
Laminin Triggers Neutrophil Extracellular Traps (NETs) and Modulates NET Release Induced by Leishmania amazonensis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Laminins
2.2. Neutrophil Isolation
2.3. NET Quantification
2.4. NET Inhibition Assays
2.5. Immunofluorescence
2.6. Integrin Receptor Expression and Inhibition Assays
2.7. Parasite Culture
2.8. Production of NET-Rich Supernatants
2.9. Modulation of NET Release
2.10. Parasite Survival Assay
2.11. Statistical Analysis
3. Results
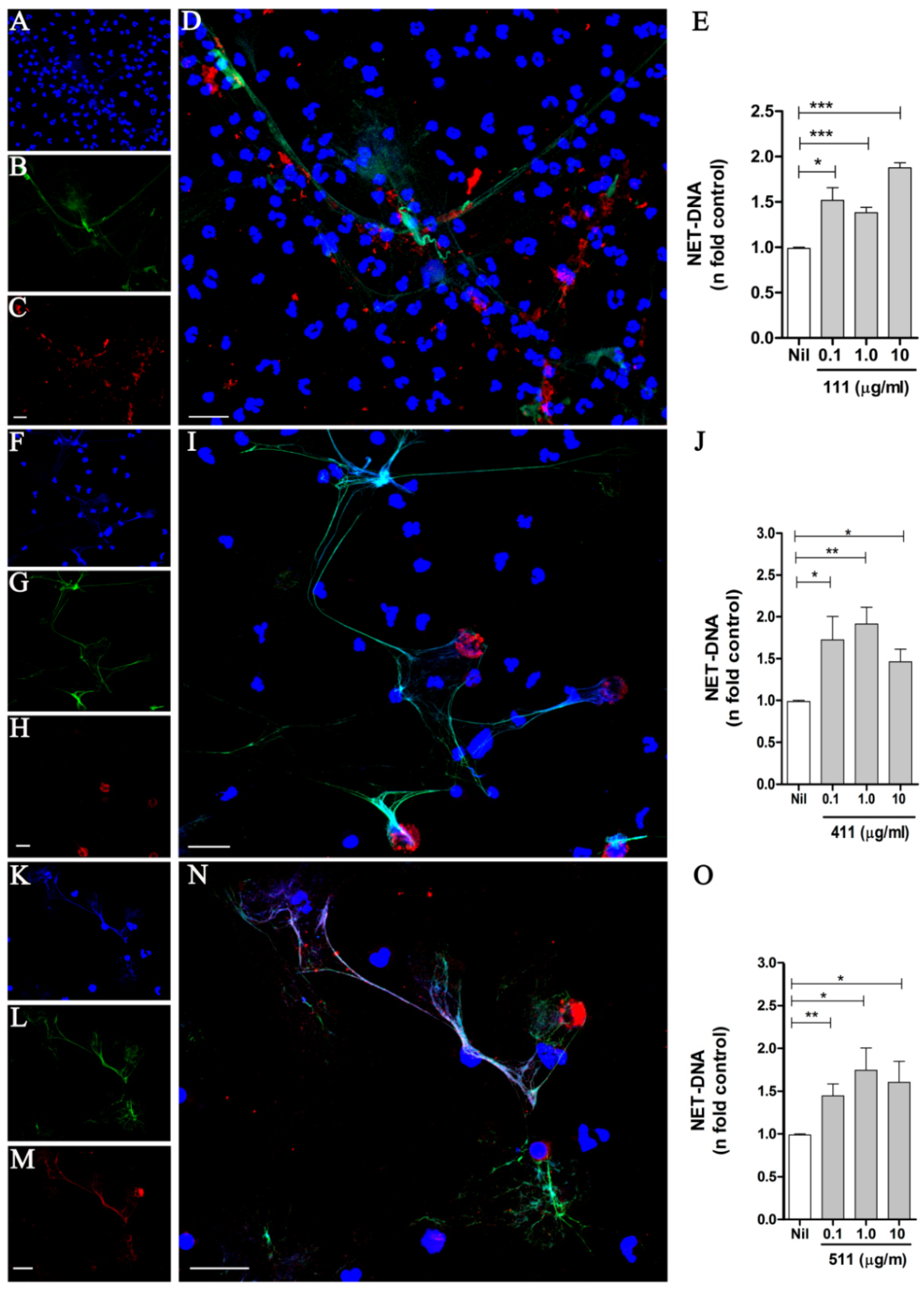
3.1. Soluble Laminins Trigger NET Release
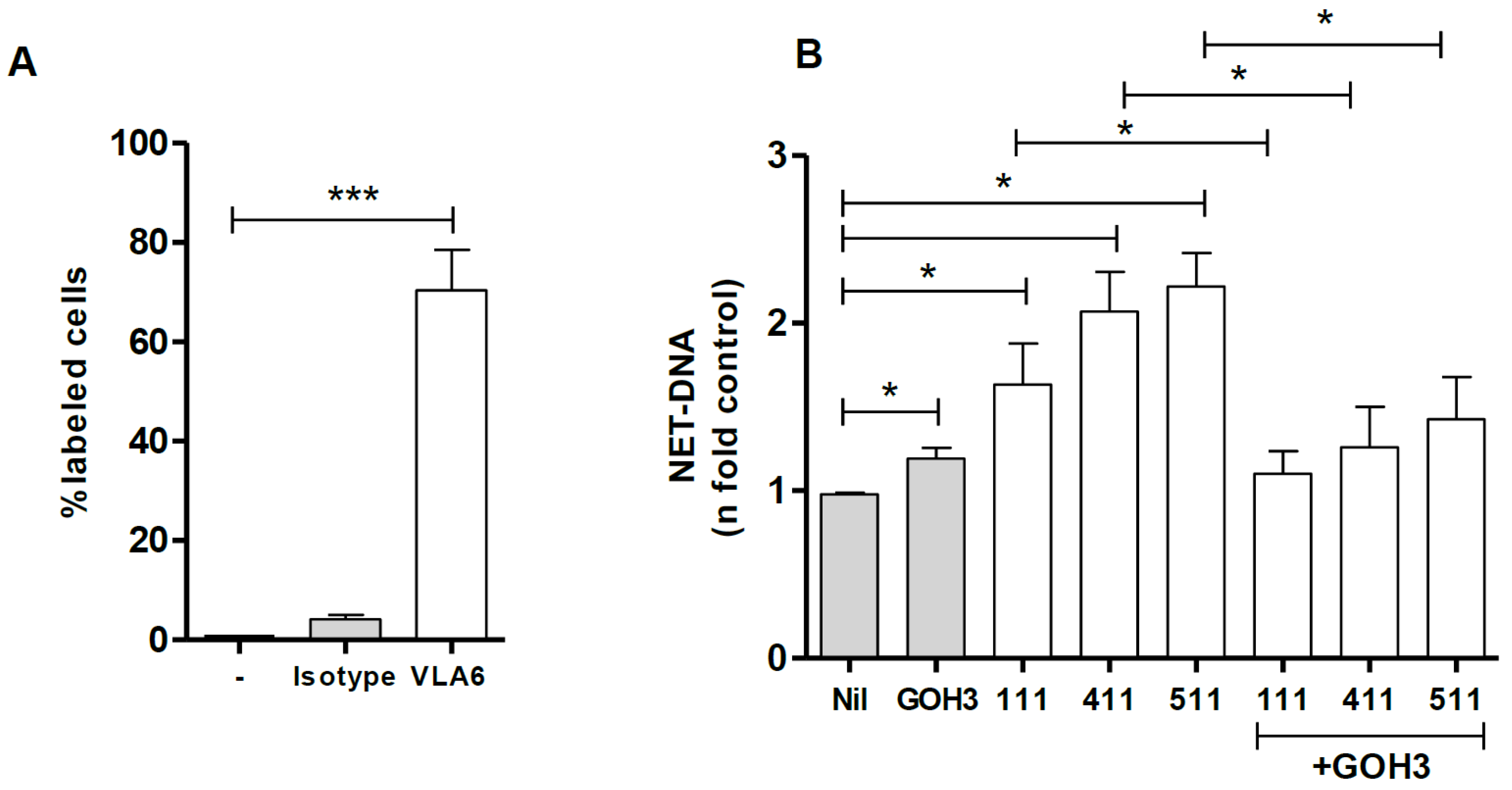
3.2. Integrin α6 Is Involved in NET Release by Laminins
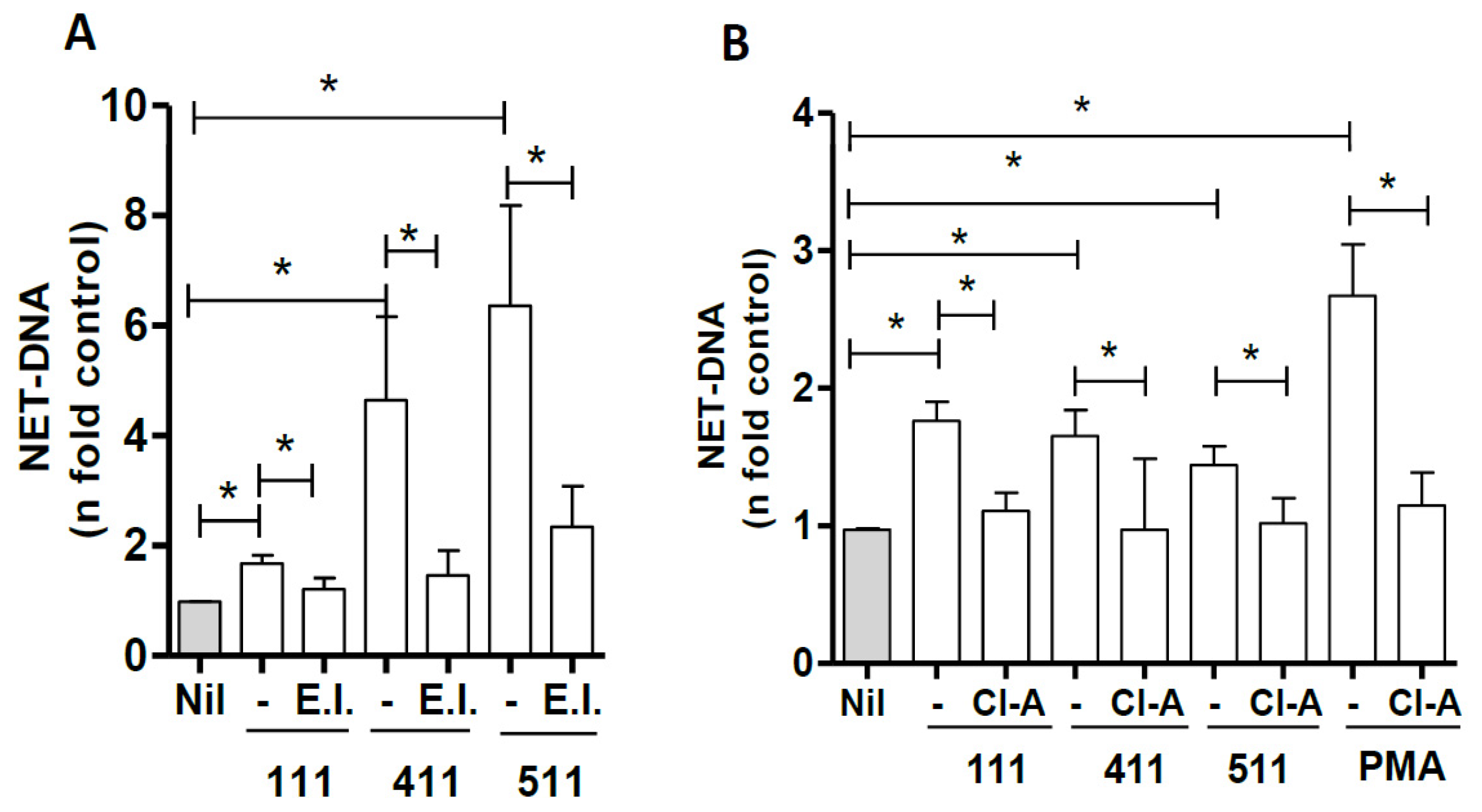
3.3. Signaling Pathways Involved in NET Induction by Laminins
3.4. NET Release by Soluble or Plastic-Adsorbed Laminins
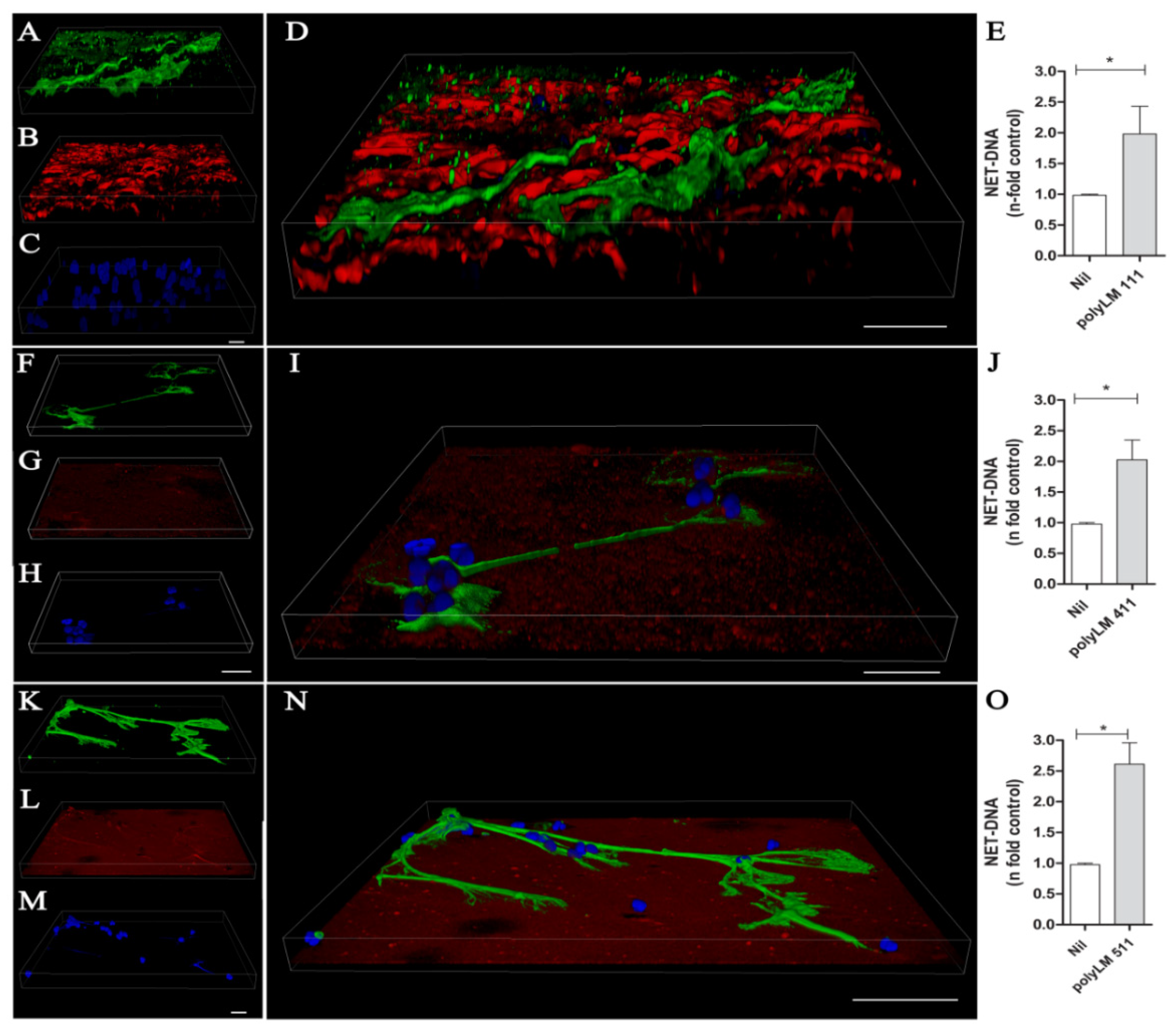
3.5. A Polymerized Form of Laminin Induces NET Release
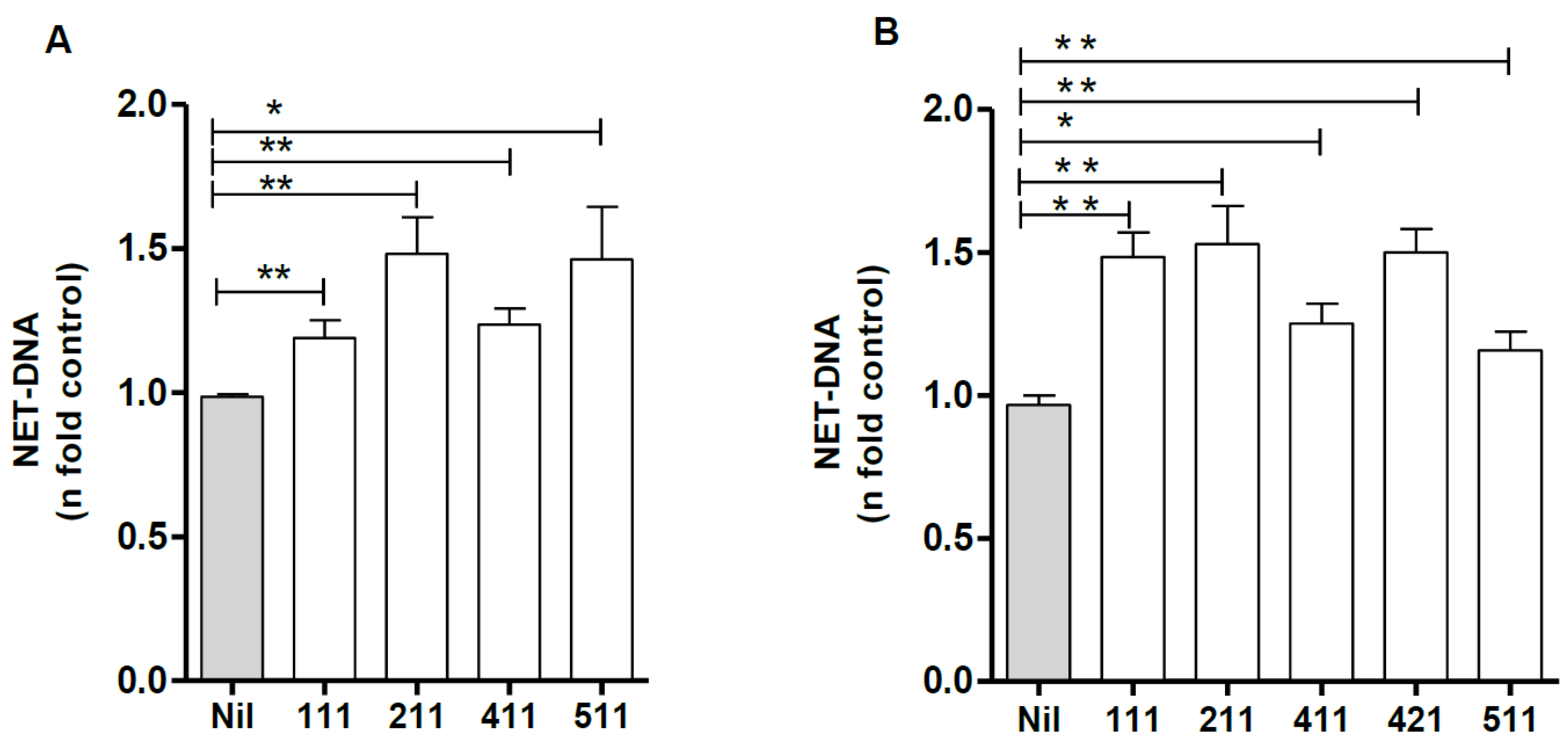
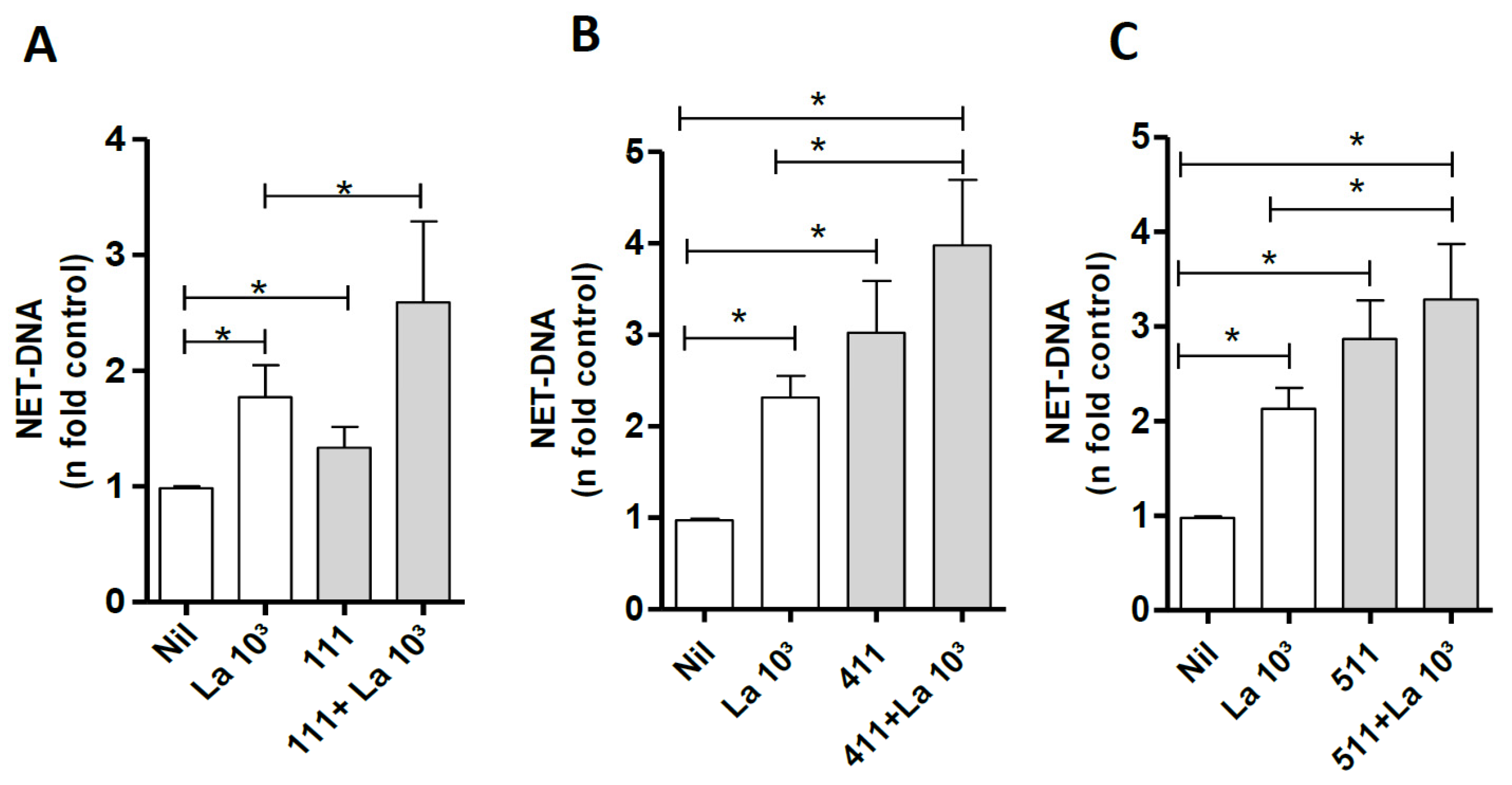
3.6. Laminin Modulates NETs Induced by Leishmania amazonensis
3.7. Leishmania Killing by NET-Rich Supernatants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayannopoulos, V. A myeloperoxidase-cointaining complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; von Bernuth, H.; Zychlinsky, A. Diverse stimuli engage different neutrophil extracellular trap pathways. eLife 2017, 6, e24437. [Google Scholar] [CrossRef]
- Wong, S.L.; Wagner, D.D. Peptidylarginine deiminase 4: A nuclear button triggering neutrophil extracellular traps in inflammatory diseases and aging. FASEB J. 2018, 32, 6358–6370. [Google Scholar] [CrossRef]
- Allen, C.; Thornton, P.; Denes, A.; McColl, B.W.; Pierozynski, A.; Monestier, M.; Pinteaux, E.; Rothwell, N.J.; Allan, S.M. Neutrophil cerebrovascular transmigration triggers rapid neurotoxicity through release of proteases associated with decondensed DNA. J. Immunol. 2012, 189, 381–392. [Google Scholar] [CrossRef]
- McCarthy, J.B.; Furcht, L.T. Laminin and fibronectin promote the haptotactic migration of B16 mouse melanoma cells in vitro. J. Cell Biol. 1984, 98, 1474–1480. [Google Scholar] [CrossRef]
- Devreotes, P.; Janetopoulos, C. Eukaryotic Chemotaxis: Distinctions between Directional Sensing and Polarization. J. Biol. Chem. 2003, 278, 20445–20448. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, D.-H.; Levchenko, A. Topotaxis: A New Mechanism of Directed Cell Migration in Topographic ECM Gradients. Biophys. J. 2018, 114, 1257–1263. [Google Scholar] [CrossRef] [Green Version]
- Aumailley, M.; Bruckner-Tuderman, L.; Carter, W.G.; Deutzmann, R.; Edgar, D.; Ekblom, P.; Engel, J.; Engvall, E.; Hohenester, E.; Jones, J.C.R.; et al. A simpliflied lamin nomenclature. Matrix Biol. 2005, 5, 326–332. [Google Scholar] [CrossRef]
- Durbeej, M. Laminin. Cell Tissue Res. 2010, 339, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, A.; Yurchenco, P.; Iozzo, R.V. The nature and bioloy of basement membranes. Matrix Biol. 2017, 58, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurchenco, P.D.; Patton, B.L. Devepmental and pathogenic mechanism of basement membrane assembly. Curr. Pharm. Des. 2009, 15, 1277–1294. [Google Scholar] [CrossRef] [Green Version]
- Nishiuchi, R.; Takagi, J.; Hayashi, M.; Ido, H.; Yagi, Y.; Sanzen, N.; Tsuji, T.; Yamada, M.; Sekiguchi, K. Ligand-binding specificities of laminin-binding integrins: A comprehensive survey of laminin-integrin interactions using recombinant α3bβ1, α6β1, α7β1 and α6β4 integrins. Matrix Biol. 2006, 25, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Layseca, P.; Icha, J.; Hamidi, H.; Ivaska, J. Integrin trafficking in cells and tissues. Nat. Cell Biol. 2019, 2, 122–132. [Google Scholar] [CrossRef]
- Rieu, P.; Lesavre, P.; Halbwachs-Mecarelli, L. Evidence for integrins other than beta 2 on polymorphonuclear neutrophils: Expression of alpha 6 beta 1 heterodimer. J. Leukoc. Biol. 1993, 53, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Roussel, E.; Gringas, M.C. Transendothelial migration induces rapid expression on neutrophils of granule-release VLA6 used for tissue infiltration. J. Leukoc. Biol. 1997, 62, 356–362. [Google Scholar] [CrossRef]
- Dangerfield, J.; Larbi, K.Y.; Huang, M.T.; Dewar, A.; Nourshargh, S. PECAM-1 (CD31) homophilic interaction up-regulates alpha6beta1 on transmigrated neutrophils in vivo and plays a functional role in the ability of alpha6 integrins to mediate leukocyte migration through the perivascular basement membrane. J. Exp. Med. 2002, 196, 1201–1211. [Google Scholar] [CrossRef]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef] [Green Version]
- Peters, N.C.; Egen, J.G.; Secundino, N.; Debrabant, A.; Kimblin, N.; Kamhawi, S.; Lawyer, P.; Fay, M.P.; Germain, R.N.; Sacks, D. In Vivo Imaging Reveals an Essential Role for Neutrophils in Leishmaniasis Transmitted by Sand Flies. Science 2008, 321, 970–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimaraes-Costa, A.B.; Nascimento, M.T.C.; Froment, G.S.; Soares, R.P.P.; Morgado, F.N.; Conceicao-Silva, F.; Saraiva, E.M. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sixt, M.; Engelhardt, B.; Pausch, F.; Hallmann, R.; Wendler, O.; Sorokin, L.M. Endothelial Cell Laminin Isoforms, Laminins 8 and 10, Play Decisive Roles in T Cell Recruitment across the Blood–Brain Barrier in Experimental Autoimmune Encephalomyelitis. J. Cell Biol. 2001, 153, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Voisin, M.B.; Larbi, K.Y.; Dangerfield, J.; Scheiermann, C.; Tran, M.; Maxwell, P.H.; Sorokin, L.; Nourshargh, S. Venular basement membranes contain specific matrix protein low expression regions that act as exit points for emigrating neutrophils. J. Exp. Med. 2006, 203, 1519–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Zhang, X.; Buscher, K.; Wang, Y.; Wang, H.; Di Russo, J.; Li, L.; Lütke-Enking, S.; Zarbock, A.; Stadtmann, A.; et al. Endothelial Basement Membrane Laminin 511 Contributes to Endothelial Junctional Tightness and Thereby Inhibits Leukocyte Transmigration. Cell Rep. 2017, 18, 1256–1269. [Google Scholar] [CrossRef] [Green Version]
- Di Russo, J.; Luik, A.L.; Yousif, L.; Budny, S.; Oberleithner, H.; Hofschröer, V.; Klingauf, J.; van Bavel, E.; Bakker, E.N.; Hellstrand, P.; et al. Endothelial basement membrane laminin 511 is essential for shear stress response. EMBO J. 2017, 36, 183–201. [Google Scholar] [CrossRef]
- Wondimu, Z.; Geberhiwot, T.; Ingerpuu, S.; Juronen, E.; Xie, X.; Lindbom, L.; Doi, M.; Kortesmaa, J.; Thyboll, J.; Tryggvason, K.; et al. An endothelial laminin isoform, laminin 8 (α4β1γ1), is secreted by blood neutrophils, promotes neutrophil migration and extravasation, and protects neutrophils from apoptosis. Blood 2004, 104, 1859–1866. [Google Scholar] [CrossRef]
- Kenne, E.; Soehnlein, O.; Genové, G.; Rotzius, P.; Eriksson, E.E.; Lindbom, L. Immune cell recruitment to inflammatory loci is impaired in mice deficient in basement membrane protein laminin alpha4. J. Leukoc. Biol. 2010, 88, 523–528. [Google Scholar] [CrossRef]
- Freire, E.; Gomes, F.; Linden, R.; Moura-Neto, V.; Coelho-Sampaio, T. Structure of laminin substrate modulates cellular signaling for neuritogenesis. J. Cell Sci. 2002, 115, 4867–4876. [Google Scholar] [CrossRef] [Green Version]
- Hochman-Mendez, C.; Cantini, M.; Moratal, D.; Salmeron-Sanchez, M.; Coelho-Sampaio, T. A Fractal Nature for Polymerized Laminin. PLoS ONE 2014, 9, e109388. [Google Scholar] [CrossRef] [Green Version]
- Barroso, M.M.; Freire, E.; Limaverde, G.S.; Rocha, G.M.; Batista, E.J.; Weissmüller, G.; Andrade, L.R.; Coelho-Sampaio, T. Artificial laminin polymers assembled in acidic pH mimic basement membrane organization. J. Biol. Chem. 2008, 283, 11714–11720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitroullis, I.; Chavakis, T. α3β1 is INTEGRAL to septic neutrophils. Blood J. Am. Soc. Hematol. 2014, 124, 3507–3509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Simon, T.; Bromberg, J.S. Regulation of the Immune System by Laminins. Trends Immunol. 2017, 38, 858–871. [Google Scholar] [CrossRef]
- Pike, M.C.; Wicha, M.S.; Yoon, P.; Mayo, L.; Boxer, L.A. Laminin promotes the oxidative burst in human neutrophils via increased chemoattractant receptor expression. J. Immunol. 1989, 142, 2004–2011. [Google Scholar]
- Ottonello, L.; Dapino, P.; Amelotti, M.; Barbera, P.; Arduino, N.; Bertolotto, M.; Dallegri, F. Activation of neutrophil respiratory burst by cytokines and chemoattractants. Regulatory role of extracellular matrix glycoproteins. Agents Actions 1998, 47, 345–350. [Google Scholar] [CrossRef]
- Yousif, L.F.; Di Russo, J.; Sorokin, L. Laminin isoforms in endothelial and perivascular basement membranes. Cell Adhes. Migr. 2013, 7, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, H.; Momota, Y.; Murata, K.; Matsushima, H.; Shinkai, H.; Utani, A.; Suzuki, N.; Nomizu, M. Localization of the Laminin α4 Chain in the Skin and Identification of a Heparin-Dependent Cell Adhesion Site Within the Laminin α4 Chain C-Terminal LG4 Module. J. Investig. Dermatol. 2004, 122, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Tsuruta, D.; Ishii, M.; Jones, J.C.; Kobayashi, H. Laminin-332 and -511 in skin. Exp. Dermatol. 2008, 17, 473–480. [Google Scholar] [CrossRef]
- Holmberg, J.; Durbeej, M. Laminin-211 in skeletal muscle function. Cell Adhes. Migr. 2013, 7, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Ekblom, P.; Lonai, P.; Talts, J.F. Expression and biological role of laminin-1. Matrix Biol. 2003, 22, 35–47. [Google Scholar] [CrossRef]
- Raftery, M.J.; Lalwani, P.; Krautkrämer, E.; Peters, T.; Scharffetter-Kochanek, K.; Krüger, R.; Hofmann, J.; Seeger, K.; Krüger, D.H.; Schönrich, G. β2 integrin mediates hantavirus-induced release of neutrophil extracellular traps. J. Exp. Med. 2014, 211, 1485–1497. [Google Scholar] [CrossRef] [PubMed]
- Rossaint, J.; Herter, J.M.; Van Aken, H.; Napirei, M.; Döring, Y.; Weber, C.; Soehnlein, O.; Zarbock, A. Synchronized integrin engagement and chemokine activation is crucial in neutrophil extracellular trap-mediated sterile inflammation. Blood J. Am. Soc. Hematol. 2014, 123, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Gillenius, E.; Urban, C.F. The adhesive protein invasin of Yersinia pseudotuberculosis induces neutrophil extracellular traps via β1 integrins. Microbes Infect. 2015, 17, 327–336. [Google Scholar] [CrossRef]
- Jenne, C.N.; Wong, C.; Zemp, F.J.; McDonald, B.; Rahman, M.M.; Forsyth, P.A.; McFadden, G.; Kubes, P. Neutrophils Recruited to Sites of Infection Protect from Virus Challenge by Releasing Neutrophil Extracellular Traps. Cell Host Microbe 2013, 13, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.C.; Rodrigues, N.C.; Thompson-Souza, G.A.; Muniz, V.S.; Neves, J.S.; Figueiredo, R.T. Mac-1 triggers neutrophil DNA extracellular trap formation to Aspergillus fumigatus independently of PAD4 histone citrullination. J. Leukoc. Biol. 2020, 107, 69–83. [Google Scholar] [CrossRef]
- Yamada, M.; Sekiguchi, K. Molucar basis of laminin-Integrin interacions. Curr. Top. Membr. 2015, 76, 197–229. [Google Scholar]
- Byrd, A.S.; O’Brien, X.M.; Johnson, C.M.; Lavigne, L.M.; Reichner, J.S. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to Candida albicans. J. Immunol. 2013, 190, 4136–4148. [Google Scholar] [CrossRef] [Green Version]
- Menezes, K.; de Menezes, J.R.; Nascimento, M.A.; Santos Rde, S.; Coelho-Sampaio, T. Polylaminin, a polymeric form of laminin, promotes regeneration after spinal cord injury. FASEB J. 2010, 24, 4513–4522. [Google Scholar] [CrossRef]
- Rochael, N.C.; Guimarães-Costa, A.B.; Nascimento, M.T.C.; DeSouza-Vieira, T.S.; De Oliveira, M.P.; Souza, L.F.G.E.; Oliveira, M.F.; Saraiva, E.M. Classical ROS-dependent and early/rapid ROS-independent release of Neutrophil Extracellular Traps triggered by Leishmania parasites. Sci. Rep. 2015, 5, 18302. [Google Scholar] [CrossRef] [Green Version]
- DeSouza-Vieira, T.; Guimarães-Costa, A.; Rochael, N.C.; Lira, M.N.; Nascimento, M.T.; Lima-Gomez, P.S.; Mariante, R.M.; Persechini, P.M.; Saraiva, E.M. Neutrophil extracellular traps release induced by Leishmania: Role of PI3Kγ, ERK, PI3Kσ, PKC, and [Ca2+]. J. Leukoc. Biol. 2016, 100, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Fleury, C.; Jalalvand, F.; Riesbeck, K. Human pathogens utilize host extracellular matrix proteins laminin and collagen for adhesion and invasion of the host. FEMS Microbiol. Rev. 2012, 36, 1122–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, K.; Karmakar, S.; Ghosh, A.; Das, P.K. Role of 67 kDa cell surface laminin binding protein of Leishmania donovani in pathogenesis. J. Biochem. 2001, 130, 141–148. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Oliveira, G.; Linhares-Lacerda, L.; Mattos, T.R.F.; Sanches, C.; Coelho-Sampaio, T.; Riederer, I.; Saraiva, E.M. Laminin Triggers Neutrophil Extracellular Traps (NETs) and Modulates NET Release Induced by Leishmania amazonensis. Biomedicines 2022, 10, 521. https://doi.org/10.3390/biomedicines10030521
Silva-Oliveira G, Linhares-Lacerda L, Mattos TRF, Sanches C, Coelho-Sampaio T, Riederer I, Saraiva EM. Laminin Triggers Neutrophil Extracellular Traps (NETs) and Modulates NET Release Induced by Leishmania amazonensis. Biomedicines. 2022; 10(3):521. https://doi.org/10.3390/biomedicines10030521
Chicago/Turabian StyleSilva-Oliveira, Gustavo, Leandra Linhares-Lacerda, Thayana R. F. Mattos, Camila Sanches, Tatiana Coelho-Sampaio, Ingo Riederer, and Elvira M. Saraiva. 2022. "Laminin Triggers Neutrophil Extracellular Traps (NETs) and Modulates NET Release Induced by Leishmania amazonensis" Biomedicines 10, no. 3: 521. https://doi.org/10.3390/biomedicines10030521