk, J.S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L.M. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv. Drug Deliv. Rev. 2016, 99, 28–51. [Google Scholar] [CrossRef] [Green Version]
- Attia, M.F.; Anton, N.; Wallyn, J.; Omran, Z.; Vandamme, T.F. An overview of active and passive targeting strategies to improve the nanocarriers efficiency to tumour sites. J. Pharm. Pharmacol. 2019, 71, 1185–1198. [Google Scholar] [CrossRef] [Green Version]
- Moros, M.; Mitchell, S.G.; Grazú, V.; de la Fuente, J.M. The fate of nanocarriers as nanomedicines in vivo: Important considerations and biological barriers to overcome. Curr. Med. Chem. 2013, 20, 2759–2778. [Google Scholar] [CrossRef]
- Zhao, Z.; Ukidve, A.; Krishnan, V.; Mitragotri, S. Effect of physicochemical and surface properties on in vivo fate of drug nanocarriers. Adv. Drug Deliv. Rev. 2019, 143, 3–21. [Google Scholar] [CrossRef]
- Couvreur, P.; Vauthier, C. Nanotechnology: Intelligent design to treat complex disease. Pharm. Res. 2006, 23, 1417–1450. [Google Scholar] [CrossRef]
- Muoth, C.; Aengenheister, L.; Kucki, M.; Wick, P.; Buerki-Thurnherr, T. Nanoparticle transport across the placental barrier: Pushing the field forward! Nanomedicine 2016, 11, 941–957. [Google Scholar] [CrossRef]
- Refuerzo, J.S.; Longo, M.; Godin, B. Targeted nanoparticles in pregnancy: A new frontier in perinatal therapeutics. Am. J. Obstet. Gynecol. 2017, 216, 204–205. [Google Scholar] [CrossRef] [Green Version]
- Alhareth, K. How should we plan the future of nanomedicine for cancer diagnosis and therapy? Int. J. Pharm. 2017, 532, 657–659. [Google Scholar] [CrossRef]
- Valero, L.; Alhareth, K.; Gil, S.; Lecarpentier, E.; Tsatsaris, V.; Mignet, N.; Fournier, T.; Andrieux, K. Nanomedicine as a potential approach to empower the new strategies for the treatment of preeclampsia. Drug Discov. Today 2018, 23, 1099–1107. [Google Scholar] [CrossRef]
- Nikitina, L.; Dohr, G.; Juch, H. Studying nanoparticle interaction with human placenta: Festina lente! Nanotoxicology 2015, 9, 133–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, L.K. Nanomedicine; Future Medicine Ltd.: London, UK, 2016; pp. 2235–2238. [Google Scholar]
- Semmler-Behnke, M.; Lipka, J.; Wenk, A.; Hirn, S.; Schäffler, M.; Tian, F.; Schmid, G.; Oberdörster, G.; Kreyling, W.G. Size dependent translocation and fetal accumulation of gold nanoparticles from maternal blood in the rat. Part. Fibre Toxicol. 2014, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Tsyganova, N.A.; Khairullin, R.M.; Terentyuk, G.S.; Khlebtsov, B.N.; Bogatyrev, V.A.; Dykman, L.A.; Erykov, S.N.; Khlebtsov, N.G. Penetration of pegylated gold nanoparticles through rat placental barrier. Bull. Exp. Biol. Med. 2014, 157, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sun, C.; Fan, Z.; Tian, X.; Yan, L.; Du, L.; Liu, Y.; Chen, C.; Liang, X.J.; Anderson, G.J.; et al. Effects of gestational age and surface modification on materno-fetal transfer of nanoparticles in murine pregnancy. Sci. Rep. 2012, 2, 847. [Google Scholar] [CrossRef] [Green Version]
- Rattanapinyopituk, K.; Shimada, A.; Morita, T.; Sakurai, M.; Asano, A.; Hasegawa, T.; Inoue, K.; Takano, H. Demonstration of the clathrin- and caveolin-mediated endocytosis at the maternal-fetal barrier in mouse placenta after intravenous administration of gold nanoparticles. J. Vet. Med. Sci. 2014, 76, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Kenesei, K.; Murali, K.; Czéh, Á.; Piella, J.; Puntes, V.; Madarász, E. Enhanced detection with spectral imaging fluorescence microscopy reveals tissue- and cell-type-specific compartmentalization of surface-modified polystyrene nanoparticles. J. Nanobiotechnol. 2016, 14, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.T.; Chen, J.H.; Hang, L.L.; Liu, S.S.; Zhong, M. Activation of PAR-1/NADPH Oxidase/ROS signaling pathways is crucial for the thrombin-induced sFlt-1 production in extravillous trophoblasts: Possible involvement in the pathogenesis of preeclampsia. Cell. Physiol. Biochem. 2015, 35, 1654–1662. [Google Scholar] [CrossRef]
- Yamashita, K.; Yoshioka, Y.; Higashisaka, K.; Mimura, K.; Morishita, Y.; Nozaki, M.; Yoshida, T.; Ogura, T.; Nabeshi, H.; Nagano, K.; et al. Silica and titanium dioxide nanoparticles cause pregnancy complications in mice. Nat. Nanotechnol. 2011, 6, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.R.; Helal-Neto, E.; Paumgartten, F.; Felzenswalb, I.; Araujo-Lima, C.F.; Martínez-Máñez, R.; Santos-Oliveira, R. Cytotoxicity, genotoxicity, transplacental transfer and tissue disposition in pregnant rats mediated by nanoparticles: The case of magnetic core mesoporous silica nanoparticles. Artif. Cell. Nanomed. Biotechnol. 2018, 46, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Pietroiusti, A.; Vecchione, L.; Malvindi, M.A.; Aru, C.; Massimiani, M.; Camaioni, A.; Magrini, A.; Bernardini, R.; Sabella, S.; Pompa, P.P.; et al. Relevance to investigate different stages of pregnancy to highlight toxic effects of nanoparticles: The example of silica. Toxicol. Appl. Pharmacol. 2018, 342, 60–68. [Google Scholar] [CrossRef]
- Sweeney, S.; Adamcakova-Dodd, A.; Thorne, P.S.; Assouline, J.G. Multifunctional nanoparticles for real-time evaluation of toxicity during fetal development. PLoS ONE 2018, 13, e0192474. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.; Wu, Q.; Yang, H.; Yuan, R.; Hou, S.; Yang, Y.; Zou, Y.; Xu, S.; Xu, K.; Ji, A.; et al. Transfer of Quantum Dots from Pregnant Mice to Pups Across the Placental Barrier. Small 2010, 6, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, S.; Qu, G.; Liu, S. The capability of quantum dots in crossing the placental barrier and the potential influence on erythrocytes. J. Nanosci. Nanotechnol. 2013, 13, 6529–6532. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, F.; Sun, X.; Choi, K.Y.; Niu, G.; Zhang, G.; Guo, J.; Lee, S.; Chen, X. The genotype-dependent influence of functionalized multiwalled carbon nanotubes on fetal development. Biomaterials 2014, 35, 856–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.P.; Hsieh, P.C.H.; Chen, C.Y.; Wang, T.Y.; Chen, P.C.; Liu, C.C.; Chen, C.C.; Chen, C.P. Nanoparticles can cross mouse placenta and induce trophoblast apoptosis. Placenta 2015, 36, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Refuerzo, J.S.; Alexander, J.F.; Leonard, F.; Leon, M.; Longo, M.; Godin, B. Liposomes: A nanoscale drug carrying system to prevent indomethacin passage to the fetus in a pregnant mouse model. Am. J. Obstet. Gynecol. 2015, 212, 508.e1–508.e7. [Google Scholar] [CrossRef] [PubMed]
- Refuerzo, J.S.; Leonard, F.; Bulayeva, N.; Gorenstein, D.; Chiossi, G.; Ontiveros, A.; Longo, M.; Godin, B. Uterus-targeted liposomes for preterm labor management: Studies in pregnant mice. Sci. Rep. 2016, 6, 34710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cureton, N.; Korotkova, I.; Baker, B.; Greenwood, S.; Wareing, M.; Kotamraju, V.R.; Teesalu, T.; Cellesi, F.; Tirelli, N.; Ruoslahti, E.; et al. Selective Targeting of a Novel Vasodilator to the Uterine Vasculature to Treat Impaired Uteroplacental Perfusion in Pregnancy. Theranostics 2017, 7, 3715–3731. [Google Scholar] [CrossRef]
- Shetty, A.N.; Pautler, R.; Ghagahda, K.; Rendon, D.; Gao, H.; Starosolski, Z.; Bhavane, R.; Patel, C.; Annapragada, A.; Yallampalli, C.; et al. A liposomal Gd contrast agent does not cross the mouse placental barrier. Sci. Rep. 2016, 6, 27863. [Google Scholar] [CrossRef]
- Zhang, B.; Liang, R.; Zheng, M.; Cai, L.; Fan, X. Surface-functionalized nanoparticles as efficient tools in targeted therapy of pregnancy complications. Int. J. Mol. Sci. 2019, 20, 3642. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Ndifon, C.; Lui, S.; Widdows, K.; Kotamraju, V.R.; Agemy, L.; Teesalu, T.; Glazier, J.D.; Cellesi, F.; Tirelli, N.; et al. Tumor-homing peptides as tools for targeted delivery of payloads to the placenta. Sci. Adv. 2016, 2, e1600349. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Tan, L.; Yu, Y.; Wang, B.; Chen, Z.; Han, J.; Li, M.; Chen, J.; Xiao, T.; Ambati, B.K.; et al. Placenta-specific drug delivery by trophoblast-targeted nanoparticles in mice. Theranostics 2018, 8, 2765–2781. [Google Scholar] [CrossRef]
- Tian, X.; Zhu, M.; Du, L.; Wang, J.; Fan, Z.; Liu, J.; Zhao, Y.; Nie, G. Intrauterine inflammation increases materno-fetal transfer of gold nanoparticles in a size-dependent manner in murine pregnancy. Small 2013, 9, 2432–2439. [Google Scholar] [CrossRef]
- Lei, J.; Rosenzweig, J.M.; Mishra, M.K.; Alshehri, W.; Brancusi, F.; McLane, M.; Almalki, A.; Bahabry, R.; Arif, H.; Rozzah, R.; et al. Maternal dendrimer-based therapy for inflammation-induced preterm birth and perinatal brain injury. Sci. Rep. 2017, 7, 6106. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jia, J.; Guo, X.; Chen, R.; Feng, L. Modulating circulating sFlt1 in an animal model of preeclampsia using PAMAM nanoparticles for siRNA delivery. Placenta 2017, 58, 1–8. [Google Scholar] [CrossRef]
- Yu, Q.; Qiu, Y.; Wang, X.; Tang, J.; Liu, Y.; Mei, L.; Li, M.; Yang, M.; Tang, L.; Gao, H.; et al. Efficient siRNA transfer to knockdown a placenta specific lncRNA using RGD-modified nano-liposome: A new preeclampsia-like mouse model. Int. J. Pharm. 2018, 546, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Doridot, L.; Passet, B.; Méhats, C.; Rigourd, V.; Barbaux, S.; Ducat, A.; Mondon, F.; Vilotte, M.; Castille, J.; Breuiller-Fouché, M.; et al. Preeclampsia-like symptoms induced in mice by fetoplacental expression of STOX1 are reversed by aspirin treatment. Hypertension 2013, 61, 662–668. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; LaMarca, B.; Reckelhoff, J.F. A model of preeclampsia in rats: The reduced uterine perfusion pressure (RUPP) model. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1–H8. [Google Scholar] [CrossRef]
- Fushima, T.; Sekimoto, A.; Minato, T.; Ito, T.; Oe, Y.; Kisu, K.; Sato, E.; Funamoto, K.; Hayase, T.; Kimura, Y.; et al. Reduced uterine perfusion pressure (RUPP) model of preeclampsia in Mice. PLoS ONE 2016, 11, e0155426. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.S.; Levasseur, J.; Ganguly, E.; Quon, A.; Kirschenman, R.; Dyck, J.R.B.; Fraser, G.M.; Davidge, S.T. Characterisation of the Selective Reduced Uteroplacental Perfusion (sRUPP) Model of Preeclampsia. Sci. Rep. 2019, 9, 9565. [Google Scholar] [CrossRef]
- Erlandsson, L.; Nääv, Å.; Hennessy, A.; Vaiman, D.; Gram, M.; Åkerström, B.; Hansson, S.R. Inventory of Novel Animal Models Addressing Etiology of Preeclampsia in the Development of New Therapeutic/Intervention Opportunities. Am. J. Reprod. Immunol. 2016, 75, 402–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wooding, P.; Burton, G. Implantation, maternofetal exchange and vascular relationships. In Comparative Placentation; Springer: Berlin/Heidelberg, Germany, 2008; pp. 47–81. [Google Scholar]
- Rinkenberger, J.; Werb, Z. The labyrinthine placenta. Nat. Genet. 2000, 25, 248–250. [Google Scholar] [CrossRef]
- Cross, J.C. Adaptability and potential for treatment of placental functions to improve embryonic development and postnatal health. Reprod. Fertil. Dev. 2016, 28, 75–82. [Google Scholar] [CrossRef]
- Bryda, E.C. The Mighty Mouse: The impact of rodents on advances in biomedical research. Mo. Med. 2013, 110, 207–211. [Google Scholar] [PubMed]
- Carter, A.M. Animal Models of Human Placentation—A Review. Placenta 2007, 28, S41–S47. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.D.; Alstrup, A.K.O.; Duvald, C.S.; Mikkelsen, E.F.R.; Vendelbo, M.H.; Ovesen, P.G.; Pedersen, M. Animal models of fetal medicine and obstetrics. In Experimental Animal Models of Human Diseases—An Effective Therapeutic Strategy; InTech: London, UK, 2018. [Google Scholar]
- Swanson, A.M.; David, A.L. Animal models of fetal growth restriction: Considerations for translational medicine. Placenta 2015, 36, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigsby, P.L. Animal Models to Study Placental Development and Function throughout Normal and Dysfunctional Human Pregnancy. Semin. Reprod. Med. 2016, 34, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Rothbauer, M.; Patel, N.; Gondola, H.; Siwetz, M.; Huppertz, B.; Ertl, P. A comparative study of five physiological key parameters between four different human trophoblast-derived cell lines. Sci. Rep. 2017, 7, 5892. [Google Scholar] [CrossRef]
- Kou, L.; Sun, J.; Zhai, Y.; He, Z. The endocytosis and intracellular fate of nanomedicines: Implication for rational design. Asian J. Pharm. Sci. 2013, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Kim, W.K.; Lee, K.; Son, M.J.; Kwak, M.; Chang, W.S.; Min, J.K.; Song, N.W.; Lee, J.; Bae, K.H. A reliable approach for assessing size-dependent effects of silica nanoparticles on cellular internalization behavior and cytotoxic mechanisms. Int. J. Nanomed. 2019, 14, 7375–7387. [Google Scholar] [CrossRef] [Green Version]
- Correia Carreira, S.; Walker, L.; Paul, K.; Saunders, M. The toxicity, transport and uptake of nanoparticles in the in vitro BeWo b30 placental cell barrier model used within NanoTEST. Nanotoxicology 2015, 9, 66–78. [Google Scholar] [CrossRef]
- Orendi, K.; Kivity, V.; Sammar, M.; Grimpel, Y.; Gonen, R.; Meiri, H.; Lubzens, E.; Huppertz, B. Placental and trophoblastic in vitro models to study preventive and therapeutic agents for preeclampsia. Placenta 2011, 32, S49–S54. [Google Scholar] [CrossRef]
- McCormick, J.; Whitley, G.S.J.; Le Bouteiller, P.; Cartwright, J.E. Soluble HLA-G regulates motility and invasion of the trophoblast-derived cell line SGHPL-4. Hum. Reprod. 2009, 24, 1339–1345. [Google Scholar] [CrossRef] [Green Version]
- Reiter, J.L.; Drendel, H.M.; Chakraborty, S.; Schellinger, M.M.; Lee, M.J.; Mor, G. Cytogenetic features of human trophoblast cell lines SWAN-71 and 3A-subE. Placenta 2017, 52, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiden, U.; Prutsch, N.; Gauster, M.; Weiss, U.; Frank, H.G.; Schmitz, U.; Fast-Hirsch, C.; Hengstschläger, M.; Pötgens, A.; Rüben, A.; et al. The first trimester human trophoblast cell line ACH-3P: A novel tool to study autocrine/paracrine regulatory loops of human trophoblast subpopulations—TNF-α stimulates MMP15 expression. BMC Dev. Biol. 2007, 7, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.H.F. Endocrine cell lines from the placenta. Mol. Cell. Endocrinol. 2004, 228, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Menezes, V.; Malek, A.; Keelan, J.A. Nanoparticulate Drug Delivery in Pregnancy: Placental Passage and Fetal Exposure. Curr. Pharm. Biotechnol. 2012, 12, 731–742. [Google Scholar] [CrossRef]
- Pattillo, R.A.; Gey, G.O. The Establishment of a Cell Line of Human Hormone-synthesizing Trophoblastic Cells in Vitro. Cancer Res. 1968, 28, 1231–1236. [Google Scholar]
- Abaidoo, C.S.; Warren, M.A.; Andrews, P.W.; Boateng, K.A. Una evaluación cuantitativa de las características morfológicas de las células BeWo como un modelo in vitro de las células de trofoblasto humano. Int. J. Morphol. 2010, 28, 1047–1058. [Google Scholar] [CrossRef]
- Grümmer, R.; Hohn, H.-P.; Denker, H.-W. Choriocarcinoma Cell Spheroids: An In Vitro Model for the Human Trophoblast. In Trophoblast Invasion and Endometrial Receptivity; Springer: Boston, MA, USA, 1990; pp. 97–111. [Google Scholar]
- Bode, C.J.; Jin, H.; Rytting, E.; Silverstein, P.S.; Young, A.M.; Audus, K.L. In vitro models for studying trophoblast transcellular transport. Methods Mol. Med. 2006, 122, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, K.; Nasu, K.; Arima, K.; Sugano, T.; Narahara, H.; Miyakawa, I. Production of interleukin (IL)-6 and IL-8 by a choriocarcinoma cell line, BeWo. Placenta 2000, 21, 354–360. [Google Scholar] [CrossRef]
- Abou-Kheir, W.; Barrak, J.; Hadadeh, O.; Daoud, G. HTR-8/SVneo cell line contains a mixed population of cells. Placenta 2017, 50, 1–7. [Google Scholar] [CrossRef]
- Wice, B.; Menton, D.; Geuze, H.; Schwartz, A.L. Modulators of cyclic AMP metabolism induce syncytiotrophoblast formation in vitro. Exp. Cell Res. 1990, 186, 306–316. [Google Scholar] [CrossRef]
- Hannan, N.J.; Paiva, P.; Dimitriadis, E.; Salamonsen, L.A. Models for Study of Human Embryo Implantation: Choice of Cell Lines? Biol. Reprod. 2010, 82, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, R.J.; Gomes, A.O.; Franco, P.S.; Pereira, A.S.; Milian, I.C.B.; Ribeiro, M.; Fiorenzani, P.; dos Santos, M.C.; Mineo, J.R.; da Silva, N.M.; et al. Enrofloxacin and toltrazuril are able to reduce Toxoplasma gondii growth in human bewo trophoblastic cells and villous explants from human third trimester pregnancy. Front. Cell. Infect. Microbiol. 2017, 7, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, M.; Franco, P.S.; Lopes-Maria, J.B.; Angeloni, M.B.; de Freitas Barbosa, B.; de Oliveira Gomes, A.; Castro, A.S.; da Silva, R.J.; de Oliveira, F.C.; Milian, I.C.B.; et al. Azithromycin treatment is able to control the infection by two genotypes of Toxoplasma gondii in human trophoblast BeWo cells. Exp. Parasitol. 2017, 181, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Dolcini, G.; Derrien, M.; Chaouat, G.; Barré-Sinoussi, F.; Menu, E. Cell-Free HIV Type 1 Infection is Restricted in the Human Trophoblast Choriocarcinoma BeWo Cell Line, Even with Expression of CD4, CXCR4 and CCR5. AIDS Res. Hum. Retrovir. 2003, 19, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, M.S.; Rytting, E.; Mose, T.; Knudsen, L.E. Modeling placental transport: Correlation of in vitro BeWo cell permeability and ex vivo human placental perfusion. Toxicol. In Vitro 2009, 23, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Aengenheister, L.; Keevend, K.; Muoth, C.; Schönenberger, R.; Diener, L.; Wick, P.; Buerki-Thurnherr, T. An advanced human in vitro co-culture model for translocation studies across the placental barrier. Sci. Rep. 2018, 8, 5388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, M.S.; Mose, T.; Maroun, L.L.; Mathiesen, L.; Knudsen, L.E.; Rytting, E. Kinetics of silica nanoparticles in the human placenta. Nanotoxicology 2015, 9, 79–86. [Google Scholar] [CrossRef]
- Faust, J.J.; Zhang, W.; Chen, Y.; Capco, D.G. Alpha-Fe2O3 elicits diameter-dependent effects during exposure to an in vitro model of the human placenta. Cell Biol. Toxicol. 2014, 30, 31–53. [Google Scholar] [CrossRef]
- Soininen, S.K.; Repo, J.K.; Karttunen, V.; Auriola, S.; Vähäkangas, K.H.; Ruponen, M. Human placental cell and tissue uptake of doxorubicin and its liposomal formulations. Toxicol. Lett. 2015, 239, 108–114. [Google Scholar] [CrossRef]
- Sezgin-Bayindir, Z.; Elcin, A.E.; Parmaksiz, M.; Elcin, Y.M.; Yuksel, N. Investigations on clonazepam-loaded polymeric micelle-like nanoparticles for safe drug administration during pregnancy. J. Microencapsul. 2018, 35, 149–164. [Google Scholar] [CrossRef]
- Albekairi, N.A.; Al-Enazy, S.; Ali, S.; Rytting, E. Transport of digoxin-loaded polymeric nanoparticles across BeWo cells, an in vitro model of human placental trophoblast. Ther. Deliv. 2015, 6, 1325–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, H.; Kalashnikova, I.; White, M.A.; Sherman, M.; Rytting, E. Preparation, characterization, and transport of dexamethasone-loaded polymeric nanoparticles across a human placental in vitro model. Int. J. Pharm. 2013, 454, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rytting, E.; Cartwright, L.; Poulsen, M.S.; Nielsen, H.M.; Pojana, G.; Knudsen, L.E.; Saunders, M. In vitro placental model optimization for nanoparticle transport studies. Int. J. Nanomed. 2012, 7, 497–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloet, S.K.; Walczak, A.P.; Louisse, J.; van den Berg, H.H.J.; Bouwmeester, H.; Tromp, P.; Fokkink, R.G.; Rietjens, I.M.C.M. Translocation of positively and negatively charged polystyrene nanoparticles in an in vitro placental model. Toxicol. In Vitro 2015, 29, 1701–1710. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Jiang, Z.; He, H.; Li, X.; Hu, H.; Zhang, N.; Dai, Y.; Zhou, Z. Uptake and transport of pullulan acetate nanoparticles in the BeWo b30 placental barrier cell model. Int. J. Nanomed. 2018, 13, 4073–4082. [Google Scholar] [CrossRef] [Green Version]
- Juch, H.; Nikitina, L.; Reimann, S.; Gauster, M.; Dohr, G.; Obermayer-Pietsch, B.; Hoch, D.; Kornmueller, K.; Haag, R. Dendritic polyglycerol nanoparticles show charge dependent bio-distribution in early human placental explants and reduce hCG secretion. Nanotoxicology 2018, 12, 90–103. [Google Scholar] [CrossRef]
- Chou, J.Y. Effects of Retinoic Acid on Differentiation of Choriocarcinoma Cells in Vitro. J. Clin. Endocrinol. Metab. 1982, 54, 1174–1180. [Google Scholar] [CrossRef]
- Sun, T. Characterization of a Region Upstream of Exon I.1 of the Human CYP19 (Aromatase) Gene That Mediates Regulation by Retinoids in Human Choriocarcinoma Cells. Endocrinology 1998, 139, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.; Hardy, D.B.; Pereira, L.E.; Yang, K. Retinoic Acid Stimulates the Expression of 11β-Hydroxysteroid Dehydrogenase Type 2 in Human Choriocarcinoma JEG-3 Cells1. Biol. Reprod. 1999, 60, 541–545. [Google Scholar] [CrossRef] [Green Version]
- Blanchon, L. Human choriocarcinoma cell line JEG-3 produces and secretes active retinoids from retinol. Mol. Hum. Reprod. 2002, 8, 485–493. [Google Scholar] [CrossRef] [Green Version]
- McConkey, C.A.; Delorme-Axford, E.; Nickerson, C.A.; Kim, K.S.; Sadovsky, Y.; Boyle, J.P.; Coyne, C.B. A three-dimensional culture system recapitulates placental syncytiotrophoblast development and microbial resistance. Sci. Adv. 2016, 2, e1501462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huining, L.; Yi, Z.; Dihong, T.; Yifeng, P.; Man, X.; Ting, Y.; Jingting, C. Inhibition of choriocarcinoma by Fe3O4-dextran-anti-β-human chorionic gonadotropin nanoparticles containing antisense oligodeoxynucleotide of heparanase. Int. J. Nanomed. 2013, 8, 4371–4377. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Cheng, G.; Zheng, M.; Han, J.; Wang, B.; Li, M.; Chen, J.; Xiao, T.; Zhang, J.; Cai, L.; et al. Targeted delivery of doxorubicin by CSA-binding nanoparticles for choriocarcinoma treatment. Drug Deliv. 2018, 25, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simian Virus 40, Poliovirus Vaccines, and Human Cancer: Research Progress Versus Media and Public Interests. Available online: https://www.ncbi.nlm.nih.gov/pubmed/10743284 (accessed on 28 April 2020).
- Liu, X.; Mu, H.; Luo, X.; Xiao, X.; Ding, Y.; Yin, N.; Deng, Q.; Qi, H. Expression of Gadd45α in human early placenta and its role in trophoblast invasion. Placenta 2014, 35, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Guirelli, P.M.; Angeloni, M.B.; Barbosa, B.F.; Gomes, A.O.; Castro, A.S.; Franco, P.S.; Silva, R.J.; Oliveira, J.G.; Martins-Filho, O.A.; Mineo, J.R.; et al. Trophoblast-macrophage crosstalk on human extravillous under Toxoplasma gondii infection. Placenta 2015, 36, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.J.; Song, B.F.; Zhang, Y.H.; Ma, Y.Y.; Shao, Q.Q.; Liu, J.; Qu, X. Expression of RGC32 in human normal and preeclamptic placentas and its role in trophoblast cell invasion and migration. Placenta 2015, 36, 350–356. [Google Scholar] [CrossRef]
- Yang, Y.; He, G.; Xu, W.; Liu, X. ENaC mediates human extravillous trophblast cell line (HTR8/SVneo) invasion by regulating levels of matrix metalloproteinase 2 (MMP2). Placenta 2015, 36, 587–593. [Google Scholar] [CrossRef]
- Wapner, R.J. Invasive prenatal diagnostic techniques. Semin. Perinatol. 2005, 29, 401–404. [Google Scholar] [CrossRef]
- Bolnick, J.M.; Kilburn, B.A.; Bajpayee, S.; Reddy, N.; Jeelani, R.; Crone, B.; Simmerman, N.; Singh, M.; Diamond, M.P.; Armant, D.R. Trophoblast retrieval and isolation from the cervix (TRIC) for noninvasive prenatal screening at 5 to 20 weeks of gestation. Fertil. Steril. 2014, 102, 135–142. [Google Scholar] [CrossRef]
- Kliman, H.J.; Strauss, J.F.; Nestler, J.E.; Sermasi, E.; Strauss, J.F.; Sanger, J.M. Purification, characterization, and in vitro differentiation of cytotrophoblasts from human term placentae. Endocrinology 1986, 118, 1567–1582. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Schust, D.J. Isolation, purification and in vitro differentiation of cytotrophoblast cells from human term placenta. Reprod. Biol. Endocrinol. 2015, 13, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serjilus, A.; Alcendor, D.J. Unique method for human villous trophoblasts isolation from placental tissue explants. Clin. Obstet. Gynecol. Reprod. Med. 2020, 6, 319. [Google Scholar] [CrossRef] [PubMed]
- Bajoria, R.; Sooranna, S.R.; Contractor, S.F. Endocytotic uptake of small unilamellar liposomes by human trophoblast cells in culture. Hum. Reprod. 1997, 12, 1343–1348. [Google Scholar] [CrossRef]
- Valero, L.; Alhareth, K.; Romero, J.E.; Viricel, W.; Leblond, J.; Chissey, A.; Dhotel, H.; Roques, C.; Arruda, D.C.; Escriou, V.; et al. Liposomes as gene delivery vectors for human placental cells. Molecules 2018, 23, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nursalim, Y.N.S.; Blenkiron, C.; Groom, K.M.; Chamley, L.W. Growing human trophoblasts in vitro: A review of the media commonly used in trophoblast cell culture. Reproduction 2020, 160, R119–R128. [Google Scholar] [CrossRef]
- Kovo, M.; Golan, A. In Vitro Models Using the Human Placenta to Study Fetal Exposure to Drugs. Clin. Med. Reprod. Health 2008, 2, S974. [Google Scholar] [CrossRef]
- Sheridan, M.A.; Zhao, X.; Fernando, R.C.; Gardner, L.; Perez-Garcia, V.; Li, Q.; Marsh, S.G.E.; Hamilton, R.; Moffett, A.; Turco, M.Y. Characterization of primary models of human trophoblast. Development 2021, 148, dev199749. [Google Scholar] [CrossRef]
- Kallol, S.; Moser-Haessig, R.; Ontsouka, C.E.; Albrecht, C. Comparative expression patterns of selected membrane transporters in differentiated BeWo and human primary trophoblast cells. Placenta 2018, 72–73, 48–52. [Google Scholar] [CrossRef]
- Giaginis, C.; Tsantili-Kakoulidou, A.; Theocharis, S. Assessing Drug Transport Across the Human Placental Barrier: From In Vivo and In Vitro Measurements to the Ex Vivo Perfusion Method and In silico Techniques. Curr. Pharm. Biotechnol. 2012, 12, 804–813. [Google Scholar] [CrossRef]
- Drwal, E.; Rak, A.; Gregoraszczuk, E. Co-culture of JEG-3, BeWo and syncBeWo cell lines with adrenal H295R cell line: An alternative model for examining endocrine and metabolic properties of the fetoplacental unit. Cytotechnology 2018, 70, 285–297. [Google Scholar] [CrossRef]
- Müller, E.K.; Gräfe, C.; Wiekhorst, F.; Bergemann, C.; Weidner, A.; Dutz, S.; Clement, J.H. Magnetic nanoparticles interact and pass an in vitro co-culture blood-placenta barrier model. Nanomaterials 2018, 8, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muoth, C.; Wichser, A.; Monopoli, M.; Correia, M.; Ehrlich, N.; Loeschner, K.; Gallud, A.; Kucki, M.; Diener, L.; Manser, P.; et al. A 3D co-culture microtissue model of the human placenta for nanotoxicity assessment. Nanoscale 2016, 8, 17322–17332. [Google Scholar] [CrossRef] [PubMed]
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.S.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal–fetal interactions during human placentation. Nature 2018, 564, 263–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.J.B.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, S.; Meinhardt, G.; Saleh, L.; Kunihs, V.; Gamperl, M.; Kaindl, U.; Ellinger, A.; Burkard, T.R.; Fiala, C.; Pollheimer, J.; et al. Self-Renewing Trophoblast Organoids Recapitulate the Developmental Program of the Early Human Placenta. Stem Cell Rep. 2018, 11, 537–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muoth, C.; Großgarten, M.; Karst, U.; Ruiz, J.; Astruc, D.; Moya, S.; Diener, L.; Grieder, K.; Wichser, A.; Jochum, W.; et al. Impact of particle size and surface modification on gold nanoparticle penetration into human placental microtissues. Nanomedicine 2017, 12, 1119–1133. [Google Scholar] [CrossRef]
- Blundell, C.; Tess, E.R.; Schanzer, A.S.R.; Coutifaris, C.; Su, E.J.; Parry, S.; Huh, D. A microphysiological model of the human placental barrier. Lab Chip 2016, 16, 3065–3073. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, F.; Wang, H.; Wang, L.; Yuan, J.; Qin, J. Placental Barrier-on-a-Chip: Modeling Placental Inflammatory Responses to Bacterial Infection. ACS Biomater. Sci. Eng. 2018, 4, 3356–3363. [Google Scholar] [CrossRef]
- Blundell, C.; Yi, Y.-S.; Ma, L.; Tess, E.R.; Farrell, M.J.; Georgescu, A.; Aleksunes, L.M.; Huh, D. Placental Drug Transport-on-a-Chip: A Microengineered In Vitro Model of Transporter-Mediated Drug Efflux in the Human Placental Barrier. Adv. Healthc. Mater. 2018, 7, 1700786. [Google Scholar] [CrossRef]
- Pemathilaka, R.L.; Caplin, J.D.; Aykar, S.S.; Montazami, R.; Hashemi, N.N. Placenta-on-a-Chip: In Vitro Study of Caffeine Transport across Placental Barrier Using Liquid Chromatography Mass Spectrometry. Glob. Chall. 2019, 3, 1800112. [Google Scholar] [CrossRef] [Green Version]
- Yin, F.; Zhu, Y.; Zhang, M.; Yu, H.; Chen, W.; Qin, J. A 3D human placenta-on-a-chip model to probe nanoparticle exposure at the placental barrier. Toxicol. In Vitro 2019, 54, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. 2017, 22, 456–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, P.; Malek, A.; Manser, P.; Meili, D.; Maeder-Althaus, X.; Diener, L.; Diener, P.A.; Zisch, A.; Krug, H.F.; Von Mandach, U. Barrier capacity of human placenta for nanosized materials. Environ. Health Perspect. 2010, 118, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Aengenheister, L.; Favaro, R.R.; Morales-Prieto, D.M.; Furer, L.A.; Gruber, M.; Wadsack, C.; Markert, U.R.; Buerki-Thurnherr, T. Research on nanoparticles in human perfused placenta: State of the art and perspectives. Placenta 2021, 104, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Panigel, M. Placental perfusion experiments. Am. J. Obstet. Gynecol. 1962, 84, 1670–1683. [Google Scholar] [CrossRef]
- Schneider, H.; Panigel, M.; Dancis, J. Transfer across the perfused human placenta of antipyrine, sodium, and leucine. Am. J. Obstet. Gynecol. 1972, 114, 822–828. [Google Scholar] [CrossRef]
- Pehrson, C.; Mathiesen, L.; Heno, K.K.; Salanti, A.; Resende, M.; Dzikowski, R.; Damm, P.; Hansson, S.R.; King, C.L.; Schneider, H.; et al. Adhesion of Plasmodium falciparum infected erythrocytes in ex vivo perfused placental tissue: A novel model of placental malaria. Malar. J. 2016, 15, 292. [Google Scholar] [CrossRef] [Green Version]
- Berveiller, P.; Gil, S.; Vialard, F. Placental perfusion: Interest and limits. J. Matern. Neonatal Med. 2017, 30, 1347–1348. [Google Scholar] [CrossRef]
- Myllynen, P.K.; Loughran, M.J.; Howard, C.V.; Sormunen, R.; Walsh, A.A.; Vähäkangas, K.H. Kinetics of gold nanoparticles in the human placenta. Reprod. Toxicol. 2008, 26, 130–137. [Google Scholar] [CrossRef]
- Menjoge, A.R.; Rinderknecht, A.L.; Navath, R.S.; Faridnia, M.; Kim, C.J.; Romero, R.; Miller, R.K.; Kannan, R.M. Transfer of PAMAM dendrimers across human placenta: Prospects of its use as drug carrier during pregnancy. J. Control. Release 2011, 150, 326–338. [Google Scholar] [CrossRef] [Green Version]
- Bajoria, R.; Contractor, S.F. Effect of the size of liposomes on the transfer and uptake of carboxyfluorescein by the perfused human term placenta. J. Pharm. Pharmacol. 1997, 49, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Bajoria, R.; Sooranna, S.; Chatterjee, R. Effect of lipid composition of cationic SUV liposomes on materno-fetal transfer of warfarin across the perfused human term placenta. Placenta 2013, 34, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Sooranna, S.R.; Oteng-Ntim, E.; Meah, R.; Ryder, T.A.; Bajoria, R. Characterization of human placental explants: Morphological, biochemical and physiological studies using first and third trimester placenta. Hum. Reprod. 1999, 14, 536–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.K.; Genbacev, O.; Turner, M.A.; Aplin, J.D.; Caniggia, I.; Huppertz, B. Human placental explants in culture: Approaches and assessments. Placenta 2005, 26, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Balan, A.; Szaingurten-Solodkin, I.; Swissa, S.S.; Feinshtein, V.; Huleihel, M.; Holcberg, G.; Dukler, D.; Beharier, O. The effects of pravastatin on the normal human placenta: Lessons from ex-vivo models. PLoS ONE 2017, 12, e0172174. [Google Scholar] [CrossRef]
- Valero, L.; Alhareth, K.; Gil, S.; Simasotchi, C.; Roques, C.; Scherman, D.; Mignet, N.; Fournier, T.; Andrieux, K. Assessment of dually labelled PEGylated liposomes transplacental passage and placental penetration using a combination of two ex-vivo human models: The dually perfused placenta and the suspended villous explants. Int. J. Pharm. 2017, 532, 729–737. [Google Scholar] [CrossRef]
- Alhareth, K.; Valero, L.; Mohamed, K.E.; Fliedel, L.; Roques, C.; Gil, S.; Mignet, N.; Fournier, T.; Andrieux, K. Qualitative and quantitative analysis of the uptake of lipoplexes by villous placenta explants. Int. J. Pharm. 2019, 567, 118479. [Google Scholar] [CrossRef]
- Kaitu’u-Lino, T.J.; Pattison, S.; Ye, L.; Tuohey, L.; Sluka, P.; MacDiarmid, J.; Brahmbhatt, H.; Johns, T.; Horne, A.W.; Brown, J.; et al. Targeted nanoparticle delivery of doxorubicin into placental tissues to treat ectopic pregnancies. Endocrinology 2013, 154, 911–919. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Rajakumar, A. Preeclampsia and soluble fms-like tyrosine kinase 1. J. Clin. Endocrinol. Metab. 2009, 94, 2252–2254. [Google Scholar] [CrossRef] [Green Version]
- Campagnolo, L.; Massimiani, M.; Vecchione, L.; Piccirilli, D.; Toschi, N.; Magrini, A.; Bonanno, E.; Scimeca, M.; Castagnozzi, L.; Buonanno, G.; et al. Silver nanoparticles inhaled during pregnancy reach and affect the placenta and the foetus. Nanotoxicology 2017, 11, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Naserzadeh, P.; Ghanbary, F.; Ashtari, P.; Seydi, E.; Ashtari, K.; Akbari, M. Biocompatibility assessment of titanium dioxide nanoparticles in mice fetoplacental unit. J. Biomed. Mater. Res. Part A 2018, 106, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Lopalco, A.; Ali, H.; Denora, N.; Rytting, E. Oxcarbazepine-loaded polymeric nanoparticles: Development and permeability studies across in vitro models of the blood–brain barrier and human placental trophoblast. Int. J. Nanomed. 2015, 10, 1985–1996. [Google Scholar] [CrossRef] [Green Version]







| Characteristics | Rodents | Human Primates | |
|---|---|---|---|
| Anatomy [60] | Morphology | Discoid: a single placenta is formed in a discoid shape (Villi are connected in a disk) | |
| Structure [61] | Labyrinthine placenta | Villous placenta | |
| Histology | Hemochorial placenta: Placental tissues are bathed in maternal blood (opposition to other placenta where fetal tissues are separated from maternal blood by 2 or 3 layers of cells). | ||
| Hemotrichorial: one layer of cytotrophoblast based on two layers of basal syncytiotrophoblast | Hemodichorial: one layer of syncytiotrophoblast upon one layer of cytotrophoblast on basal layer | ||
| Physiological functions—major differences [62] | Progesterone production provided by corpus luteum indispensable during the whole pregnancy. Chorionic gonadotropin hormone presence has not been demonstrated | Progesterone production provided by corpus luteum and after placental production takes over gradually. Chorionic gonadotropin hormone presence | |
| Immortalized Human Trophoblasts | Trophoblasts Derived from Human Choriocarcinoma | Hybrid Cell Line |
|---|---|---|
| HTR-8/Svneo—1st T (SV 40) | BeWo | ACH-3P |
| Swan 71—1st T (SV 40) | JEG-3 | |
| JAR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fliedel, L.; Alhareth, K.; Mignet, N.; Fournier, T.; Andrieux, K. Placental Models for Evaluation of Nanocarriers as Drug Delivery Systems for Pregnancy Associated Disorders. Biomedicines 2022, 10, 936. https://doi.org/10.3390/biomedicines10050936
Fliedel L, Alhareth K, Mignet N, Fournier T, Andrieux K. Placental Models for Evaluation of Nanocarriers as Drug Delivery Systems for Pregnancy Associated Disorders. Biomedicines. 2022; 10(5):936. https://doi.org/10.3390/biomedicines10050936
Chicago/Turabian StyleFliedel, Louise, Khair Alhareth, Nathalie Mignet, Thierry Fournier, and Karine Andrieux. 2022. "Placental Models for Evaluation of Nanocarriers as Drug Delivery Systems for Pregnancy Associated Disorders" Biomedicines 10, no. 5: 936. https://doi.org/10.3390/biomedicines10050936