
The Role of Hub and Spoke Regions in Theory of Mind in Early Alzheimer’s Disease and Frontotemporal Dementia

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
- (i)
- MMSE score ≥26 and CDR score ≤1 at the time of enrollment;
- (ii)
- Symptoms duration less than two years at enrollment;
- (iii)
- Confirmation of clinical diagnosis after at least two years of follow-up;
- (iv)
- No current or past diagnosis of a comorbid psychiatric or neurological condition, or other major medical conditions;
- (v)
- No regular use of psychoactive drugs;
- (vi)
- A brain 18F-FDG-PET scan performed no later than three months after neuropsychological testing.
2.2. Theory of Mind Assessment
2.3. 18F-FDG-PET Acquisition
2.4. Reading the Mind in the Eyes Task Regions of Interest
2.5. Statistical Analysis
3. Results
3.1. Relative Hypometabolism in Behavioral Variant of Frontotemporal Dementia Patients vs. Controls
3.2. Relative Hypometabolism in Mild Cognitive Impairment due to Alzheimer’s Disease Patients vs. Controls
3.3. Hub and Spoke Definition
3.4. Correlation between Regional Metabolism of the Reading the Mind in the Eyes Task Regions of Interest and Reading the Mind in the Eyes Task Performance: Behavioral Variant of Frontotemporal Dementia
3.5. Correlation between Regional Metabolism in the Reading the Mind in the Eyes Task Regions of Interest and Reading the Mind in the Eyes Task Performance: Mild Cognitive Impairment Due to Alzheimer’s Disease
3.6. Hub–Spoke Interactions and Reading the Mind in the Eyes Task Performance: Behavioral Variant of Frontotemporal Dementia
3.7. Hub–Spoke Interactions and Reading the Mind in the Eyes Task Performance: Mild Cognitive Impairment due to Alzheimer’s Disease
4. Discussion
5. Conclusions
6. Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baron-Cohen, S.; Campbell, R.; Karmiloff-Smith, A.; Grant, J.; Walker, J. Are children with autism blind to the mentalistic significance of the eyes? Br. J. Dev. Psychol. 1995, 13, 379–398. [Google Scholar] [CrossRef]
- Orso, B.; Arnaldi, D.; Famà, F.; Girtler, N.; Brugnolo, A.; Doglione, E.; Filippi, L.; Massa, F.; Peira, E.; Bauckneht, M.; et al. Anatomical and neurochemical bases of theory of mind in de novo Parkinson’s Disease. Cortex 2020, 130, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Pardini, M.; Gialloreti, L.E.; Mascolo, M.; Benassi, F.; Abate, L.; Guida, S.; Viani, E.; Monte, O.D.; Schintu, S.; Krueger, F.; et al. Isolated theory of mind deficits and risk for frontotemporal dementia: A longitudinal pilot study. J. Neurol. Neurosurg. Psychiatry 2013, 84, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Schurz, M.; Radua, J.; Aichhorn, M.; Richlan, F.; Perner, J. Fractionating theory of mind: A meta-analysis of functional brain imaging studies. Neurosci. Biobehav. Rev. 2014, 42, 9–34. [Google Scholar] [CrossRef] [Green Version]
- Schurz, M.; Perner, J. An evaluation of neurocognitive models of theory of mind. Front. Psychol. 2015, 6, 1610. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Zhao, Y.; Zhang, T.; Zhao, D.; Zhao, F.; Lu, E. A Brain-Inspired Model of Theory of Mind. Front. Neurorobotics 2020, 14, 60. [Google Scholar] [CrossRef]
- Ricci, M.; Cimini, A.; Camedda, R.; Chiaravalloti, A.; Schillaci, O. Tau Biomarkers in Dementia: Positron Emission Tomography Radiopharmaceuticals in Tauopathy Assessment and Future Perspective. Int. J. Mol. Sci. 2021, 22, 13002. [Google Scholar] [CrossRef]
- Chiaravalloti, A.; Barbagallo, G.; Ricci, M.; Martorana, A.; Ursini, F.; Sannino, P.; Karalis, G.; Schillaci, O. Brain metabolic correlates of CSF Tau protein in a large cohort of Alzheimer’s disease patients: A CSF and FDG PET study. Brain Res. 2018, 1678, 116–122. [Google Scholar] [CrossRef]
- Balogh, L.; Tanaka, M.; Török, N.; Vécsei, L.; Taguchi, S. Crosstalk between Existential Phenomenological Psychotherapy and Neurological Sciences in Mood and Anxiety Disorders. Biomedicines 2021, 9, 340. [Google Scholar] [CrossRef]
- Buckner, R.L.; Andrews-Hanna, E.J.R.; Schactera, D.L. The Brain’s Default Network: Anatomy, Function, and Relevance to Disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Kyeong, S.; Kim, J.; Kim, J.; Kim, E.J.; Kim, H.E.; Kim, J.-J. Differences in the modulation of functional connectivity by self-talk tasks between people with low and high life satisfaction. NeuroImage 2020, 217, 116929. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Peng, Y.; Chong, T.; Zhang, Y.; Von Deneen, K.M.; Huang, L.; Cai, Y.P.S. Differentiated effective connectivity patterns of the executive control network in progressive MCI: A potential biomarker for predicting AD. Curr. Alzheimer Res. 2017, 14, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Lu, H.; Metmer, H.; Li, W.X.; Lu, J. Evaluating functional connectivity of executive control network and frontoparietal network in Alzheimer’s disease. Brain Res. 2018, 1678, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Cieri, F.; Esposito, R.; Cera, N.; Pieramico, V.; Tartaro, A.; Di Giannantonio, M. Late-Life Depression: Modifications of Brain Resting State Activity. J. Geriatr. Psychiatry Neurol. 2017, 30, 140–150. [Google Scholar] [CrossRef]
- Seeley, W.W. The Salience Network: A Neural System for Perceiving and Responding to Homeostatic Demands. J. Neurosci. 2019, 39, 9878–9882. [Google Scholar] [CrossRef]
- Dai, L.; Zhou, H.; Xu, X.; Zuo, Z. Brain structural and functional changes in patients with major depressive disorder: A literature review. PeerJ 2019, 7, e8170. [Google Scholar] [CrossRef]
- Battaglia, S.; Harrison, B.J.; Fullana, M.A. Does the human ventromedial prefrontal cortex support fear learning, fear extinction or both? A commentary on subregional contributions. Mol. Psychiatry 2021, 1–3. [Google Scholar] [CrossRef]
- Van Den Heuvel, M.P.; Stam, C.J.; Kahn, R.S.; Pol, H.E.H. Efficiency of functional brain networks and intellectual performance. J. Neurosci. 2009, 29, 7619–7624. [Google Scholar] [CrossRef] [Green Version]
- Hwang, K.; Hallquist, M.N.; Luna, B. The development of hub architecture in the human functional brain network. Cereb. Cortex 2013, 23, 2380–2393. [Google Scholar] [CrossRef]
- Albert, R.; Jeong, H.; Barabási, A.L. Error and attack tolerance of complex networks. Nature 2000, 406, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Aben, H.P.; Biessels, G.J.; Weaver, N.A.; Spikman, J.M.; Visser-Meily, J.M.; de Kort, P.L.; Reijmer, Y.D.; PROCRAS Study Group. Extent to Which Network Hubs Are Affected by Ischemic Stroke Predicts Cognitive Recovery. Stroke 2019, 50, 2768–2774. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.S.; DeKosky, S.T.; Dickson, D.; Dubois, B.; Feldman, H.H.; Fox, N.C.; Gamst, A.; Holtzman, D.M.; Jagust, W.J.; Petersen, R.C.; et al. The Diagnosis of Mild Cognitive Impairment due to Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Focus 2013, 11, 96–106. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Rascovsky, K.; Hodges, J.R.; Kipps, C.; Johnson, J.K.; Seeley, W.W.; Mendez, M.F.; Knopman, D.S.; Kertesz, A.; Mesulam, M.M.; Salmon, D.P.; et al. Diagnostic Criteria for the Behavioral Variant of Frontotemporal Dementia (bvFTD): Current Limitations and Future Directions. Alzheimer Dis. Assoc. Disord. 2007, 21, S14–S18. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Wheelwright, S.; Hill, J.; Raste, Y.; Plumb, I. The “Reading the Mind in the Eyes” Test revised version: A study with normal adults, and adults with Asperger syndrome or high-functioning autism. J. Child Psychol. Psychiatry 2001, 42, 241–251. [Google Scholar] [CrossRef]
- Harkness, K.L.; Sabbagh, M.A.; Jacobson, J.A.; Chowdrey, N.K.; Chen, T. Enhanced accuracy of mental state decoding in dysphoric college students. Cogn. Emot. 2005, 19, 999–1025. [Google Scholar] [CrossRef]
- Pardini, M.; Nichelli, P.F. Age-related decline in mentalizing skills across adult life span. Exp. Aging Res. 2009, 35, 98–106. [Google Scholar] [CrossRef]
- Varrone, A.; Asenbaum, S.; Borght, T.V.; Booij, J.; Nobili, F.; Någren, K.; Darcourt, J.; Kapucu, Ö.L.; Tatsch, K.; Bartenstein, P.; et al. EANM procedure guidelines for PET brain imaging using [18F] FDG, version 2. Eur. J. Nucl. Med. Mol. Imaging 2009, 36, 2103–2110. [Google Scholar] [CrossRef]
- Della Rosa, P.A.; the EADC-PET Consortium; Cerami, C.; Gallivanone, F.; Prestia, A.; Caroli, A.; Castiglioni, I.; Gilardi, M.C.; Frisoni, G.; Friston, K.; et al. A Standardized [18F]-FDG-PET Template for Spatial Normalization in Statistical Parametric Mapping of Dementia. Neuroinformatics 2014, 12, 575–593. [Google Scholar] [CrossRef]
- Molenberghs, P.; Johnson, H.; Henry, J.D.; Mattingley, J.B. Understanding the minds of others: A neuroimaging meta-analysis. Neurosci. Biobehav. Rev. 2016, 65, 276–291. [Google Scholar] [CrossRef] [Green Version]
- Abu-Akel, A. A neurobiological mapping of theory of mind. Brain Res. Rev. 2003, 43, 29–40. [Google Scholar] [CrossRef]
- Cotter, J.; Granger, K.; Backx, R.; Hobbs, M.; Looi, C.Y.; Barnett, J.H. Social cognitive dysfunction as a clinical marker: A systematic review of meta-analyses across 30 clinical conditions. Neurosci. Biobehav. Rev. 2018, 84, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Koff, E.; Brownell, H.; Winner, E.; Albert, M.; Zaitchik, D. Inference of mental states in patients with Alzheimer’s disease. Cogn. Neuropsychiatry 2004, 9, 301–313. [Google Scholar] [CrossRef] [PubMed]
- De Lucena, A.T.; Bhalla, R.K.; Dos Santos, T.T.B.A.; Dourado, M.C.N. The relationship between theory of mind and cognition in Alzheimer’s disease: A systematic review. J. Clin. Exp. Neuropsychol. 2020, 42, 223–239. [Google Scholar] [CrossRef]
- Orso, B.; Mattei, C.; Arnaldi, D.; Massa, F.; Serafini, G.; Plantone, D.; Doglione, E.; Grafman, J.; Nobili, F.; Pardini, M. Clinical and MRI Predictors of Conversion From Mild Behavioural Impairment to Dementia. Am. J. Geriatr. Psychiatry 2020, 28, 755–763. [Google Scholar] [CrossRef]
- Dal Monte, O.; Schintu, S.; Pardini, M.; Berti, A.; Wassermann, E.M.; Grafman, J.; Krueger, F. The left inferior frontal gyrus is crucial for reading the mind in the eyes: Brain lesion evidence. Cortex 2014, 58, 9–17. [Google Scholar] [CrossRef]
- Rowe, A.D.; Bullock, P.R.; Polkey, C.E.; Morris, R.G. ‘Theory of mind’ impairments and their relationship to executive functioning following frontal lobe excisions. Brain 2001, 124, 600–616. [Google Scholar] [CrossRef]
- Le Bouc, R.; Lenfant, P.; Delbeuck, X.; Ravasi, L.; Lebert, F.; Semah, F.; Pasquier, F. My belief or yours? Differential theory of mind deficits in frontotemporal dementia and Alzheimer’s disease. Brain 2012, 135, 3026–3038. [Google Scholar] [CrossRef] [Green Version]
- Ellena, G.; Battaglia, S.; Làdavas, E. The spatial effect of fearful faces in the autonomic response. Exp. Brain Res. 2020, 238, 2009–2018. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Battaglia, S.; Avenanti, A. Early Right Motor Cortex Response to Happy and Fearful Facial Expressions: A TMS Motor-Evoked Potential Study. Brain Sci. 2021, 11, 1203. [Google Scholar] [CrossRef]
- Kim, J.; Kim, Y.-K. Crosstalk between Depression and Dementia with Resting-State fMRI Studies and Its Relationship with Cognitive Functioning. Biomedicines 2021, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Vécsei, L. Editorial of Special Issue “Crosstalk between Depression, Anxiety, and Dementia: Comorbidity in Behavioral Neurology and Neuropsychiatry”. Biomedicines 2021, 9, 517. [Google Scholar] [CrossRef] [PubMed]
- Bryll, A.; Krzyściak, W.; Karcz, P.; Pilecki, M.; Śmierciak, N.; Szwajca, M.; Skalniak, A.; Popiela, T. Determinants of Schizophrenia Endophenotypes Based on Neuroimaging and Biochemical Parameters. Biomedicines 2021, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Li, S.; Ye, L.; Guan, J. Gene Module Analysis Reveals Cell-Type Specificity and Potential Target Genes in Autism’s Pathogenesis. Biomedicines 2021, 9, 410. [Google Scholar] [CrossRef]
- Chiarelli, A.M.; Perpetuini, D.; Croce, P.; Filippini, C.; Cardone, D.; Rotunno, L.; Anzoletti, N.; Zito, M.; Zappasodi, F.; Merla, A. Evidence of Neurovascular Un-Coupling in Mild Alzheimer’s Disease through Multimodal EEG-fNIRS and Multivariate Analysis of Resting-State Data. Biomedicines 2021, 9, 337. [Google Scholar] [CrossRef]
- Hirao, K.; Miyata, J.; Fujiwara, H.; Yamada, M.; Namiki, C.; Shimizu, M.; Sawamoto, N.; Fukuyama, H.; Hayashi, T.; Murai, T. Theory of mind and frontal lobe pathology in schizophrenia: A voxel-based morphometry study. Schizophr. Res. 2008, 105, 165–174. [Google Scholar] [CrossRef]
- Yildirim, E.; Buyukiscan, E.S.; Demirtas-Tatlidede, A.; Bilgiç, B.; Gurvit, H. An investigation of affective theory of mind ability and its relation to neuropsychological functions in Alzheimer’s disease. J. Neuropsychol. 2020, 14, 399–415. [Google Scholar] [CrossRef]
- Overgaauw, S.; Van Duijvenvoorde, A.C.K.; Moor, B.G.; Crone, E. A longitudinal analysis of neural regions involved in reading the mind in the eyes. Soc. Cogn. Affect. Neurosci. 2014, 10, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef]
- Abu-Akel, A.; Shamay-Tsoory, S. Neuroanatomical and neurochemical bases of theory of mind. Neuropsychologia 2011, 49, 2971–2984. [Google Scholar] [CrossRef]
- Irish, M.; Hodges, J.R.; Piguet, O. Right anterior temporal lobe dysfunction underlies theory of mind impairments in semantic dementia. Brain 2014, 137, 1241–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodden, M.E.; Dodel, R.; Kalbe, E. Theory of mind in Parkinson’s disease and related basal ganglia disorders: A systematic review. Mov. Disord. 2009, 25, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Brunet, E.; Sarfati, Y.; Hardy-Baylé, M.-C.; Decety, J. Abnormalities of brain function during a nonverbal theory of mind task in schizophrenia. Neuropsychologia 2003, 41, 1574–1582. [Google Scholar] [CrossRef]
- Isernia, S.; Cabinio, M.; Pirastru, A.; Mendozzi, L.; Di Dio, C.; Marchetti, A.; Massaro, D.; Baglio, F. Theory of mind network in multiple Sclerosis: A double disconnection mechanism. Soc. Neurosci. 2020, 15, 544–557. [Google Scholar] [CrossRef] [PubMed]
- Honey, C.J.; Sporns, O. Dynamical consequences of lesions in cortical networks. Hum. Brain Mapp. 2008, 29, 802–809. [Google Scholar] [CrossRef]
- Foulon, C.; Cerliani, L.; Kinkingnéhun, S.; Levy, R.; Rosso, C.; Urbanski, M.; Volle, E.; De Schotten, M.T. Advanced lesion symptom mapping analyses and implementation as BCBtoolkit. GigaScience 2018, 7, giy004. [Google Scholar] [CrossRef] [Green Version]
- Pardini, M.; Yaldizli, Ö.; Sethi, V.; Muhlert, N.; Liu, Z.; Samson, R.S.; Altmann, D.R.; Ron, M.A.; Wheeler-Kingshott, C.A.; Miller, D.H.; et al. Motor network efficiency and disability in multiple sclerosis. Neurology 2015, 85, 1115–1122. [Google Scholar] [CrossRef] [Green Version]



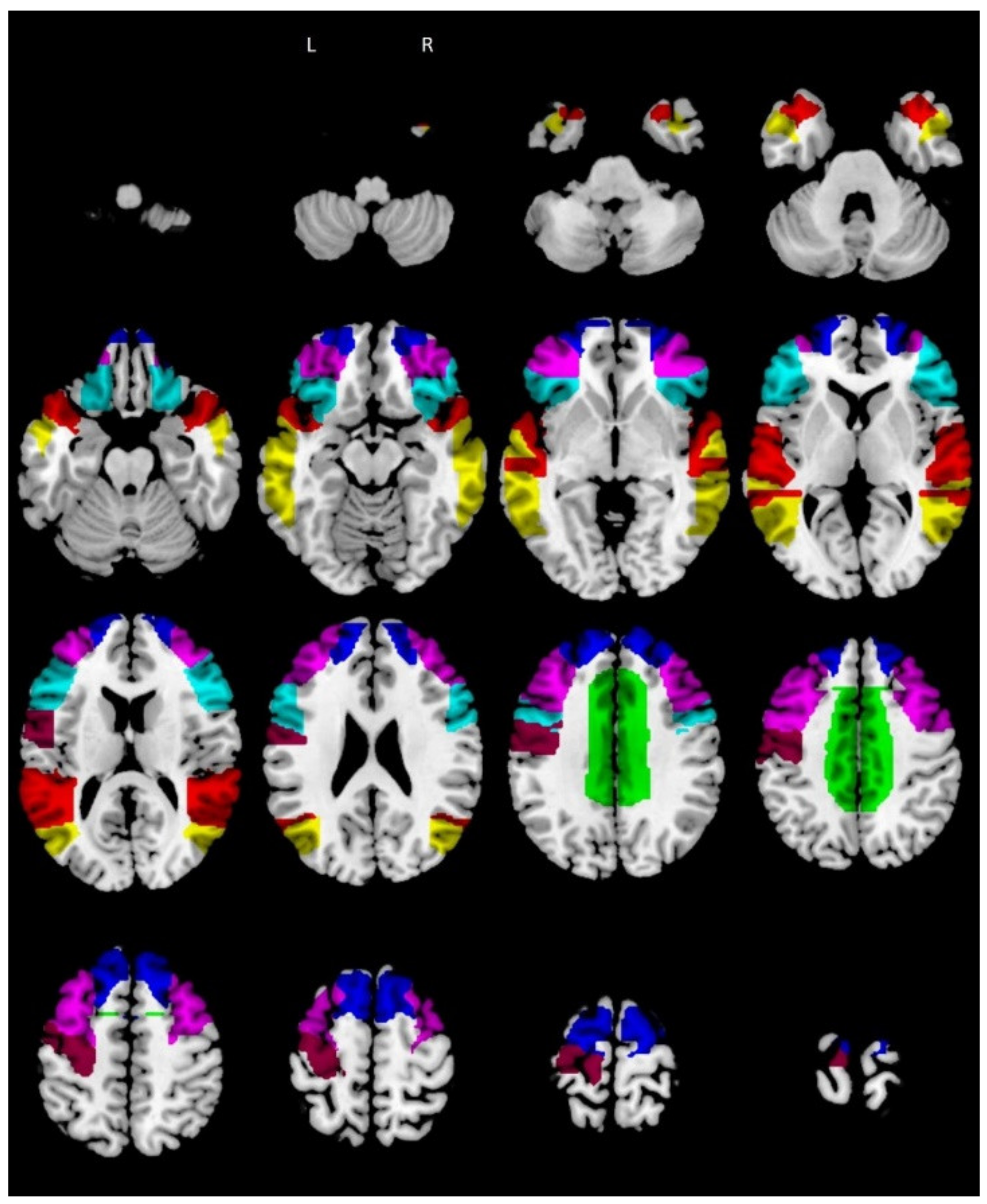{kind=link}
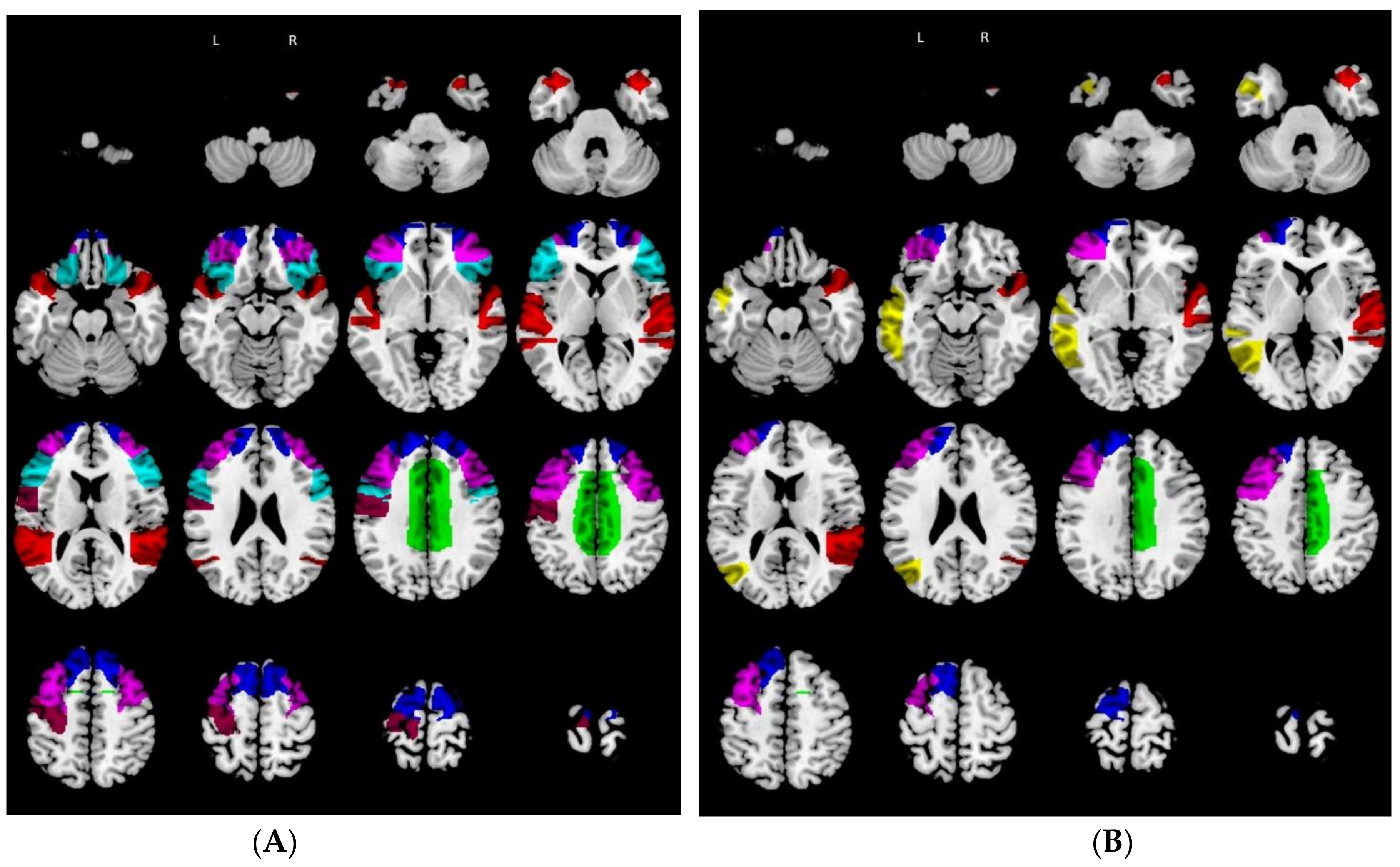{kind=link}
| bvFTD (n = 24) | MCI–AD (n = 25) | HC (n = 40) | p-Value | |
|---|---|---|---|---|
| Age (yr) | 75.58 ± 7.66 | 80.72 ± 5.61 | 78.5 ± 9.4 | p = 0.09 |
| Education (yr) | 9.86 ± 4.66 | 10.32 ± 4.07 | 9.9 ± 3.2 | p = 0.714 |
| Gender (M:F) | 6:18 | 9:16 | 15:25 | p = 0.568 |
| MMSE score | 26.41 ± 2.84 | 26.13 ± 3.01 | 28.0 ± 3.2 | p = 0.031 |
| RMET total score | 16.25 ± 4.89 | 17.6 ± 6.8 | 28.9 ± 2.1 | p = 0.000 |
| GDS-15 score | 3.78 ± 2.69 | 3.86 ± 2.79 | 2.0 ± 3.0 | p = 0.014 |
| bvFTD | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Connection | Connections among Hypometabolic Areas in bvFTD | |||||||||||||
| LMTG (*) | - | - | * | * | - | * | - | - | - | - | - | - | 0 | |
| LCG (X) | - | - | - | - | X | - | - | - | - | - | - | - | 1 | |
| LIFG (X) | - | - | X | X | X | - | - | - | - | - | - | - | 3 | |
| LMFG (X) | * | - | X | X | X | - | - | - | - | X | X | - | 5 | |
| LPG (X) | * | - | X | X | X | - | - | - | - | - | X | - | 4 | |
| LSFG (X) | - | X | X | X | X | - | - | - | - | X | X | - | 6 | |
| LSTG (X) | * | - | - | - | - | - | - | - | - | - | - | - | 0 | |
| RMTG (*) | - | - | - | - | - | - | - | - | - | X | - | X | 0 | |
| RCG (X) | - | - | - | - | - | - | - | - | - | - | X | - | 1 | |
| RIFG (X) | - | - | - | - | - | - | - | - | - | X | X | - | 2 | |
| RMFG (X) | - | - | - | X | - | X | - | * | - | X | X | - | 4 | |
| RSFG (X) | - | - | - | X | X | X | - | - | X | X | X | - | 6 | |
| RSTG (X) | - | - | - | - | - | - | - | * | - | - | - | - | 0 | |
| LMTG (*) | LCG | LIFG | LMFG | LPG | LSFG | LSTG | RMTG (*) | RCG | RIFG | RMFG | RSFG | RSTG | ||
| MCI–AD | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Connection | Connections among Hypometabolic Areas in MCI–AD | |||||||||||||
| LMTG (X) | - | - | X | * | - | * | - | - | - | - | - | - | 1 | |
| LCG (*) | - | - | - | - | * | - | - | - | - | - | - | - | 0 | |
| LIFG (*) | - | - | * | * | * | - | - | - | - | - | - | - | 0 | |
| LMFG (X) | X | - | * | * | X | - | - | - | - | * | * | - | 2 | |
| LPG (*) | * | - | * | * | * | - | - | - | - | - | * | - | 0 | |
| LSFG (X) | - | * | * | X | * | - | - | - | - | * | * | - | 1 | |
| LSTG (*) | * | - | - | - | - | - | - | - | - | - | - | - | 0 | |
| RMTG (*) | - | - | - | - | - | - | - | * | - | * | 0 | |||
| RCG (X) | - | - | - | - | - | - | - | - | * | - | 0 | |||
| RIFG (*) | - | - | - | - | - | - | - | - | - | * | * | - | 0 | |
| RMFG (*) | - | - | - | * | * | - | * | - | * | * | - | 0 | ||
| RSFG (*) | - | - | - | * | * | * | - | - | * | * | * | - | 0 | |
| RSTG (X) | - | - | - | - | - | - | - | * | - | - | - | - | 0 | |
| LMTG | LCG (*) | LIFG (*) | LMFG (X) | LPG (*) | LSFG (X) | LSTG (*) | RMTG (*) | RCG (X) | RIFG (*) | RMFG (*) | RSFG (*) | RSTG (X) | ||
| bvFTD | MCI–AD | |||
|---|---|---|---|---|
| r Values | p Values (FDR Corrected) | r Values | p Values (FDR Corrected) | |
| LMTG | r = 0.278 | p = 0.280 | r = 0.43 | p = 0.033 |
| LCG | r = 0.48 | p = 0.033 | r = −0.358 | p = 0.150 |
| LIFG | r = 0.53 | p = 0.010 | r = −0.027 | p = 0.906 |
| LMFG | r = 0.65 | p = 0.004 | r = 0.69 | p = 0.01 |
| LPG | r = 0.56 | p = 0.005 | r = 0.010 | p = 0.966 |
| LSFG | r = 0.72 | p = 0.004 | r = 0.50 | p = 0.039 |
| LSTG | r = −0.205 | p = 0.431 | r = −0.190 | p = 0.398 |
| RMTG | r = −0.101 | p = 0.699 | r = 0.078 | p = 0.729 |
| RCG | r = 0.45 | p = 0.032 | r = −0.478 | p = 0.120 |
| RIFG | r = 0.52 | p = 0.013 | r = 0.064 | p = 0.776 |
| RMFG | r = 0.57 | p = 0.005 | r = 0.196 | p = 0.383 |
| RSFG | r = 0.70 | p = 0.004 | r = 0.086 | p = 0.703 |
| RSTG | r = 0.226 | p = 0.382 | r = −0.241 | p = 0.281 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orso, B.; Lorenzini, L.; Arnaldi, D.; Girtler, N.; Brugnolo, A.; Doglione, E.; Mattioli, P.; Biassoni, E.; Massa, F.; Peira, E.; et al. The Role of Hub and Spoke Regions in Theory of Mind in Early Alzheimer’s Disease and Frontotemporal Dementia. Biomedicines 2022, 10, 544. https://doi.org/10.3390/biomedicines10030544
Orso B, Lorenzini L, Arnaldi D, Girtler N, Brugnolo A, Doglione E, Mattioli P, Biassoni E, Massa F, Peira E, et al. The Role of Hub and Spoke Regions in Theory of Mind in Early Alzheimer’s Disease and Frontotemporal Dementia. Biomedicines. 2022; 10(3):544. https://doi.org/10.3390/biomedicines10030544
Chicago/Turabian StyleOrso, Beatrice, Luigi Lorenzini, Dario Arnaldi, Nicola Girtler, Andrea Brugnolo, Elisa Doglione, Pietro Mattioli, Erica Biassoni, Federico Massa, Enrico Peira, and et al. 2022. "The Role of Hub and Spoke Regions in Theory of Mind in Early Alzheimer’s Disease and Frontotemporal Dementia" Biomedicines 10, no. 3: 544. https://doi.org/10.3390/biomedicines10030544