Proteasome Subtypes and Regulators in the Processing of Antigenic Peptides Presented by Class I Molecules of the Major Histocompatibility Complex

Abstract
:1. Introduction to the Ubiquitin Proteasome System
2. Role of the Proteasome in Antigen Processing

{kind=link}
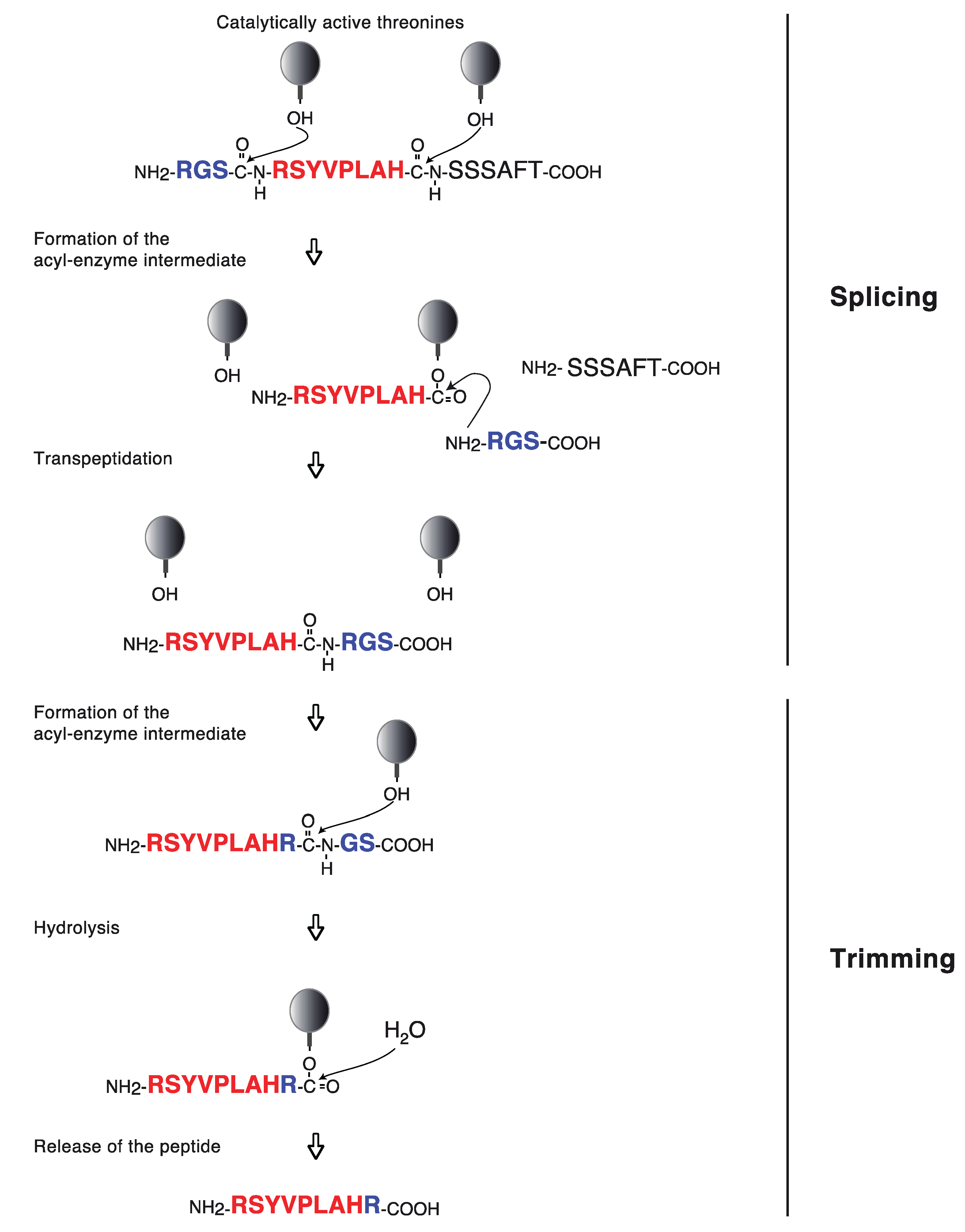{kind=link}
| Peptide source | MHC restriction | Peptide Sequence | Standard Proteasome | Immuno-proteasome | Intermediate Proteasome β5i | Intermediate Proteasome β1i–β5i | References |
|---|---|---|---|---|---|---|---|
| Self | |||||||
| RU134–42 | HLA-B51 | VPYGSFKHV | + + § | +/− | n.d. | n.d. | [20,31] |
| FGF-5172–176 and 217–220 * | HLA-A3 | NTYAS_PRFK | + + | − | n.d. | n.d. | [55,56] |
| Differentiation | |||||||
| gp10040–42 and 47–52 * | HLA-A32 | RTK_QLYPEW | + + | +/− | n.d. | n.d. | [22,56] |
| gp100209–217 | HLA-A2 | ITDQVPFSV | + + | +/− | +/− | +/− | [12,31,38] |
| gp100195–202 and 191 or 192 * | HLA-A3 | RSYVPLAH_R | + + | n.d. | n.d. | n.d. | [57] |
| Tyrosinase369−377 | HLA-A2 | YMDGTMSQV | + + | +/− | − | +/− | [31,38,58] |
| Tyrosinase368–373 and 336–340 * | HLA-A24 | IYMDGT_ADFSF | + + | − | n.d. | n.d. | [59] |
| Melan-A26–35 | HLA-A2 | EAAGIGILTV | + + | − | +/− | +/− | [38,60] |
| Cancer germline | |||||||
| MAGE-A3114–122 | HLA-B40 | AELVHFLLL | − | + + | + + | + + | [31,38,61] |
| MAGE-A3271–279 | HLA-A2 | FLWGPRALV | − | − | + + | − | [32,62] |
| MAGE-A10254–262 | HLA-A2 | GLYDGMEHL | − | − | − | + + | [32,63] |
| MAGE-C2336–344 | HLA-A2 | ALKDVEERV | − | + + | − | + + | [31,38,64] |
| MAGE-C242–50 | HLA-B57 | ASSTLYLVF | − | + + | + + | + + | [38,65] |
| MAGE-C2191–200 | HLA-A2 | LLFGLALIEV | − | − | − | + + | [38,64] |
| Mutation | |||||||
| CLPP240–248 | HLA-A2 | ILDKVLVHL | +/− | + + | n.d. | n.d. | [66] |
| HAUS3154–162 | HLA-A2 | ILNAMIAKI | − | n.d. | n.d. | + + | [67] |
| Polymorphism (mHC ||) | |||||||
| SP110296–301 and 286−289 * | HLA-A3 | SLPRGT_STPK | +/− | + + | n.d. | n.d. | [56,68] |
3. The 20S Proteasome


4. Regulation of the Proteasome Activity
4.1. Proteasome Subtypes
4.1.1. Standard Proteasome and Immunoproteasome
4.1.2. Intermediate Proteasomes
4.1.3. Thymoproteasome
4.1.4. Proteasome Subtypes and Negative Selection of T-Cell Precursors in the Thymus
4.1.5. Structure-Related Properties
4.1.6. Other Functions of the Immunoproteasome
4.2. Proteasome Regulators
4.2.1. The 19S Regulator
4.2.2. The PA28αβ Regulator
| Peptide Source | MHC Restriction | Peptide Sequence | Dependence on PA28αβ | References |
|---|---|---|---|---|
| Viral antigens | ||||
| Murine cytomegalovirus pp89168–176 | H-2Ld | YPHFMPTNL | + | [167,168] |
| Murine Moloney MuLV Gag-leader75–83 | H-2Db | CCLCLTVFL | + | [169] |
| Murine Moloney MuLV gp75-env189–196 | H-2Kb | SSWDFITV | − | [169] |
| LCMV nuceoprotein118–126 | H-2Ld | RPQASGVYM | − | [174] |
| JAK1 tyrosine kinase355–363 | H-2Kd | SYFPEITHI | + | [165] |
| Influenza A/PR/8 nucleoprotein146–154 | H-2Kd | TYQRTRALV | + | [167] |
| Murine tumor Antigens | ||||
| TRP2181–188 | H-2Kb | VYDFFVWL | + | [164] |
| Human Tumor Antigen | ||||
| TRP2360–368 | HLA-A2 | TLDSQVMSL | + | [171,172] |
| TRP2288–296 | HLA-A2 | SLDDYNHLV | − | [172] |
4.2.3. The PA28γ
4.2.4. The PA200 Regulator
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Rock, K.L.; Goldberg, A.L. Degradation of cell proteins and the generation of MHC class I-presented peptides. Annu. Rev. Immunol. 1999, 17, 739–779. [Google Scholar] [CrossRef] [PubMed]
- Tomko, R.J., Jr.; Hochstrasser, M. Molecular architecture and assembly of the eukaryotic proteasome. Annu. Rev. Biochem. 2013, 82, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Akopian, T.N.; Woo, K.M.; Goldberg, A.L. The sizes of peptides generated from protein by mammalian 26 and 20S proteasomes. J. Biol. Chem. 1999, 274, 3363–3371. [Google Scholar] [CrossRef] [PubMed]
- Reits, E.; Griekspoor, A.; Neijssen, J.; Groothuis, T.; Jalink, K.; van Veelen, P.; Janssen, H.; Calafat, J.; Drijfhout, J.W.; Neefjes, J. Peptide diffusion, protection, and degradation in nuclear and cytoplasmic compartments before antigen presentation by MHC class I. Immunity 2003, 18, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Androlewicz, M.J.; Anderson, K.S.; Cresswell, P. Evidence that transporters associated with antigen processing translocate a major histocompatibility complex class I-binding peptide into the endoplasmic reticulum in an ATP-dependent manner. Proc. Natl. Acad. Sci. USA 1993, 90, 9130–9134. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Tsomides, T.J.; Sykulev, Y.; Eisen, H.N. Variations in the number of peptide-MHC class I complexes required to activate cytotoxic T-cell responses. J. Immunol. 1995, 154, 567–576. [Google Scholar] [PubMed]
- Coulie, P.G.; Van den Eynde, B.J.; van der Bruggen, P.; Boon, T. Tumour antigens recognized by T-lymphocytes: At the core of cancer immunotherapy. Nat. Rev. Cancer 2014, 14, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, N.; Van den Eynde, B.J. Insights into the processing of MHC class I ligands gained from the study of human tumor epitopes. Cell. Mol. Life Sci. 2011, 68, 1503–1520. [Google Scholar] [CrossRef] [PubMed]
- Wölfel, T.; Hauer, M.; Schneider, J.; Serrano, M.; Wölfel, C.; Klehmann-Hieb, E.; de Plaen, E.; Hankeln, T.; Meyer zum Büschenfelde, K.-H.; Beach, D. A p16INK4a-insensitive CDK4 mutant targeted by cytolytic T-lymphocytes in a human melanoma. Science 1995, 269, 1281–1284. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, N.; Ooms, A.; Morel, S.; Degiovanni, G.; Van den Eynde, B. Identification of a new peptide recognized by autologous cytolytic T-lymphocytes on a human melanoma. Cancer Immun. 2002, 2, 9–18. [Google Scholar] [PubMed]
- Coulie, P.G.; Brichard, V.; van Pel, A.; Wölfel, T.; Schneider, J.; Traversari, C.; Mattei, S.; de Plaen, E.; Lurquin, C.; Szikora, J.-P.; et al. A new gene coding for a differentiation antigen recognized by autologous cytolytic T-lymphocytes on HLA-A2 melanomas. J. Exp. Med. 1994, 180, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Eliyahu, S.; Jennings, C.; Sakaguchi, K.; Kang, X.; Southwood, S.; Robbins, P.F.; Sette, A.; Appella, E.; Rosenberg, S.A. Recognition of multiple epitopes in the human melanoma antigen gp100 by tumor-infiltrating T-lymphocytes associated with in vivo tumor regression. J. Immunol. 1995, 154, 3961–3968. [Google Scholar] [PubMed]
- Van der Bruggen, P.; Traversari, C.; Chomez, P.; Lurquin, C.; de Plaen, E.; Van den Eynde, B.; Knuth, A.; Boon, T. A gene encoding an antigen recognized by cytolytic T-lymphocytes on a human melanoma. Science 1991, 254, 1643–1647. [Google Scholar]
- Michalek, M.T.; Grant, E.P.; Gramm, C.; Goldberg, A.L.; Rock, K.L. A role for the ubiquitin-dependent proteolytic pathway in MHC class I-restricted antigen presentation. Nature 1993, 363, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shahar, S.; Cassouto, B.; Novak, L.; Porgador, A.; Reiss, Y. Production of a specific major histocompatibility complex class I-restricted epitope by ubiquitin-dependent degradation of modified ovalbumin in lymphocyte lysate. J. Biol. Chem. 1997, 272, 21060–21066. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.P.; Michalek, M.T.; Goldberg, A.L.; Rock, K.L. Rate of antigen degradation by the ubiquitin-proteasome pathway influences MHC class I presentation. J. Immunol. 1995, 155, 3750–3758. [Google Scholar] [PubMed]
- Townsend, A.; Bastin, J.; Gould, K.; Brownlee, G.; Andrew, M.; Coupar, B.; Boyle, D.; Chan, S.; Smith, G. Defective presentation to class I-restricted cytotoxic T-lymphocytes in vaccinia-infected cells is overcome by enhanced degradation of antigen. J. Exp. Med. 1988, 168, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.T.; Reiss, Y.; Fried, V.A.; Hershko, A.; Yoon, J.K.; Gonda, D.K.; Sangan, P.; Copeland, N.G.; Jenkins, N.A.; Varshavsky, A. The mouse and human genes encoding the recognition component of the N-end rule pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 7898–7903. [Google Scholar] [CrossRef] [PubMed]
- Rock, K.L.; Gramm, C.; Rothstein, L.; Clark, K.; Stein, R.; Dick, L.; Hwang, D.; Goldberg, A.L. Inhibitors of the proteasome block the degradation of most cell proteins and the generation of peptides presented on MHC class I molecules. Cell 1994, 78, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Lévy, F.; Burlet-Schiltz, O.; Brasseur, F.; Probst-Kepper, M.; Peitrequin, A.-L.; Monsarrat, B.; van Velthoven, R.; Cerottini, J.-C.; Boon, T.; et al. Processing of some antigens by the standard proteasome but not by the immunoproteasome results in poor presentation by dendritic cells. Immunity 2000, 12, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, S.; Zhang, Y.; Boon, T.; van der Bruggen, P. A MAGE-1 antigenic peptide recognized by human cytolytic T-lymphocytes on HLA-A2 tumor cells. Cancer Immunol. Immunother. 2005, 54, 1214–1220. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, N.; Stroobant, V.; Chapiro, J.; Ooms, A.; Degiovanni, G.; Morel, S.; van der Bruggen, P.; Boon, T.; Van den Eynde, B. An antigenic peptide produced by peptide splicing in the proteasome. Science 2004, 304, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Cerundolo, V.; Benham, A.; Braud, V.; Mukherjee, S.; Gould, K.; Macino, B.; Neefjes, J.; Townsend, A. The proteasome-specific inhibitor lactacystin blocks presentation of cytotoxic T-lymphocyte epitopes in human and murine cells. Eur. J. Immunol. 1997, 27, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.V.; France, J.; Song, R.; Farah, J.M.; Chatterjee, S.; Iqbal, M.; Siman, R. Novel dipeptide aldehydes are proteasome inhibitors and block the MHC-I antigen-processing pathway. J. Immunol. 1995, 155, 1767–1775. [Google Scholar] [PubMed]
- Yang, B.; Hahn, Y.S.; Hahn, C.S.; Braciale, T.J. The requirement for proteasome activity class I major histocompatibility complex antigen presentation is dictated by the length of preprocessed antigen. J. Exp. Med. 1996, 183, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Niedermann, G.; Butz, S.; Ihlenfeldt, H.G.; Grimm, R.; Lucchiari, M.; Hoschutzky, H.; Jung, G.; Maier, B.; Eichmann, K. Contribution of proteasome-mediated proteolysis to the hierarchy of epitopes presented by major histocompatibility complex class I molecules. Immunity 1995, 2, 289–299. [Google Scholar] [CrossRef]
- Vinitsky, A.; Anton, L.C.; Snyder, H.L.; Orlowski, M.; Bennink, J.R.; Yewdell, J.W. The generation of MHC class I-associated peptides is only partially inhibited by proteasome inhibitors-involvement of nonproteasomal cytosolic proteases in antigen processing? J. Immunol. 1997, 159, 554–564. [Google Scholar]
- Craiu, A.; Akopian, T.; Goldberg, A.; Rock, K.L. Two distinct proteolytic processes in the generation of a major histocompatibility complex class I-presented peptide. Proc. Natl. Acad. Sci. USA 1997, 94, 10850–10855. [Google Scholar] [CrossRef] [PubMed]
- Milner, E.; Gutter-Kapon, L.; Bassani-Strenberg, M.; Barnea, E.; Beer, I.; Admon, A. The effect of proteasome inhibition on the generation of the human leukocyte antigen (HLA) peptidome. Mol. Cell. Proteomics 2013, 12, 1853–1864. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, S.T.; Carew, J.S.; Dunner, K., Jr.; Boise, L.H.; Chiao, P.J.; Huang, P.; Abbruzzese, J.L.; McConkey, D.J. Bortezomib inhibits PKR-like endoplasmic reticulum (ER) kinase and induces apoptosis via ER stress in human pancreatic cancer cells. Cancer. Res. 2005, 65, 11510–11519. [Google Scholar] [CrossRef] [PubMed]
- Chapiro, J.; Claverol, S.; Piette, F.; Ma, W.; Stroobant, V.; Guillaume, B.; Gairin, J.-E.; Morel, S.; Burlet-Schiltz, O.; Monsarrat, B.; et al. Destructive cleavage of antigenic peptides either by the immunoproteasome or by the standard proteasome results in differential antigen presentation. J. Immunol. 2006, 176, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, B.; Chapiro, J.; Stroobant, V.; Colau, D.; van Holle, B.; Parvizi, G.; Bousquet-Dubouch, M.P.; Theate, I.; Parmentier, N.; Van den Eynde, B.J. Two abundant proteasome subtypes that uniquely process some antigens presented by HLA class I molecules. Proc. Natl. Acad. Sci. USA 2010, 107, 18599–18604. [Google Scholar] [CrossRef] [PubMed]
- Gelman, J.S.; Sironi, J.; Berezniuk, I.; Dasgupta, S.; Castro, L.M.; Gozzo, F.C.; Ferro, E.S.; Fricker, L.D. Alterations of the intracellular peptidome in response to the proteasome inhibitor bortezomib. PLoS One 2013, 8, e53263. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.Y.; Cascio, P.; Lemerise, K.; Goldberg, A.L.; Rock, K. Distinct proteolytic processes generate the C and N termini of MHC class I-binding peptides. J. Immunol. 1999, 163, 5851–5859. [Google Scholar] [PubMed]
- Reits, E.; Neijssen, J.; Herberts, C.; Benckhuijsen, W.; Janssen, L.; Drijfhout, J.W.; Neefjes, J. A major role for TPPII in trimming proteasomal degradation products for MHC class I antigen presentation. Immunity 2004, 20, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Saric, T.; Chang, S.C.; Hattori, A.; York, I.A.; Markant, S.; Rock, K.L.; Tsujimoto, M.; Goldberg, A.L. An IFN-γ-induced aminopeptidase in the ER, ERAP1, trims precursors to MHC class I-presented peptides. Nat. Immunol. 2002, 3, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Kanaseki, T.; Blanchard, N.; Hammer, G.E.; Gonzalez, F.; Shastri, N. ERAAP synergizes with MHC class I molecules to make the final cut in the antigenic peptide precursors in the endoplasmic reticulum. Immunity 2006, 25, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, B.; Stroobant, V.; Bousquet-Dubouch, M.P.; Colau, D.; Chapiro, J.; Parmentier, N.; Dalet, A.; Van den Eynde, B.J. Analysis of the processing of seven human tumor antigens by intermediate proteasomes. J. Immunol. 2012, 189, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Luckey, C.J.; King, G.M.; Marto, J.A.; Venketeswaran, S.; Maier, B.F.; Crotzer, V.L.; Colella, T.A.; Shabanowitz, J.; Hunt, D.F.; Engelhard, V.H. Proteasomes can either generate or destroy MHC class I epitopes: Evidence for nonproteasomal epitope generation in the cytosol. J. Immunol. 1998, 161, 112–121. [Google Scholar] [PubMed]
- Seifert, U.; Maranon, C.; Shmueli, A.; Desoutter, J.F.; Wesoloski, L.; Janek, K.; Henklein, P.; Diescher, S.; Andrieu, M.; de la Salle, H.; et al. An essential role for tripeptidyl peptidase in the generation of an MHC class I epitope. Nat. Immunol. 2003, 4, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Guil, S.; Rodriguez-Castro, M.; Aguilar, F.; Villasevil, E.M.; Anton, L.C.; del Val, M. Need for tripeptidyl-peptidase II in major histocompatibility complex class I viral antigen processing when proteasomes are detrimental. J. Biol. Chem. 2006, 281, 39925–39934. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, J.; Adamopoulou, E.; Beck, O.; Rauser, G.; Lurati, S.; Tenzer, S.; Einsele, H.; Rammensee, H.G.; Schild, H.; Topp, M.S. Processing of two latent membrane protein 1 MHC class I epitopes requires tripeptidyl peptidase II involvement. J. Immunol. 2009, 183, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, N.; Stroobant, V.; Colau, D.; de Diesbach, P.; Morel, S.; Chapiro, J.; Van Endert, P.; Van den Eynde, B.J. Production of an antigenic peptide by insulin-degrading enzyme. Nat. Immunol. 2010, 11, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Kessler, J.H.; Khan, S.; Seifert, U.; le Gall, S.; Chow, K.M.; Paschen, A.; Bres-Vloemans, S.A.; de Ru, A.; van Montfoort, N.; Franken, K.L.; et al. Antigen processing by nardilysin and thimet oligopeptidase generates cytotoxic T-cell epitopes. Nat. Immunol. 2011, 12, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kessler, B.; Hong, X.; Petrovic, J.; Borodovsky, A.; Dantuma, N.P.; Bogyo, M.; Overkleeft, H.S.; Ploegh, H.; Glas, R. Pathways accessory to proteasomal proteolysis are less efficient in major histocompatibility complex class I antigen production. J. Biol. Chem. 2003, 278, 10013–10021. [Google Scholar] [CrossRef] [PubMed]
- Van Endert, P. Role of tripeptidyl peptidase II in MHC class I antigen processing—The end of controversies? Eur. J. Immunol. 2008; 38, 609–613. [Google Scholar]
- Culina, S.; Mauvais, F.X.; Hsu, H.T.; Burgevin, A.; Guenette, S.; Moser, A.; van Endert, P. No major role for insulin-degrading enzyme in antigen presentation by MHC molecules. PLoS One 2014, 9, e88365. [Google Scholar] [CrossRef] [PubMed]
- El Hage, F.; Stroobant, V.; Vergnon, I.; Baurain, J.F.; Echchakir, H.; Lazar, V.; Chouaib, S.; Coulie, P.G.; Mami-Chouaib, F. Preprocalcitonin signal peptide generates a cytotoxic T-lymphocyte-defined tumor epitope processed by a proteasome-independent pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 10119–10124. [Google Scholar]
- Wölfel, C.; Drexler, I.; van Pel, A.; Thres, T.; Leister, N.; Herr, W.; Sutter, G.; Huber, C.; Wölfel, T. Transporter (TAP)- and proteasome-independent presentation of a melanoma-associated tyrosinase epitope. Int. J. Cancer 2000, 88, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, R.M.; Fiegl, D.; Rufer, E.; Karger, A.; Bettin, B.; Knittler, M.R. Post-endoplasmic reticulum rescue of unstable MHC class I requires proprotein convertase PC7. J. Immunol. 2010, 184, 2985–2998. [Google Scholar] [CrossRef] [PubMed]
- Gil-Torregrosa, B.C.; Raul Castano, A.; del Val, M. Major histocompatibility complex class I viral antigen processing in the secretory pathway defined by the trans-Golgi network protease furin. J. Exp. Med. 1998, 188, 1105–1116. [Google Scholar] [PubMed]
- Tey, S.K.; Khanna, R. Autophagy mediates transporter associated with antigen processing-independent presentation of viral epitopes through MHC class I pathway. Blood 2012, 120, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- English, L.; Chemali, M.; Duron, J.; Rondeau, C.; Laplante, A.; Gingras, D.; Alexander, D.; Leib, D.; Norbury, C.; Lippe, R.; et al. Autophagy enhances the presentation of endogenous viral antigens on MHC class I molecules during HSV-1 infection. Nat. Immunol. 2009, 10, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Demachi-Okamura, A.; Torikai, H.; Akatsuka, Y.; Miyoshi, H.; Yoshimori, T.; Kuzushima, K. Autophagy creates a CTL epitope that mimics tumor-associated antigens. PLoS One 2012, 7, e47126. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Yewdell, J.W.; Yang, J.C. Immune recognition of a human renal cancer antigen through post-translational protein splicing. Nature 2004, 427, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Dalet, A.; Stroobant, V.; Vigneron, N.; Van den Eynde, B.J. Differences in the production of spliced antigenic peptides by the standard proteasome and the immunoproteasome. Eur. J. Immunol. 2011, 41, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Michaux, A.; Larrieu, P.; Stroobant, V.; Fonteneau, J.F.; Jotereau, F.; Van den Eynde, B.J.; Moreau-Aubry, A.; Vigneron, N. A spliced antigenic peptide comprising a single spliced amino acid is produced in the proteasome by reverse splicing of a longer peptide fragment followed by trimming. J. Immunol. 2014, 192, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Wölfel, T.; van Pel, A.; Brichard, V.; Schneider, J.; Seliger, B.; Meyer zum Büschenfelde, K.-H.; Boon, T. Two tyrosinase nonapeptides recognized on HLA-A2 melanomas by autologous cytolytic T-lymphocytes. Eur. J. Immunol. 1994, 24, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Dalet, A.; Robbins, P.F.; Stroobant, V.; Vigneron, N.; Li, Y.F.; El-Gamil, M.; Hanada, K.; Yang, J.C.; Rosenberg, S.A.; Van den Eynde, B.J. An antigenic peptide produced by reverse splicing and double asparagine deamidation. Proc. Natl. Acad. Sci. USA 2011, 108, E323–E331. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Eliyahu, S.; Sakaguchi, K.; Robbins, P.F.; Rivoltini, L.; Yannelli, J.R.; Appella, E.; Rosenberg, S.A. Identification of the immunodominant peptides of the MART-1 human melanoma antigen recognized by the majority of HLA-A2-restricted tumor infiltrating lymphocytes. J. Exp. Med. 1994, 180, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Schultz, E.S.; Chapiro, J.; Lurquin, C.; Claverol, S.; Burlet-Schiltz, O.; Warnier, G.; Russo, V.; Morel, S.; Levy, F.; Boon, T.; et al. The production of a new MAGE-3 peptide presented to cytolytic T-lymphocytes by HLA-B40 requires the immunoproteasome. J. Exp. Med. 2002, 195, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Van der Bruggen, P.; Bastin, J.; Gajewski, T.; Coulie, P.G.; Boël, P.; de Smet, C.; Traversari, C.; Townsend, A.; Boon, T. A peptide encoded by human gene MAGE-3 and presented by HLA-A2 induces cytolytic T-lymphocytes that recognize tumor cells expressing MAGE-3. Eur. J. Immunol. 1994, 24, 3038–3043. [Google Scholar]
- Huang, L.-Q.; Brasseur, F.; Serrano, A.; de Plaen, E.; van der Bruggen, P.; Boon, T.; van Pel, A. Cytolytic T-lymphocytes recognize an antigen encoded by MAGE-A10 on a human melanoma. J. Immunol. 1999, 162, 6849–6854. [Google Scholar] [PubMed]
- Ma, W.; Germeau, C.; Vigneron, N.; Maernoudt, A.-S.; Morel, S.; Boon, T.; Coulie, P.G.; Van den Eynde, B. Two new tumor-specific antigenic peptides encoded by gene MAGE-C2 and presented to cytolytic T-lymphocytes by HLA-A2. Int. J. Cancer 2004, 109, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Vigneron, N.; Chapiro, J.; Stroobant, V.; Germeau, C.; Boon, T.; Coulie, P.G.; van den Eynde, B.J. A MAGE-C2 antigenic peptide processed by the immunoproteasome is recognized by cytolytic T-cells isolated from a melanoma patient after successful immunotherapy. Int. J. Cancer 2011, 129, 2427–2434. [Google Scholar] [CrossRef] [PubMed]
- Corbière, V.; Chapiro, J.; Stroobant, V.; Ma, W.; Lurquin, C.; Lethe, B.; Van Baren, N.; van den Eynde, B.J.; Boon, T.; Coulie, P.G. Antigen spreading contributes to MAGE vaccination-induced regression of melanoma metastases. Cancer. Res. 2011, 71, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Robbins, P.F.; Lu, Y.C.; El-Gamil, M.; Li, Y.F.; Gross, C.; Gartner, J.; Lin, J.C.; Teer, J.K.; Cliften, P.; Tycksen, E.; et al. Mining exomic sequencing data to identify mutated antigens recognized by adoptively transferred tumor-reactive T-cells. Nat. Med. 2013, 19, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Warren, E.H.; Vigneron, N.J.; Gavin, M.A.; Coulie, P.G.; Stroobant, V.; Dalet, A.; Tykodi, S.S.; Xuereb, S.M.; Mito, J.K.; Riddell, S.R.; et al. An antigen produced by splicing of noncontiguous peptides in the reverse order. Science 2006, 313, 1444–1447. [Google Scholar] [CrossRef] [PubMed]
- Löwe, J.; Stock, D.; Jap, B.; Zwickl, P.; Baumeister, W.; Huber, R. Crystal structure of the 20S proteasome from the archaeon T. acidophilum at 3.4 Å resolution. Science 1995, 268, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Groll, M.; Ditzel, L.; Löwe, J.; Stock, D.; Bochtler, M.; Bartunik, H.D.; Huber, R. Structure of 20S proteasome from yeast at 2.4 Å resolution. Nature 1997, 386, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Seemuller, E.; Lupas, A.; Stock, D.; Lowe, J.; Huber, R.; Baumeister, W. Proteasome from Thermoplasma acidophilum: A threonine protease. Science 1995, 268, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Heinemeyer, W.; Fischer, M.; Krimmer, T.; Stachon, U.; Wolf, D.H. The active sites of the eukaryotic 20 S proteasome and their involvement in subunit precursor processing. J. Biol. Chem. 1997, 272, 25200–25209. [Google Scholar] [CrossRef] [PubMed]
- Groll, M.; Huber, R. Substrate access and processing by the 20S proteasome core particle. Int. J. Biochem. Cell Biol. 2003, 35, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Dalet, A.; Vigneron, N.; Stroobant, V.; Hanada, K.; Van den Eynde, B.J. Splicing of distant peptide fragments occurs in the proteasome by transpeptidation and produces the spliced antigenic peptide derived from fibroblast growth factor-5. J. Immunol. 2010, 184, 3016–3024. [Google Scholar] [CrossRef] [PubMed]
- Mishto, M.; Goede, A.; Taube, K.T.; Keller, C.; Janek, K.; Henklein, P.; Niewienda, A.; Kloss, A.; Gohlke, S.; Dahlmann, B.; et al. Driving forces of proteasome-catalyzed peptide splicing in yeast and humans. Mol. Cell. Proteomics 2012, 11, 1008–1023. [Google Scholar] [CrossRef] [PubMed]
- Liepe, J.; Mishto, M.; Textoris-Taube, K.; Janek, K.; Keller, C.; Henklein, P.; Kloetzel, P.M.; Zaikin, A. The 20S proteasome splicing activity discovered by SpliceMet. PLoS Comput. Biol. 2010, 6, e1000830. [Google Scholar] [CrossRef] [PubMed]
- Van der Bruggen, P.; Zhang, Y.; Chaux, P.; Stroobant, V.; Panichelli, C.; Schultz, E.S.; Chapiro, J.; Van den Eynde, B.J.; Brasseur, F.; Boon, T. Tumor-specific shared antigenic peptides recognized by human T-cells. Immunol. Rev. 2002, 188, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, W.; Walz, J.; Zühl, F.; Seemüller, E. The proteasome: Paradigm of a self-compartmentalizing protease. Cell 1998, 92, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Unno, M.; Mizushima, T.; Morimoto, Y.; Tomisugi, Y.; Tanaka, K.; Yasuoka, N.; Tsukihara, T. The structure of the mammalian 20S proteasome at 2.75 Å resolution. Structure 2002, 10, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J. Degradation of oxidized proteins by the 20S proteasome. Biochimie 2001, 83, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Pickering, A.M.; Koop, A.L.; Teoh, C.Y.; Ermak, G.; Grune, T.; Davies, K.J. The immunoproteasome, the 20S proteasome and the PA28αβ proteasome regulator are oxidative-stress-adaptive proteolytic complexes. Biochem. J. 2010, 432, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.; Fuertes, G.; Murray, R.Z.; Bose, S.; Knecht, E.; Rechsteiner, M.C.; Hendil, K.B.; Tanaka, K.; Dyson, J.; Rivett, J. Subcellular localization of proteasomes and their regulatory complexes in mammalian cells. Biochem. J. 2000, 346, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M.; Franke, W.W.; Kleinschmidt, J.A. Distinct 19S and 20S subcomplexes of the 26S proteasome and their distribution in the nucleus and the cytoplasm. J. Biol. Chem. 1994, 269, 7709–7718. [Google Scholar] [PubMed]
- Baugh, J.M.; Viktorova, E.G.; Pilipenko, E.V. Proteasomes can degrade a significant proportion of cellular proteins independent of ubiquitination. J. Mol. Biol. 2009, 386, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Whittier, J.E.; Xiong, Y.; Rechsteiner, M.C.; Squier, T.C. Hsp90 enhances degradation of oxidized calmodulin by the 20S proteasome. J. Biol. Chem. 2004, 279, 46135–46142. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, M. Selective activation of the 20S proteasome (multicatalytic proteinase complex) by histone H3. Biochemistry 2001, 40, 15318–15326. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Kaganovich, D.; Goldberg, A.L. Binding of hydrophobic peptides to several non-catalytic sites promotes peptide hydrolysis by all active sites of 20S proteasomes. Evidence for peptide-induced channel opening in the α-rings. J. Biol. Chem. 2002, 277, 22260–22270. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yen, J.; Kaiser, P.; Huang, L. Regulation of the 26S proteasome complex during oxidative stress. Sci. Signal. 2010. [Google Scholar] [CrossRef]
- Grune, T.; Catalgol, B.; Licht, A.; Ermak, G.; Pickering, A.M.; Ngo, J.K.; Davies, K.J. Hsp70 mediates dissociation and reassociation of the 26S proteasome during adaptation to oxidative stress. Free Radic. Biol. Med. 2011, 51, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Bajorek, M.; Finley, D.; Glickman, M.H. Proteasome disassembly and downregulation is correlated with viability during stationary phase. Curr. Biol. 2003, 13, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Aiken, C.T.; Kaake, R.M.; Wang, X.; Huang, L. Oxidative stress-mediated regulation of proteasome complexes. Mol. Cell. Proteomics 2011. [Google Scholar] [CrossRef]
- Li, J.; Powell, S.R.; Wang, X. Enhancement of proteasome function by PA28α; overexpression protects against oxidative stress. FASEB J. 2011, 25, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Griffin, T.A.; Nandi, D.; Cruz, M.; Fehling, H.J.; van Kaer, L.; Monaco, J.J.; Colbert, R.A. Immunoproteasome assembly: Cooperative incorporation of interferon γ (IFN-γ)-inducible subunits. J. Exp. Med. 1998, 187, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Enenkel, C.; Lehmann, H.; Kipper, J.; Guckel, R.; Hilt, W.; Wolf, D.H. PRE3, highly homologous to the human major histocompatibility complex-linked LMP2 (RING12) gene, codes for a yeast proteasome subunit necessary for the peptidylglutamyl-peptide hydrolyzing activity. FEBS Lett. 1994, 341, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Dick, T.P.; Nussbaum, A.K.; Deeg, M.; Heinemeyer, W.; Groll, M.; Schirle, M.; Keilholz, W.; Stevanovic, S.; Wolf, D.H.; Huber, R.; et al. Contribution of proteasomal β-subunits to the cleavage of peptide substrates analyzed with yeast mutants. J. Biol. Chem. 1998, 273, 25637–25646. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, A.K.; Dick, T.P.; Keilholz, W.; Schirle, M.; Stevanovic, S.; Dietz, K.; Heinemeyer, W.; Groll, M.; Wolf, D.H.; Huber, R.; et al. Cleavage motifs of the yeast 20S proteasome β subunits deduced from digests of enolase 1. Proc. Natl. Acad. Sci. USA 1998, 95, 12504–12509. [Google Scholar] [CrossRef] [PubMed]
- Toes, R.E.M.; Nussbaum, A.K.; Degermann, S.; Schirle, M.; Emmerich, N.P.N.; Kraft, M.; Laplace, C.; Zwinderman, A.; Dick, T.P.; Müller, J.; et al. Discrete cleavage motifs of constitutive and immunoproteasomes revealed by quantitative analysis of cleavage products. J. Exp. Med. 2001, 194, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gaczynska, M.; Rock, K.L.; Goldberg, A.L. γ-interferon and expression of MHC genes regulate peptide hydrolysis by proteasomes. Nature 1993, 365, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Gaczynska, M.; Rock, K.L.; Spies, T.; Goldberg, A.L. Peptidase activities of proteasomes are differentially regulated by the major histocompatibility complex-encoded genes for LMP2 and LMP7. Proc. Natl. Acad. Sci. USA 1994, 91, 9213–9217. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, J.; Brown, M.; Finley, D.; Monaco, J. MHC-linked LMP gene products specifically alter peptidase activities of the proteasome. Nature 1993, 365, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Boes, B.; Hengel, H.; Ruppert, T.; Multhaup, G.; Koszinowski, U.H.; Kloetzel, P.M. Interferon γ stimulation modulates the proteolytic activity and cleavage site preference of 20S mouse proteasomes. J. Exp. Med. 1994, 179, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Fehling, H.J.; Swat, W.; Laplace, C.; Kühn, R.; Rajewsky, K.; Müller, U.; von Boehmer, H. MHC class I expression in mice lacking the proteasome subunit LMP-7. Science 1994, 265, 1234–1237. [Google Scholar] [CrossRef] [PubMed]
- Van Kaer, L.; Ashton-Rickardt, P.G.; Eichelberger, M.; Gaczynska, M.; Nagashima, K.; Rock, K.L.; Goldberg, A.L.; Doherty, P.C.; Tonegawa, S. Altered peptidase and viral-specific T-cell response in LMP2 mutant mice. Immunity 1994, 1, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Kincaid, E.Z.; Che, J.W.; York, I.; Escobar, H.; Reyes-Vargas, E.; Delgado, J.C.; Welsh, R.M.; Karow, M.L.; Murphy, A.J.; Valenzuela, D.M.; et al. Mice completely lacking immunoproteasomes show major changes in antigen presentation. Nat. Immunol. 2012, 13, 129–135. [Google Scholar] [CrossRef]
- De Verteuil, D.; Muratore-Schroeder, T.L.; Granados, D.P.; Fortier, M.H.; Hardy, M.P.; Bramoulle, A.; Caron, E.; Vincent, K.; Mader, S.; Lemieux, S.; et al. Deletion of immunoproteasome subunits imprints on the transcriptome and has a broad impact on peptides presented by major histocompatibility complex I molecules. Mol. Cell. Proteomics 2010, 9, 2034–2047. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, N.; Van den Eynde, B.J. Proteasome subtypes and the processing of tumor antigens: Increasing antigenic diversity. Curr. Opin. Immunol. 2012, 24, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Lauer, C.; Moebius, J.; Weber, R.; Przybylski, M.; Kisselev, A.F.; Tsu, C.; Groettrup, M. Why the structure but not the activity of the immunoproteasome subunit low molecular mass polypeptide 2 rescues antigen presentation. J. Immunol. 2012, 189, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Norbury, C.C.; Cho, Y.; Yewdell, J.W.; Bennink, J.R. Immunoproteasomes shape immunodominance hierarchies of antiviral CD8+ T-cells at the levels of T-cell repertoire and presentation of viral antigens. J. Exp. Med. 2001, 193, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Chapatte, L.; Ayyoub, M.; Morel, S.; Peitrequin, A.L.; Levy, N.; Servis, C.; Van den Eynde, B.J.; Valmori, D.; Levy, F. Processing of tumor-associated antigen by the proteasomes of dendritic cells controls in vivo T-cell responses. Cancer Res. 2006, 66, 5461–5468. [Google Scholar] [CrossRef]
- Dannull, J.; Haley, N.R.; Archer, G.; Nair, S.; Boczkowski, D.; Harper, M.; de Rosa, N.; Pickett, N.; Mosca, P.J.; Burchette, J.; et al. Melanoma immunotherapy using mature DCs expressing the constitutive proteasome. J. Clin. Invest. 2013, 123, 3135–3145. [Google Scholar] [CrossRef] [PubMed]
- Dannull, J.; Lesher, D.T.; Holzknecht, R.; Qi, W.; Hanna, G.; Seigler, H.; Tyler, D.S.; Pruitt, S.K. Immunoproteasome down-modulation enhances the ability of dendritic cells to stimulate antitumor immunity. Blood 2007, 110, 4341–4350. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Moebius, J.; Elenich, L.; Groettrup, M.; Monaco, J.J. An altered T-cell repertoire in MECL-1-deficient mice. J. Immunol. 2006, 176, 6665–6672. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Youhnovski, N.; van den Broek, M.; Przybylski, M.; Groettrup, M. Immunoproteasomes down-regulate presentation of a subdominant T-cell epitope from lymphocytic choriomeningitis virus. J. Immunol. 2004, 173, 3925–3934. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.C.; Sanders, M.T.; Monaco, J.J.; Doherty, P.C.; Turner, S.J.; Chen, W. Immunoproteasome subunit deficiencies impact differentially on two immunodominant influenza virus-specific CD8+ T-cell responses. J. Immunol. 2006, 177, 7680–7688. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, S.; Sims, S.; O’Hara, G.; Silk, J.; Gileadi, U.; Cerundolo, V.; Klenerman, P. A dominant role for the immunoproteasome in CD8+ T-cell responses to murine cytomegalovirus. PLoS One 2011, 6, e14646. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, A.; Trilling, M.; Wagner, M.; Wilborn, M.; Bubic, I.; Jonjic, S.; Koszinowski, U.; Hengel, H. A cytomegaloviral protein reveals a dual role for STAT2 in IFN-γ signaling and antiviral responses. J. Exp. Med. 2005, 201, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Dahlmann, B.; Ruppert, T.; Kuehn, L.; Merforth, S.; Kloetzel, P.-M. Different proteasome subtypes in a single tissue exhibit different enzymatic properties. J. Mol. Biol. 2000, 303, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Groettrup, M.; Standera, S.; Stohwasser, R.; Kloetzel, P.M. The subunits MECL-1 and LMP2 are mutually required for incorporation into the 20S proteasome. Proc. Natl. Acad. Sci. USA 1997, 94, 8970–8975. [Google Scholar] [CrossRef] [PubMed]
- De, M.; Jayarapu, K.; Elenich, L.; Monaco, J.J.; Colbert, R.A.; Griffin, T.A. β2 subunit propeptides influence cooperative proteasome assembly. J. Biol. Chem. 2003, 278, 6153–6159. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, D.J.; Griffin, T.A.; Colbert, R.A. Novel propeptide function in 20S proteasome assembly influences β subunit composition. J. Biol. Chem. 2000, 275, 24156–24162. [Google Scholar] [CrossRef] [PubMed]
- Groll, M.; Heinemeyer, W.; Jäger, S.; Ullrich, T.; Bochtler, M.; Wolf, D.H.; Huber, R. The catalytic sites of 20S proteasomes and their role in subunit maturation: A mutational and crystallographic study. Proc. Natl. Acad. Sci. USA 1999, 96, 10976–10983. [Google Scholar] [CrossRef] [PubMed]
- Klare, N.; Seeger, M.; Janek, K.; Jungblut, P.R.; Dahlmann, B. Intermediate-type 20S proteasomes in HeLa cells: “Asymmetric” subunit composition, diversity and adaptation. J. Mol. Biol. 2007, 373, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Sasaki, K.; Kishimoto, T.; Niwa, S.; Hayashi, H.; Takahama, Y.; Tanaka, K. Regulation of CD8+ T-cell development by thymus-specific proteasomes. Science 2007, 316, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Tomaru, U.; Ishizu, A.; Murata, S.; Miyatake, Y.; Suzuki, S.; Takahashi, S.; Kazamaki, T.; Ohara, J.; Baba, T.; Iwasaki, S.; et al. Exclusive expression of proteasome subunit β5t in the human thymic cortex. Blood 2009, 113, 5186–5191. [Google Scholar] [CrossRef] [PubMed]
- Nitta, T.; Murata, S.; Sasaki, K.; Fujii, H.; Ripen, A.M.; Ishimaru, N.; Koyasu, S.; Tanaka, K.; Takahama, Y. Thymoproteasome shapes immunocompetent repertoire of CD8+ T-cells. Immunity 2010, 32, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Muller, C.A.; Bockmann, R.A.; Uchanska-Ziegler, B. Low-affinity peptides and T-cell selection. Trends Immunol. 2009, 30, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Jameson, S.C.; Hogquist, K.A. Thymoproteasome subunit-β5t generates peptide-MHC complexes specialized for positive selection. Proc. Natl. Acad. Sci. USA 2013, 110, 6979–6984. [Google Scholar] [CrossRef] [PubMed]
- Groettrup, M.; Kirk, C.J.; Basler, M. Proteasomes in immune cells: more than peptide producers? Nat. Rev. Immunol. 2010, 10, 73–78. [Google Scholar] [CrossRef]
- Gotter, J.; Brors, B.; Hergenhahn, M.; Kyewski, B. Medullary epithelial cells of the human thymus express a highly diverse selection of tissue-specific genes colocalized in chromosomal clusters. J. Exp. Med. 2004, 199, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Huijbers, I.J.; Soudja, S.M.; Uyttenhove, C.; Buferne, M.; Inderberg-Suso, E.M.; Colau, D.; Pilotte, L.; Chomez, P.; Brasseur, F.; Schmitt-Verhulst, A.M.; et al. Minimal tolerance to a tumor antigen encoded by a cancer-germline gene. J. Immunol. 2012, 188, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Huber, E.M.; Basler, M.; Schwab, R.; Heinemeyer, W.; Kirk, C.J.; Groettrup, M.; Groll, M. Immuno- and constitutive proteasome crystal structures reveal differences in substrate and inhibitor specificity. Cell 2012, 148, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Sijts, A.J.A.M.; Ruppert, T.; Rehermann, B.; Schmidt, M.; Koszinowski, U.; Kloetzel, P.-M. Efficient generation of a hepatitis B virus cytotoxic T-lymphocyte epitope requires the structural features of immunoproteasomes. J. Exp. Med. 2000, 191, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Seifert, U.; Bialy, L.P.; Ebstein, F.; Bech-Otschir, D.; Voigt, A.; Schroter, F.; Prozorovski, T.; Lange, N.; Steffen, J.; Rieger, M.; et al. Immunoproteasomes preserve protein homeostasis upon interferon-induced oxidative stress. Cell 2010, 142, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Nathan, J.A.; Spinnenhirn, V.; Schmidtke, G.; Basler, M.; Groettrup, M.; Goldberg, A.L. Immuno- and constitutive proteasomes do not differ in their abilities to degrade ubiquitinated proteins. Cell 2013, 152, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Faustman, D. NOD mice are defective in proteasome production and activation of NF-κB. Mol. Cell. Biol. 1999, 19, 8646–8659. [Google Scholar] [PubMed]
- Kessler, B.M.; Lennon-Dumenil, A.M.; Shinohara, M.L.; Lipes, M.A.; Ploegh, H.L. LMP2 expression and proteasome activity in NOD mice. Nat. Med. 2000, 6, 1064. [Google Scholar] [PubMed]
- Runnels, H.A.; Watkins, W.A.; Monaco, J.J. LMP2 expression and proteasome activity in NOD mice. Nat. Med. 2000, 6, 1064–1065. [Google Scholar] [PubMed]
- Hensley, S.E.; Zanker, D.; Dolan, B.P.; David, A.; Hickman, H.D.; Embry, A.C.; Skon, C.N.; Grebe, K.M.; Griffin, T.A.; Chen, W.; et al. Unexpected role for the immunoproteasome subunit LMP2 in antiviral humoral and innate immune responses. J. Immunol. 2010, 184, 4115–4122. [Google Scholar] [CrossRef] [PubMed]
- Moebius, J.; van den Broek, M.; Groettrup, M.; Basler, M. Immunoproteasomes are essential for survival and expansion of T-cells in virus-infected mice. Eur. J. Immunol. 2010, 40, 3439–3449. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Kapphahn, R.J.; Terluk, M.R.; Heuss, N.D.; Yuan, C.; Gregerson, D.S.; Ferrington, D.A. Immunoproteasome deficiency modifies the alternative pathway of NF-κB signaling. PLoS One 2013, 8, e56187. [Google Scholar] [CrossRef] [PubMed]
- Muchamuel, T.; Basler, M.; Aujay, M.A.; Suzuki, E.; Kalim, K.W.; Lauer, C.; Sylvain, C.; Ring, E.R.; Shields, J.; Jiang, J.; et al. A selective inhibitor of the immunoproteasome subunit LMP7 blocks cytokine production and attenuates progression of experimental arthritis. Nat. Med. 2009, 15, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M. Mechanisms underlying ubiquitination. Annu. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef] [PubMed]
- Forster, F.; Unverdorben, P.; Sledz, P.; Baumeister, W. Unveiling the long-held secrets of the 26S proteasome. Structure 2013, 21, 1551–1562. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Cohen, R.E. A cryptic protease couples deubiquitination and degradation by the proteasome. Nature 2002, 419, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Aravind, L.; Oania, R.; McDonald, W.H.; Yates, J.R., 3rd; Koonin, E.V.; Deshaies, R.J. Role of Rpn11 metalloprotease in deubiquitination and degradation by the 26S proteasome. Science 2002, 298, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Pathare, G.R.; Nagy, I.; Bohn, S.; Unverdorben, P.; Hubert, A.; Korner, R.; Nickell, S.; Lasker, K.; Sali, A.; Tamura, T.; et al. The proteasomal subunit Rpn6 is a molecular clamp holding the core and regulatory subcomplexes together. Proc. Natl. Acad. Sci. USA 2012, 109, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, N.; Murakami, Y.; Minami, Y.; Shimbara, N.; Hendil, K.B.; Tanaka, K. Hybrid proteasomes. Induction by interferon-γ and contribution to ATP-dependent proteolysis. J. Biol. Chem. 2000, 275, 14336–14345. [Google Scholar] [CrossRef] [PubMed]
- Ustrell, V.; Hoffman, L.; Pratt, G.; Rechsteiner, M. PA200, a nuclear proteasome activator involved in DNA repair. EMBO J. 2002, 21, 3516–3525. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.P.; Slaughter, C.A.; DeMartino, G.N. Identification, purification, and characterization of a protein activator (PA28) of the 20 S proteasome (macropain). J. Biol. Chem. 1992, 267, 10515–10523. [Google Scholar] [PubMed]
- Dubiel, W.; Pratt, G.; Ferrell, K.; Rechsteiner, M. Purification of an 11S regulator of the multicatalytic protease. J. Biol. Chem. 1992, 267, 22369–22377. [Google Scholar] [PubMed]
- Rechsteiner, M.; Realini, C.; Ustrell, V. The proteasome activator 11S REG (PA28) and class I antigen presentation. Biochem. J. 2000, 345, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Realini, C.; Dubiel, W.; Pratt, G.; Ferrell, K.; Rechsteiner, M. Molecular cloning and expression of a γ-interferon-inducible activator of the multicatalytic protease. J. Biol. Chem. 1994, 269, 20727–20732. [Google Scholar] [PubMed]
- Ahn, J.Y.; Tanahashi, N.; Akiyama, K.; Hisamatsu, H.; Noda, C.; Tanaka, K.; Chung, C.H.; Shibmara, N.; Willy, P.J.; Mott, J.D.; et al. Primary structures of two homologous subunits of PA28, a γ-interferon-inducible protein activator of the 20S proteasome. FEBS Lett. 1995, 366, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.W.; Slaughter, C.A.; DeMartino, G.N. PA28 activator protein forms regulatory caps on proteasome stacked rings. J. Mol. Biol. 1994, 236, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, J.R.; Johnston, S.C.; Whitby, F.G.; Realini, C.; Zhang, Z.; Rechsteiner, M.; Hill, C.P. Structure of the proteasome activator REGα (PA28α). Nature 1997, 390, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Krutchinsky, A.; Endicott, S.; Realini, C.; Rechsteiner, M.; Standing, K.G. Proteasome activator 11S REG or PA28: Recombinant REG α/REG β hetero-oligomers are heptamers. Biochemistry 1999, 38, 5651–5658. [Google Scholar] [CrossRef] [PubMed]
- Hendil, K.B.; Khan, S.; Tanaka, K. Simultaneous binding of PA28 and PA700 activators to 20S proteasomes. Biochem. J. 1998, 332, 749–754. [Google Scholar] [PubMed]
- Cascio, P.; Call, M.; Petre, B.M.; Walz, T.; Goldberg, A.L. Properties of the hybrid form of the 26S proteasome containing both 19S and PA28 complexes. EMBO J. 2002, 21, 2636–2645. [Google Scholar] [CrossRef] [PubMed]
- DeMartino, G.N.; Slaughter, C.A. The proteasome, a novel protease regulated by multiple mechanisms. J. Biol. Chem. 1999, 274, 22123–22126. [Google Scholar] [CrossRef] [PubMed]
- Whitby, F.G.; Masters, E.I.; Kramer, L.; Knowlton, J.R.; Yao, Y.; Wang, C.C.; Hill, C.P. Structural basis for the activation of 20S proteasomes by 11S regulators. Nature 2000, 408, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Kohler, A.; Cascio, P.; Leggett, D.S.; Woo, K.M.; Goldberg, A.L.; Finley, D. The axial channel of the proteasome core particle is gated by the Rpt2 ATPase and controls both substrate entry and product release. Mol. Cell 2001, 7, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Murata, S.; Shimbara, N.; Tanaka, N.; Chiba, T.; Tanaka, K.; Yui, K.; Udono, H. Two distinct pathways mediated by PA28 and hsp90 in major histocompatibility complex class I antigen processing. J. Exp. Med. 2002, 196, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Preckel, T.; Fung-Leung, W.-P.; Cai, Z.; Vitiello, A.; Salter-Cid, L.; Winqvist, O.; Wolfe, T.G.; von Herrath, M.; Angulo, A.; Ghazal, P.; et al. Impaired immunoproteasome assembly and immune responses in PA28−/− mice. Science 1999, 286, 2162–2165. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Udono, H.; Tanahashi, N.; Hamada, H.; Watanabe, K.; Adachi, K.; Yamano, T.; Yui, K.; Kobayashi, H.; Kasahara, M.; et al. Immunoproteasome assembly and antigen presentation in mice lacking both PA28α and PA28β. EMBO J. 2001, 20, 5898–5907. [Google Scholar] [CrossRef] [PubMed]
- Dick, T.P.; Ruppert, T.; Groettrup, M.; Kloetzel, P.M.; Kuehn, L.; Koszinowski, U.H.; Stevanovic, S.; Schild, H.; Rammensee, H.-G. Coordinated dual cleavages induced by the proteasome regulator PA28 lead to dominant MHC ligands. Cell 1996, 86, 253–262. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, N.; van Helden, M.J.; Textoris-Taube, K.; Chiba, T.; Topham, D.J.; Kloetzel, P.M.; Zaiss, D.M.; Sijts, A.J. PA28 and the proteasome immunosubunits play a central and independent role in the production of MHC class I-binding peptides in vivo. Eur. J. Immunol. 2011, 41, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Groettrup, M.; Soza, A.; Eggers, M.; Kuehn, L.; Dick, T.P.; Schild, H.; Rammensee, H.-G.; Koszinowski, U.H.; Kloetzel, P.M. A role for the proteasome regulator PA28α in antigen presentation. Nature 1996, 381, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; Eggers, M.; Soza, A.; Koszinowski, U.H.; Kloetzel, P.M.; Groettrup, M. The proteasome regulator PA28α/β can enhance antigen presentation without affecting 20S proteasome subunit composition. Eur. J. Immunol. 2000, 30, 3672–3679. [Google Scholar] [CrossRef] [PubMed]
- Van Hall, T.; Sijts, A.; Camps, M.; Offringa, R.; Melief, C.; Kloetzel, P.-M.; Ossendorp, F. Differential influence on cytotoxic T-lymphocyte epitope presentation by controlled expression of either proteasome immunosubunits or PA28. J. Exp. Med. 2000, 192, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Sijts, A.; Sun, Y.; Janek, K.; Kral, S.; Paschen, A.; Schadendorf, D.; Kloetzel, P.M. The role of the proteasome activator PA28 in MHC class I antigen processing. Mol. Immunol. 2002, 39, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sijts, A.J.A.M.; Song, M.; Janek, K.; Nussbaum, A.K.; Kral, S.; Schirle, M.; Stevanovic, S.; Paschen, A.; Schild, H.; et al. Expression of the proteasome activator PA28 rescues the presentation of a cytotoxic T-lymphocyte epitope on melanoma cells. Cancer Res. 2002, 62, 2875–2882. [Google Scholar] [PubMed]
- Textoris-Taube, K.; Henklein, P.; Pollmann, S.; Bergann, T.; Weisshoff, H.; Seifert, U.; Drung, I.; Mugge, C.; Sijts, A.; Kloetzel, P.M.; et al. The N-terminal flanking region of the TRP2360–368 melanoma antigen determines proteasome activator PA28 requirement for epitope liberation. J. Biol. Chem. 2007, 282, 12749–12754. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Sugahara, H.; Mizukami, S.; Murata, S.; Chiba, T.; Tanaka, K.; Yui, K.; Udono, H. Allele-selective effect of PA28 in MHC class I antigen processing. J. Immunol. 2008, 181, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; van den Broek, M.; Kostka, S.; Kraft, R.; Soza, A.; Schmidtke, G.; Kloetzel, P.-M.; Groettrup, M. Overexpression of the proteasome subunits LMP2, LMP7, and MECL-1, but not PA28α/β, enhances the presentation of an immunodominant lymphocytic choriomeningitis virus T-cell epitope. J. Immunol. 2000, 165, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, T.; Shimada, K.; Shibata, M.; Hata, M.; Sakamoto, M.; Takasaki, Y.; Sato, C.; Takahashi, T.; Nishida, Y. Cloning and nucleotide sequence of cDNA for Ki antigen, a highly conserved nuclear protein detected with sera from patients with systemic lupus erythematosus. Clin. Exp. Immunol. 1990, 79, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Wilk, S.; Chen, W.E.; Magnusson, R.P. Properties of the nuclear proteasome activator PA28γ (REGγ). Arch. Biochem. Biophys. 2000, 383, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Kawahara, H.; Tohma, S.; Yamamoto, K.; Kasahara, M.; Nabeshima, Y.; Tanaka, K.; Chiba, T. Growth retardation in mice lacking the proteasome activator PA28γ. J. Biol. Chem. 1999, 274, 38211–38215. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.F.; Runnels, H.A.; Schell, T.D.; Cho, Y.; Gibbons, R.; Tevethia, S.S.; Deepe, G.S.; Monaco, J.J. Immune defects in 28-kDa proteasome activator γ-deficient mice. J. Immunol. 2004, 172, 3948–3954. [Google Scholar] [CrossRef] [PubMed]
- Araya, R.; Takahashi, R.; Nomura, Y. Yeast two-hybrid screening using constitutive-active caspase-7 as bait in the identification of PA28γ as an effector caspase substrate. Cell Death Differ. 2002, 9, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, C.; Patel, R.; Blank, J.L. MEKK3 interacts with the PA28γ regulatory subunit of the proteasome. Biochem. J. 2003, 373, 71–79. [Google Scholar] [CrossRef]
- Li, X.; Lonard, D.M.; Jung, S.Y.; Malovannaya, A.; Feng, Q.; Qin, J.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. The SRC-3/AIB1 coactivator is degraded in a ubiquitin- and ATP-independent manner by the REGγ proteasome. Cell 2006, 124, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Amazit, L.; Long, W.; Lonard, D.M.; Monaco, J.J.; O’Malley, B.W. Ubiquitin- and ATP-independent proteolytic turnover of p21 by the REGγ-proteasome pathway. Mol. Cell 2007, 26, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Levy-Barda, A.; Lerenthal, Y.; Davis, A.J.; Chung, Y.M.; Essers, J.; Shao, Z.; van Vliet, N.; Chen, D.J.; Hu, M.C.; Kanaar, R.; et al. Involvement of the nuclear proteasome activator PA28γ in the cellular response to DNA double-strand breaks. Cell Cycle 2011, 10, 4300–4310. [Google Scholar] [CrossRef]
- Zannini, L.; Lecis, D.; Buscemi, G.; Carlessi, L.; Gasparini, P.; Fontanella, E.; Lisanti, S.; Barton, L.; Delia, D. REGγ proteasome activator is involved in the maintenance of chromosomal stability. Cell Cycle 2008, 7, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Baldin, V.; Militello, M.; Thomas, Y.; Doucet, C.; Fic, W.; Boireau, S.; Jariel-Encontre, I.; Piechaczyk, M.; Bertrand, E.; Tazi, J.; et al. A novel role for PA28γ-proteasome in nuclear speckle organization and SR protein trafficking. Mol. Biol. Cell 2008, 19, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R. Proteasome activator PA28γ regulates p53 by enhancing its MDM2-mediated degradation. EMBO J. 2008, 27, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, G.; Zhao, Y.; Zhao, D.; Wang, Y.; Wang, L.; Li, L.; Zeng, Y.; Dang, Y.; Wang, C.; et al. REGγ modulates p53 activity by regulating its cellular localization. J. Cell. Sci. 2010, 123, 4076–4084. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, D.; Wei, H.; Yao, L.; Dang, Y.; Amjad, A.; Xu, J.; Liu, J.; Guo, L.; Li, D.; et al. REGγ deficiency promotes premature aging via the casein kinase 1 pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 11005–11010. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.X.; Yuan, D.M.; Zhuo, Y.M.; Yi, X.; Zhou, J.; Xu, Z.X.; Zhou, J.L. The proteasome activator PA28γ, a negative regulator of p53, is transcriptionally up-regulated by p53. Int. J. Mol. Sci. 2014, 15, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Realini, C.; Jensen, C.C.; Zhang, Z.; Johnston, S.C.; Knowlton, J.R.; Hill, C.P.; Rechsteiner, M. Characterization of recombinant REGα, REGβ, and REGγ proteasome activators. J. Biol. Chem. 1997, 272, 25483–25492. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.; Pratt, G.; Rechsteiner, M. Multiple forms of the 20S multicatalytic and the 26S ubiquitin/ATP-dependent proteases from rabbit reticulocyte lysate. J. Biol. Chem. 1992, 267, 22362–22368. [Google Scholar] [PubMed]
- Blickwedehl, J.; Agarwal, M.; Seong, C.; Pandita, R.K.; Melendy, T.; Sung, P.; Pandita, T.K.; Bangia, N. Role for proteasome activator PA200 and postglutamyl proteasome activity in genomic stability. Proc. Natl. Acad. Sci. USA 2008, 105, 16165–16170. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigneron, N.; Van den Eynde, B.J. Proteasome Subtypes and Regulators in the Processing of Antigenic Peptides Presented by Class I Molecules of the Major Histocompatibility Complex. Biomolecules 2014, 4, 994-1025. https://doi.org/10.3390/biom4040994
Vigneron N, Van den Eynde BJ. Proteasome Subtypes and Regulators in the Processing of Antigenic Peptides Presented by Class I Molecules of the Major Histocompatibility Complex. Biomolecules. 2014; 4(4):994-1025. https://doi.org/10.3390/biom4040994
Chicago/Turabian StyleVigneron, Nathalie, and Benoît J. Van den Eynde. 2014. "Proteasome Subtypes and Regulators in the Processing of Antigenic Peptides Presented by Class I Molecules of the Major Histocompatibility Complex" Biomolecules 4, no. 4: 994-1025. https://doi.org/10.3390/biom4040994