UHPLC-HRMSn Analysis Reveals the Dynamic Metabonomic Responses of Salvia miltiorrhiza Hairy Roots to Polysaccharide Fraction from Trichoderma atroviride

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Hairy Root Culture
2.3. Polysaccharide Fraction Preparation and Induction
2.4. Sample Extraction Procedures
2.5. UHPLC-MS/MS Measurements
2.6. Data Reduction and Multivariate Data Analysis
3. Results
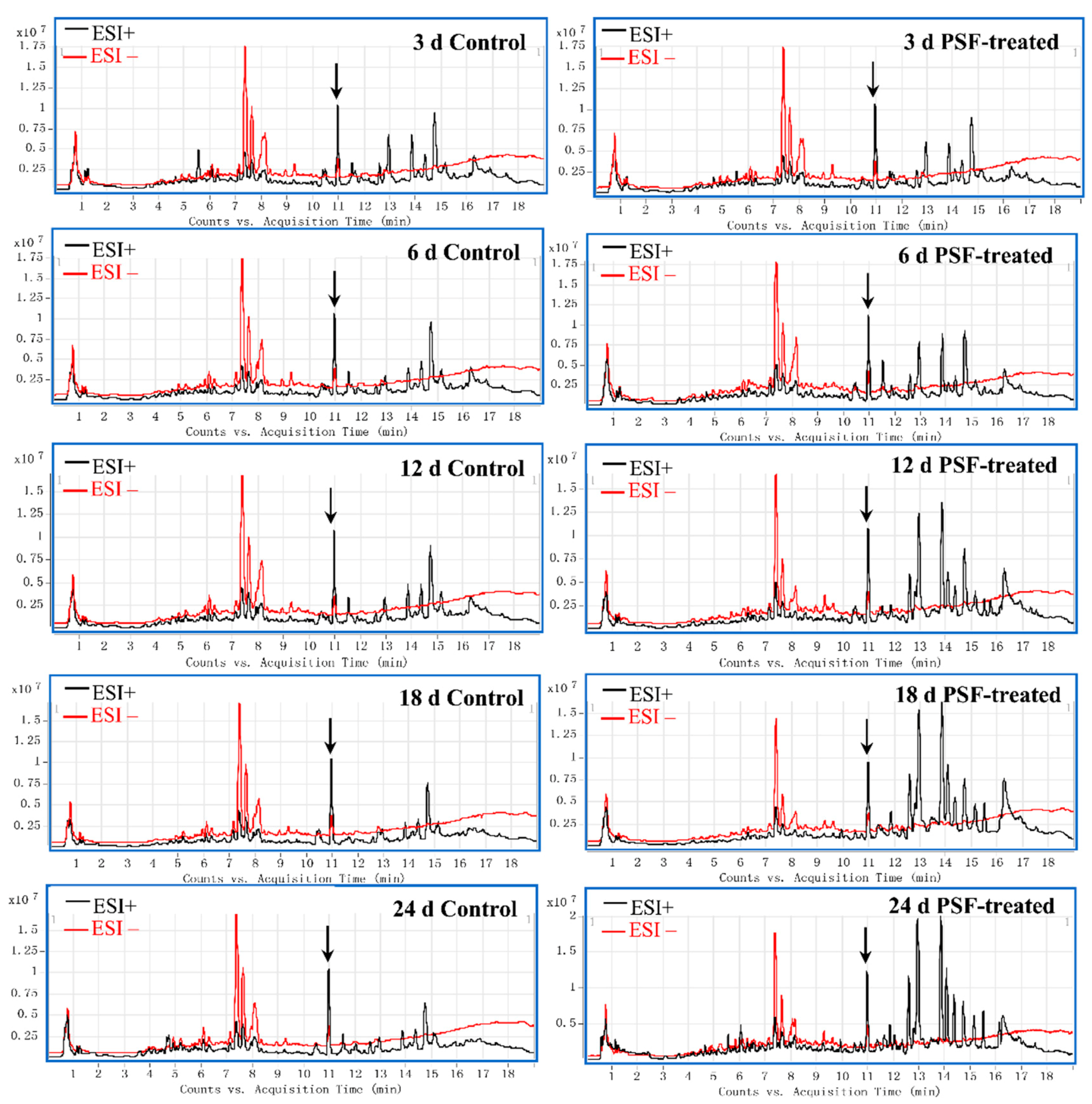
3.1. Global Metabolite Profiling of the Elicitation Response
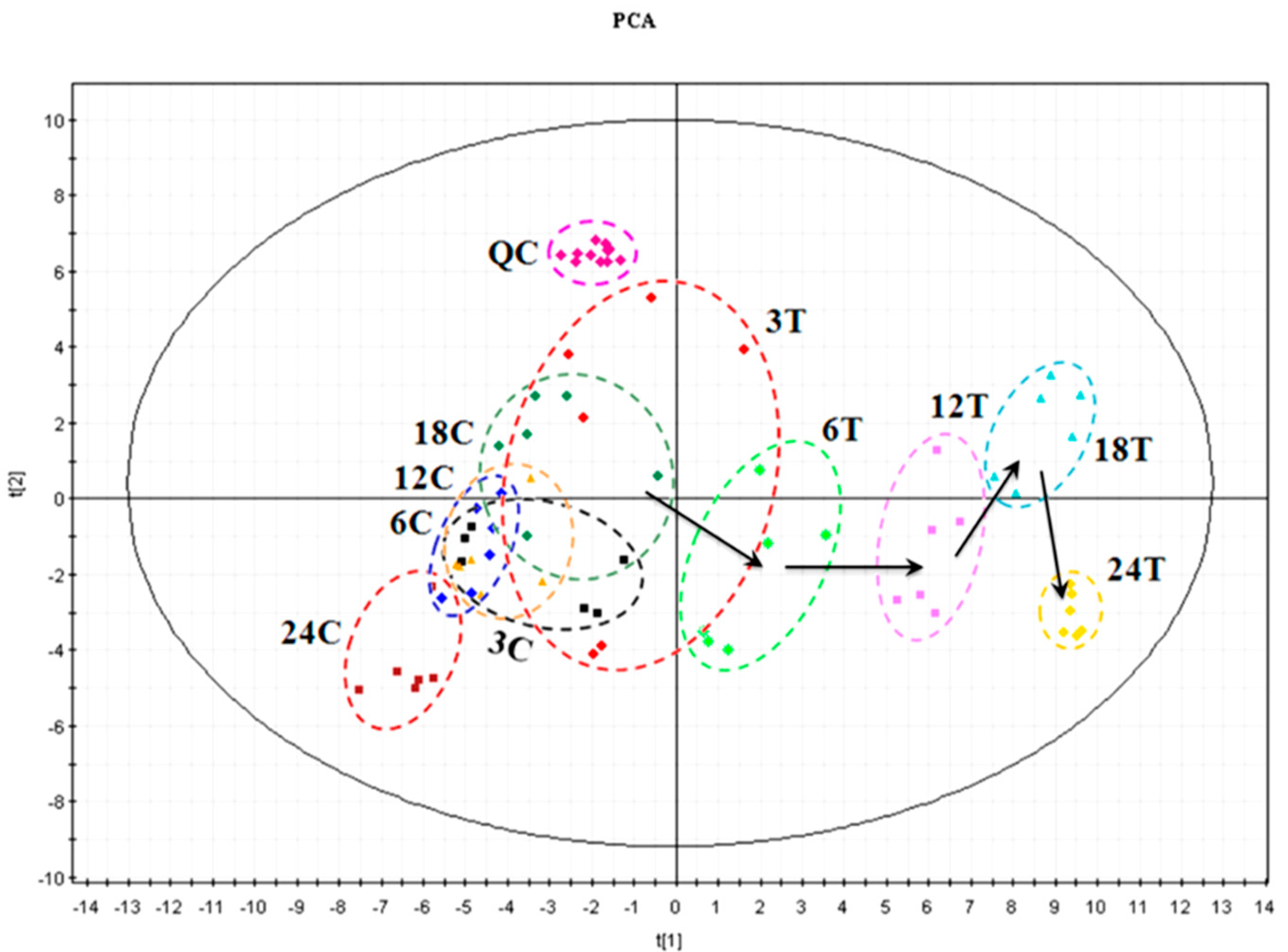
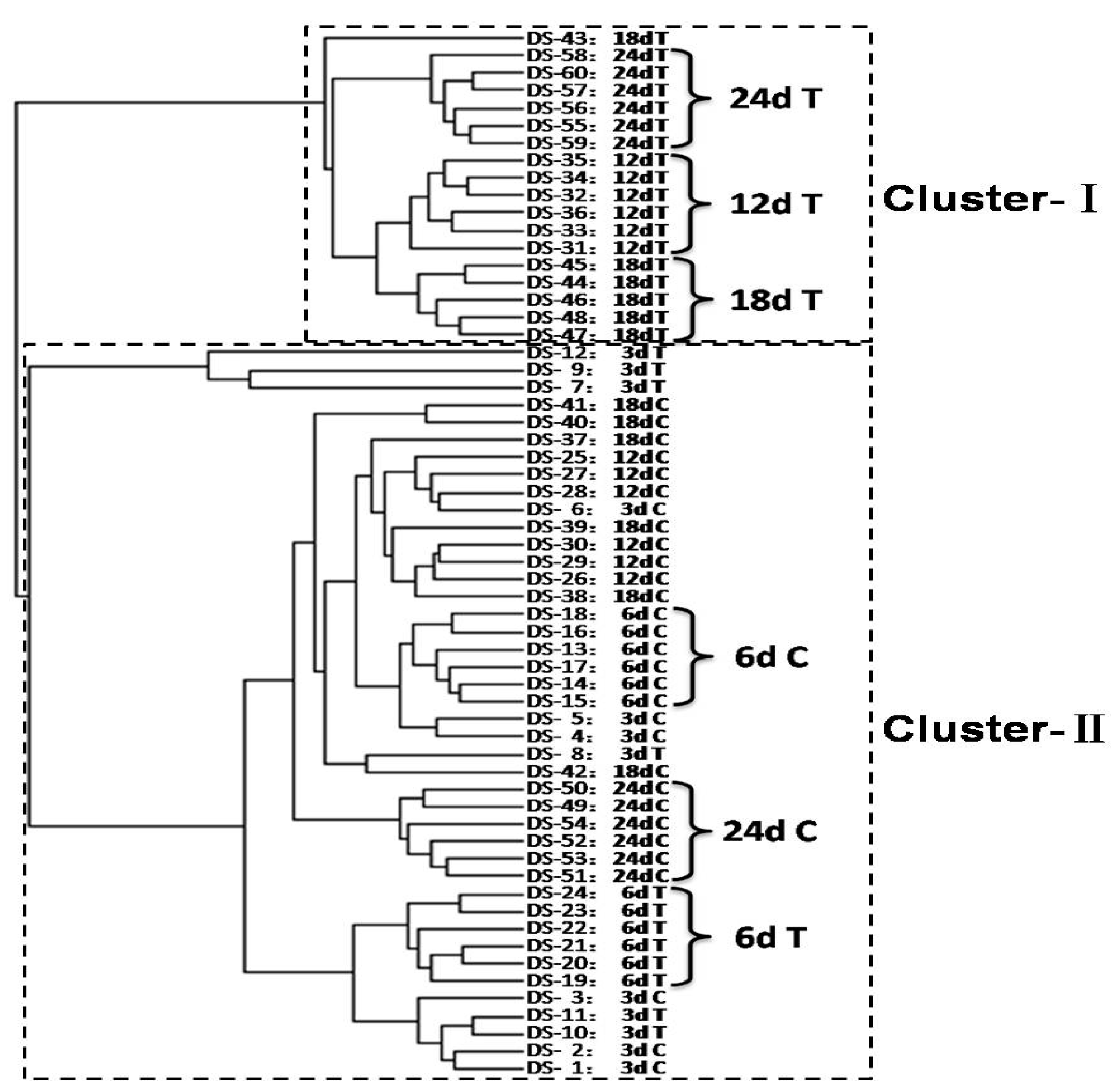
3.2. Modelling for Metabolic Discrimination
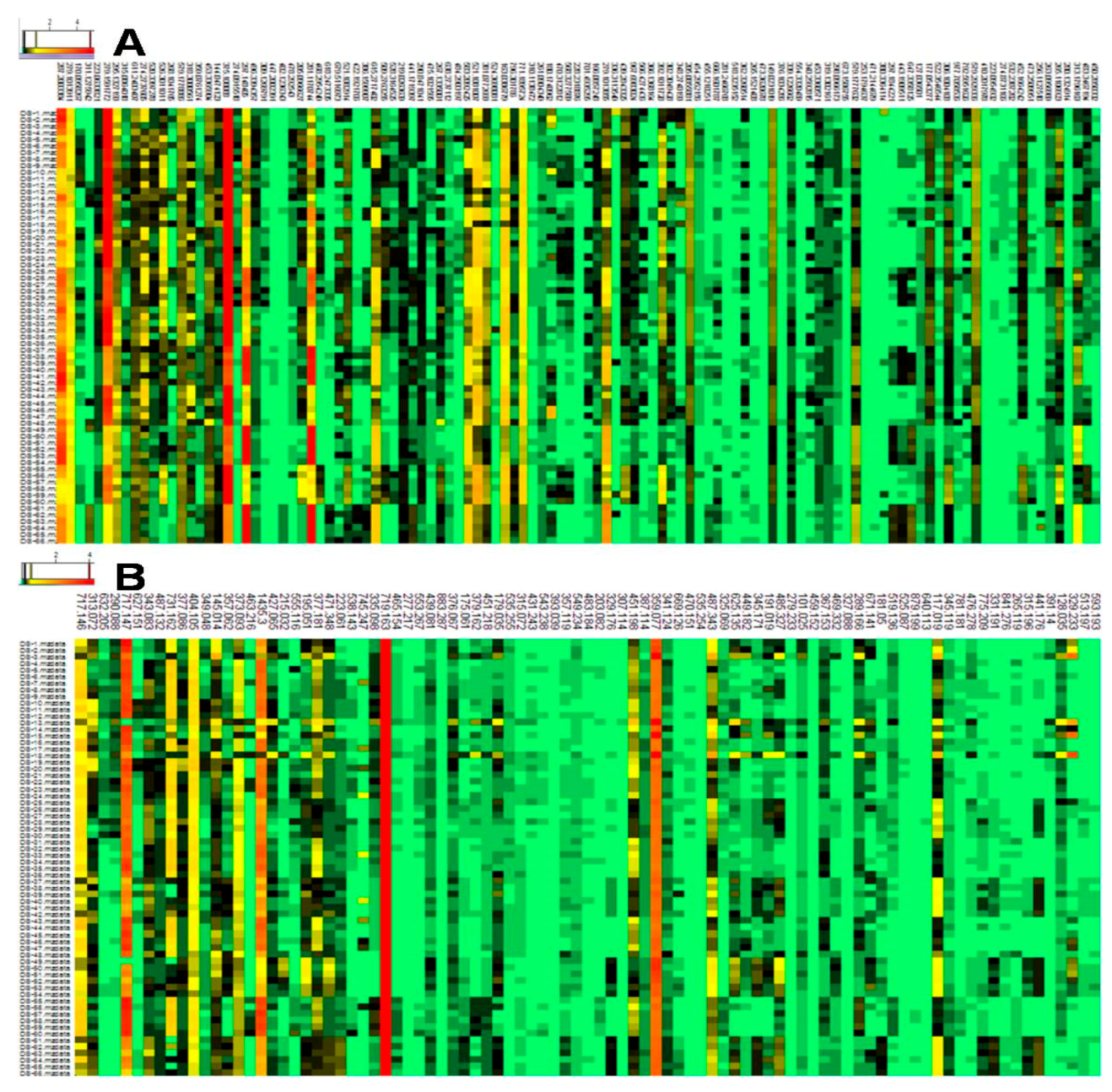
3.3. Screening and Identification of the Differential Metabolites
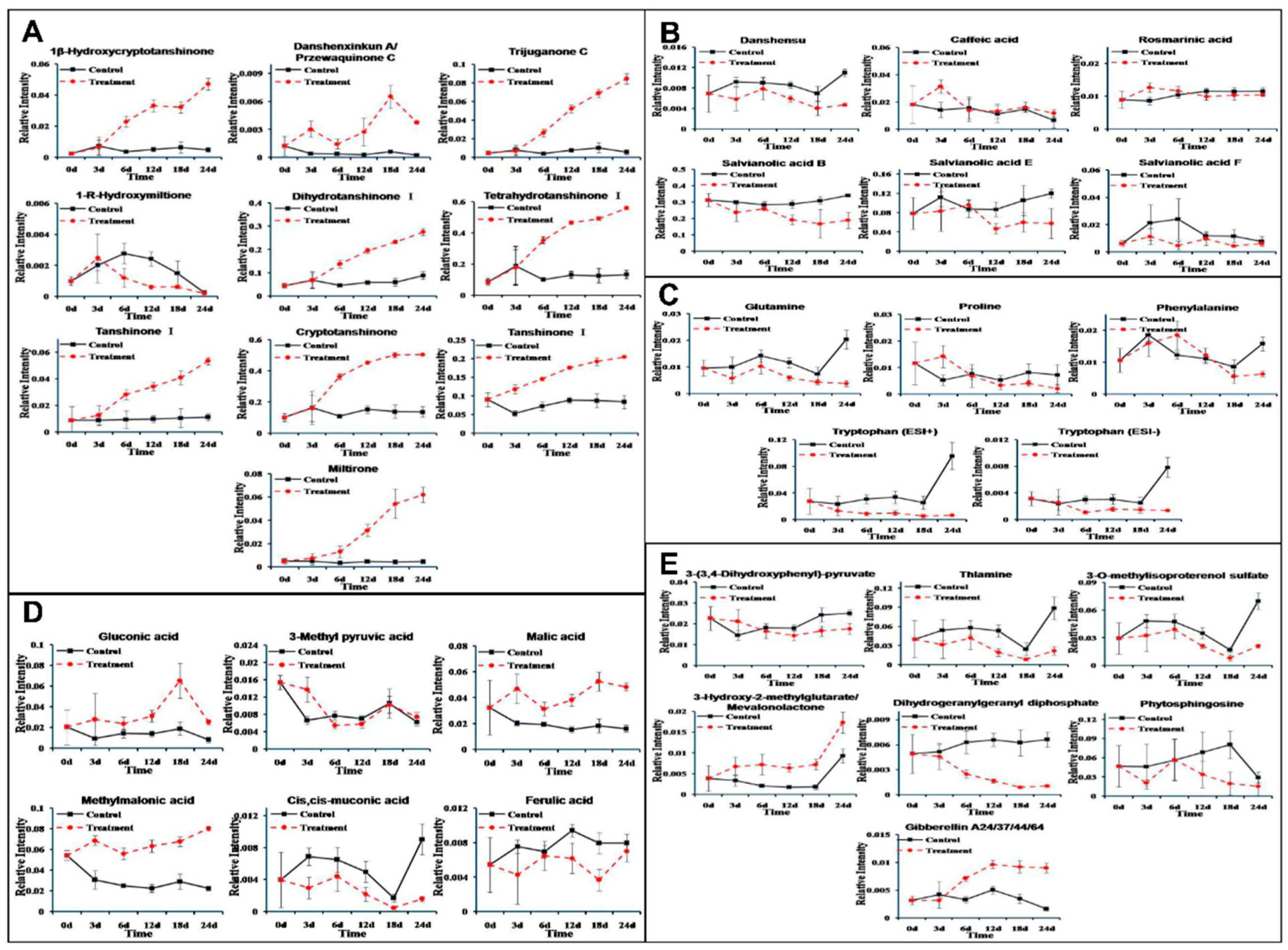
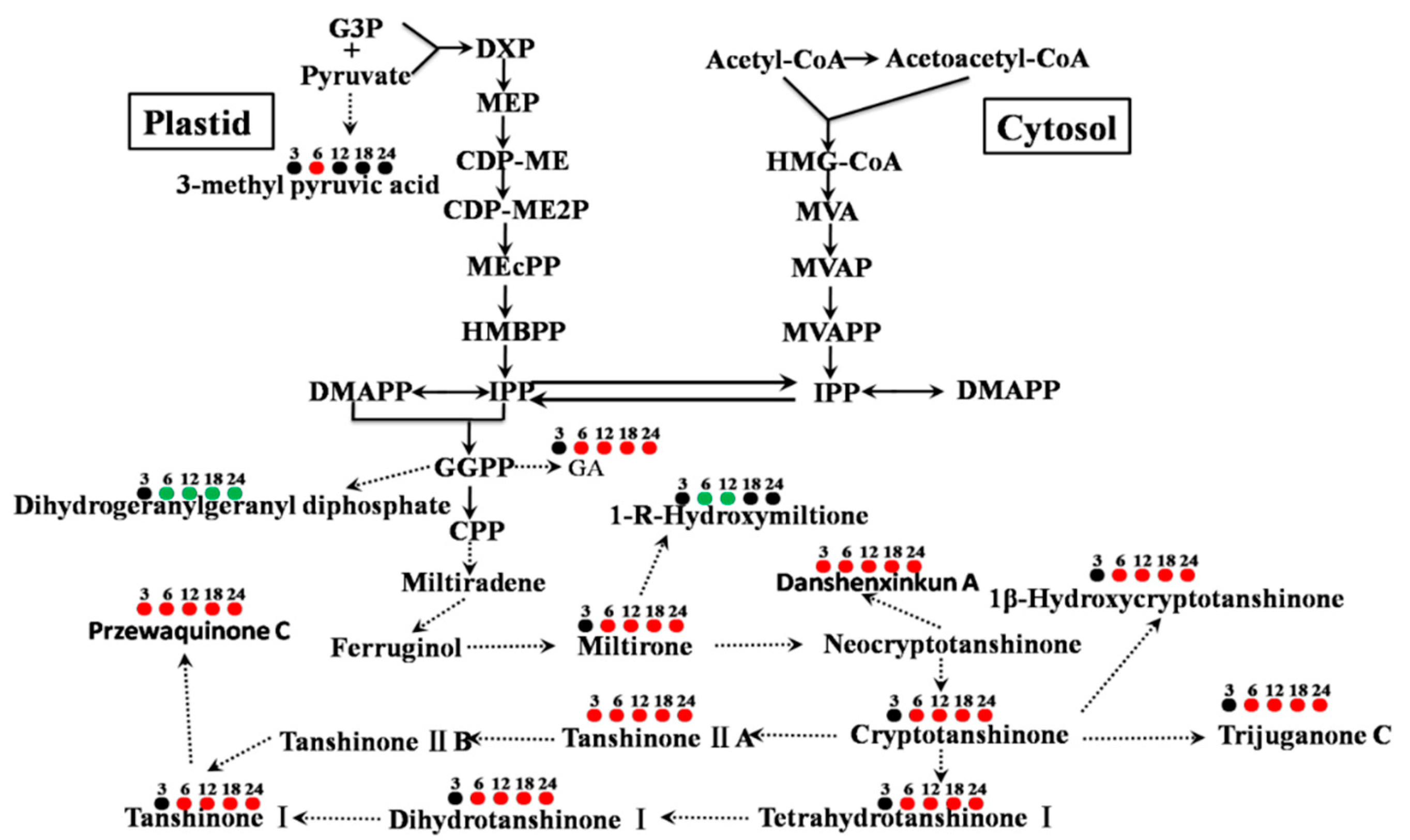
3.4. Metabolite-Specific Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Giauque, H.; Connor, E.W.; Hawkes, C.V. Endophyte Traits Relevant to Stress Tolerance, Resource Use and Habitat of Origin Predict Effects on Host Plants. New Phytol. 2019, 221, 2239–2249. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial Effects of Endophytic Fungi Colonization on Plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef] [PubMed]
- Krings, M.; Taylor, T.N.; Hass, H.; Kerp, H.; Dotzler, N.; Hermsen, E.J. Fungal Endophytes in a 400-million-yr-old Land Plant: Infection Pathways, Spatial Distribution, and Host Responses. New Phytol. 2007, 174, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; Redman, R. More than 400 Million Years of Evolution and Some Plants Still Can’t Make it on their Own: Plant Stress Tolerance via Fungal Symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.G.; Dai, C.C. Jasmonic Acid is Involved in the Signaling Pathway for Fungal Endophyte-induced Volatile Oil Accumulation of Atractylodes lancea Plantlets. BMC Plant Biol. 2012, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-Derived Bioactive Compounds Produced by Endophytic Fungi. Mini Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Su, C.Y.; Ming, Q.L.; Rahman, K.; Han, T.; Qin, L.P. Salvia miltiorrhiza: Traditional Medicinal Uses, Chemistry, and Pharmacology. Chin. J. Nat. Med. 2015, 13, 163–182. [Google Scholar] [CrossRef]
- Wei, T.; Gao, Y.; Deng, K.; Zhang, L.; Yang, M.; Liu, X.; Qi, C.; Wang, C.; Song, W.; Zhang, Y.; et al. Enhancement of Tanshinone Production in Salvia miltiorrhiza Hairy Root Cultures by Metabolic Engineering. Plant Methods 2019, 15, 53. [Google Scholar] [CrossRef]
- Ding, K.; Pei, T.; Bai, Z.; Jia, Y.; Ma, P.; Liang, Z. SmMYB36, a Novel R2R3-MYB Transcription Factor, Enhances Tanshinone Accumulation and Decreases Phenolic Acid Content in Salvia miltiorrhiza Hairy Roots. Sci. Rep. 2017, 7, 5104. [Google Scholar] [CrossRef]
- Xiao, Y.; Gao, S.; Di, P.; Chen, J.; Chen, W.; Zhang, L. Lithospermic Acid B is More Responsive to Silver Ions (Ag+) than Rosmarinic Acid in Salvia miltiorrhiza Hairy Root Cultures. Biosci. Rep. 2010, 30, 33–40. [Google Scholar] [CrossRef]
- Ming, Q.; Han, T.; Li, W.; Zhang, Q.; Zhang, H.; Zheng, C.; Huang, F.; Rahman, K.; Qin, L. Tanshinone IIA and Tanshinone I Production by Trichoderma atroviride D16, an Endophytic Fungus in Salvia miltiorrhiza. Phytomedicine 2012, 19, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Ming, Q.; Su, C.; Zheng, C.; Jia, M.; Zhang, Q.; Zhang, H.; Rahman, K.; Han, T.; Qin, L. Elicitors from the Endophytic Fungus Trichoderma atroviride Promote Salvia miltiorrhiza Hairy Root Growth and Tanshinone Biosynthesis. J. Exp. Bot. 2013, 64, 5687–5694. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.L.; Rudaz, S.; Choi, Y.H.; Kim, H.K. Plant Metabolomics: From Holistic Data to Relevant Biomarkers. Curr. Med. Chem. 2013, 20, 1056–1090. [Google Scholar] [CrossRef] [PubMed]
- Allwood, J.W.; Ellis, D.I.; Goodacre, R. Metabolomic Technologies and their Application to the Study of Plants and Plant–host Interactions. Physiol. Plant. 2008, 132, 117–135. [Google Scholar] [CrossRef] [PubMed]
- T’Kindt, R.; Morreel, K.; Deforce, D.; Boerjan, W.; Van Bocxlaer, J. Joint GC-MS and LC-MS Platforms for Comprehensive Plant Metabolomics: Repeatability and Sample Pre-treatment. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 3572–3580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Peng, W.; Wei, S.; Wei, D.; Li, R.; Liu, J.; Peng, L.; Yang, S.; Gao, Y.; Wu, C.; et al. Guizhi-Shaoyao-Zhimu Decoction Possesses Anti-arthritic Effects on Type II Collagen-induced Arthritis in Rats via Suppression of Inflammatory Reactions, Inhibition of Invasion & Migration and Induction of Apoptosis in Synovial Fibroblasts. Biomed. Pharmacother. 2019, 118, 109367. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhao, X.; Bai, C.; Zhao, C.; Lu, G.; Xu, G. LC-MS-based Metabonomics Analysis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 866, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Gray, N.; Heaton, J.; Musenga, A.; Cowan, D.A.; Plumb, R.S.; Smith, N.W. Comparison of Reversed-phase and Hydrophilic Interaction Liquid Chromatography for the Quantification of Ephedrines using Medium-resolution Accurate Mass Spectrometry. J. Chromatogr. A 2013, 1289, 37–46. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, P.; Ye, M.; Kim, S.H.; Jiang, C.; Lu, J. Tanshinones: Sources, Pharmacokinetics and Anti-cancer Activities. Int. J. Mol. Sci. 2012, 13, 13621–13666. [Google Scholar] [CrossRef]
- Shang, Q.; Xu, H.; Huang, L. Tanshinone IIA: A Promising Natural Cardioprotective Agent. Evid. Based Complement. Altern. Med. 2012, 2012, 716459. [Google Scholar] [CrossRef]
- Jeon, S.J.; Son, K.H.; Kim, Y.S.; Choi, Y.H.; Kim, H.P. Inhibition of Prostaglandin and Nitric Oxide Production in Lipopolysaccharide-treated RAW 264.7 Cells by Tanshinones from the Roots of Salvia miltiorrhiza bunge. Arch. Pharm. Res. 2008, 31, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lou, J.; Mou, Y.; Li, P.; Wu, J.; Zhou, L. Diterpenoid Tanshinones and Phenolic Acids from Cultured Hairy Roots of Salvia miltiorrhiza Bunge and Their Antimicrobial Activities. Molecules 2011, 16, 2259–2267. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kwon, D.Y.; Kim, Y.S.; Kim, Y.C. Peroxyl Radical Scavenging Capacity of Extracts and Isolated Components from Selected Medicinal Plants. Arch. Pharm. Res. 2010, 33, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Morris-Natschke, S.L.; Lee, K.H. Biosynthesis, Total Syntheses, and Antitumor Activity of Tanshinones and Their Analogs as Potential Therapeutic Agents. Nat. Prod. Rep. 2011, 28, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Woo, E.R.; Lee, H.W.; Park, H.R.; Kim, H.N.; Jung, Y.K.; Choi, J.Y.; Chae, S.W.; Kim, H.R.; Chae, H.J. The Correlation of Salvia miltiorrhiza Extract-induced Regulation of Osteoclastogenesis with the Amount of Components Tanshinone I, Tanshinone IIA, Cryptotanshinone, and Dihydrotanshinone. Immunopharmacol. Immunotoxicol. 2008, 30, 347–364. [Google Scholar] [CrossRef]
- Damasceno, S.S.; Dantas, B.B.; Ribeiro-Filho, J.; Antonio, M.A.D.; Galberto, M.d.C.J. Chemical Properties of Caffeic and Ferulic Acids in Biological System: Implications in Cancer Therapy. A Review. Curr. Pharm. Des. 2017, 23, 3015–3023. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point | Ion Model | Type | A | R2X | R2Y | Q2 |
|---|---|---|---|---|---|---|
| 3 d | ESI+ | PCA | 2 | 0.658 | 0.495 | |
| PLS-DA | 4 | 0.761 | 0.991 | 0.864 | ||
| ESI− | PCA | 2 | 0.825 | 0.501 | ||
| PLS-DA | 2 | 0.777 | 0.593 | 0.239 | ||
| 6 d | ESI+ | PCA | 2 | 0.695 | 0.557 | |
| PLS-DA | 2 | 0.675 | 0.991 | 0.959 | ||
| ESI− | PCA | 2 | 0.655 | 0.470 | ||
| PLS-DA | 2 | 0.525 | 0.997 | 0.951 | ||
| 12 d | ESI+ | PCA | 2 | 0.788 | 0.683 | |
| PLS-DA | 2 | 0.744 | 0.996 | 0.974 | ||
| ESI− | PCA | 2 | 0.766 | 0.645 | ||
| PLS-DA | 2 | 0.760 | 0.992 | 0.981 | ||
| 18d | ESI+ | PCA | 3 | 0.880 | 0.635 | |
| PLS-DA | 2 | 0.755 | 0.993 | 0.958 | ||
| ESI− | PCA | 3 | 0.820 | 0.635 | ||
| PLS-DA | 2 | 0.68 | 0.994 | 0.923 | ||
| 24 d | ESI+ | PCA | 2 | 0.846 | 0.729 | |
| PLS-DA | 2 | 0.834 | 0.999 | 0.994 | ||
| ESI− | PCA | 2 | 0.819 | 0.682 | ||
| PLS-DA | 2 | 0.796 | 0.999 | 0.994 | ||
| 3–24 d | ESI+ | PCA | 12 | 0.889 | 0.759 | |
| PLS-DA | 19 | 0.925 | 0.941 | 0.627 | ||
| ESI− | PCA | 10 | 0.879 | 0.757 | ||
| PLS-DA | 13 | 0.897 | 0.966 | 0.79 | ||
| 3–24 d | ESI± | PCA | 11 | 0.876 | 0.771 | |
| PLS-DA | 18 | 0.912 | 0.953 | 0.654 |
| NO. | RT(min) | mz | Ion | Formula | Identification | Fragments |
|---|---|---|---|---|---|---|
| Tanshinones | ||||||
| 1 △ | 9.88 | 313.143 | [M + H]+ | C19H20O4 | 1β-Hydroxycryptotanshinone | 313.1432, 295.1315, 277.1233, 267.1360, 253.1219, 249.1264, 225.0903, 209.0952, 183.0796, 165.0690, 141.0688 |
| 2 ● | 11.74 | 295.098 | [M − H]− | C18H16O4 | Danshenxinkun A/Przewaquinone C | 295.0985, 279.0658, 267.1064, 249.0924, 237.0926, 222.0690, 209.0628, 183.0086, 165.0712, 148.4306 |
| 3 △ | 11.80 | 341.138 | [M + H]+ | C20H20O5 | Trijuganone C | 341.1380, 281.1165, 263.1069, 253.1197, 235.1113, 220.0881, 207.1166, 192.0923, 179.0850, 169.0639, 153.0697, 145.1000, 135.1176, 124.0449, 107.0836 |
| 4 △ | 12.47 | 343.155 | [M − H]− | C19H22O3 | 1-R-Hydroxymiltione | 343.1603, 299.1671, 284.1423, 256.1112, 243.1040, 228.0799, 216.0804, 199.0765, 186.0690, 173.0253, 160.0522, 127.6810, 106.5153 |
| 5 * | 12.55 | 279.101 | [M + H]+ | C18H14O3 | Dihydrotanshinone I | 279.1020, 261.0910, 251.1060, 233.0960, 218.0730, 209.0960, 205.1010, 190.0770, 169.0650 |
| 6 △ | 12.91 | 281.117 | [M + H]+ | C18H16O3 | Tetrahydrotanshinone 1 | 281.1172, 263.1067, 248.0825, 235.1106, 220.0880, 207.1165, 192.0931, 179.0850, 169.0645, 153.0701, 141.0698, 115.0554 |
| 7 * | 13.78 | 277.086 | [M + H]+ | C18H12O3 | Tanshinone I | 277.0860, 262.0620, 249.0910, 234.0670, 221.0960, 203.0850, 193.1010, 178.0770, 169.0650 |
| 8 * | 13.81 | 297.148 | [M + H]+ | C19H20O3 | Cryptotanshinone | 297.1490, 282.1250, 279.1380, 268.1090, 254.0940, 251.1430, 236.1187, 221.0954, 209.0958, 193.1014, 181.1004, 165.0698, 143.0495 |
| 9 * | 15.10 | 295.133 | [M + H]+ | C19H18O3 | Tanshinone IIA | 295.1330, 280.1090, 277.1230, 266.0940, 262.0990, 252.0780, 249.1270, 235.0750, 221.1320, 207.0810 |
| 10 △ | 15.45 | 283.169 | [M + H]+ | C19H22O2 | Miltirone | 283.1710, 268.1492, 253.1211, 240.1145, 225.0914, 207.0813, 195.1163, 180.0922, 165.0698, 153.0711, 139.0041 |
| Phenolic acids | ||||||
| 11 * | 4.07 | 197.045 | [M − H]− | C9H10O5 | Danshensu | 197.0463, 179.0353, 135.0453, 123.0452, 109.0292, 89.0402, 72.9930, 67.0192, 53.0392 |
| 12 * | 5.61 | 179.035 | [M − H]− | C9H8O4 | Caffeic acid | 179.0353, 135.0445, 117.0340, 108.0218, 89.0398, 79.0556, 65.0038 |
| 13 * | 7.34 | 359.078 | [M − H]− | C18H16O8 | Rosmarinic acid | 359.0780, 197.0460, 179.0350, 161.0240, 135.0450, 72.9930 |
| 14 * | 7.78 | 717.147 | [M − H]− | C36H30O16 | Salvianolic acid B | 717.1480,519.0950,321.0420 |
| 15 △ | 8.21 | 717.147 | [M − H]− | C36H30O16 | Salvianolic acid E | 717.1480, 519.0950, 339.0530, 321.0420 |
| 16 △ | 9.24 | 315.086 | [M + H]+ | C17H14O6 | Salvianolic acid F | 163.0390, 145.0285, 135.0440, 117.0335, 107.0493, 89.0387 |
| Amino acids | ||||||
| 17 | 0.70 | 147.076 | [M + H]+ | C5H10N2O3 | Glutamine | 147.0766, 130.0501, 102.0551, 84.0449, 56.0496 |
| 18 | 2.50 | 116.071 | [M + H]+ | C5H9NO4 | Proline | 116.0707, 99.0437, 84.0442, 71.0494, 56.0494 |
| 19 # | 3.54 | 166.086 | [M + H]+ | C9H11NO2 | Phenylalanine | 166.0861, 149.0601, 131.0486, 120.0807, 103.0540, 84.9599, 79.0530 |
| 20 # | 4.71 | 205.097 | [M + H]+ | C11H12N2O2 | Tryptophan | 205.0966, 188.0700, 159.0917, 146.0599, 132.0805, 118.0650 |
| 4.71 | 203.082 | [M − H]− | 203.0382, 186.0552, 159.0930, 142.0665, 116.0510, 74.0253 | |||
| Organic acids | ||||||
| 21 # | 0.71 | 195.051 | [M − H]− | C6H12O7 | Gluconic acid | 195.0512, 177.0394, 159.0294, 129.0194, 99.0288, 75.0092, 59.0139 |
| 22 # | 0.92 | 101.025 | [M − H]− | C4H6O3 | 3-methyl pyruvic acid | 101.0248, 83.0138, 73.0286, 57.0351 |
| 23 # | 0.96 | 133.014 | [M − H]− | C4H6O5 | Malic acid | 133.0143, 115.0039, 89.0249, 71.0142, 59.0144 |
| 24 # | 1.42 | 117.019 | [M − H]− | C4H6O4 | Methylmalonic acid | 117.0193, 99.0089, 73.0302, 55.019 |
| 25 # | 4.09 | 143.034 | [M + Na]+ | C6H6O4 | cis, cis-Muconic acid | 143.0341, 126.0342, 113.0291, 97.9684, 69.0337, 55.0181 |
| 26 | 5.17 | 239.056 | [M + FA − H]− | C10H10O4 | Ferulic acid | 197.0457, 179.035, 135.0452 |
| Other compounds | ||||||
| 27 | 0.69 | 219.026 | [M + Na]+ | C9H8O5 | 3-(3,4-Dihydroxyphenyl)pyruvate | 219.1202, 202.0934, 174.098, 156.0889, 130.0724, 104.0946, 84.0695 |
| 28 # | 0.72 | 265.111 | [M + H]+ | C12H16N4OS | Thiamine | 265.1114, 248.1132, 144.0479, 122.0716 |
| 29 | 3.57 | 306.100 | [M + H]+ | C12H19NO6S | 3-O-Methylisoproterenol Sulfate | 306.0955, 186.0577, 144.0476, 126.037, 113.0291, 99.0261, 85.0284, 69.03367 |
| 30 ● | 5.14 | 175.061 | [M + FA − H]− | C6H10O3 | 3-hydroxy-2-methylglutarate/ Mevalonolactone | 175.0612, 157.0512, 131.0721, 115.0403, 85.0664, 59.0148 |
| 31 | 7.90 | 475.196 | [M + Na]+ | C20H38O7P2 | Dihydrogeranylgeranyl diphosphate | 475.1922, 393.2455, 342.2300, 297.1884, 164.1471, 209.1525, 181.0498, 163.0377, 135.1163, 107.0857, 93.0700, 85.0286, 71.0491, 57.0698 |
| 32 | 10.45 | 318.300 | [M + H]+ | C18H39NO3 | Phytosphingosine | 318.3003, 300.2896, 256.2629, 212.2368, 146.1174, 132.1017, 102.0917, 88.0753, 70.0652, 57.0702 |
| 33 ● | 11.26 | 347.185 | [M + H]+ | C20H26O5 | Gibberellin A44/24/37/64 | 347.1850, 329.1753, 301.1793, 283.1688, 233.0808, 187.0736, 169.0630, 163.0761, 141.0692, 121.1000, 95.0854, 55.0540 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ming, Q.; Dong, X.; Wu, S.; Zhu, B.; Jia, M.; Zheng, C.; Rahman, K.; Han, T.; Qin, L. UHPLC-HRMSn Analysis Reveals the Dynamic Metabonomic Responses of Salvia miltiorrhiza Hairy Roots to Polysaccharide Fraction from Trichoderma atroviride. Biomolecules 2019, 9, 541. https://doi.org/10.3390/biom9100541
Ming Q, Dong X, Wu S, Zhu B, Jia M, Zheng C, Rahman K, Han T, Qin L. UHPLC-HRMSn Analysis Reveals the Dynamic Metabonomic Responses of Salvia miltiorrhiza Hairy Roots to Polysaccharide Fraction from Trichoderma atroviride. Biomolecules. 2019; 9(10):541. https://doi.org/10.3390/biom9100541
Chicago/Turabian StyleMing, Qianliang, Xin Dong, Sijia Wu, Bo Zhu, Min Jia, Chengjian Zheng, Khalid Rahman, Ting Han, and Luping Qin. 2019. "UHPLC-HRMSn Analysis Reveals the Dynamic Metabonomic Responses of Salvia miltiorrhiza Hairy Roots to Polysaccharide Fraction from Trichoderma atroviride" Biomolecules 9, no. 10: 541. https://doi.org/10.3390/biom9100541