
Immunohistochemical Analysis of Neurotransmitters in Neurosecretory Protein GL-Producing Neurons of the Mouse Hypothalamus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Immunohistochemistry
2.3. NPGL-Immunoreactive Cells Co-Expressing with GAD67 Counting
3. Results
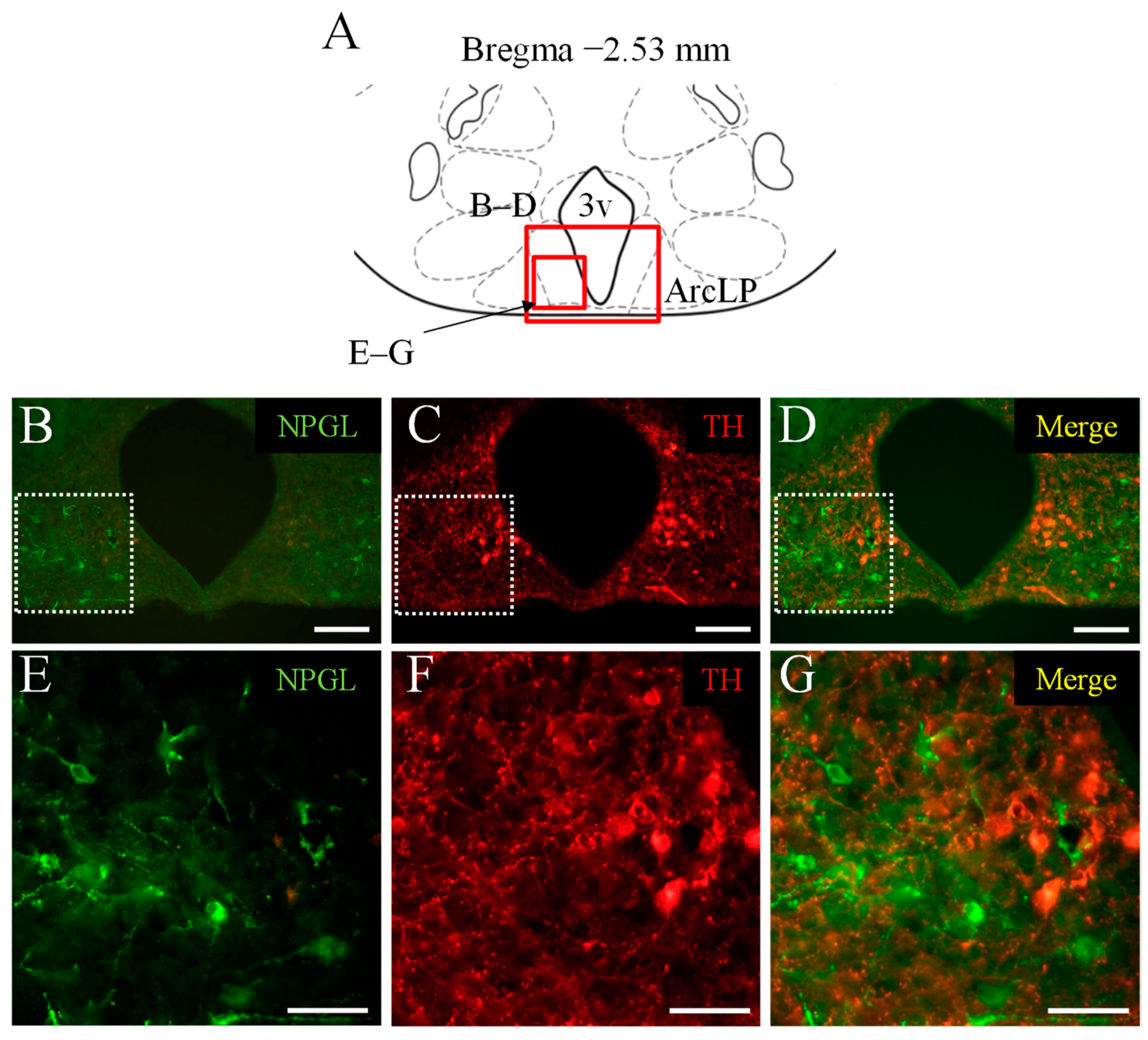
3.1. Double Immunofluorescence Staining of NPGL and TH as a Marker of Catecholaminergic Neurons
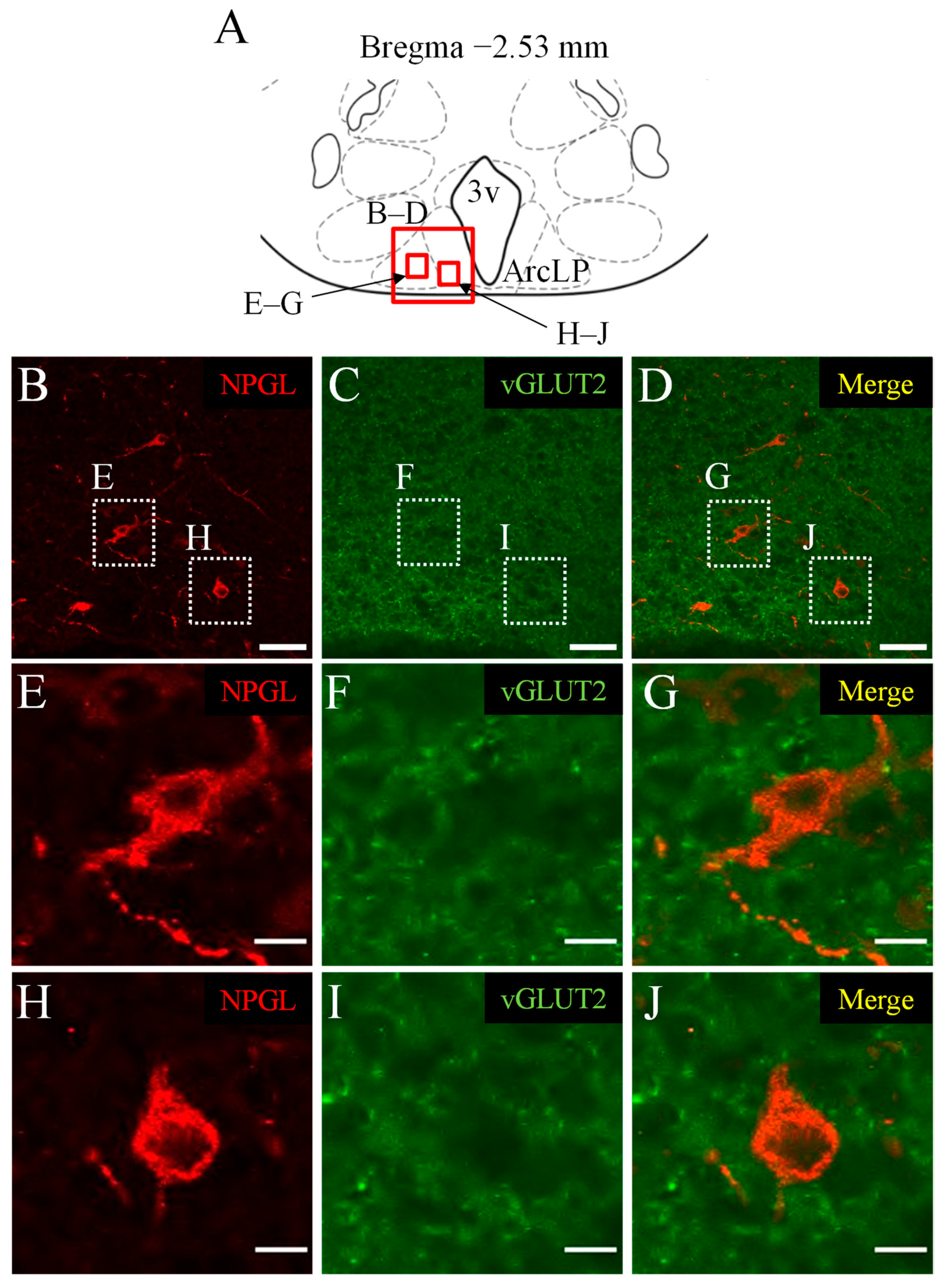
3.2. Double Immunofluorescence Staining of NPGL and vGLUT2 as a Marker of Glutamatergic Neurons
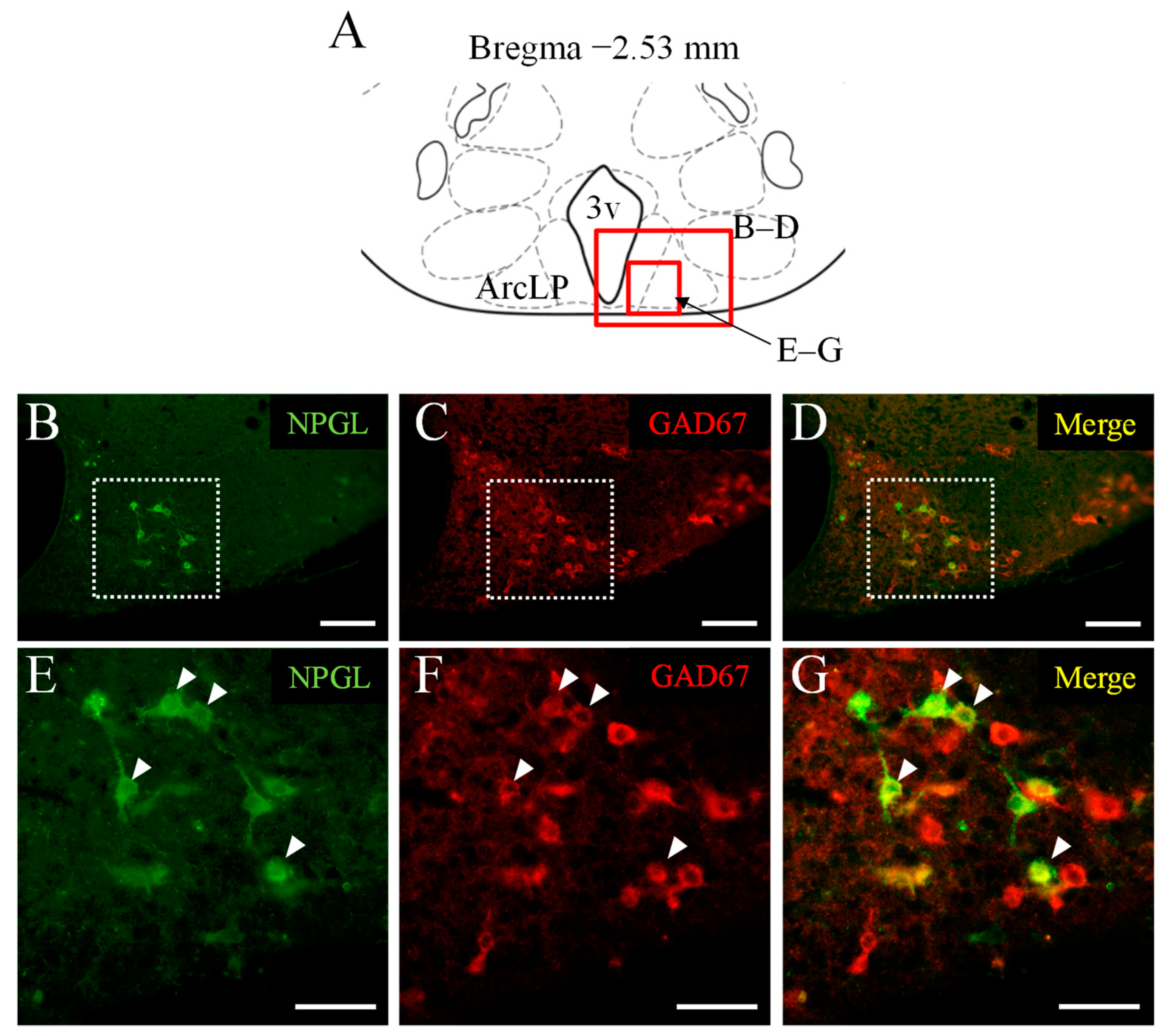
3.3. Double Immunofluorescence Staining of NPGL and GAD67 as a Marker of GABAergic Neurons
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saper, C.B.; Lowell, B.B. The Hypothalamus. Curr. Biol. 2014, 24, R1111–R1116. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.-W. Network of Hypothalamic Neurons That Control Appetite. BMB Rep. 2015, 48, 229–233. [Google Scholar] [CrossRef]
- Wilson, B.D.; Bagnol, D.; Kaelin, C.B.; Ollmann, M.M.; Gantz, I.; Watson, S.J.; Barsh, G.S. Physiological and Anatomical Circuitry between Agouti-Related Protein and Leptin Signaling. Endocrinology 1999, 140, 2387–2397. [Google Scholar] [CrossRef]
- Loh, K.; Herzog, H.; Shi, Y.-C. Regulation of Energy Homeostasis by the NPY System. Trends Endocrinol. Metab. TEM 2015, 26, 125–135. [Google Scholar] [CrossRef]
- Aja, S.; Sahandy, S.; Ladenheim, E.E.; Schwartz, G.J.; Moran, T.H. Intracerebroventricular CART Peptide Reduces Food Intake and Alters Motor Behavior at a Hindbrain Site. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1862–R1867. [Google Scholar] [CrossRef] [Green Version]
- Valassi, E.; Scacchi, M.; Cavagnini, F. Neuroendocrine Control of Food Intake. Nutr. Metab. Cardiovasc. Dis. NMCD 2008, 18, 158–168. [Google Scholar] [CrossRef]
- Williams, G.; Cai, X.J.; Elliott, J.C.; Harrold, J.A. Anabolic Neuropeptides. Physiol. Behav. 2004, 81, 211–222. [Google Scholar] [CrossRef]
- Geary, N. Endocrine Controls of Eating: CCK, Leptin, and Ghrelin. Physiol. Behav. 2004, 81, 719–733. [Google Scholar] [CrossRef]
- Gutzwiller, J.P.; Drewe, J.; Ketterer, S.; Hildebrand, P.; Krautheim, A.; Beglinger, C. Interaction between CCK and a Preload on Reduction of Food Intake Is Mediated by CCK-A Receptors in Humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R189–R195. [Google Scholar] [CrossRef]
- Katsurada, K.; Maejima, Y.; Nakata, M.; Kodaira, M.; Suyama, S.; Iwasaki, Y.; Kario, K.; Yada, T. Endogenous GLP-1 Acts on Paraventricular Nucleus to Suppress Feeding: Projection from Nucleus Tractus Solitarius and Activation of Corticotropin-Releasing Hormone, Nesfatin-1 and Oxytocin Neurons. Biochem. Biophys. Res. Commun. 2014, 451, 276–281. [Google Scholar] [CrossRef]
- Halatchev, I.G.; Ellacott, K.L.J.; Fan, W.; Cone, R.D. Peptide YY3-36 Inhibits Food Intake in Mice through a Melanocortin-4 Receptor-Independent Mechanism. Endocrinology 2004, 145, 2585–2590. [Google Scholar] [CrossRef] [Green Version]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A.; et al. Gut Hormone PYY(3-36) Physiologically Inhibits Food Intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C. Gastrointestinal Satiety Signals I. An Overview of Gastrointestinal Signals That Influence Food Intake. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G7–G13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin Is a Growth-Hormone-Releasing Acylated Peptide from Stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-R.; Chen, H.; Zhou, J.-J.; Pradhan, G.; Sun, Y.; Pan, H.-L.; Li, D.-P. Ghrelin Receptors Mediate Ghrelin-Induced Excitation of Agouti-Related Protein/Neuropeptide Y but Not pro-Opiomelanocortin Neurons. J. Neurochem. 2017, 142, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Beutler, L.R.; Chen, Y.; Ahn, J.S.; Lin, Y.C.; Essner, R.A.; Knight, Z.A. Dynamics of Gut-Brain Communication Underlying Hunger. Neuron 2017, 96, 461–475. [Google Scholar] [CrossRef] [Green Version]
- Vong, L.; Ye, C.; Yang, Z.; Choi, B.; Chua, S.; Lowell, B.B. Leptin Action on GABAergic Neurons Prevents Obesity and Reduces Inhibitory Tone to POMC Neurons. Neuron 2011, 71, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional Cloning of the Mouse Obese Gene and Its Human Homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Koch, L.; Wunderlich, F.T.; Seibler, J.; Könner, A.C.; Hampel, B.; Irlenbusch, S.; Brabant, G.; Kahn, C.R.; Schwenk, F.; Brüning, J.C. Central Insulin Action Regulates Peripheral Glucose and Fat Metabolism in Mice. J. Clin. Investig. 2008, 118, 2132–2147. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef] [Green Version]
- Meister, B. Neurotransmitters in Key Neurons of the Hypothalamus That Regulate Feeding Behavior and Body Weight. Physiol. Behav. 2007, 92, 263–271. [Google Scholar] [CrossRef]
- Meguid, M.M.; Fetissov, S.O.; Varma, M.; Sato, T.; Zhang, L.; Laviano, A.; Rossi-Fanelli, F. Hypothalamic Dopamine and Serotonin in the Regulation of Food Intake. Nutr. Burbank Los Angel. Cty. Calif 2000, 16, 843–857. [Google Scholar] [CrossRef]
- Ramos, E.J.B.; Meguid, M.M.; Campos, A.C.L.; Coelho, J.C.U. Neuropeptide Y, α-Melanocyte-Stimulating Hormone, and Monoamines in Food Intake Regulation. Nutrition 2005, 21, 269–279. [Google Scholar] [CrossRef]
- Yonemochi, N.; Ardianto, C.; Yang, L.; Yamamoto, S.; Ueda, D.; Kamei, J.; Waddington, J.L.; Ikeda, H. Dopaminergic Mechanisms in the Lateral Hypothalamus Regulate Feeding Behavior in Association with Neuropeptides. Biochem. Biophys. Res. Commun. 2019, 519, 547–552. [Google Scholar] [CrossRef]
- Cansell, C.; Denis, R.G.P.; Joly-Amado, A.; Castel, J.; Luquet, S. Arcuate AgRP Neurons and the Regulation of Energy Balance. Front. Endocrinol. 2012, 3, 169. [Google Scholar] [CrossRef] [Green Version]
- Suyama, S.; Yada, T. New Insight into GABAergic Neurons in the Hypothalamic Feeding Regulation. J. Physiol. Sci. JPS 2018, 68, 717–722. [Google Scholar] [CrossRef]
- Ukena, K.; Iwakoshi-Ukena, E.; Taniuchi, S.; Bessho, Y.; Maejima, S.; Masuda, K.; Shikano, K.; Kondo, K.; Furumitsu, M.; Tachibana, T. Identification of a cDNA Encoding a Novel Small Secretory Protein, Neurosecretory Protein GL, in the Chicken Hypothalamic Infundibulum. Biochem. Biophys. Res. Commun. 2014, 446, 298–303. [Google Scholar] [CrossRef]
- Iwakoshi-Ukena, E.; Shikano, K.; Kondo, K.; Taniuchi, S.; Furumitsu, M.; Ochi, Y.; Sasaki, T.; Okamoto, S.; Bentley, G.E.; Kriegsfeld, L.J.; et al. Neurosecretory Protein GL Stimulates Food Intake, de Novo Lipogenesis, and Onset of Obesity. eLife 2017, 6, e28527. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, D.; Shikano, K.; Saito, T.; Iwakoshi-Ukena, E.; Furumitsu, M.; Ochi, Y.; Sato, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Neurosecretory Protein GL, a Hypothalamic Small Secretory Protein, Participates in Energy Homeostasis in Male Mice. Endocrinology 2017, 158, 1120–1129. [Google Scholar] [CrossRef]
- Narimatsu, Y.; Iwakoshi-Ukena, E.; Fukumura, K.; Shikano, K.; Furumitsu, M.; Morishita, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Hypothalamic Overexpression of Neurosecretory Protein GL Leads to Obesity in Male C57BL/6J Mice. Neuroendocrinology 2021. [Google Scholar] [CrossRef]
- Shikano, K.; Iwakoshi-Ukena, E.; Saito, T.; Narimatsu, Y.; Kadota, A.; Furumitsu, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Neurosecretory Protein GL Induces Fat Accumulation in Mice. J. Endocrinol. 2020, 244, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, S.F.; Rossakis, C. Analysis of Feeding Suppression Produced by Perifornical Hypothalamic Injection of Catecholamines, Amphetamines and Mazindol. Eur. J. Pharmacol. 1978, 53, 69–81. [Google Scholar] [CrossRef]
- Shah, B.P.; Vong, L.; Olson, D.P.; Koda, S.; Krashes, M.J.; Ye, C.; Yang, Z.; Fuller, P.M.; Elmquist, J.K.; Lowell, B.B. MC4R-Expressing Glutamatergic Neurons in the Paraventricular Hypothalamus Regulate Feeding and Are Synaptically Connected to the Parabrachial Nucleus. Proc. Natl. Acad. Sci. USA 2014, 111, 13193–13198. [Google Scholar] [CrossRef] [Green Version]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdán, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin Activates Anorexigenic POMC Neurons through a Neural Network in the Arcuate Nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef]
- Trotta, M.; Bello, E.P.; Alsina, R.; Tavella, M.B.; Ferrán, J.L.; Rubinstein, M.; Bumaschny, V.F. Hypothalamic Pomc Expression Restricted to GABAergic Neurons Suppresses Npy Overexpression and Restores Food Intake in Obese Mice. Mol. Metab. 2020, 37, 100985. [Google Scholar] [CrossRef]
- Otgon-Uul, Z.; Suyama, S.; Onodera, H.; Yada, T. Optogenetic Activation of Leptin- and Glucose-Regulated GABAergic Neurons in Dorsomedial Hypothalamus Promotes Food Intake via Inhibitory Synaptic Transmission to Paraventricular Nucleus of Hypothalamus. Mol. Metab. 2016, 5, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Krahn, D.D.; Gosnell, B.A.; Levine, A.S.; Morley, J.E. Behavioral Effects of Corticotropin-Releasing Factor: Localization and Characterization of Central Effects. Brain Res. 1988, 443, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Iigaya, K.; Minoura, Y.; Onimaru, H.; Kotani, S.; Izumizaki, M. Effects of Feeding-Related Peptides on Neuronal Oscillation in the Ventromedial Hypothalamus. J. Clin. Med. 2019, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Bomberg, E.; Levine, A.; Billington, C.; Kotz, C.M. Brain-Derived Neurotrophic Factor in the Ventromedial Nucleus of the Hypothalamus Reduces Energy Intake. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1037–R1045. [Google Scholar] [CrossRef] [Green Version]
- Viskaitis, P.; Irvine, E.E.; Smith, M.A.; Choudhury, A.I.; Alvarez-Curto, E.; Glegola, J.A.; Hardy, D.G.; Pedroni, S.M.A.; Paiva Pessoa, M.R.; Fernando, A.B.P.; et al. Modulation of SF1 Neuron Activity Coordinately Regulates Both Feeding Behavior and Associated Emotional States. Cell Rep. 2017, 21, 3559–3572. [Google Scholar] [CrossRef] [Green Version]
- Orlando, G.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Mollica, A.; Stefanucci, A.; Macedonio, G.; Dimmito, M.P.; Leporini, L.; Menghini, L.; et al. Effects of Kisspeptin-10 on Hypothalamic Neuropeptides and Neurotransmitters Involved in Appetite Control. Mol. Basel Switz. 2018, 23, 3071. [Google Scholar] [CrossRef] [Green Version]
- Leone, S.; Ferrante, C.; Recinella, L.; Chiavaroli, A.; Mollica, A.; Tömböly, C.; Stefanucci, A.; Dimmito, M.P.; Dvorácskó, S.; Verratti, V.; et al. Effects of RVD-Hemopressin (α) on Feeding and Body Weight after Standard or Cafeteria Diet in Rats. Neuropeptides 2018, 72, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Stefanucci, A.; Mollica, A.; Macedonio, G.; Zengin, G.; Ahmed, A.A.; Novellino, E. Exogenous Opioid Peptides Derived from Food Proteins and Their Possible Uses as Dietary Supplements: A Critical Review. Food Rev. Int. 2018, 34, 70–86. [Google Scholar] [CrossRef]
- Skowron, K.; Kurnik-Łucka, M.; Dadański, E.; Bętkowska-Korpała, B.; Gil, K. Backstage of Eating Disorder—About the Biological Mechanisms behind the Symptoms of Anorexia Nervosa. Nutrients 2020, 12, 2604. [Google Scholar] [CrossRef]
- Tanaka, M.; Kádár, K.; Tóth, G.; Telegdy, G. Antidepressant-like Effects of Urocortin 3 Fragments. Brain Res. Bull. 2011, 84, 414–418. [Google Scholar] [CrossRef]
- Tanaka, M.; Schally, A.V.; Telegdy, G. Neurotransmission of the Antidepressant-like Effects of the Growth Hormone-Releasing Hormone Antagonist MZ-4-71. Behav. Brain Res. 2012, 228, 388–391. [Google Scholar] [CrossRef]
- Tanaka, M.; Telegdy, G. Neurotransmissions of Antidepressant-Like Effects of Neuromedin U-23 in Mice. Behav. Brain Res. 2014, 259, 196–199. [Google Scholar] [CrossRef]
- Ibos, K.E.; Bodnár, É.; Bagosi, Z.; Bozsó, Z.; Tóth, G.; Szabó, G.; Csabafi, K. Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats. Biomedicines 2021, 9, 112. [Google Scholar] [CrossRef]
- Małgorzata, P.; Paweł, K.; Iwona, M.L.; Brzostek, T.; Andrzej, P. Glutamatergic Dysregulation in Mood Disorders: Opportunities for the Discovery of Novel Drug Targets. Expert Opin. Ther. Targets 2020, 24, 1187–1209. [Google Scholar] [CrossRef]
- Czapski, G.A.; Strosznajder, J.B. Glutamate and GABA in Microglia-Neuron Cross-Talk in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 11677. [Google Scholar] [CrossRef]
- Tempel, D.L.; Leibowitz, K.J.; Leibowitz, S.F. Effects of PVN Galanin on Macronutrient Selection. Peptides 1988, 9, 309–314. [Google Scholar] [CrossRef]
- Qualls-Creekmore, E.; Yu, S.; Francois, M.; Hoang, J.; Huesing, C.; Bruce-Keller, A.; Burk, D.; Berthoud, H.-R.; Morrison, C.D.; Münzberg, H. Galanin-Expressing GABA Neurons in the Lateral Hypothalamus Modulate Food Reward and Noncompulsive Locomotion. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 6053–6065. [Google Scholar] [CrossRef] [Green Version]
- Melander, T.; Hökfelt, T.; Rökaeus, A.; Cuello, A.C.; Oertel, W.H.; Verhofstad, A.; Goldstein, M. Coexistence of Galanin-like Immunoreactivity with Catecholamines, 5-Hydroxytryptamine, GABA and Neuropeptides in the Rat CNS. J. Neurosci. Off. J. Soc. Neurosci. 1986, 6, 3640–3654. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.A.; Basiri, M.L.; Liu, Y.; Hashikawa, Y.; Hashikawa, K.; Fenno, L.E.; Kim, Y.S.; Ramakrishnan, C.; Deisseroth, K.; Stuber, G.D. Transcriptional and Functional Divergence in Lateral Hypothalamic Glutamate Neurons Projecting to the Lateral Habenula and Ventral Tegmental Area. Neuron 2021, 109, 3823–3837.e6. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| NPGL-immunoreactive cells | 373 ± 22.1 |
| NPGL-immunoreactive cells containing GAD67 (No.) | 188 ± 18.2 |
| NPGL-immunoreactive cells containing GAD67 (%) | 50.1 ± 3.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naito, M.; Iwakoshi-Ukena, E.; Moriwaki, S.; Narimatsu, Y.; Kato, M.; Furumitsu, M.; Miyamoto, Y.; Esumi, S.; Ukena, K. Immunohistochemical Analysis of Neurotransmitters in Neurosecretory Protein GL-Producing Neurons of the Mouse Hypothalamus. Biomedicines 2022, 10, 454. https://doi.org/10.3390/biomedicines10020454
Naito M, Iwakoshi-Ukena E, Moriwaki S, Narimatsu Y, Kato M, Furumitsu M, Miyamoto Y, Esumi S, Ukena K. Immunohistochemical Analysis of Neurotransmitters in Neurosecretory Protein GL-Producing Neurons of the Mouse Hypothalamus. Biomedicines. 2022; 10(2):454. https://doi.org/10.3390/biomedicines10020454
Chicago/Turabian StyleNaito, Mana, Eiko Iwakoshi-Ukena, Shogo Moriwaki, Yuki Narimatsu, Masaki Kato, Megumi Furumitsu, Yuta Miyamoto, Shigeyuki Esumi, and Kazuyoshi Ukena. 2022. "Immunohistochemical Analysis of Neurotransmitters in Neurosecretory Protein GL-Producing Neurons of the Mouse Hypothalamus" Biomedicines 10, no. 2: 454. https://doi.org/10.3390/biomedicines10020454