CRISPR/Cas9-Mediated Knock-Out of dUTPase in Mice Leads to Early Embryonic Lethality

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
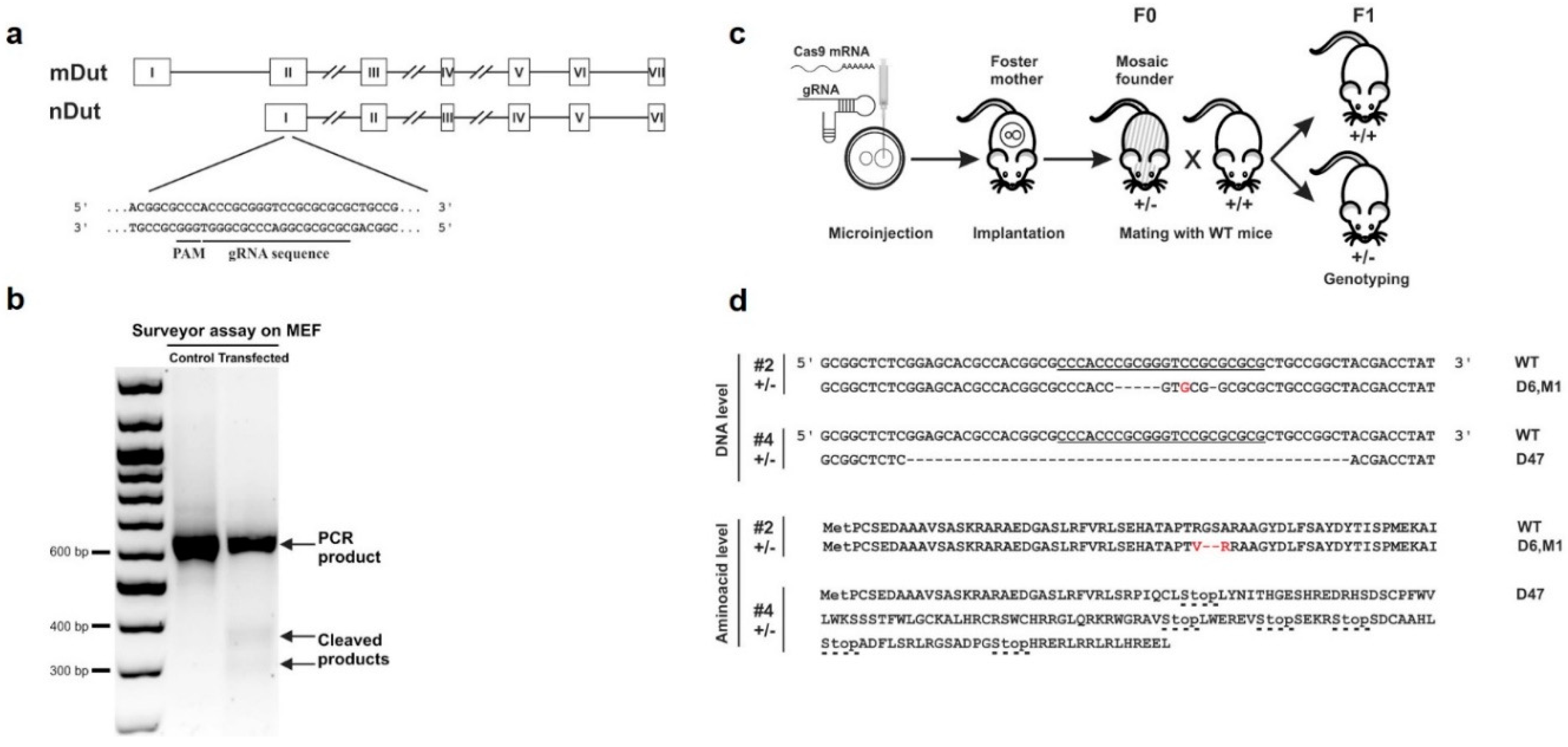
2.1. CRISPR Constructs
2.2. CRISPR/Cas9 Efficiency Test in Mouse Embryonic Fibroblast (MEF) Cells
2.3. Animals
2.4. Micromanipulation and Detection of Gene Targeting
2.5. Cloning and Sequencing
2.6. Off-Target Analysis
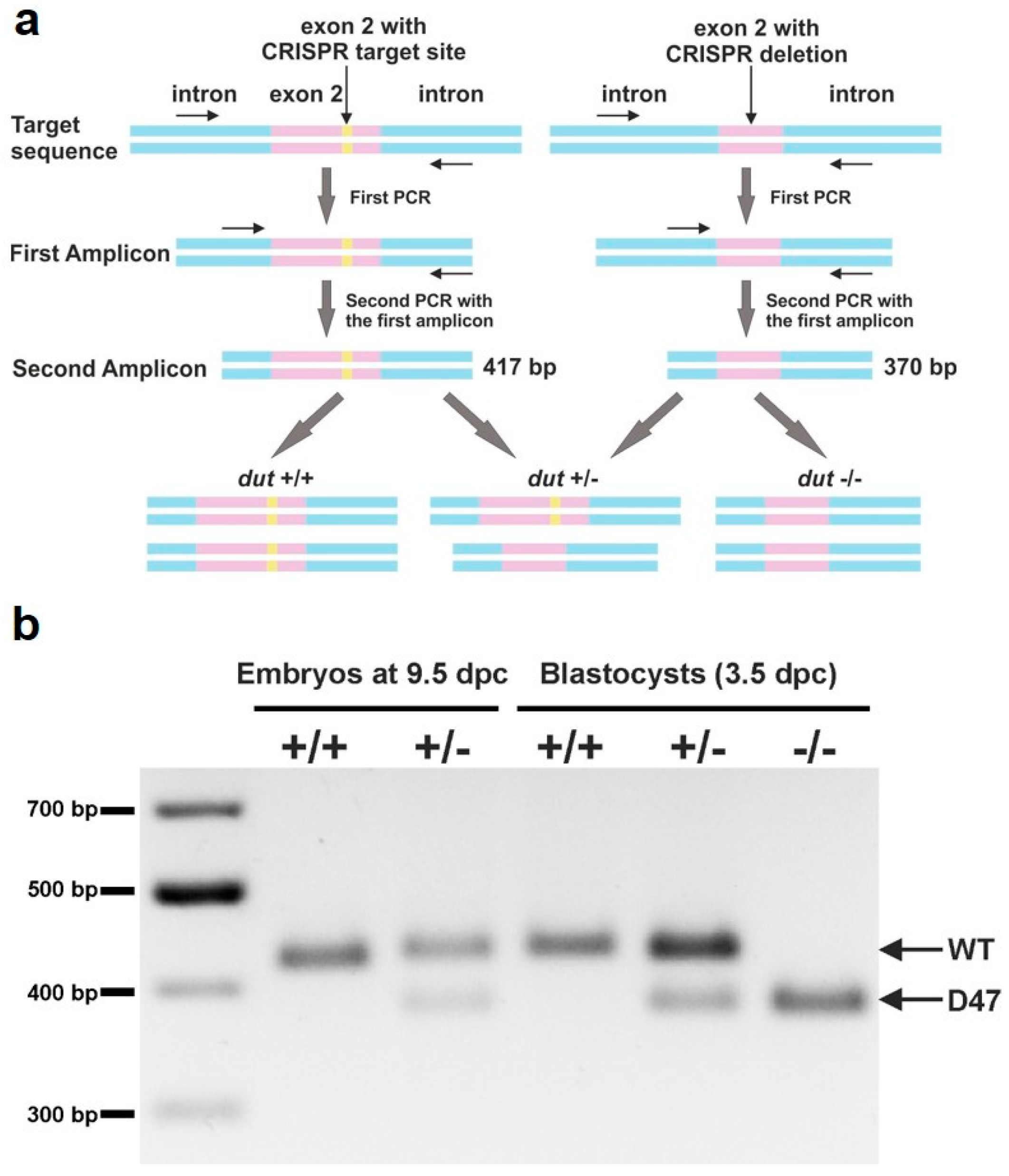
2.7. Genotyping
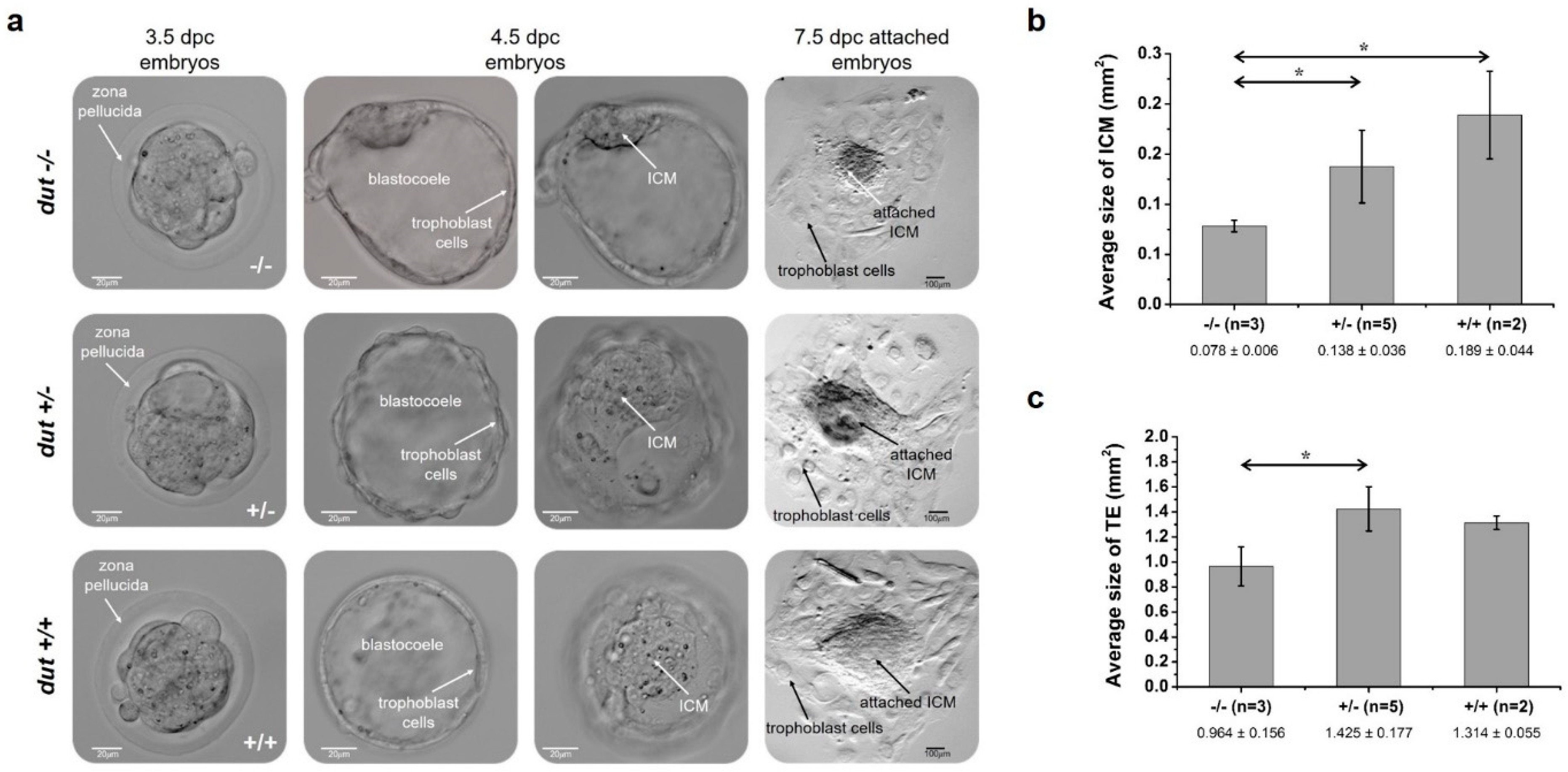
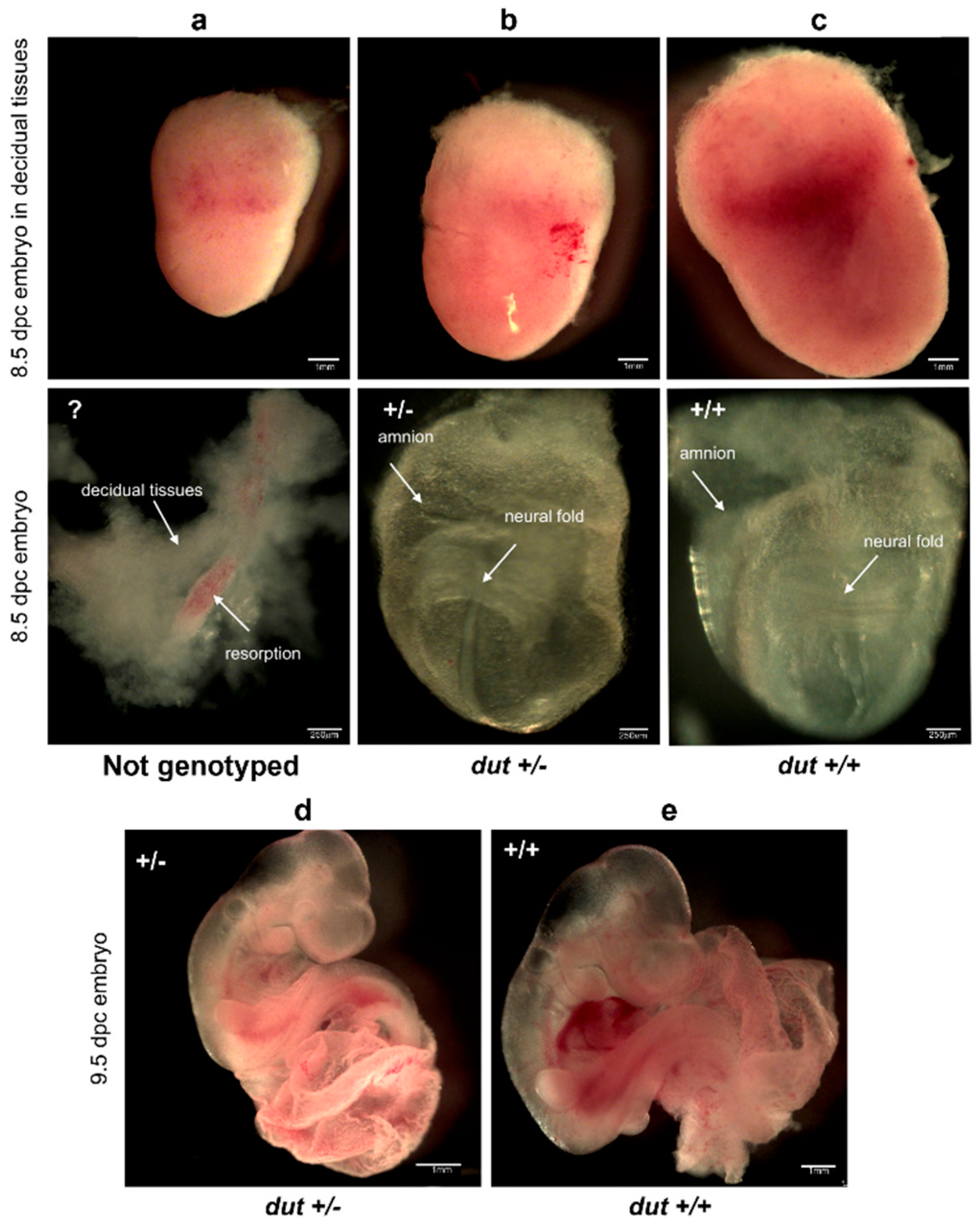
2.8. Analysis of Dissected Embryos
2.9. Analysis of Blastocyst Outgrowth
2.10. Western Blot
2.11. Statistical Analysis
3. Results
3.1. Targeted Knock-Out of Mouse dUTPase by CRISPR/Cas9 Gene Editing
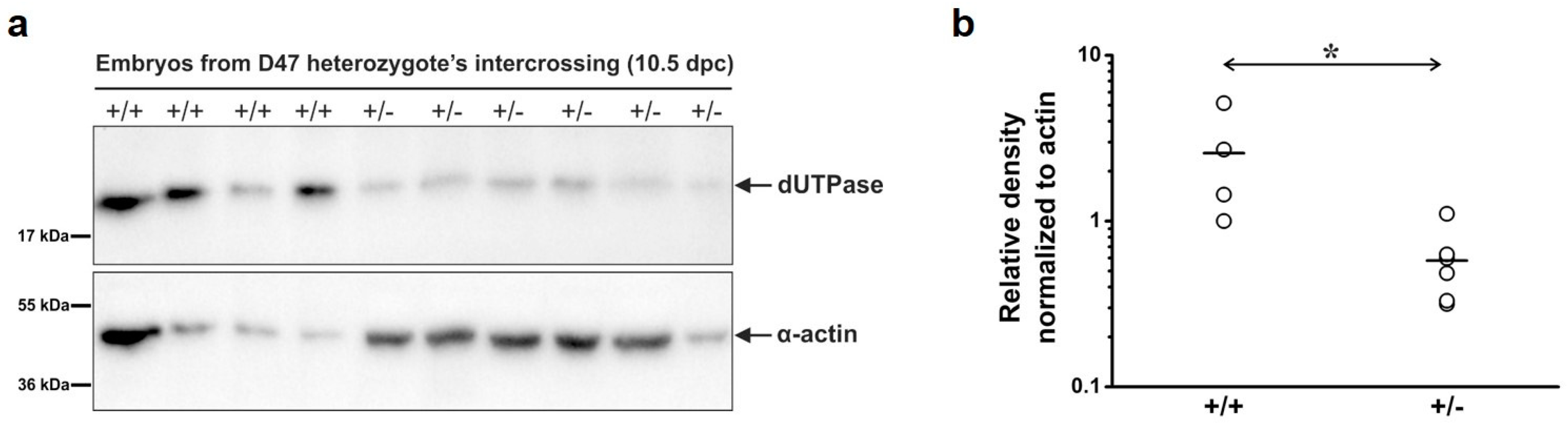
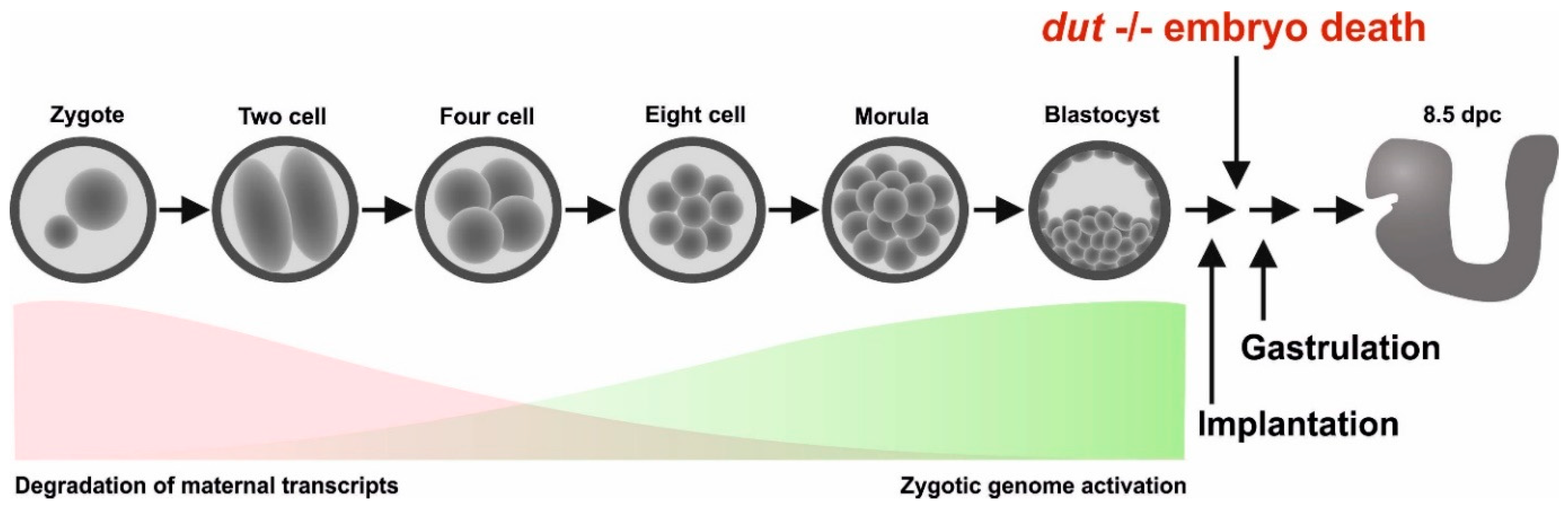
3.2. Analysis of Developmental Effects of dUTPase Knock-Out
3.3. Embryonic Development in the dUTPase Knock-Out as Compared to the Heterozygous and Wild-Type Animals
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mjelle, R.; Hegre, S.A.; Aas, P.A.; Slupphaug, G.; Drabløs, F.; Sætrom, P.; Krokan, H.E. Cell cycle regulation of human DNA repair and chromatin remodeling genes. DNA Repair (Amst). 2015, 30, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Moroz, O.V.; Wilson, K.S.; Murzin, A.G. House cleaning, a part of good housekeeping. Mol. Microbiol. 2006, 59, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakabeppu, Y.; Ohta, E.; Abolhassani, N. MTH1 as a nucleotide pool sanitizing enzyme: Friend or foe? Free Radic. Biol. Med. 2017, 107, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Requena, C.E.; Pérez-Moreno, G.; Ruiz-Pérez, L.M.; Vidal, A.E.; González-Pacanowska, D. The NTP pyrophosphatase DCTPP1 contributes to the homoeostasis and cleansing of the dNTP pool in human cells. Biochem. J. 2014, 459, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.N.; Leveles, I.; Vértessy, B.G. Preventive DNA repair by sanitizing the cellular (deoxy)nucleoside triphosphate pool. FEBS J. 2014, 281, 4207–4223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vértessy, B.G.; Tóth, J. Keeping Uracil Out of DNA: Physiological Role, Structure and Catalytic Mechanism of dUTPases. Acc. Chem. Res. 2009, 42, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Hirmondo, R.; Lopata, A.; Suranyi, E.V.; Vertessy, B.G.; Toth, J. Differential control of dNTP biosynthesis and genome integrity maintenance by the dUTPase superfamily enzymes. Sci. Rep. 2017, 7, 6043. [Google Scholar] [CrossRef]
- Hizi, A.; Herzig, E. dUTPase: The frequently overlooked enzyme encoded by many retroviruses. Retrovirology 2015, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Gaidamaviciute, E.; Tauraite, D.; Gagilas, J.; Lagunavicius, A. Site-directed chemical modification of archaeal Thermococcus litoralis Sh1B DNA polymerase: Acquired ability to read through template-strand uracils. Biochim. Biophys. Acta-Proteins Proteomics 2010, 1804, 1385–1393. [Google Scholar] [CrossRef]
- Hemsworth, G.R.; González-Pacanowska, D.; Wilson, K.S. On the catalytic mechanism of dimeric dUTPases. Biochem. J. 2013, 456, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemsworth, G.R.; Moroz, O.V.; Fogg, M.J.; Scott, B.; Bosch-Navarrete, C.; González-Pacanowska, D.; Wilson, K.S. The crystal structure of the Leishmania major deoxyuridine triphosphate nucleotidohydrolase in complex with nucleotide analogues, dUMP, and deoxyuridine. J. Biol. Chem. 2011, 286, 16470–16481. [Google Scholar] [CrossRef] [PubMed]
- Kerepesi, C.; Szabó, J.E.; Papp-Kádár, V.; Dobay, O.; Szabó, D.; Grolmusz, V.; Vértessy, B.G. Life without dUTPase. Front. Microbiol. 2016, 7, 1768. [Google Scholar] [CrossRef] [PubMed]
- Benedek, A.; Horváth, A.; Hirmondó, R.; Ozohanics, O.; Békési, A.; Módos, K.; Révész, Á.; Vékey, K.; Nagy, G.N.; Vértessy, B.G. Potential steps in the evolution of a fused trimeric all-β dUTPase involve a catalytically competent fused dimeric intermediate. FEBS J. 2016, 283, 3268–3286. [Google Scholar] [CrossRef] [Green Version]
- Varga, B.; Barabás, O.; Takács, E.; Nagy, N.; Nagy, P.; Vértessy, B.G. Active site of mycobacterial dUTPase: Structural characteristics and a built-in sensor. Biochem. Biophys. Res. Commun. 2008, 373, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.; Segelke, B.; Lekin, T.; Krupka, H.; Cho, U.S.; Kim, M.; So, M.; Kim, C.-Y.; Naranjo, C.M.; Rogers, Y.C.; et al. Crystal structure of the Mycobacterium tuberculosis dUTPase: Insights into the catalytic mechanism. J. Mol. Biol. 2004, 341, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Mol, C.D.; Harris, J.M.; McIntosh, E.M.; Tainer, J.A. Human dUTP pyrophosphatase: Uracil recognition by a β hairpin and active sites formed by three separate subunits. Structure 1996, 4, 1077–1092. [Google Scholar] [CrossRef]
- Williams, M.V.; Cox, B.; Ariza, M.E. Herpesviruses dUTPases: A New Family of Pathogen-Associated Molecular Pattern (PAMP) Proteins with Implications for Human Disease. Pathogens 2017, 6, 2. [Google Scholar] [CrossRef]
- Bergman, A.-C.; Nyman, P.O.; Larsson, G. Kinetic properties and stereospecificity of the monomeric dUTPase from herpes simplex virus type 1. FEBS Lett. 1998, 441, 327–330. [Google Scholar] [CrossRef] [Green Version]
- Tarbouriech, N.; Buisson, M.; Seigneurin, J.-M.; Cusack, S.; Burmeister, W.P. The monomeric dUTPase from Epstein-Barr virus mimics trimeric dUTPases. Structure 2005, 13, 1299–1310. [Google Scholar] [CrossRef]
- El-Hajj, H.H.; Zhang, H.; Weiss, B. Lethality of a dut (deoxyuridine triphosphatase) mutation in Escherichia coli. J. Bacteriol. 1988, 170, 1069–1075. [Google Scholar] [CrossRef]
- Gadsden, M.H.; McIntosh, E.M.; Game, J.C.; Wilson, P.J.; Haynes, R.H. dUTP pyrophosphatase is an essential enzyme in Saccharomyces cerevisiae. EMBO J. 1993, 12, 4425–4431. [Google Scholar] [CrossRef] [PubMed]
- Dengg, M.; Garcia-Muse, T.; Gill, S.G.; Ashcroft, N.; Boulton, S.J.; Nilsen, H. Abrogation of the CLK-2 checkpoint leads to tolerance to base-excision repair intermediates. EMBO Rep. 2006, 7, 1046–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.S.; Moriyama, H.; Matsumoto, M. Inhibition of Dr-dut gene causes DNA damage in planarian. Mol. Reprod. Dev. 2018, 85, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Muha, V.; Horváth, A.; Békési, A.; Pukáncsik, M.; Hodoscsek, B.; Merényi, G.; Róna, G.; Batki, J.; Kiss, I.; Jankovics, F.; et al. Uracil-containing DNA in Drosophila: Stability, stage-specific accumulation, and developmental involvement. PLoS Genet. 2012, 8, e1002738. [Google Scholar] [CrossRef] [PubMed]
- Békési, A.; Zagyva, I.; Hunyadi-Gulyás, É.; Pongrácz, V.; Kovári, J.; Nagy, Á.O.; Erdei, A.; Medzihradszky, K.F.; Vértessy, B.G. Developmental regulation of dUTPase in Drosophila melanogaster. J. Biol. Chem. 2004, 279, 22362–22370. [Google Scholar] [CrossRef] [PubMed]
- Dubois, E.; Córdoba-Cañero, D.; Massot, S.; Siaud, N.; Gakière, B.; Domenichini, S.; Guérard, F.; Roldan-Arjona, T.; Doutriaux, M.P. Homologous recombination is stimulated by a decrease in dUTPase in Arabidopsis. PLoS ONE 2011, 6, e18658. [Google Scholar] [CrossRef] [PubMed]
- Koehler, S.E.; Ladner, R.D. Small Interfering RNA-Mediated Suppression of dUTPase Sensitizes Cancer Cell Lines to Thymidylate Synthase Inhibition. Mol. Pharmacol. 2004, 66, 620–626. [Google Scholar]
- Studebaker, A.W.; Lafuse, W.P.; Kloesel, R.; Williams, M.V. Modulation of human dUTPase using small interfering RNA. Biochem. Biophys. Res. Commun. 2005, 327, 306–310. [Google Scholar] [CrossRef]
- Merényi, G.; Kovári, J.; Tóth, J.; Takács, E.; Zagyva, I.; Erdei, A.; Vértessy, B.G. Cellular response to efficient dUTPase RNAi silencing in stable HeLa cell lines perturbs expression levels of genes involved in thymidylate metabolism. Nucleosides Nucleotides Nucleic Acids 2011, 30, 369–390. [Google Scholar] [CrossRef]
- Wilson, P.M.; Danenberg, P.V.; Johnston, P.G.; Lenz, H.-J.; Ladner, R.D. Standing the test of time: Targeting thymidylate biosynthesis in cancer therapy. Nat. Rev. Clin. Oncol. 2014, 11, 282–298. [Google Scholar] [CrossRef]
- Yano, W.; Yokogawa, T.; Wakasa, T.; Yamamura, K.; Fujioka, A.; Yoshisue, K.; Matsushima, E.; Miyahara, S.; Miyakoshi, H.; Taguchi, J.; et al. TAS-114, a First-in-Class Dual dUTPase/DPD Inhibitor, Demonstrates Potential to Improve Therapeutic Efficacy of Fluoropyrimidine-based Chemotherapy. Mol. Cancer Ther. 2018, 17, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Nagashima, H.; Noguchi, K.; Yoshisue, K.; Yokogawa, T.; Matsushima, E.; Tahara, T.; Takagi, S. First-in-human, phase I dose-escalation study of single and multiple doses of a first-in-class enhancer of fluoropyrimidines, a dUTPase inhibitor (TAS-114) in healthy male volunteers. Cancer Chemother. Pharmacol. 2014, 73, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Behringer, R.; Gertsenstein, M.; Nagy, K.V.; Nagy, A. Manipulating the Mouse Embryo: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2014. [Google Scholar]
- Stemmer, M.; Thumberger, T.; Del Sol Keyer, M.; Wittbrodt, J.; Mateo, J.L. CCTop: An intuitive, flexible and reliable CRISPR/Cas9 target prediction tool. PLoS One 2015, 10, e0124633. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2014, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zheng, P.; Dean, J. Maternal control of early mouse development. Development 2010, 137, 859–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadros, W.; Lipshitz, H.D. The maternal-to-zygotic transition: A play in two acts. Development 2009, 136, 3033–3042. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Coordinates | MM | Target Sequence | PAM |
|---|---|---|---|---|
| Dut | chr2:125247853-125247874 | 0 | CGCGCGCGGAC[CCGCGGGT] | GGG |
| Off-1 | chr13:88680616-88680637 | 4 | TGAGTGGGGAC[CCGCGGGT] | TGG |
| Off-2 | chr4:120746882-120746903 | 4 | CTGGAGCGGCC[CCGCGGGT] | GGG |
| Off-3 | chr15:28025084-28025105 | 4 | CGGCCACGGCC[CCGCGGGT] | AGG |
| Off-4 | chr11:23306808-23306829 | 4 | GGGGCGGGGAG[CCGCGGGT] | GGG |
| Off-5 | chr2:118372992-118373013 | 4 | CGCTGGTGGCC[CCGCGGGT] | TGG |
| Off-6 | chr5:64803646-64803667 | 4 | ACCGCACGGAC[GCGCGGGT] | GGG |
| Off-7 | chr18:85179644-85179665 | 3 | CGTGCGCGCAC[GCGCGGGT] | GGG |
| Off-8 | chr9:77319229-77319250 | 4 | CGCGCTTACAC[CCGCGGGT] | GGG |
| Off-9 | chr2:104319532-104319553 | 3 | CGCGTGCGCAC[ACGCGGGT] | AGG |
| Off-10 | chr19:36918725-36918746 | 4 | CTCGCTGGGAC[GCGCGGGT] | AGG |
| Off-11 | chr5:75044665-75044686 | 4 | TGGGCGCGGGC[GCGCGGGT] | GGG |
| Off-12 | chr4:152086570-152086591 | 4 | CGCACCCAGAC[ACGCGGGT] | CGG |
| Off-13 | chr3:41563582-41563603 | 4 | GGCGCGGGGGC[GCGCGGGT] | CGG |
| Off-14 | chr8:60640130-60640151 | 4 | GGCGCGTGGGC[ACGCGGGT] | TGG |
| Off-15 | chr9:40192333-40192354 | 3 | CGCGCGGGGCC[CAGCGGGT] | CGG |
| Off-16 | chr2:174438958-174438979 | 4 | AGCGCGTGGGC[CTGCGGGT] | CGG |
| Off-17 | chr17:88792070-88792091 | 4 | CGGGCGGGGGC[CGGCGGGT] | GGG |
| Off-18 | chr2:28641663-28641684 | 4 | GGCACGGGGAC[CCGGGGGT] | GGG |
| Off-19 | chr17:28350853-28350874 | 4 | GGCGGGCGGGC[CCACGGGT] | GGG |
| Off-20 | chr5:107597539-107597560 | 4 | GGCGCGTGGAT[CGGCGGGT] | AGG |
| DNA Source | Genotype | Resorbed | No. Total | ||
|---|---|---|---|---|---|
| +/+ | +/− | −/− | |||
| Postnatal | 21 | 42 | 0 | NA a | 63 |
| 10.5 dpc | 3 | 5 | 0 | 3 | 11 |
| 9.5 dpc | 5 | 5 | 0 | 0 | 10 |
| 8.5 dpc | 10 | 5 | 0 | 5 | 20 |
| 3.5 dpc | 11 | 13 | 7 | NA | 31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pálinkás, H.L.; Rácz, G.A.; Gál, Z.; Hoffmann, O.I.; Tihanyi, G.; Róna, G.; Gócza, E.; Hiripi, L.; Vértessy, B.G. CRISPR/Cas9-Mediated Knock-Out of dUTPase in Mice Leads to Early Embryonic Lethality. Biomolecules 2019, 9, 136. https://doi.org/10.3390/biom9040136
Pálinkás HL, Rácz GA, Gál Z, Hoffmann OI, Tihanyi G, Róna G, Gócza E, Hiripi L, Vértessy BG. CRISPR/Cas9-Mediated Knock-Out of dUTPase in Mice Leads to Early Embryonic Lethality. Biomolecules. 2019; 9(4):136. https://doi.org/10.3390/biom9040136
Chicago/Turabian StylePálinkás, Hajnalka Laura, Gergely Attila Rácz, Zoltán Gál, Orsolya Ivett Hoffmann, Gergely Tihanyi, Gergely Róna, Elen Gócza, László Hiripi, and Beáta G. Vértessy. 2019. "CRISPR/Cas9-Mediated Knock-Out of dUTPase in Mice Leads to Early Embryonic Lethality" Biomolecules 9, no. 4: 136. https://doi.org/10.3390/biom9040136