
Effects of Complete and Partial Loss of the 24S-Hydroxycholesterol-Generating Enzyme Cyp46a1 on Behavior and Hippocampal Transcription in Mouse
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Behavioral Testing
2.3. Tissue Harvest and Sequencing
2.4. Measurement of 24S-HC in Hippocampus
3. Results
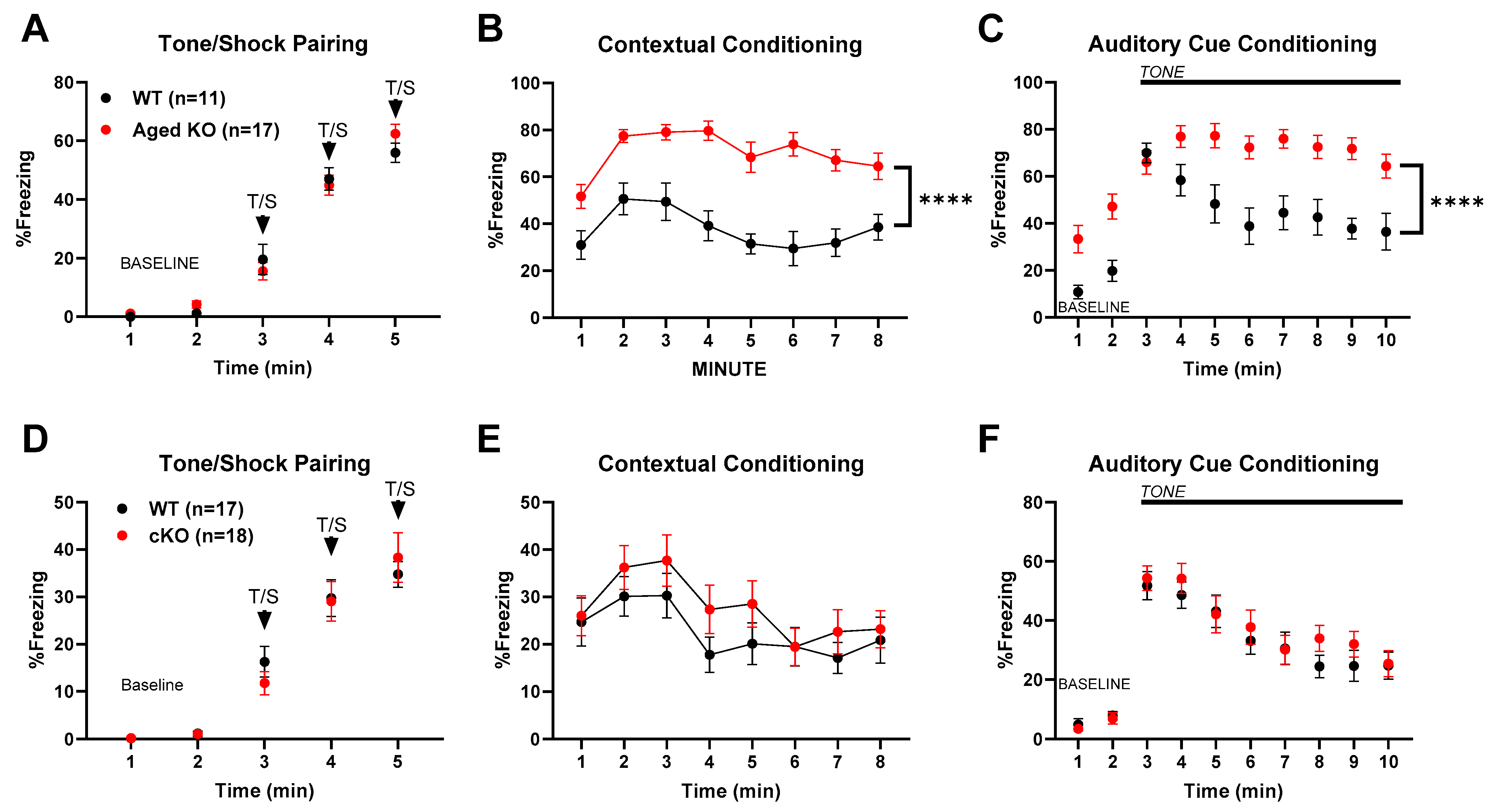
3.1. Behavioral Changes with Global and Partial Cyp46a1 Loss
3.2. Transcriptional Changes with Complete and Selective Cyp46a1 Loss
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pikuleva, I.A. Targeting Cytochrome P450 46A1 and Brain Cholesterol 24-Hydroxylation to Treat Neurodegenerative Diseases. Explor. Neuroprot. Ther. 2021, 1, 159–172. [Google Scholar] [CrossRef]
- Leoni, V.; Caccia, C. Study of Cholesterol Metabolism in Huntington’s Disease. Biochem. Biophys. Res. Commun. 2014, 446, 697–701. [Google Scholar] [CrossRef]
- Alavi, M.S.; Karimi, G.; Ghanimi, H.A.; Roohbakhsh, A. The Potential of CYP46A1 as a Novel Therapeutic Target for Neurological Disorders: An Updated Review of Mechanisms. Eur. J. Pharmacol. 2023, 949, 175726. [Google Scholar] [CrossRef]
- Ishikawa, M.; Yoshitomi, T.; Covey, D.F.; Zorumski, C.F.; Izumi, Y. Neurosteroids and Oxysterols as Potential Therapeutic Agents for Glaucoma and Alzheimer’s Disease. Neuropsychiatry 2018, 8, 344–359. [Google Scholar] [CrossRef]
- Sun, M.-Y.Y.; Linsenbardt, A.J.J.; Emnett, C.M.M.; Eisenman, L.N.N.; Izumi, Y.; Zorumski, C.F.F.; Mennerick, S. 24(S)-Hydroxycholesterol as a Modulator of Neuronal Signaling and Survival. Neuroscientist 2015, 22, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Covey, D.F.; Evers, A.S.; Izumi, Y.; Maguire, J.L.; Mennerick, S.J.; Zorumski, C.F. Neurosteroid Enantiomers as Potentially Novel Neurotherapeutics. Neurosci. Biobehav. Rev. 2023, 149, 105191. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.W.; Halford, R.W.; Ramirez, D.M.; Shah, R.; Kotti, T. Cholesterol 24-Hydroxylase: An Enzyme of Cholesterol Turnover in the Brain. Annu. Rev. Biochem. 2009, 78, 1017–1040. [Google Scholar] [CrossRef] [PubMed]
- Lund, E.G.; Guileyardo, J.M.; Russell, D.W. CDNA Cloning of Cholesterol 24-Hydroxylase, a Mediator of Cholesterol Homeostasis in the Brain. Proc. Natl. Acad. Sci. USA 1999, 96, 7238–7243. [Google Scholar] [CrossRef] [PubMed]
- Popiolek, M.; Izumi, Y.; Hopper, A.T.; Dai, J.; Miller, S.; Shu, H.-J.; Zorumski, C.F.; Mennerick, S.J. Effects of CYP46A1 Inhibition on Long-Term-Depression in Hippocampal Slices Ex Vivo and 24S-Hydroxycholesterol Levels in Mice in Vivo. Front. Mol. Neurosci. 2020, 13, 206. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-Y.M.-Y.; Taylor, A.; Zorumski, C.F.; Mennerick, S. 24S-Hydroxycholesterol and 25-Hydroxycholesterol Differentially Impact Hippocampal Neuronal Survival Following Oxygen-Glucose Deprivation. PLoS ONE 2017, 12, e0174416. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.M.; Doherty, J.J.; Robichaud, A.J.; Belfort, G.M.; Chow, B.Y.; Hammond, R.S.; Crawford, D.C.; Linsenbardt, A.J.; Shu, H.-J.; Izumi, Y.; et al. The Major Brain Cholesterol Metabolite 24(S)-Hydroxycholesterol Is a Potent Allosteric Modulator of N-Methyl-D-Aspartate Receptors. J. Neurosci. 2013, 33, 17290–17300. [Google Scholar] [CrossRef] [PubMed]
- Linsenbardt, A.J.; Taylor, A.; Emnett, C.M.; Doherty, J.J.; Krishnan, K.; Covey, D.F.; Paul, S.M.; Zorumski, C.F.; Mennerick, S. Different Oxysterols Have Opposing Actions at N-Methyl-D-Aspartate Receptors. Neuropharmacology 2014, 85, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Kotti, T.J.; Ramirez, D.M.; Pfeiffer, B.E.; Huber, K.M.; Russell, D.W. Brain Cholesterol Turnover Required for Geranylgeraniol Production and Learning in Mice. Proc. Natl. Acad. Sci. USA 2006, 103, 3869–3874. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-Y.; Izumi, Y.; Benz, A.; Zorumski, C.F.; Mennerick, S. Endogenous 24S-Hydroxycholesterol Modulates NMDAR-Mediated Function in Hippocampal Slices. J. Neurophysiol. 2016, 115, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Kliewer, S.A.; Moore, L.B.; Smith-Oliver, T.A.; Oliver, B.B.; Su, J.-L.; Sundseth, S.S.; Winegar, D.A.; Blanchard, D.E.; Spencer, T.A.; et al. Activation of the Nuclear Receptor LXR by Oxysterols Defines a New Hormone Response Pathway. J. Biol. Chem. 1997, 272, 3137–3140. [Google Scholar] [CrossRef] [PubMed]
- Janowski, B.A.; Willy, P.J.; Devi, T.R.; Falck, J.R.; Mangelsdorf, D.J. An Oxysterol Signalling Pathway Mediated by the Nuclear Receptor LXR Alpha. Nature 1996, 383, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Mast, N.; Lin, J.B.; Anderson, K.W.; Bjorkhem, I.; Pikuleva, I.A. Transcriptional and Post-Translational Changes in the Brain of Mice Deficient in Cholesterol Removal Mediated by Cytochrome P450 46A1 (CYP46A1). PLoS ONE 2017, 12, e0187168. [Google Scholar] [CrossRef] [PubMed]
- Lund, E.G.; Xie, C.; Kotti, T.; Turley, S.D.; Dietschy, J.M.; Russell, D.W. Knockout of the Cholesterol 24-Hydroxylase Gene in Mice Reveals a Brain-Specific Mechanism of Cholesterol Turnover. J. Biol. Chem. 2003, 278, 22980–22988. [Google Scholar] [CrossRef]
- Meljon, A.; Wang, Y.; Griffiths, W.J. Oxysterols in the Brain of the Cholesterol 24-Hydroxylase Knockout Mouse. Biochem. Biophys. Res. Commun. 2014, 446, 768–774. [Google Scholar] [CrossRef]
- Moutinho, M.; Nunes, M.J.; Gomes, A.Q.; Gama, M.J.; Cedazo-Minguez, A.; Rodrigues, C.M.P.; Björkhem, I.; Rodrigues, E. Cholesterol 24S-Hydroxylase Overexpression Inhibits the Liver X Receptor (LXR) Pathway by Activating Small Guanosine Triphosphate-Binding Proteins (SGTPases) in Neuronal Cells. Mol. Neurobiol. 2015, 51, 1489–1503. [Google Scholar] [CrossRef]
- Ramirez, D.M.; Andersson, S.; Russell, D.W. Neuronal Expression and Subcellular Localization of Cholesterol 24-Hydroxylase in the Mouse Brain. J. Comp. Neurol. 2008, 507, 1676–1693. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef]
- Miura, H.; Quadros, R.M.; Gurumurthy, C.B.; Ohtsuka, M. Easi-CRISPR for Creating Knock-in and Conditional Knockout Mouse Models Using Long SsDNA Donors. Nat. Protoc. 2018, 13, 195–215. [Google Scholar] [CrossRef]
- Maloney, S.E.; Yuede, C.M.; Creeley, C.E.; Williams, S.L.; Huffman, J.N.; Taylor, G.T.; Noguchi, K.N.; Wozniak, D.F. Repeated Neonatal Isoflurane Exposures in the Mouse Induce Apoptotic Degenerative Changes in the Brain and Relatively Mild Long-Term Behavioral Deficits. Sci. Rep. 2019, 9, 2779. [Google Scholar] [CrossRef] [PubMed]
- Dearborn, J.T.; Harmon, S.K.; Fowler, S.C.; O’Malley, K.L.; Taylor, G.T.; Sands, M.S.; Wozniak, D.F. Comprehensive Functional Characterization of Murine Infantile Batten Disease Including Parkinson-like Behavior and Dopaminergic Markers. Sci. Rep. 2015, 5, 12752. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential Expression Analysis of Multifactor RNA-Seq Experiments with Respect to Biological Variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. Differential Expression Analysis of Complex RNA-Seq Experiments Using EdgeR. In Statistical Analysis of Next Generation Sequence Data; Datta, S., Nettleton, D.S., Eds.; Springer: New York, NY, USA, 2014. [Google Scholar]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kutmon, M.; van Iersel, M.P.; Bohler, A.; Kelder, T.; Nunes, N.; Pico, A.R.; Evelo, C.T. PathVisio 3: An Extendable Pathway Analysis Toolbox. PLoS Comput. Biol. 2015, 11, e1004085. [Google Scholar] [CrossRef]
- van Iersel, M.P.; Kelder, T.; Pico, A.R.; Hanspers, K.; Coort, S.; Conklin, B.R.; Evelo, C. Presenting and Exploring Biological Pathways with PathVisio. BMC Bioinform. 2008, 9, 399. [Google Scholar] [CrossRef]
- Yao, Z.; van Velthoven, C.T.J.; Nguyen, T.N.; Goldy, J.; Sedeno-Cortes, A.E.; Baftizadeh, F.; Bertagnolli, D.; Casper, T.; Chiang, M.; Crichton, K.; et al. A Taxonomy of Transcriptomic Cell Types across the Isocortex and Hippocampal Formation. Cell 2021, 184, 3222–3241.e26. [Google Scholar] [CrossRef]
- Izumi, Y.; Mennerick, S.J.; Doherty, J.J.; Zorumski, C.F. Oxysterols Modulate the Acute Effects of Ethanol on Hippocampal N-Methyl-d-Aspartate Receptors, Long-Term Potentiation, and Learning. J. Pharmacol. Exp. Ther. 2021, 377, 181–188. [Google Scholar] [CrossRef]
- Radosevic, M.; Planagumà, J.; Mannara, F.; Mellado, A.; Aguilar, E.; Sabater, L.; Landa, J.; García-Serra, A.; Maudes, E.; Gasull, X.; et al. Allosteric Modulation of NMDARs Reverses Patients’ Autoantibody Effects in Mice. Neurol. Neuroimmunol. Neuroinflamm. 2022, 9, e1122. [Google Scholar] [CrossRef]
- Chen, W.; Chen, G.; Head, D.L.; Mangelsdorf, D.J.; Russell, D.W. Enzymatic Reduction of Oxysterols Impairs LXR Signaling in Cultured Cells and the Livers of Mice. Cell Metab. 2007, 5, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.M.; Lam, M.; Mast, N.; Moon, J.; Li, Y.; Maxfield, E.; Pikuleva, I.A. CYP46A1 Activation by Efavirenz Leads to Behavioral Improvement without Significant Changes in Amyloid Plaque Load in the Brain of 5XFAD Mice. Neurotherapeutics 2019, 16, 710–724. [Google Scholar] [CrossRef] [PubMed]
- Mast, N.; Saadane, A.; Valencia-Olvera, A.; Constans, J.; Maxfield, E.; Arakawa, H.; Li, Y.; Landreth, G.; Pikuleva, I.A. Cholesterol-Metabolizing Enzyme Cytochrome P450 46A1 as a Pharmacologic Target for Alzheimer’s Disease. Neuropharmacology 2017, 123, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega, C.; Mendonça, L.; Marcelo, A.; Lamazière, A.; Tomé, S.; Despres, G.; Matos, C.A.; Mechmet, F.; Langui, D.; den Dunnen, W.; et al. Restoring Brain Cholesterol Turnover Improves Autophagy and Has Therapeutic Potential in Mouse Models of Spinocerebellar Ataxia. Acta Neuropathol. 2019, 138, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Pikuleva, I.A.; Cartier, N. Cholesterol Hydroxylating Cytochrome P450 46A1: From Mechanisms of Action to Clinical Applications. Front. Aging Neurosci. 2021, 13, 696778. [Google Scholar] [CrossRef] [PubMed]
- Kacher, R.; Lamazière, A.; Heck, N.; Kappes, V.; Mounier, C.; Despres, G.; Dembitskaya, Y.; Perrin, E.; Christaller, W.; Sasidharan Nair, S.; et al. CYP46A1 Gene Therapy Deciphers the Role of Brain Cholesterol Metabolism in Huntington’s Disease. Brain 2019, 142, 2432–2450. [Google Scholar] [CrossRef] [PubMed]
- Burlot, M.A.; Braudeau, J.; Michaelsen-Preusse, K.; Potier, B.; Ayciriex, S.; Varin, J.; Gautier, B.; Djelti, F.; Audrain, M.; Dauphinot, L.; et al. Cholesterol 24-Hydroxylase Defect Is Implicated in Memory Impairments Associated with Alzheimer-like Tau Pathology. Hum. Mol. Genet. 2015, 24, 5965–5976. [Google Scholar] [CrossRef]
- Hudry, E.; Van Dam, D.; Kulik, W.; De Deyn, P.P.; Stet, F.S.; Ahouansou, O.; Benraiss, A.; Delacourte, A.; Bougnères, P.; Aubourg, P.; et al. Adeno-Associated Virus Gene Therapy with Cholesterol 24-Hydroxylase Reduces the Amyloid Pathology before or after the Onset of Amyloid Plaques in Mouse Models of Alzheimer’s Disease. Mol. Ther. 2010, 18, 44–53. [Google Scholar] [CrossRef]
- Boussicault, L.; Alves, S.; Lamazière, A.; Planques, A.; Heck, N.; Moumné, L.; Despres, G.; Bolte, S.; Hu, A.; Pagès, C.; et al. CYP46A1, the Rate-Limiting Enzyme for Cholesterol Degradation, Is Neuroprotective in Huntington’s Disease. Brain 2016, 139, 953–970. [Google Scholar] [CrossRef] [PubMed]
- Djelti, F.; Braudeau, J.; Hudry, E.; Dhenain, M.; Varin, J.; Bieche, I.; Marquer, C.; Chali, F.; Ayciriex, S.; Auzeil, N.; et al. CYP46A1 Inhibition, Brain Cholesterol Accumulation and Neurodegeneration Pave the Way for Alzheimer’s Disease. Brain 2015, 138, 2383–2398. [Google Scholar] [CrossRef] [PubMed]
- Parente, M.; Tonini, C.; Buzzelli, V.; Carbone, E.; Trezza, V.; Pallottini, V. Brain Cholesterol Biosynthetic Pathway Is Altered in a Preclinical Model of Fragile X Syndrome. Int. J. Mol. Sci. 2022, 23, 3408. [Google Scholar] [CrossRef] [PubMed]
- Hochgerner, H.; Singh, S.; Tibi, M.; Lin, Z.; Skarbianskis, N.; Admati, I.; Ophir, O.; Reinhardt, N.; Netser, S.; Wagner, S.; et al. Neuronal Types in the Mouse Amygdala and Their Transcriptional Response to Fear Conditioning. Nat. Neurosci. 2023, 26, 2237–2249. [Google Scholar] [CrossRef] [PubMed]
- Litvinchuk, A.; Suh, J.H.; Guo, J.L.; Lin, K.; Davis, S.S.; Bien-Ly, N.; Tycksen, E.; Tabor, G.T.; Remolina Serrano, J.; Manis, M.; et al. Amelioration of Tau and ApoE4-Linked Glial Lipid Accumulation and Neurodegeneration with an LXR Agonist. Neuron 2023, 112, 384–403.e8. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | LogFC | LogCPM | p Value | FDR | Gene Name |
|---|---|---|---|---|---|
| Cyp46a1 | −4.749 | 6.600 | 1.78 × 10−141 | 2.58 × 10−137 | cytochrome P450, family 46, subfamily a, polypeptide 1 |
| Gm8420 | 7.112 | 1.062 | 1.88 × 10−22 | 1.36 × 10−18 | predicted gene 8420 |
| Gm14094 | 7.278 | −0.200 | 6.78 × 10−13 | 3.27 × 10−9 | predicted gene 14094 |
| Olfr31 | 5.142 | −0.603 | 1.57 × 10−8 | 5.65 × 10−5 | olfactory receptor 31 |
| Rbm43 | 1.109 | 4.735 | 2.53 × 10−6 | 0.007305 | RNA binding motif protein 43 |
| Gene | LogFC | LogCPM | p Value | FDR | Gene Name |
|---|---|---|---|---|---|
| Cyp46a1 | −6.040 | 6.527 | 5.55 × 10−112 | 8.19 × 10−108 | cytochrome P450, family 46, subfamily a, polypeptide 1 |
| Trim21 | 3.453 | 0.427 | 2.10 × 10−7 | 0.001548 | tripartite motif containing 21 |
| 4930481B07Rik | 2.744 | 1.040 | 1.89 × 10−6 | 0.009277 | 4930481B07Rik RIKEN cDNA |
| Chst3 | −2.570 | 0.367 | 3.67 × 10−6 | 0.013563 | carbohydrate sulfotransferase 3 |
| Insig1 | −1.260 | 4.886 | 8.31 × 10−6 | 0.024536 | insulin induced gene 1 |
| Hhe× | −2.333 | −0.026 | 1.32 × 10−5 | 0.032547 | hematopoietically expressed homeobox |
| Tox3 | 1.811 | 1.779 | 2.03 × 10−5 | 0.038145 | tox high mobility group box family member 3 |
| Otos | −3.473 | 1.457 | 2.07 × 10−5 | 0.038145 | otospiralin |
| Gene | LogFC | LogCPM | p Value | FDR | Gene Name |
|---|---|---|---|---|---|
| Cyp46a1 | −4.472 | 3.202 | 1.15 × 10−47 | 1.64 × 10−43 | cytochrome P450, family 46, subfamily a, polypeptide 1 |
| Gm8730 | 4.835 | 2.075 | 8.31 × 10−33 | 5.93 × 10−29 | predicted pseudogene 8730 |
| Gm8420 | 6.367 | 1.185 | 4.22 × 10−31 | 2.01 × 10−27 | predicted gene 8420 |
| Tagap | 2.952 | 1.641 | 3.75 × 10−18 | 1.34 × 10−14 | T-cell activation Rho GTPase-activating protein |
| Tmem181b.ps | 2.101 | 3.050 | 1.72 × 10−17 | 4.91 × 10−14 | transmembrane protein 181B, pseudogene |
| Gm9625 | 2.509 | 2.528 | 2.20 × 10−17 | 5.22 × 10−14 | predicted gene 9625 |
| Ccr6 | 5.854 | 0.071 | 1.05 × 10−15 | 2.14 × 10−12 | Mus musculus chemokine (C-C motif) receptor 6 |
| Gm14094 | 4.628 | 0.098 | 6.65 × 10−15 | 1.19 × 10−11 | predicted gene 14094 |
| Ubc | −1.025 | 8.212 | 4.82 × 10−14 | 7.64 × 10−11 | ubiquitin C |
| Fdft1 | −1.172 | 6.065 | 2.96 × 10−11 | 4.22 × 10−8 | Squalene synthase |
| Msmo1 | −1.188 | 7.134 | 8.37 × 10−10 | 1.09 × 10−6 | Methylsterol monooxygenase 1 |
| Rplp0 | −0.909 | 7.425 | 2.68 × 10−9 | 3.18 × 10−6 | ribosomal protein, large, P0 |
| Rplp0.ps1 | 4.744 | −0.799 | 8.43 × 10−9 | 8.94 × 10−6 | ribosomal protein, large, P0, pseudogene 1 |
| Nsdhl | −0.868 | 5.274 | 8.77 × 10−9 | 8.94 × 10−6 | Sterol-4-alpha-carboxylate 3-dehydrogenase, decarboxylating |
| Gm29340 | 2.834 | 1.906 | 1.18 × 10−7 | 0.00011183 | predicted gene 29340 |
| Sqle | −1.012 | 4.827 | 4.30 × 10−7 | 0.000383502 | squalene epoxidase |
| Ptprg | 1.003 | 5.433 | 1.09 × 10−6 | 0.000912133 | receptor-type tyrosine-protein phosphatase gamma isoform 2 |
| Hipk1 | 1.091 | 6.961 | 1.73 × 10−6 | 0.001368802 | Homeodomain-interacting protein kinase 1 |
| Fdps | −0.758 | 5.250 | 1.87 × 10−6 | 0.001387266 | Farnesyl pyrophosphate synthase |
| Dbi | −0.861 | 7.751 | 1.94 × 10−6 | 0.001387266 | diazepam binding inhibitor |
| Ly86 | −0.828 | 4.908 | 4.14 × 10−6 | 0.002783167 | Lymphocyte antigen 86 |
| Atf4 | −0.818 | 6.185 | 4.29 × 10−6 | 0.002783167 | Cyclic AMP-dependent transcription factor ATF-4 |
| Asb1 | 2.003 | 6.010 | 5.17 × 10−6 | 0.003108183 | Ankyrin repeat and SOCS box protein 1 |
| Fn1 | 1.088 | 4.560 | 5.23 × 10−6 | 0.003108183 | Fibronectin Anastellin |
| Arhgef37 | 1.913 | 0.135 | 6.31 × 10−6 | 0.003604708 | Rho guanine nucleotide exchange factor 37 |
| Sc5d | −0.735 | 6.988 | 8.22 × 10−6 | 0.004513714 | sterol-C5-desaturase |
| Gm7180 | 1.790 | 0.740 | 1.16 × 10−5 | 0.006143589 | predicted pseudogene 7180 |
| Clasp1 | 0.822 | 6.013 | 1.25 × 10−5 | 0.00638559 | Mus musculus CLIP associating protein 1 |
| Htr1a | 0.603 | 6.326 | 1.31 × 10−5 | 0.006427525 | 5-hydroxytryptamine (serotonin) receptor 1A |
| Ilf3 | 0.774 | 7.299 | 2.96 × 10−5 | 0.014060774 | Interleukin enhancer-binding factor 3 |
| Abca1 | 0.926 | 5.119 | 3.96 × 10−5 | 0.01825061 | ATP-binding cassette, sub-family A member 1 |
| Gene | LogFC | LogCPM | p Value | FDR | Gene Name |
|---|---|---|---|---|---|
| Cyp46a1 | −3.770 | 2.482 | 5.07 × 10−36 | 7.49 × 10−32 | cytochrome P450, family 46, subfamily a, polypeptide 1 |
| Ccr6 | 4.937 | 0.363 | 2.82 × 10−18 | 2.08 × 10−14 | Mus musculus chemokine (C-C motif) receptor 6 |
| Hmgcs1 | −1.423 | 7.614 | 2.51 × 10−15 | 1.24 × 10−11 | Hydroxymethylglutaryl-CoA synthase, cytoplasmic |
| Tagap | 2.713 | 1.343 | 1.45 × 10−14 | 5.34 × 10−11 | T-cell activation Rho GTPase-activating protein |
| Ldlr | −1.370 | 4.099 | 6.47 × 10−14 | 1.72 × 10−10 | Low-density lipoprotein receptor |
| Msmo1 | −1.184 | 6.772 | 6.98 × 10−14 | 1.72 × 10−10 | Methylsterol monooxygenase 1 |
| Scd1 | −1.156 | 6.407 | 4.11 × 10−12 | 8.67 × 10−9 | stearoyl-Coenzyme A desaturase 1 |
| 1700023H06Rik | 4.020 | 0.221 | 2.71 × 10−11 | 5.00 × 10−8 | RIKEN cDNA 1700023H06 gene |
| Aplnr | −4.997 | −0.024 | 3.06 × 10−10 | 5.01 × 10−7 | Apelin receptor |
| Foxn3 | −0.731 | 6.114 | 1.88 × 10−8 | 2.66 × 10−5 | Forkhead box protein N3 |
| Fdft1 | −0.978 | 5.866 | 1.98 × 10−8 | 2.66 × 10−5 | Squalene synthase |
| Gm29340 | 3.305 | 1.711 | 2.24 × 10−7 | 0.000275929 | predicted gene 29340 |
| Rgs13 | −1.373 | 2.612 | 3.93 × 10−7 | 0.000431055 | Regulator of G-protein signaling 13 |
| Slc27a3 | 1.245 | 2.314 | 4.09 × 10−7 | 0.000431055 | solute carrier family 27, member 3 |
| Erap1 | −1.083 | 3.175 | 5.77 × 10−7 | 0.000568015 | Endoplasmic reticulum aminopeptidase 1 |
| Sqle | −0.984 | 4.735 | 1.50 × 10−6 | 0.001384955 | squalene epoxidase |
| Lrrc55 | −0.867 | 4.527 | 2.35 × 10−6 | 0.002044269 | Leucine-rich repeat-containing protein 55 |
| Pik3r1 | −0.635 | 6.415 | 3.26 × 10−6 | 0.002674551 | phosphoinositide-3-kinase regulatory subunit 1 |
| Acvr1c | −0.887 | 5.867 | 4.81 × 10−6 | 0.003737493 | Activin receptor type-1C |
| Slc7a11 | −0.948 | 4.993 | 7.18 × 10−6 | 0.005301361 | solute carrier family 7, member 11 |
| Nhlh1 | −1.539 | 1.424 | 8.16 × 10−6 | 0.005740722 | nescient helix loop helix 1 |
| Prkd3 | −0.780 | 4.093 | 1.28 × 10−5 | 0.008590708 | protein kinase D3 |
| Ddn | 0.626 | 7.228 | 1.99 × 10−5 | 0.012805329 | Dendrin |
| Icam1 | −1.971 | 0.929 | 2.27 × 10−5 | 0.013449952 | intercellular adhesion molecule 1 |
| Fst | 1.216 | 3.318 | 2.28 × 10−5 | 0.013449952 | follistatin FST315 |
| Idh1 | −0.569 | 5.968 | 2.51 × 10−5 | 0.014240691 | Isocitrate dehydrogenase [NADP] cytoplasmic |
| Arsi | 1.359 | 1.213 | 2.70 × 10−5 | 0.014767094 | Arylsulfatase I |
| Ube2t | 1.207 | 1.824 | 2.98 × 10−5 | 0.015715329 | ubiquitin-conjugating enzyme E2T |
| Insig1 | −0.753 | 3.350 | 3.28 × 10−5 | 0.01672288 | Insulin-induced 1 protein |
| Gene | LogFC | LogCPM | p Value | FDR | Gene Name |
|---|---|---|---|---|---|
| Cyp46a1 | −4.283 | 5.941 | 9.79 × 10−52 | 1.47 × 10−47 | cytochrome P450, family 46, subfamily a, polypeptide 1 |
| Msmo1 | −1.382 | 6.848 | 1.81 × 10−20 | 1.37 × 10−16 | methylsterol monoxygenase 1 |
| Fdft1 | −1.457 | 5.659 | 5.61 × 10−18 | 2.81 × 10−14 | farnesyl diphosphate farnesyl transferase 1 |
| Sqle | −1.515 | 5.548 | 5.37 × 10−16 | 2.02 × 10−12 | squalene epoxidase |
| Hmgcs1 | −1.24 | 7.655 | 1.35 × 10−14 | 4.06 × 10−11 | 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 1 |
| Cyp51 | −1.082 | 5.776 | 9.99 × 10−13 | 2.50 × 10−9 | cytochrome P450, family 51 |
| Hmgcr | −1.099 | 6.661 | 1.86 × 10−11 | 4.00 × 10−8 | 3-hydroxy-3-methylglutaryl-Coenzyme A reductase |
| Insig1 | −1.711 | 5.536 | 4.44 × 10−10 | 8.35 × 10−7 | insulin induced gene 1 |
| Dhcr24 | −1.354 | 4.014 | 2.49 × 10−9 | 4.17 × 10−6 | 24-dehydrocholesterol reductase |
| Ldlr | −2.279 | 3.86 | 6.37 × 10−8 | 9.59 × 10−5 | low density lipoprotein receptor |
| Cdr1 | −0.743 | 6.427 | 7.39 × 10−8 | 0.000101 | cerebellar degeneration related antigen 1 |
| Nsdhl | −1.047 | 4.737 | 2.70 × 10−7 | 0.000338 | NAD(P) dependent steroid dehydrogenase-like |
| Abca1 | 1.049 | 5.172 | 3.38 × 10−7 | 0.000391 | ATP-binding cassette, sub-family A (ABC1), member 1 |
| Adarb1 | −0.992 | 5.413 | 3.98 × 10−7 | 0.000421 | adenosine deaminase, RNA-specific, B1 |
| Rgs4 | −1.128 | 7.309 | 4.24 × 10−7 | 0.000421 | regulator of G-protein signaling 4 |
| Sostdc1 | −6.799 | 3.501 | 4.48 × 10−7 | 0.000421 | sclerostin domain containing 1 |
| Ccbe1 | −5.451 | 1.559 | 1.28 × 10−6 | 0.001129 | collagen and calcium binding EGF domains 1 |
| Mvd | −0.979 | 4.083 | 2.25 × 10−6 | 0.001871 | mevalonate (diphospho) decarboxylase |
| Sh2d3c | −1.142 | 3.226 | 2.36 × 10−6 | 0.001871 | SH2 domain containing 3C |
| Sc5d | −0.675 | 6.354 | 2.75 × 10−6 | 0.001975 | sterol-C5-desaturase |
| Dnajc6 | −0.626 | 6.977 | 2.76 × 10−6 | 0.001975 | DnaJ heat shock protein family (Hsp40) member C6 |
| Sv2b | −0.658 | 8.158 | 2.91 × 10−6 | 0.00199 | synaptic vesicle glycoprotein 2 b |
| Gm10384 | 2.935 | −0.252 | 4.33 × 10−6 | 0.002834 | predicted gene 10384 |
| Npr3 | −1.129 | 3.34 | 5.82 × 10−6 | 0.003652 | natriuretic peptide receptor 3 |
| Kcng2 | −1.532 | 2.191 | 6.82 × 10−6 | 0.004102 | potassium voltage-gated channel, subfamily G, member 2 |
| C4b | 1.041 | 5.116 | 7.12 × 10−6 | 0.004123 | complement component 4B (Chido blood group) |
| Cfap45 | −2.203 | 1.157 | 8.71 × 10−6 | 0.004855 | cilia and flagella associated protein 45 |
| Pcp4 | −3.375 | 4.502 | 1.21 × 10−5 | 0.006514 | Purkinje cell protein 4 |
| Nppc | −1.673 | 3.004 | 1.28 × 10−5 | 0.006631 | natriuretic peptide type C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, H.-J.; Ziolkowski, L.H.; Salvatore, S.V.; Benz, A.M.; Wozniak, D.F.; Yuede, C.M.; Paul, S.M.; Zorumski, C.F.; Mennerick, S. Effects of Complete and Partial Loss of the 24S-Hydroxycholesterol-Generating Enzyme Cyp46a1 on Behavior and Hippocampal Transcription in Mouse. Biomolecules 2024, 14, 254. https://doi.org/10.3390/biom14030254
Shu H-J, Ziolkowski LH, Salvatore SV, Benz AM, Wozniak DF, Yuede CM, Paul SM, Zorumski CF, Mennerick S. Effects of Complete and Partial Loss of the 24S-Hydroxycholesterol-Generating Enzyme Cyp46a1 on Behavior and Hippocampal Transcription in Mouse. Biomolecules. 2024; 14(3):254. https://doi.org/10.3390/biom14030254
Chicago/Turabian StyleShu, Hong-Jin, Luke H. Ziolkowski, Sofia V. Salvatore, Ann M. Benz, David F. Wozniak, Carla M. Yuede, Steven M. Paul, Charles F. Zorumski, and Steven Mennerick. 2024. "Effects of Complete and Partial Loss of the 24S-Hydroxycholesterol-Generating Enzyme Cyp46a1 on Behavior and Hippocampal Transcription in Mouse" Biomolecules 14, no. 3: 254. https://doi.org/10.3390/biom14030254