
Site-Specific Transient Receptor Potential Channel Mechanisms and Their Characteristics for Targeted Chronic Itch Treatment


1
Gachon Pain Center and Department of Physiology, College of Medicine, Gachon University, Incheon 21999, Republic of Korea
2
Department of Anesthesiology and Pain Medicine, Gil Medical Center, Gachon University, Incheon 21565, Republic of Korea
*
Authors to whom correspondence should be addressed.
Biomolecules 2024, 14(1), 107; https://doi.org/10.3390/biom14010107
Submission received: 28 December 2023
/
Revised: 10 January 2024
/
Accepted: 13 January 2024
/
Published: 15 January 2024
(This article belongs to the Section Molecular Biology)
Abstract
:Chronic itch is a debilitating condition with limited treatment options, severely affecting quality of life. The identification of pruriceptors has sparked a growing interest in the therapeutic potential of TRP channels in the context of itch. In this regard, we provided a comprehensive overview of the site-specific expression of TRP channels and their associated functions in response to a range of pruritogens. Although several potent antipruritic compounds that target specific TRP channels have been developed and have demonstrated efficacy in various chronic itch conditions through experimental means, a more thorough understanding of the potential for adverse effects or interactions with other TRP channels or GPCRs is necessary to develop novel and selective therapeutics that target TRP channels for treating chronic itch. This review focuses on the mechanism of itch associated with TRP channels at specific sites, from the skin to the sensory neuron, with the aim of suggesting specific therapeutic targets for treating this condition.
1. Introduction
Itch, or pruritus, is defined as an unpleasant sensation that elicits an innate scratching response. Irritants such as insect bites and contact with poisonous plants, leading to acute itch, can be readily alleviated by scratching. This response may be natural; however, it can become pathological if immense suffering from chronic itch lasts more than 6 weeks [1]. Chronic itch is classified into four categories: dermatologic, systemic, neuropathic, and psychogenic [2,3,4]. Dermatologic itch conditions in atopic dermatitis (AD), psoriasis, and xerosis stem from skin diseases [3]. Systemic itch arises from organs other than the skin and always accompanies diseases of organs, such as conditions in cholestatic pruritus and uremic pruritus [5]. Neuropathic itch results from nerve injury and arises from diseases of the central or peripheral nervous system [6]. Examples of neuropathic itch include neuropathy, nerve compression or irritation, multiple sclerosis, and brain tumors. Psychogenic itch is caused by psychological or psychiatric disorders [7]. Though chronic itch has been significantly impairing the quality of patients’ lives by interrupting their sleep or causing anxiety or depression [8,9], an understanding of the molecular and neural mechanisms of chronic itch remains limited.
Itch has been regarded as a sub-modality or a mild form of pain [10] due to the similarities in the two sensations [11]. In 2009, Chen et al. discovered itch-specific neurons in mice and claimed that itch and pain are two distinct sensations [12,13]. Though not identical, the two sensations are closely related, as itch-sensing nerve fibers express two families of receptors: G protein-coupled receptors (GPCRs) and the transient receptor potential (TRP) [14,15]. TRP channels are non-selective ion channels primarily located on the plasma membrane in various cells and are divided into six families: TRPV (vanilloid), TRPA (ankyrin), TRPM (melastatin), TRPP (polycystin), TRPC (canonical), and TRPML (mucolipin) [16]. Among the members of the TRP channel family, TRPV1, TRPA1, TRPV3, and TRPV4 are especially known to be involved in itch transduction [17,18,19].
In this review, we provide an overview of the current understanding of the mechanisms of itch, beginning with the initiation of itch at the site of the skin and tracing its progression to the itch sensory neurons where the sensation is perceived and conveyed. Additionally, we discuss the role of TRP channels as key players in chronic itch, aiming to suggest specific therapeutic targets for treating this condition.
2. Mechanism of Itch from Skin to Peripheral Sensory Neuron
During an episode of acute itch, the skin is distributed by a breach or chemical insult (such as chemical mediators or insect bites), which in turn follows a pruriceptive itch [20]. Free nerve endings of peripheral sensory nerve fibers terminate in the skin and detect the changes in the local chemical environment. These changes involve various chemical mediators, including histamine, serotonin, proteases, chemokines, and cytokines released by keratinocytes and local immune cells [21,22].
The peripheral sensory nerve fibers conveying the itch sensation are broadly classified into unmyelinated C-fibers and lightly myelinated Aδ-fibers [23,24]. Unmyelinated C-fibers detect and transmit pruriceptive information more slowly than myelinated Aδ-fibers. Unmyelinated C-fibers are subdivided into peptidergic and non-peptidergic fibers [24,25,26]. Peptidergic C-fibers contain large vesicles that release inflammatory neuropeptides such as substance P (SP) and calcitonin gene-related peptide (CGRP) at both central and peripheral terminals, whereas non-peptidergic fibers express the purinergic receptor P2 X3 and isolectin B4 (IB4) [26]. Ringkamp et al. reported the involvement of A fibers in itch perception, demonstrating attenuated itch sensation when the conduction of myelinated fibers was selectively blocked [23]. Among many subtypes of primary afferent nerves originating from the distal, a single nerve subtype consisting of an itch receptor is responsible for transducing pruriceptive stimuli to primary afferent dorsal root ganglion (DRG) neurons and then to the higher centers [27]. In the itch signal transduction, the TRP family is recognized as a key player. Among the TRP family in mammals, the five subgroups, the TRP vanilloids 1 (TRPV1), TRPV3, TRPV4, TRPA 1, and TRPM 8, are predominantly implicated in itch transduction [28].
One of the most commonly studied pruritogen, contributing to the initiation and modulation of itch sensation, is histamine, secreted by various cells, including T-cells, mast cells, and keratinocytes [29]. Based on this mechanism, itch is generally classified as either histamine-dependent or histamine-independent [30]. Acute itch, manifested as changes in the local chemical environment, can be detected through histamine-mediated mechanisms; however, chronic itch typically involves histamine-independent mechanisms [14]. Understanding how pruritogens interact with specific TRP channels provides insights into the molecular mechanisms of itch transduction. Furthermore, targeting these interactions holds potential for developing therapeutic interventions to alleviate itching associated with various skin conditions.
2.1. Detection of Itch through the Skin
When the skin barrier is damaged due to genetic, inflammatory, and environmental causes, passive water loss through the skin increases and induces the sensation of itch through non-myelinated C-fiber activation [31]. The itch–scratch cycle initiation leads to epidermal damage, perpetuating the itch sensation [31]. Various itch-sensing receptors are characterized in the skin and immune cells. Histamine receptors H1R, H2R, H3R, and H4R are distributed across various tissues [32]. H1R binds to Gq/G11 proteins and activates phospholipase A2 [33], phospholipase Cβ3 (PLCβ3) [34], protein kinase Cδ (PCKδ) [35], and TRPV1 [35,36], resulting in calcium influx and firing action potentials in primary sensory neurons. Mas-related G protein-coupled receptors (Mrgprs) are broadly required for detecting both exogenous pruritogens and endogenous itch mediators from keratinocytes and immune cells [37]. Serotonin (5-hydroxytryptamine, 5-HT) receptors (HTRs) are expressed in the skin by immune cells and sensory neurons, influencing immune response and sensory perception [38]. In mice, 5-HT is released by mast cells, acting as a pruritogen at lower doses than histamine [39]. Toll-like receptors (TLRs) are expressed across keratinocytes, immune cells, and neurons, and they detect pathogen-associated molecular patterns [40]. Among TLRs, TLR3, TLR4, TLR5, and TLR7 have been known to mediate itch in mice [41,42]. Protease-activated receptors (PARs), especially PAR2 and PAR4, have been implicated in itch initiation by detecting various exogenous and endogenous proteases [43,44,45]. They are expressed by keratinocytes, immune cells, and neurons [46]. Finally, cytokines-related type 2 immune responses are increasingly recognized for their role in itch perception. Keratinocytes release TSLP, a major instigator of the T helper (Th) 2 response, in response to various stimuli, including allergy and proteolytic PAR2 activation [47,48,49]. High TSLP expression in the skin is reported as a feature of atopic dermatitis (AD) [50].
2.2. Transduction of the Itch Signal through the Peripheral Sensory Neuron
Two theories for pruriceptive sensory processing in the nervous system have been postulated: the specificity and pattern theories [51,52,53]. The former asserts the existence of specific types of sensory nerve fibers and spinal cord neurons that transmit itch-specific information to the central nervous system; however, the latter states that the itch sensation is encoded by many sensory receptors and spinal cord neurons that compose the collective pattern of neuronal activity, determining the ultimate sensation experienced. Cumulative evidence increasingly supports the specificity theory over the pattern theory, although much remains to be learned.
As multiple nerve subtypes are activated, the specific subtypes of sensory neurons transducing pruriceptive itch become complex. For example, cutaneous C-fibers respond to both histamine and capsaicin [54,55]. Similar to C-fibers, an evaluation of the sensory nerves that express MrgprA3-expressing DRG neurons also express receptors for histamine, gastrin-releasing peptide (GRP) [56], and capsaicin (TRPV1) [57]. Han et al. found that the ablation of MrgprA3-expressing neurons in the DRG reduced itch behavior without disturbing pain [17]. Moreover, the expression of TLR7 in primary sensory neurons is required specifically for inducing itch, excluding pain. [41]. However, given the limitations of these studies, the potential role of the investigated molecules or neurons in mediating pain cannot be dismissed [14]. Pruriceptors on free nerve endings in cutaneous primary sensory neurons are activated by pruritogens and evoke an itch sensation [58]. Besides pruritogens, various inflammatory mediators, including adenosine 5′-triphosphate (ATP), thymic stromal lymphopoietin, endothelins, prostaglandins, nitric oxide, histamine, and serotonin, are released by keratinocytes and directly sensitize or activate primary sensory neurons to initiate itch signal [59]. Similarly, pruritogenic inflammatory mediators are also released by innate immune cells, including mast cells, macrophages, neutrophils, and dendritic cells [60], and the interplay between adaptive immune cells and neurons then plays a crucial role in initiating the itch signal [61,62,63].
3. Functional Roles of TRP Channels in the Skin
TRP channels are expressed in various excitable and unexcitable cells [64,65,66,67,68,69]. Studies have also revealed that TRP channels are involved in regulating skin physiology [70,71,72,73,74,75]. Despite the limited studies on TRP channels associated with skin and epidermis in itch, TRPV3 and TRPV4 channels (Figure 1) have been studied.
3.1. Skin TRPV3 in Itch
TRPV3 is a warm temperature (>33 °C)-sensitive non-selective cation channel expressed in skin keratinocytes [73,76,77,78] and poorly detected in DRG neurons or the spinal cord [73]. Similar to other polymodal TRP channels, TRPV3 is also activated by chemicals such as eugenol, carvacrol, thymol, camphor, and 2-APB [76,79]. Notably, TRPV3 is expressed in keratinocytes in mice and not in the peripheral or central nervous system, like DRG neurons or the spinal cord [73]. Studies using TRPV3 knockout mice showed markedly diminished heat responses, whereas other sensory stimuli were maintained [76]. Moreover, hair abnormalities were also observed in TRPV3 knockout mice [76,80]. Therefore, the physiological role of TRPV3 for thermosensation, hair regulation, and dermatitis has been highlighted.
In the context of itch, in vivo studies using gain-of-function TRPV3 mutation (Gly573 to Ser or Cys) in DS-Nh mice showed hairless and AD-like symptoms with spontaneous scratching [80,81]. AD is an inflammatory skin disease with intractable, chronic itch [82,83]. Moreover, gain-of-function TRPV3 Gly573Ser mutant in mice keratinocytes led to similarities with the clinical symptoms of human AD, such as skin inflammation, pruritus, immune cell infiltration, hyperkeratosis, upregulation of nerve growth factors, and systemic symptoms with elevated proinflammatory cytokines [84]. While TRPV3 Gly573 mutation mice indicated the involvement of TRPV3 in itch, the Gly573 missense mutation was also discovered in Olmsted syndrome, which is a rare congenital disease characterized by skin hyperplasia, diffuse palmoplantar keratoderma, alopecia, and severe pruritus [85]. TRPV3 knockout mice showed skin-related loss-of-function phenotypes, including curly whiskers, wavy hair, a thin stratum corneum, and misaligned hair follicles; however, they did not show itch-related scratching behaviors [86]. Moreover, TRPV3 lacking keratinocytes impaired protease-activated receptor 2 (PAR2) function in response to PAR2 agonists, leading to reduced neuronal activation and scratching behaviors [87]. Overall, these studies suggest a major role of TRPV3 in pruriceptive itch through skin keratinocytes.
3.2. Skin TRPV4 in Itch
TRPV4 is another temperature (27 and 35 °C)-activated channel [88], mainly expressed in keratinocytes, although less than TRPV3 in keratinocytes and in much higher levels than those found in DRG neurons [89]. TRPV4 is involved in skin barrier recovery [90], intercellular junction formation in keratinocytes [91], and intracellular calcium concentration; thus, accelerating barrier recovery after stratum corneum disruption [92,93].
A recent study on human chronic pruritus found increased expression in epidermal TRPV4 [94], suggesting a crucial role of TRPV4 in pruritus. Several TRPV4 knockout studies showed the role of TRPV4 in 5-HT- or histamine-induced itch. Reduced scratching behaviors in response to 5-HT were displayed in TRPV4 knockout mice, compared to wild-type mice [95]. In global and keratinocyte-specific TRPV4 knockout mice, scratching responses induced by histaminergic pruritogens, such as histamine, compound 48/80, and ET-1, were significantly reduced [96,97], suggesting an important role of TRPV4 in histamine-induced itch. Although controversial, the number of non-histaminergic pruritogen chloroquine (CQ)-induced scratching behaviors was increased in global TRPV4 knockout mice [95]. On the contrary, while one study found that both global or skin-specific TRPV4 knockout mice showed no effect of CQ-induced scratching [97], another study showed a significant attenuation in CQ-induced scratching behaviors in global TRPV4 knockout mice [96]. Recently, lysophosphatidylcholine (LPC), a cholestatic pruritogen, was found to directly activate TRPV4 in skin keratinocytes, triggering micro-RNA-146a release to activate TRPV1 pruriceptor neurons [98]. This study proposed the critical role of skin as a sensory organ with a new pathway of pruriception. Furthermore, the involvement of TRPV4 is supported by the deletion of TRPV4 in macrophages and keratinocytes, which showed reductions in both allergic and nonallergic chronic itch in mice [99,100]. Thus, the evidence represents TRPV4 as a potential therapeutic target for both allergic and nonallergic chronic itch conditions.
4. Functional Roles of TRP Channel in Sensory Neurons
Before the signal transduction of itch from the periphery in the skin to the central nervous system (i.e., spinal cord and brain), the signals pass through the DRG sensory neurons. Various TRP channels in sensory neurons are activated by external and internal itch mediators and metabolic byproducts via GPCRs, TLRs, integrin receptors, and immune receptors complex [101], and the sensory neurons are depolarized to transmit signals. Among the TRP channels, TRPA1, TRPV1, TRPV4, TRPM8, TRPC3, and TRPC4 (Figure 1) are generally known for their itch generation and transduction from sensory nerves such as DRG neurons.
4.1. Sensory TRPA1 in Itch
TRPA1 is a non-selective cation channel named after cytosolic N-termini with 14 ankyrin repeats and is activated by noxious cold temperature, mechanical sensation, electrophilic compounds (allyl isothiocyanate, cinnamaldehyde, diallyl disulfide, and allicin) as well as endogenous reactive oxygen species (hydrogen peroxide and 4-hydroxynonenal) [102]. Besides the physiological role of sensory detection through thermal and chemical stimuli, TRPA1 also contributes to the chronic [103] and acute histamine-independent pruritis evoked by CQ [104] and proenkephalin product, BAM8-22 [104,105,106]. Overexpression of TRPA1 in mast cells, keratinocytes, and dermal sensory neurons was found in human and murine AD models [103].
TRPA1 is an essential downstream mediator of GPCR signaling involved in histamine-independent itch [104]. Pruritogen-sensing GPCRs include the TSLP receptor, the bile acid receptor TGR5, and the MrgprA3 and MrgprC11, which can modulate TRPA1 positively [57,107,108]. In response to histaminergic signaling, TSLP released by keratinocytes activates TRPA1 downstream of the TSLP receptor and allows for calcium entry into the sensory neurons to promote itch signaling [107]. Similarly, a release of the pruritogenic neuropeptide GRP in response to the bile acids is diminished in pharmacological inhibition and genetic ablation of TRPA1 [108]. Thus, TRPA1 is activated and sensitized by TGR5 by a Gβγ- and PCK-dependent mechanism, and the overexpression of TGR5 that induces exacerbated spontaneous scratching is prevented with the treatment of a TRPA1 antagonist [108]. Histamine-independent itch mediators such as CQ and BAM8-22 peptide activate MrgprA3 with Gβγ signaling and MrgprC11 with PLC signaling, respectively [104]. One study, however, reported the lack of TRPA1 or TRPV1 involvement in the activation of GPCRs with CQ or histamine in inducing membrane depolarization and action potential at the peripheral C-fiber terminals of itch nerves [109]. Notably, this study found that, while the increase in intracellular calcium by CQ in nerve cell bodies dissociated from DRG is strictly dependent on TRPA1, TRPA1 is not required for the action potential elicited by CQ at the C-fiber nerve terminals [109]. These findings indicate that TRPA1 plays a distinct role at specific sites in the scratching response.
In addition to its relevance to GPCRs, TRPA1 has also been shown to interact with another ion channel, specifically the Nav1.7 channel. Methylglyoxal, an endogenous reactive carbonyl compound that plays a crucial role in the pathogenesis of diabetic neuropathy, has been found to activate both TRPA1 and Nav1.7 channels, leading to the induction of scratching behavior in response to methylglyoxal treatment [110]. Furthermore, the physiological role of extracellular microRNAs (miRNAs) as potential disease biomarkers has also been investigated in the context of itch signaling. Extracellular miR-711 has been shown to directly bind and activate TRPA1 in TRPA1-expressing heterologous cells and primary sensory neurons, resulting in scratching behavior in response to intradermal cheek injection, but not pain-related behavior. Interestingly, this effect was found to be independent of TLR7, a known modulator of itch sensation [111]. On the other hand, TLR7 agonist imiquimod (IQ) has been identified as another pruritogen via direct interaction with TRPA1, inducing itch-like behaviors in both zebrafish larvae and mice [112].
In TRPA1-deficient mice, scratching and general responses to AD seem diminished [113]. Additionally, both TRPA1 and TRPV1 channels are crucial for generating spontaneous scratching in a mouse ACD model [114]. A proinflammatory role for TRPA1 has been observed, evidenced by a suppressed cytokine response, epidermal thickening, and reductions in other features of inflammation [113]. Thus, the current understating of the pathology associated with TRPA1 in the skin requires further investigations. However, its involvement in pruriception, acting downstream of the GPCRs, makes it a potential drug target for alleviating the itch.
4.2. Sensory TRPV1 in Itch
TRPV1 is a subfamily of temperature-sensitive TRP channels activated by noxious temperatures greater than 43 °C, capsaicin, and low pH [115]. Furthermore, this channel can be directly and indirectly sensitized by various pro-inflammatory or pruritogenic agents, including histamine, proteases, sphingosine 1-phosphate (S1P), and IQ, which can endogenously mediate phosphorylation of intracellular domains by PKC, PKA, and other kinases [116,117]. The primary function of TRPV1 is the sensation of pain, with its expression mostly found in peripheral sensory nerves in the skin and central nerve endings in the DRG [118]. However, recent findings have reported TRPV1 distribution in other non-neuronal cells such as epidermal keratinocytes, T-cells, mast cells, leukocytes, macrophages, and sweat gland cells [119]. Therefore, when developing therapeutics for this channel, considering the impact on all populations of this channel distributed widely in various tissues and organs is essential.
TRPV1 is involved in histamine-dependent itch, which is facilitated by the co-expression of H1R signaling in sensory neurons [35], and histamine-induced scratching behaviors are abolished in TRPV1 knockout mice [36]. The molecular mechanism of the transduction of the histaminergic itch signal to evoke the scratching response has recently been discovered. It has been found that H1R directly binds to the deSUMOylated TRPV1 C terminus, facilitated by histamine treatment [120]. TRPV1 sensitization can also be induced by histamine-independent itch signaling, such as endogenous proteases that activate PAR-1 and PAR-4, which have been implicated in AD [121]. Thus, TRPV1 plays a critical role in itch signals associated with histamine-dependent and -independent itch.
IQ, an agonist of TLR7, also evokes itch-associated responses with the presence of TRPV1 [122]. Although research has found sex differences in TRPV1 and TRPA1 knockout mice regarding IQ-induced psoriatic dermatitis [123], these two channels may play a role in developing itch sensation. Lysophosphatidic acid (LPA) is another itch mediator associated with TRPV1 and TRPA1. Either TRPV1 or TRPA1 knockout mice show reduced LPA-induced DRG neuron activations and scratching responses after intradermal LPA administration [124]. Moreover, a low concentration of S1P evokes acute itch, whereas a high concentration causes pain and itch [125]. Studies have revealed that the co-expression of S1P receptor 3 (S1PR3) with TRPV1 leads to the induction of acute pain and heat hypersensitivity in response to S1P. However, when S1PR3 and TRPA1 are co-expressed, they mediate S1P-induced itch responses [120], suggesting that distinct mechanisms are involved in the signaling of itch and pain, and the interplay between TRPV1 and TRPA1 may play a role in the itch sensation.
Although the downstream signaling of histamine receptor subtypes H1R and H4R involves TRPV1, TRPA1 is also associated with histamine-induced pruritus transduction via H4R [126]. Periostin, a fasciclin extracellular matrix protein, potentially involves the activation of both TRPV1 and TRPA1 in the context of chronic allergic itch conditions [127]. This study revealed that periostin binds to the integrin αVβ3 in DRG sensory neurons, leading to the release of the neuropeptide NPPB due to TRPV1- and TRPA1-induced neuronal depolarization, which in turn contributes to the itch signal. In TRPV1 and TRPV4 knockout mice, cinnamaldehyde (CA)-induced scratching behavior was diminished, compared to wild-type mice, while TRPA1 knockout mice did not show the similar reduction in scratching behavior [128]. These findings suggest crosstalk between TRP channels and GPCRs, and the subsequent itch signaling transduction.
4.3. Sensory TRPV4 in Itch
TRPV4 is another temperature-sensitive TRP channel activated at 27 and 35 °C [88]. While TRPV4 is primarily expressed in keratinocytes, research has shown that the expression of TRPV4 in subsets of DRG sensory neurons is required for both CQ- and histamine-induced itch transmission [96]. In this study, the knockdown of TRPV4 in DRG neurons using intrathecal small interfering ribonucleic acid (siRNA) injection significantly reduced CQ or histamine-induced scratching behaviors compared to mice injected with negative control. Furthermore, the function of TRPV4 in complexes in DRG neurons and HEK293 cells depends on TRPV1, indicating that TRPV1 facilitates the TRPV4 responses to pruritogenic agents [96]. Thus, TRPV4 in sensory neurons plays a differential role in regulating itch signals.
4.4. Sensory TRPM8 in Itch
The activation of TRPM8 is induced by temperatures in the range of 8–28 °C and by natural compounds that can cause cooling sensations, such as menthone, menthol, and eucalyptol [129]. Cooling by applying cold water or ice has been commonly used to relieve itch, suggesting that TRPM8 activation can regulate itch transmission [130]. Moreover, cooling and menthol-induced TRPM8 activation can inhibit histamine-dependent and -independent itch pathways in neurons expressing TRPM8 [131]. Importantly, a TRPM8 agonist cooling compound, formed by combining with (1R,2S,5R)-N-(2-(2-pyridinyl)ethyl)-2-ispropyl-5-methylcyclohexancarboxamide and menthoxypropanediol, show a stronger activation than menthol [132]. The application of a lotion containing this compound ameliorated severe pruritus in a dry skin-associated chronic itch model, indicating that TRPM8 may play a role as an itch modulator. However, further research is required to identify a specific neural circuit and potential cellular mechanism for treating chronic itch.
4.5. Sensory TRPC3 and TRPC4 in Itch
TRPC channels are also calcium-permeable nonselective cation channels [133]. TRPC3 is particularly prevalent in DRG neurons, and its role in the histamine-independent itch pathway has been demonstrated through the reduction in scratching responses induced by TRPC3 agonist GSK1702934A, as well as endothelin-1 and SLIGRL in TRPC3 knockout mice [134], highlighting the role of TRPC3 in the histamine-independent itch pathway. Furthermore, research has demonstrated that TRPC3 is also expressed in MRGPRD-positive non-peptidergic C fiber nociceptors, where MRGPRD is a GPCR member D that mediates β-alanine-induced itch sensations [135]. However, the role of TRPC3 downstream of MRGPRD-positive neurons remains to be further investigated.
In DRG neurons, TRPC4 is co-expressed with serotonin receptor subtype 2B (HTR-2B) and mediates selective serotonin reuptake inhibitors (SSRIs)-evoked pruritus [136]. Sertraline, a commonly prescribed SSRI medication, elicited a robust itch response when administered subcutaneously. However, this response was significantly diminished in TRPC4 knockout mice without affecting the function of TRPA1 or TRPV1. Although TRPC4 antagonists, such as ML204, have been discovered [137], further research for its impact on itch responses is needed.
5. Antipruritic Compounds Targeting TRP Channels and Future Perspective
Several potential antipruritic compounds have been developed to target various TRP channels. Table 1 shows a list of published studies in which the effect of antipruritic compounds is attained through the modulation of TRP channels in diverse itch-related models. While some of these compounds are applied locally to the skin, others are delivered systemically. The expression of TRP channels is not limited to the skin or sensory neurons, as they are present in various tissues throughout the body. Therefore, systemically applied antipruritic compounds may have off-target effects on TRP channels in other physiological processes in non-itch-related tissues. For instance, the TRPV1 antagonist AMG-9810 was discontinued in the phase I clinical trial due to the side effect of hyperthermia, an abnormal rise in body temperature from a failure of the body’s heat-regulating mechanism [138]. Furthermore, the majority of antipruritic compounds have been investigated via restricted administration routes. Notably, some compounds (A-967079 and Resolvin D3) have demonstrated their efficacy through local and systemic administration (Figure 1).
As TRP channels have distinct functional roles in specific sites, global knockout studies may not provide tissue-specific information. Only studies on TRPV4 have demonstrated the effects of global or conditional knockout of TRPV4, including in skin keratinocytes, sensory neurons, and macrophages [19]. The use of global knockout studies to investigate the behavioral aspects of itch may be limited due to the dynamic nature of this process and the potential for the role of TRP channels to change over time. Therefore, to gain a more specific understanding of the tissue-specific role of TRP channels in itch, complementary approaches, such as tissue-specific knockouts and conditional knockouts, are considered.
Author Contributions
Writing–original draft and visualization, E.J.G.; Writing––review and editing, J.Y.L. and Y.H.K.; Supervision, C.-K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Bio and Medical Technology Development Program of the National Research Foundation of Korea (NRF-2022M3E5E8081191 to C.-K.P.) and by the National Research Foundation of Korea (NRF-2022R1F1A1076127 to J.Y.L.), both of which are funded by the Korean government.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Conflicts of Interest
The authors of this study have declared that they have no competing interests.
References
- Stander, S.; Weisshaar, E.; Mettang, T.; Szepietowski, J.C.; Carstens, E.; Ikoma, A.; Bergasa, N.V.; Gieler, U.; Misery, L.; Wallengren, J.; et al. Clinical classification of itch: A position paper of the International Forum for the Study of Itch. Acta Derm. Venereol. 2007, 87, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, J.D. Itch and pruritus: What are they, and how should itches be classified? Dermatol. Ther. 2005, 18, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Yosipovitch, G.; Greaves, M.W.; Schmelz, M. Itch. Lancet 2003, 361, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Twycross, R.; Greaves, M.W.; Handwerker, H.; Jones, E.A.; Libretto, S.E.; Szepietowski, J.C.; Zylicz, Z. Itch: Scratching more than the surface. QJM 2003, 96, 7–26. [Google Scholar] [CrossRef] [PubMed]
- Galatian, A.; Stearns, G.; Grau, R. Pruritus in connective tissue and other common systemic disease states. Cutis 2009, 84, 207–214. [Google Scholar]
- Binder, A.; Koroschetz, J.; Baron, R. Disease mechanisms in neuropathic itch. Nat. Clin. Pract. Neurol. 2008, 4, 329–337. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Samuel, L.S. Neuropathic and psychogenic itch. Dermatol. Ther. 2008, 21, 32–41. [Google Scholar] [CrossRef]
- Stander, S.; Zeidler, C.; Augustin, M.; Darsow, U.; Kremer, A.E.; Legat, F.J.; Koschmieder, S.; Kupfer, J.; Mettang, T.; Metz, M.; et al. S2k guideline: Diagnosis and treatment of chronic pruritus. J. Dtsch. Dermatol. Ges. 2022, 20, 1387–1402. [Google Scholar] [CrossRef]
- Song, J.; Xian, D.; Yang, L.; Xiong, X.; Lai, R.; Zhong, J. Pruritus: Progress toward Pathogenesis and Treatment. BioMed Res. Int. 2018, 2018, 9625936. [Google Scholar] [CrossRef]
- Graham, D.T.; Goodell, H.; Wolff, H.G. Neural mechanisms involved in itch, itchy skin, and tickle sensations. J. Clin. Investig. 1951, 30, 37–49. [Google Scholar] [CrossRef]
- Ikoma, A.; Steinhoff, M.; Stander, S.; Yosipovitch, G.; Schmelz, M. The neurobiology of itch. Nat. Rev. Neurosci. 2006, 7, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.G.; Zhao, Z.Q.; Meng, X.L.; Yin, J.; Liu, X.Y.; Chen, Z.F. Cellular basis of itch sensation. Science 2009, 325, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Q.; Liu, X.Y.; Jeffry, J.; Karunarathne, W.K.; Li, J.L.; Munanairi, A.; Zhou, X.Y.; Li, H.; Sun, Y.G.; Wan, L.; et al. Descending control of itch transmission by the serotonergic system via 5-HT1A-facilitated GRP-GRPR signaling. Neuron 2014, 84, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Dong, X. Itch mechanisms and circuits. Annu. Rev. Biophys. 2014, 43, 331–355. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Dong, X. Trp channels and itch. Semin. Immunopathol. 2016, 38, 293–307. [Google Scholar] [CrossRef]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. In Subcellular Biochemistry; Springer: Berlin/Heidelberg, Germany, 2018; Volume 87, pp. 141–165. [Google Scholar] [CrossRef]
- Han, L.; Ma, C.; Liu, Q.; Weng, H.J.; Cui, Y.; Tang, Z.; Kim, Y.; Nie, H.; Qu, L.; Patel, K.N.; et al. A subpopulation of nociceptors specifically linked to itch. Nat. Neurosci. 2013, 16, 174–182. [Google Scholar] [CrossRef]
- Bautista, D.M.; Wilson, S.R.; Hoon, M.A. Why we scratch an itch: The molecules, cells and circuits of itch. Nat. Neurosci. 2014, 17, 175–182. [Google Scholar] [CrossRef]
- Zhang, Q.; Henry, G.; Chen, Y. Emerging Role of Transient Receptor Potential Vanilloid 4 (TRPV4) Ion Channel in Acute and Chronic Itch. Int. J. Mol. Sci. 2021, 22, 7591. [Google Scholar] [CrossRef]
- Green, D.; Dong, X. The cell biology of acute itch. J. Cell Biol. 2016, 213, 155–161. [Google Scholar] [CrossRef]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289–301. [Google Scholar] [CrossRef]
- Rao, K.N.; Brown, M.A. Mast cells: Multifaceted immune cells with diverse roles in health and disease. Ann. N. Y. Acad. Sci. 2008, 1143, 83–104. [Google Scholar] [CrossRef] [PubMed]
- Ringkamp, M.; Schepers, R.J.; Shimada, S.G.; Johanek, L.M.; Hartke, T.V.; Borzan, J.; Shim, B.; LaMotte, R.H.; Meyer, R.A. A role for nociceptive, myelinated nerve fibers in itch sensation. J. Neurosci. 2011, 31, 14841–14849. [Google Scholar] [CrossRef] [PubMed]
- Lay, M.; Dong, X. Neural Mechanisms of Itch. Annu. Rev. Neurosci. 2020, 43, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Moniaga, C.S.; Tominaga, M.; Takamori, K. Mechanisms and Management of Itch in Dry Skin. Acta Derm. Venereol. 2020, 100, adv00024. [Google Scholar] [CrossRef] [PubMed]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Potenzieri, C.; Undem, B.J. Basic mechanisms of itch. Clin. Exp. Allergy 2012, 42, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Kittaka, H.; Tominaga, M. The molecular and cellular mechanisms of itch and the involvement of TRP channels in the peripheral sensory nervous system and skin. Allergol. Int. 2017, 66, 22–30. [Google Scholar] [CrossRef]
- Jeffry, J.; Kim, S.; Chen, Z.F. Itch signaling in the nervous system. Physiology 2011, 26, 286–292. [Google Scholar] [CrossRef]
- Papoiu, A.D.; Coghill, R.C.; Kraft, R.A.; Wang, H.; Yosipovitch, G. A tale of two itches. Common features and notable differences in brain activation evoked by cowhage and histamine induced itch. Neuroimage 2012, 59, 3611–3623. [Google Scholar] [CrossRef]
- Shirolkar, P.; Mishra, S.K. Role of TRP ion channels in pruritus. Neurosci. Lett. 2022, 768, 136379. [Google Scholar] [CrossRef]
- Hill, S.J.; Ganellin, C.R.; Timmerman, H.; Schwartz, J.C.; Shankley, N.P.; Young, J.M.; Schunack, W.; Levi, R.; Haas, H.L. International Union of Pharmacology. XIII. Classification of histamine receptors. Pharmacol. Rev. 1997, 49, 253–278. [Google Scholar] [PubMed]
- Kim, B.M.; Lee, S.H.; Shim, W.S.; Oh, U. Histamine-induced Ca2+ influx via the PLA2/lipoxygenase/TRPV1 pathway in rat sensory neurons. Neurosci. Lett. 2004, 361, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Han, S.K.; Mancino, V.; Simon, M.I. Phospholipase Cbeta 3 mediates the scratching response activated by the histamine H1 receptor on C-fiber nociceptive neurons. Neuron 2006, 52, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Shim, W.S.; Tak, M.H.; Lee, M.H.; Kim, M.; Kim, M.; Koo, J.Y.; Lee, C.H.; Kim, M.; Oh, U. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J. Neurosci. 2007, 27, 2331–2337. [Google Scholar] [CrossRef] [PubMed]
- Imamachi, N.; Park, G.H.; Lee, H.; Anderson, D.J.; Simon, M.I.; Basbaum, A.I.; Han, S.K. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11330–11335. [Google Scholar] [CrossRef]
- Guo, C.J.; Grabinski, N.S.; Liu, Q. Peripheral Mechanisms of Itch. J. Invest. Dermatol. 2022, 142, 31–41. [Google Scholar] [CrossRef]
- Mossner, R.; Lesch, K.P. Role of serotonin in the immune system and in neuroimmune interactions. Brain Behav. Immun. 1998, 12, 249–271. [Google Scholar] [CrossRef]
- Morita, T.; McClain, S.P.; Batia, L.M.; Pellegrino, M.; Wilson, S.R.; Kienzler, M.A.; Lyman, K.; Olsen, A.S.; Wong, J.F.; Stucky, C.L.; et al. HTR7 Mediates Serotonergic Acute and Chronic Itch. Neuron 2015, 87, 124–138. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Liu, T.; Xu, Z.Z.; Park, C.K.; Berta, T.; Ji, R.R. Toll-like receptor 7 mediates pruritus. Nat. Neurosci. 2010, 13, 1460–1462. [Google Scholar] [CrossRef]
- Liu, T.; Berta, T.; Xu, Z.Z.; Park, C.K.; Zhang, L.; Lu, N.; Liu, Q.; Liu, Y.; Gao, Y.J.; Liu, Y.C.; et al. TLR3 deficiency impairs spinal cord synaptic transmission, central sensitization, and pruritus in mice. J. Clin. Invest. 2012, 122, 2195–2207. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Lerner, E.A.; Carstens, E. Protease-activated receptors and itch. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 226, pp. 219–235. [Google Scholar] [CrossRef]
- Akiyama, T.; Merrill, A.W.; Zanotto, K.; Carstens, M.I.; Carstens, E. Scratching behavior and Fos expression in superficial dorsal horn elicited by protease-activated receptor agonists and other itch mediators in mice. J. Pharmacol. Exp. Ther. 2009, 329, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Neisius, U.; Ikoma, A.; Fartasch, M.; Heyer, G.; Skov, P.S.; Luger, T.A.; Schmelz, M. Proteinase-activated receptor-2 mediates itch: A novel pathway for pruritus in human skin. J. Neurosci. 2003, 23, 6176–6180. [Google Scholar] [CrossRef] [PubMed]
- Bohm, S.K.; Khitin, L.M.; Grady, E.F.; Aponte, G.; Payan, D.G.; Bunnett, N.W. Mechanisms of desensitization and resensitization of proteinase-activated receptor-2. J. Biol. Chem. 1996, 271, 22003–22016. [Google Scholar] [CrossRef] [PubMed]
- Kouzaki, H.; O’Grady, S.M.; Lawrence, C.B.; Kita, H. Proteases induce production of thymic stromal lymphopoietin by airway epithelial cells through protease-activated receptor-2. J. Immunol. 2009, 183, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Moniaga, C.S.; Jeong, S.K.; Egawa, G.; Nakajima, S.; Hara-Chikuma, M.; Jeon, J.E.; Lee, S.H.; Hibino, T.; Miyachi, Y.; Kabashima, K. Protease activity enhances production of thymic stromal lymphopoietin and basophil accumulation in flaky tail mice. Am. J. Pathol. 2013, 182, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Siracusa, M.C.; Saenz, S.A.; Noti, M.; Monticelli, L.A.; Sonnenberg, G.F.; Hepworth, M.R.; Van Voorhees, A.S.; Comeau, M.R.; Artis, D. TSLP elicits IL-33-independent innate lymphoid cell responses to promote skin inflammation. Sci. Transl. Med. 2013, 5, 170ra116. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, S.P.; Abrams, E.; Benson, A.; Fodeman, J.; Zheng, T. The role of thymic stromal lymphopoietin in the immunopathogenesis of atopic dermatitis. Clin. Exp. Allergy 2011, 41, 1515–1520. [Google Scholar] [CrossRef]
- Schmelz, M. Itch and pain. Neurosci. Biobehav. Rev. 2010, 34, 171–176. [Google Scholar] [CrossRef]
- Greaves, M.W.; Wall, P.D. Pathophysiology of itching. Lancet 1997, 349, 133. [Google Scholar] [CrossRef]
- McMahon, S.B.; Koltzenburg, M. Itching for an explanation. Trends Neurosci. 1992, 15, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Namer, B.; Carr, R.; Johanek, L.M.; Schmelz, M.; Handwerker, H.O.; Ringkamp, M. Separate peripheral pathways for pruritus in man. J. Neurophysiol. 2008, 100, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, M.; Hilliges, M.; Schmidt, R.; Orstavik, K.; Vahlquist, C.; Weidner, C.; Handwerker, H.O.; Torebjork, H.E. Active “itch fibers” in chronic pruritus. Neurology 2003, 61, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.G.; Chen, Z.F. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007, 448, 700–703. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.J.; Geng, Y.; et al. Sensory neuron-specific GPCR Mrgprs are itch receptors mediating chloroquine-induced pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Dong, X. Peripheral and Central Mechanisms of Itch. Neuron 2018, 98, 482–494. [Google Scholar] [CrossRef]
- Luo, J.; Feng, J.; Liu, S.; Walters, E.T.; Hu, H. Molecular and cellular mechanisms that initiate pain and itch. Cell Mol. Life Sci. 2015, 72, 3201–3223. [Google Scholar] [CrossRef]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef]
- Werfel, T.; Allam, J.P.; Biedermann, T.; Eyerich, K.; Gilles, S.; Guttman-Yassky, E.; Hoetzenecker, W.; Knol, E.; Simon, H.U.; Wollenberg, A.; et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 336–349. [Google Scholar] [CrossRef]
- Feld, M.; Garcia, R.; Buddenkotte, J.; Katayama, S.; Lewis, K.; Muirhead, G.; Hevezi, P.; Plesser, K.; Schrumpf, H.; Krjutskov, K.; et al. The pruritus- and TH2-associated cytokine IL-31 promotes growth of sensory nerves. J. Allergy Clin. Immunol. 2016, 138, 500–508.e24. [Google Scholar] [CrossRef]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef]
- Nassini, R.; Pedretti, P.; Moretto, N.; Fusi, C.; Carnini, C.; Facchinetti, F.; Viscomi, A.R.; Pisano, A.R.; Stokesberry, S.; Brunmark, C.; et al. Transient receptor potential ankyrin 1 channel localized to non-neuronal airway cells promotes non-neurogenic inflammation. PLoS ONE 2012, 7, e42454. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. TRPA1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef]
- Atoyan, R.; Shander, D.; Botchkareva, N.V. Non-neuronal expression of transient receptor potential type A1 (TRPA1) in human skin. J. Invest. Dermatol. 2009, 129, 2312–2315. [Google Scholar] [CrossRef] [PubMed]
- Earley, S.; Gonzales, A.L.; Crnich, R. Endothelium-dependent cerebral artery dilation mediated by TRPA1 and Ca2+-Activated K+ channels. Circ. Res. 2009, 104, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, I.; Gomes, P.; Aranake, S.; Shetty, M.; Karnik, P.; Damle, M.; Kuruganti, S.; Thorat, S.; Khairatkar-Joshi, N. Expression of functional TRPA1 receptor on human lung fibroblast and epithelial cells. J. Recept. Signal Transduct. 2011, 31, 350–358. [Google Scholar] [CrossRef] [PubMed]
- De Logu, F.; Li Puma, S.; Landini, L.; Portelli, F.; Innocenti, A.; de Araujo, D.S.M.; Janal, M.N.; Patacchini, R.; Bunnett, N.W.; Geppetti, P.; et al. Schwann cells expressing nociceptive channel TRPA1 orchestrate ethanol-evoked neuropathic pain in mice. J. Clin. Investig. 2019, 129, 5424–5441. [Google Scholar] [CrossRef]
- Guler, A.D.; Lee, H.; Iida, T.; Shimizu, I.; Tominaga, M.; Caterina, M. Heat-evoked activation of the ion channel, TRPV4. J. Neurosci. 2002, 22, 6408–6414. [Google Scholar] [CrossRef]
- Fujii, N.; Kenny, G.P.; McGarr, G.W.; Amano, T.; Honda, Y.; Kondo, N.; Nishiyasu, T. TRPV4 channel blockade does not modulate skin vasodilation and sweating during hyperthermia or cutaneous postocclusive reactive and thermal hyperemia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R563–R573. [Google Scholar] [CrossRef]
- Cai, S.; Fatherazi, S.; Presland, R.B.; Belton, C.M.; Izutsu, K.T. TRPC channel expression during calcium-induced differentiation of human gingival keratinocytes. J. Dermatol. Sci. 2005, 40, 21–28. [Google Scholar] [CrossRef]
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.B.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Lehen’kyi, V.; Beck, B.; Polakowska, R.; Charveron, M.; Bordat, P.; Skryma, R.; Prevarskaya, N. TRPV6 is a Ca2+ entry channel essential for Ca2+-induced differentiation of human keratinocytes. J. Biol. Chem. 2007, 282, 22582–22591. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Feng, J.; Luo, J.; Madison, M.; Hu, H. A Critical Role for TRP Channels in the Skin. In Neurobiology of TRP Channels; Emir, T.L.R., Ed.; Frontiers in Neuroscience: Boca Raton, FL, USA, 2017; pp. 95–111. [Google Scholar]
- Moqrich, A.; Hwang, S.W.; Earley, T.J.; Petrus, M.J.; Murray, A.N.; Spencer, K.S.; Andahazy, M.; Story, G.M.; Patapoutian, A. Impaired thermosensation in mice lacking TRPV3, a heat and camphor sensor in the skin. Science 2005, 307, 1468–1472. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ramsey, I.S.; Kotecha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef]
- Smith, G.D.; Gunthorpe, M.J.; Kelsell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.P.; Ooi, L.; et al. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef]
- Chung, M.K.; Lee, H.; Mizuno, A.; Suzuki, M.; Caterina, M.J. 2-aminoethoxydiphenyl borate activates and sensitizes the heat-gated ion channel TRPV3. J. Neurosci. 2004, 24, 5177–5182. [Google Scholar] [CrossRef]
- Asakawa, M.; Yoshioka, T.; Matsutani, T.; Hikita, I.; Suzuki, M.; Oshima, I.; Tsukahara, K.; Arimura, A.; Horikawa, T.; Hirasawa, T.; et al. Association of a mutation in TRPV3 with defective hair growth in rodents. J. Invest. Dermatol. 2006, 126, 2664–2672. [Google Scholar] [CrossRef]
- Yoshioka, T.; Hikita, I.; Asakawa, M.; Hirasawa, T.; Deguchi, M.; Matsutani, T.; Oku, H.; Horikawa, T.; Arimura, A. Spontaneous scratching behaviour in DS-Nh mice as a possible model for pruritus in atopic dermatitis. Immunology 2006, 118, 293–301. [Google Scholar] [CrossRef]
- Hong, J.; Buddenkotte, J.; Berger, T.G.; Steinhoff, M. Management of itch in atopic dermatitis. Semin. Cutan. Med. Surg. 2011, 30, 71–86. [Google Scholar] [CrossRef]
- Mack, M.R.; Kim, B.S. The Itch-Scratch Cycle: A Neuroimmune Perspective. Trends Immunol. 2018, 39, 980–991. [Google Scholar] [CrossRef]
- Imura, K.; Yoshioka, T.; Hirasawa, T.; Sakata, T. Role of TRPV3 in immune response to development of dermatitis. J. Inflamm. 2009, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Chen, Q.; Lee, M.; Cao, X.; Zhang, J.; Ma, D.; Chen, L.; Hu, X.; Wang, H.; Wang, X.; et al. Exome sequencing reveals mutations in TRPV3 as a cause of Olmsted syndrome. Am. J. Hum. Genet. 2012, 90, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Jin, J.; Hu, L.; Shen, D.; Dong, X.P.; Samie, M.A.; Knoff, J.; Eisinger, B.; Liu, M.L.; Huang, S.M.; et al. TRP channel regulates EGFR signaling in hair morphogenesis and skin barrier formation. Cell 2010, 141, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Munanairi, A.; Liu, X.Y.; Zhang, J.; Hu, L.; Hu, M.; Bu, D.; Liu, L.; Xie, Z.; Kim, B.S.; et al. PAR2 Mediates Itch via TRPV3 Signaling in Keratinocytes. J. Invest. Dermatol. 2020, 140, 1524–1532. [Google Scholar] [CrossRef]
- Shibasaki, K.; Sugio, S.; Takao, K.; Yamanaka, A.; Miyakawa, T.; Tominaga, M.; Ishizaki, Y. TRPV4 activation at the physiological temperature is a critical determinant of neuronal excitability and behavior. Pflügers Arch.-Eur. J. Physiol. 2015, 467, 2495–2507. [Google Scholar] [CrossRef]
- Sokabe, T.; Tominaga, M. The TRPV4 cation channel: A molecule linking skin temperature and barrier function. Commun. Integr. Biol. 2010, 3, 619–621. [Google Scholar] [CrossRef]
- Denda, M.; Sokabe, T.; Fukumi-Tominaga, T.; Tominaga, M. Effects of skin surface temperature on epidermal permeability barrier homeostasis. J. Investig. Dermatol. 2007, 127, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Sokabe, T.; Fukumi-Tominaga, T.; Yonemura, S.; Mizuno, A.; Tominaga, M. The TRPV4 channel contributes to intercellular junction formation in keratinocytes. J. Biol. Chem. 2010, 285, 18749–18758. [Google Scholar] [CrossRef]
- Kida, N.; Sokabe, T.; Kashio, M.; Haruna, K.; Mizuno, Y.; Suga, Y.; Nishikawa, K.; Kanamaru, A.; Hongo, M.; Oba, A.; et al. Importance of transient receptor potential vanilloid 4 (TRPV4) in epidermal barrier function in human skin keratinocytes. Pflugers Arch. 2012, 463, 715–725. [Google Scholar] [CrossRef]
- Akazawa, Y.; Yuki, T.; Yoshida, H.; Sugiyama, Y.; Inoue, S. Activation of TRPV4 strengthens the tight-junction barrier in human epidermal keratinocytes. Skin Pharmacol. Physiol. 2013, 26, 15–21. [Google Scholar] [CrossRef]
- Hidding, J.; Agelopoulos, K.; Pereira, M.P.; Conrad, H.; Hatt, H.; Lotts, T.; Osada, N.; Pogatzki-Zahn, E.; Schmelz, M.; Stander, S. Sensory Qualities Point to Different Structural and Functional Skin Patterns in Chronic Pruritus Patients. A Translational Explorative Study. Acta Derm. Venereol. 2019, 99, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Ivanov, M.; Nagamine, M.; Davoodi, A.; Carstens, M.I.; Ikoma, A.; Cevikbas, F.; Kempkes, C.; Buddenkotte, J.; Steinhoff, M.; et al. Involvement of TRPV4 in Serotonin-Evoked Scratching. J. Investig. Dermatol. 2016, 136, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Barry, D.M.; Liu, X.Y.; Yin, S.; Munanairi, A.; Meng, Q.T.; Cheng, W.; Mo, P.; Wan, L.; Liu, S.B.; et al. Facilitation of TRPV4 by TRPV1 is required for itch transmission in some sensory neuron populations. Sci. Signal. 2016, 9, ra71. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fang, Q.; Wang, Z.; Zhang, J.Y.; MacLeod, A.S.; Hall, R.P.; Liedtke, W.B. Transient Receptor Potential Vanilloid 4 Ion Channel Functions as a Pruriceptor in Epidermal Keratinocytes to Evoke Histaminergic Itch. J. Biol. Chem. 2016, 291, 10252–10262. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Z.L.; Yeo, M.; Zhang, Q.J.; Lopez-Romero, A.E.; Ding, H.P.; Zhang, X.; Zeng, Q.; Morales-Lazaro, S.L.; Moore, C.; et al. Epithelia-Sensory Neuron Cross Talk Underlies Cholestatic Itch Induced by Lysophosphatidylcholine. Gastroenterology 2021, 161, 301–317.e16. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Feng, J.; Yu, G.; Yang, P.; Mack, M.R.; Du, J.; Yu, W.; Qian, A.; Zhang, Y.; Liu, S.; et al. Transient receptor potential vanilloid 4-expressing macrophages and keratinocytes contribute differentially to allergic and nonallergic chronic itch. J. Allergy Clin. Immunol. 2018, 141, 608–619.e7. [Google Scholar] [CrossRef]
- Dutta, B.; Arya, R.K.; Goswami, R.; Alharbi, M.O.; Sharma, S.; Rahaman, S.O. Role of macrophage TRPV4 in inflammation. Lab. Invest. 2020, 100, 178–185. [Google Scholar] [CrossRef]
- Reddy, V.B.; Iuga, A.O.; Shimada, S.G.; LaMotte, R.H.; Lerner, E.A. Cowhage-evoked itch is mediated by a novel cysteine protease: A ligand of protease-activated receptors. J. Neurosci. 2008, 28, 4331–4335. [Google Scholar] [CrossRef]
- Zygmunt, P.M.; Hogestatt, E.D. Trpa1. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 583–630. [Google Scholar] [CrossRef]
- Oh, M.H.; Oh, S.Y.; Lu, J.; Lou, H.; Myers, A.C.; Zhu, Z.; Zheng, T. TRPA1-dependent pruritus in IL-13-induced chronic atopic dermatitis. J. Immunol. 2013, 191, 5371–5382. [Google Scholar] [CrossRef]
- Wilson, S.R.; Gerhold, K.A.; Bifolck-Fisher, A.; Liu, Q.; Patel, K.N.; Dong, X.; Bautista, D.M. TRPA1 is required for histamine-independent, Mas-related G protein-coupled receptor-mediated itch. Nat. Neurosci. 2011, 14, 595–602. [Google Scholar] [CrossRef]
- Liu, T.; Ji, R.R. Oxidative stress induces itch via activation of transient receptor potential subtype ankyrin 1 in mice. Neurosci. Bull. 2012, 28, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Vong, C.T.; Quek, S.; Cheong, J.; Awal, S.; Gentry, C.; Aubdool, A.A.; Liang, L.; Bodkin, J.V.; Bevan, S.; et al. Superoxide generation and leukocyte accumulation: Key elements in the mediation of leukotriene B4-induced itch by transient receptor potential ankyrin 1 and transient receptor potential vanilloid 1. FASEB J. 2013, 27, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; The, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef]
- Lieu, T.; Jayaweera, G.; Zhao, P.; Poole, D.P.; Jensen, D.; Grace, M.; McIntyre, P.; Bron, R.; Wilson, Y.M.; Krappitz, M.; et al. The bile acid receptor TGR5 activates the TRPA1 channel to induce itch in mice. Gastroenterology 2014, 147, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Ru, F.; Sun, H.; Jurcakova, D.; Herbstsomer, R.A.; Meixong, J.; Dong, X.; Undem, B.J. Mechanisms of pruritogen-induced activation of itch nerves in isolated mouse skin. J. Physiol. 2017, 595, 3651–3666. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.X.; Feng, Y.; Liu, D.; Wang, Z.H.; Zhang, J.T.; Chen, L.H.; Su, C.J.; Wang, B.; Huang, Y.; Ji, R.R.; et al. The role of Na(v)1.7 and methylglyoxal-mediated activation of TRPA1 in itch and hypoalgesia in a murine model of type 1 diabetes. Theranostics 2019, 9, 4287–4307. [Google Scholar] [CrossRef]
- Han, Q.; Liu, D.; Convertino, M.; Wang, Z.; Jiang, C.; Kim, Y.H.; Luo, X.; Zhang, X.; Nackley, A.; Dokholyan, N.V.; et al. miRNA-711 Binds and Activates TRPA1 Extracellularly to Evoke Acute and Chronic Pruritus. Neuron 2018, 99, 449–463.e6. [Google Scholar] [CrossRef] [PubMed]
- Esancy, K.; Condon, L.; Feng, J.; Kimball, C.; Curtright, A.; Dhaka, A. A zebrafish and mouse model for selective pruritus via direct activation of TRPA1. Elife 2018, 7, e32036. [Google Scholar] [CrossRef]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef]
- Feng, J.; Yang, P.; Mack, M.R.; Dryn, D.; Luo, J.; Gong, X.; Liu, S.; Oetjen, L.K.; Zholos, A.V.; Mei, Z.; et al. Sensory TRP channels contribute differentially to skin inflammation and persistent itch. Nat. Commun. 2017, 8, 980. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Bhave, G.; Hu, H.J.; Glauner, K.S.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau, R.W.t. Protein kinase C phosphorylation sensitizes but does not activate the capsaicin receptor transient receptor potential vanilloid 1 (TRPV1). Proc. Natl. Acad. Sci. USA 2003, 100, 12480–12485. [Google Scholar] [CrossRef]
- Varga, A.; Bolcskei, K.; Szoke, E.; Almasi, R.; Czeh, G.; Szolcsanyi, J.; Petho, G. Relative roles of protein kinase A and protein kinase C in modulation of transient receptor potential vanilloid type 1 receptor responsiveness in rat sensory neurons in vitro and peripheral nociceptors in vivo. Neuroscience 2006, 140, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Bagood, M.D.; Isseroff, R.R. TRPV1: Role in Skin and Skin Diseases and Potential Target for Improving Wound Healing. Int. J. Mol. Sci. 2021, 22, 6135. [Google Scholar] [CrossRef] [PubMed]
- Bevan, S.; Quallo, T.; Andersson, D.A. Trpv1. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 207–245. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, R.; Weng, W.; Zhang, H.; Wang, Y.; Guo, R.; Gu, X.; Yang, Y.; Yang, F.; Zhou, A.; et al. TRPV1 SUMOylation suppresses itch by inhibiting TRPV1 interaction with H1 receptors. Cell Rep. 2022, 39, 110972. [Google Scholar] [CrossRef]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhe, V.; Plee-Gautier, E.; Carre, J.L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: Pro-inflammatory response induced by their activation and their sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Park, G.H.; Kim, D.; Lee, J.; Min, H.; Wall, E.; Lee, C.J.; Simon, M.I.; Lee, S.J.; Han, S.K. Analysis of cellular and behavioral responses to imiquimod reveals a unique itch pathway in transient receptor potential vanilloid 1 (TRPV1)-expressing neurons. Proc. Natl. Acad. Sci. USA 2011, 108, 3371–3376. [Google Scholar] [CrossRef]
- Follansbee, T.; Zhou, Y.; Wu, X.; Delahanty, J.; Nguyen, A.; Domocos, D.; Carstens, M.I.; Hwang, S.T.; Carstens, E. Signs of chronic itch in the mouse imiquimod model of psoriasiform dermatitis: Sex differences and roles of TRPV1 and TRPA1. Itch 2019, 4, e25. [Google Scholar] [CrossRef]
- Kittaka, H.; Uchida, K.; Fukuta, N.; Tominaga, M. Lysophosphatidic acid-induced itch is mediated by signalling of LPA5 receptor, phospholipase D and TRPA1/TRPV1. J. Physiol. 2017, 595, 2681–2698. [Google Scholar] [CrossRef]
- Hill, R.Z.; Hoffman, B.U.; Morita, T.; Campos, S.M.; Lumpkin, E.A.; Brem, R.B.; Bautista, D.M. The signaling lipid sphingosine 1-phosphate regulates mechanical pain. Elife 2018, 7, e33285. [Google Scholar] [CrossRef]
- Wilzopolski, J.; Kietzmann, M.; Mishra, S.K.; Stark, H.; Baumer, W.; Rossbach, K. TRPV1 and TRPA1 Channels Are Both Involved Downstream of Histamine-Induced Itch. Biomolecules 2021, 11, 1166. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Wheeler, J.J.; Pitake, S.; Ding, H.; Jiang, C.; Fukuyama, T.; Paps, J.S.; Ralph, P.; Coyne, J.; Parkington, M.; et al. Periostin Activation of Integrin Receptors on Sensory Neurons Induces Allergic Itch. Cell Rep. 2020, 31, 107472. [Google Scholar] [CrossRef] [PubMed]
- Domocos, D.; Follansbee, T.; Nguyen, A.; Nguyen, T.; Carstens, M.I.; Carstens, E. Cinnamaldehyde elicits itch behavior via TRPV1 and TRPV4 but not TRPA1. Itch 2020, 5, e36. [Google Scholar] [CrossRef] [PubMed]
- Raddatz, N.; Castillo, J.P.; Gonzalez, C.; Alvarez, O.; Latorre, R. Temperature and voltage coupling to channel opening in transient receptor potential melastatin 8 (TRPM8). J. Biol. Chem. 2014, 289, 35438–35454. [Google Scholar] [CrossRef]
- Liu, B.; Jordt, S.E. Cooling the Itch via TRPM8. J. Invest. Dermatol. 2018, 138, 1254–1256. [Google Scholar] [CrossRef]
- Palkar, R.; Ongun, S.; Catich, E.; Li, N.; Borad, N.; Sarkisian, A.; McKemy, D.D. Cooling Relief of Acute and Chronic Itch Requires TRPM8 Channels and Neurons. J. Invest. Dermatol. 2018, 138, 1391–1399. [Google Scholar] [CrossRef]
- Stander, S.; Augustin, M.; Roggenkamp, D.; Blome, C.; Heitkemper, T.; Worthmann, A.C.; Neufang, G. Novel TRPM8 agonist cooling compound against chronic itch: Results from a randomized, double-blind, controlled, pilot study in dry skin. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1064–1068. [Google Scholar] [CrossRef]
- Vazquez, G.; Wedel, B.J.; Aziz, O.; Trebak, M.; Putney, J.W., Jr. The mammalian TRPC cation channels. Biochim. Biophys. Acta 2004, 1742, 21–36. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Limjunyawong, N.; Narang, C.; Jamaldeen, H.; Yu, S.; Patiram, S.; Nie, H.; Caterina, M.J.; Dong, X.; et al. Sensory neuron-expressed TRPC3 mediates acute and chronic itch. Pain 2023, 164, 98–110. [Google Scholar] [CrossRef]
- Liu, Q.; Sikand, P.; Ma, C.; Tang, Z.; Han, L.; Li, Z.; Sun, S.; LaMotte, R.H.; Dong, X. Mechanisms of itch evoked by beta-alanine. J. Neurosci. 2012, 32, 14532–14537. [Google Scholar] [CrossRef]
- Lee, S.H.; Cho, P.S.; Tonello, R.; Lee, H.K.; Jang, J.H.; Park, G.Y.; Hwang, S.W.; Park, C.K.; Jung, S.J.; Berta, T. Peripheral serotonin receptor 2B and transient receptor potential channel 4 mediate pruritus to serotonergic antidepressants in mice. J. Allergy Clin. Immunol. 2018, 142, 1349–1352.e16. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Shi, J.; Zhu, Y.; Kustov, M.; Tian, J.B.; Stevens, A.; Wu, M.; Xu, J.; Long, S.; Yang, P.; et al. Identification of ML204, a novel potent antagonist that selectively modulates native TRPC4/C5 ion channels. J. Biol. Chem. 2011, 286, 33436–33446. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Treanor, J.J.; Garami, A.; Fang, L.; Surapaneni, S.; Akrami, A.; Alvarez, F.; Bak, A.; Darling, M.; Gore, A.; et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain 2008, 136, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Chen, C.; Zhou, W.; Ma, X.; Pu, X.; Zeng, Y.; Zhou, W.; Lv, F. TRPA1 deficiency alleviates inflammation of atopic dermatitis by reducing macrophage infiltration. Life Sci. 2021, 266, 118906. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.S.; Otsuka, A.; Yamamoto, Y.; Nonomura, Y.; Nakashima, C.; Kitayama, N.; Usui, K.; Honda, T.; Kabashima, K. TRPA1 channel participates in tacrolimus-induced pruritus in a chronic contact hypersensitivity murine model. J. Dermatol. Sci. 2018, 89, 207–209. [Google Scholar] [CrossRef]
- Lee, S.H.; Tonello, R.; Im, S.T.; Jeon, H.; Park, J.; Ford, Z.; Davidson, S.; Kim, Y.H.; Park, C.K.; Berta, T. Resolvin D3 controls mouse and human TRPV1-positive neurons and preclinical progression of psoriasis. Theranostics 2020, 10, 12111–12126. [Google Scholar] [CrossRef]
- Yun, J.W.; Seo, J.A.; Jang, W.H.; Koh, H.J.; Bae, I.H.; Park, Y.H.; Lim, K.M. Antipruritic effects of TRPV1 antagonist in murine atopic dermatitis and itching models. J. Investig. Dermatol. 2011, 131, 1576–1579. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, C.S.; Bae, I.H.; Choi, J.K.; Park, Y.H.; Park, M. A novel, topical, nonsteroidal, TRPV1 antagonist, PAC-14028 cream improves skin barrier function and exerts anti-inflammatory action through modulating epidermal differentiation markers and suppressing Th2 cytokines in atopic dermatitis. J. Dermatol. Sci. 2018, 91, 184–194. [Google Scholar] [CrossRef]
- Park, C.W.; Kim, B.J.; Lee, Y.W.; Won, C.; Park, C.O.; Chung, B.Y.; Lee, D.H.; Jung, K.; Nam, H.J.; Choi, G.; et al. Asivatrep, a TRPV1 antagonist, for the topical treatment of atopic dermatitis: Phase 3, randomized, vehicle-controlled study (CAPTAIN-AD). J. Allergy Clin. Immunol. 2022, 149, 1340–1347.e4. [Google Scholar] [CrossRef]
- Metz, M.; Krause, K.; Maurer, M.; Magerl, M. Treatment of notalgia paraesthetica with an 8% capsaicin patch. Br. J. Dermatol. 2011, 165, 1359–1361. [Google Scholar] [CrossRef]
- Zeidler, C.; Luling, H.; Dieckhofer, A.; Osada, N.; Schedel, F.; Steinke, S.; Augustin, M.; Stander, S. Capsaicin 8% cutaneous patch: A promising treatment for brachioradial pruritus? Br. J. Dermatol. 2015, 172, 1669–1671. [Google Scholar] [CrossRef]
- Gibson, R.A.; Robertson, J.; Mistry, H.; McCallum, S.; Fernando, D.; Wyres, M.; Yosipovitch, G. A randomised trial evaluating the effects of the TRPV1 antagonist SB705498 on pruritus induced by histamine, and cowhage challenge in healthy volunteers. PLoS ONE 2014, 9, e100610. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Luo, A.; Kamau, P.M.; Takomthong, P.; Hu, J.; Boonyarat, C.; Luo, L.; Lai, R. A plant-derived TRPV3 inhibitor suppresses pain and itch. Br. J. Pharmacol. 2021, 178, 1669–1683. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Sun, L.L.; Qi, H.; Gao, Q.; Wang, G.X.; Wei, N.N.; Wang, K. Antipruritic Effect of Natural Coumarin Osthole through Selective Inhibition of Thermosensitive TRPV3 Channel in the Skin. Mol. Pharmacol. 2018, 94, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, X.; Qi, H.; Ma, Q.; Zhou, Q.; Wang, W.; Wang, K. Pharmacological Inhibition of the Temperature-Sensitive and Ca2+-Permeable Transient Receptor Potential Vanilloid TRPV3 Channel by Natural Forsythoside B Attenuates Pruritus and Cytotoxicity of Keratinocytes. J. Pharmacol. Exp. Ther. 2019, 368, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Qi, H.; Wu, H.; Qu, Y.; Wang, K. Anti-pruritic and anti-inflammatory effects of natural verbascoside through selective inhibition of temperature-sensitive Ca2+-permeable TRPV3 channel. J. Dermatol. Sci. 2020, 97, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Shi, Y.; Wu, H.; Niu, C.; Sun, X.; Wang, K. Inhibition of temperature-sensitive TRPV3 channel by two natural isochlorogenic acid isomers for alleviation of dermatitis and chronic pruritus. Acta Pharm. Sin. B 2022, 12, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, L.; Jiao, K.; Xue, C.; Tang, Q.; Jiang, S.; Ren, Y.; Chen, H.; El-Aziz, T.M.A.; Abdelazeem, K.N.M.; et al. Scutellarein attenuates atopic dermatitis by selectively inhibiting transient receptor potential vanilloid 3 channels. Br. J. Pharmacol. 2022, 179, 4792–4808. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; Wei, X.; Hu, J.; Ping, C.; Gao, Y.; Xie, C.; Wang, P.; Cao, P.; Cao, Z.; et al. Therapeutic inhibition of keratinocyte TRPV3 sensory channel by local anesthetic dyclonine. Elife 2021, 10, e68128. [Google Scholar] [CrossRef]
- Fan, J.; Hu, L.; Yue, Z.; Liao, D.; Guo, F.; Ke, H.; Jiang, D.; Yang, Y.; Lei, X. Structural basis of TRPV3 inhibition by an antagonist. Nat. Chem. Biol. 2023, 19, 81–90. [Google Scholar] [CrossRef]
- Qin, Z.; Xiang, L.; Zheng, S.; Zhao, Y.; Qin, Y.; Zhang, L.; Zhou, L. Vitexin inhibits pain and itch behavior via modulating TRPV4 activity in mice. Biomed. Pharmacother. 2023, 165, 115101. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Ye, F.; Ju, Y.; Wang, D.; Chen, J.; Zhang, X.; Yin, Z.; Wang, C.; Yang, Y.; Zhu, C.; et al. Cimifugin relieves pruritus in psoriasis by inhibiting TRPV4. Cell Calcium 2021, 97, 102429. [Google Scholar] [CrossRef] [PubMed]
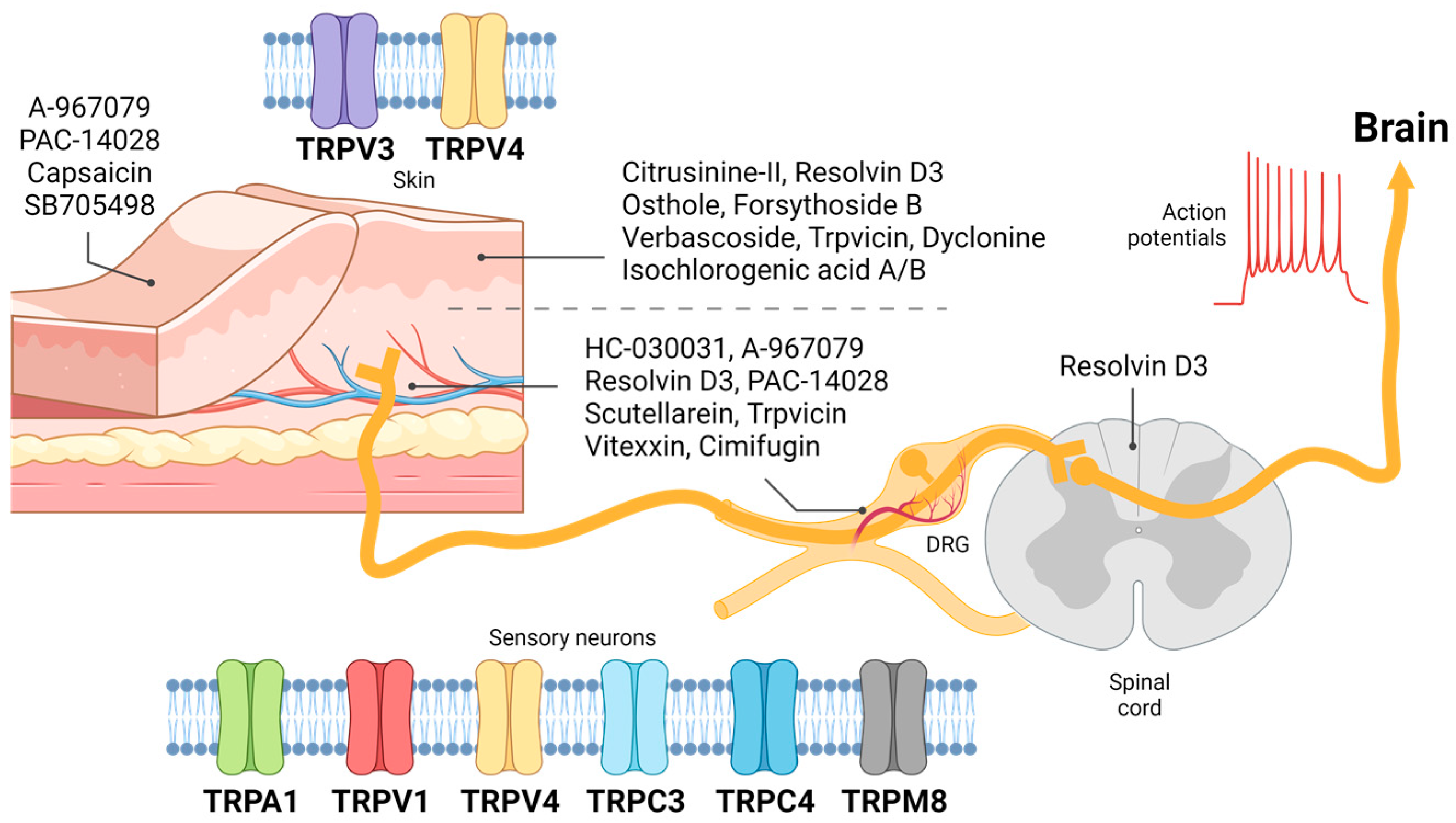
Figure 1.
Expression of transient receptor potential (TRP) receptors in skin keratinocytes and sensory neurons concerning pharmaceutical administration routes. The expression of TRPV3 and TRPV4 has been observed in the skin, where the efficacy of TRPA1 antagonist A-967079, TRPV1 antagonist PAC-14028 and SB705498, and TRPV1 agonist capsaicin has been examined. TRPV3 antagonist citrusinine-II, osthole, forsythoside B, verbascoside, isochlorogenic acid A and B, dyclonine, trpvicin, and TRPV1 antagonist resolvin D3 are locally administered via intradermal or transdermal injection. Sensory neurons express TRPA1, TRPV1, TRPV4, TRPC3, TRPC4, and TRPM8. Systemic deliveries of TRPA1 antagonists HC-030031 and A-967079, TRPV1 antagonists resolvin D3 and PAC-14028, TRPV3 antagonists scutellarein and trpvicin, and TRPV4 antagonists vitexin and cimifugin have been investigated for their potential antipruritic effects. These compounds modulate the transmission of itch signals to the brain.
Figure 1.
Expression of transient receptor potential (TRP) receptors in skin keratinocytes and sensory neurons concerning pharmaceutical administration routes. The expression of TRPV3 and TRPV4 has been observed in the skin, where the efficacy of TRPA1 antagonist A-967079, TRPV1 antagonist PAC-14028 and SB705498, and TRPV1 agonist capsaicin has been examined. TRPV3 antagonist citrusinine-II, osthole, forsythoside B, verbascoside, isochlorogenic acid A and B, dyclonine, trpvicin, and TRPV1 antagonist resolvin D3 are locally administered via intradermal or transdermal injection. Sensory neurons express TRPA1, TRPV1, TRPV4, TRPC3, TRPC4, and TRPM8. Systemic deliveries of TRPA1 antagonists HC-030031 and A-967079, TRPV1 antagonists resolvin D3 and PAC-14028, TRPV3 antagonists scutellarein and trpvicin, and TRPV4 antagonists vitexin and cimifugin have been investigated for their potential antipruritic effects. These compounds modulate the transmission of itch signals to the brain.


{kind=link}
Table 1.
Antipruritic compounds through TRP channels in various itch-related models.
| Targets | Pharmaceuticals | Models | Route | Ref. |
|---|---|---|---|---|
| TRPA1 | HC-030031 (antagonist) | DNCB-induced AD in mice | Intraperitoneal (100 mg/kg) | [139] |
| Oxazolone-induced chronic dermatitis in mice | Intraperitoneal (60 mg/kg) | [113] | ||
| A-967079 (antagonist) | Oxazolone-induced chronic dermatitis in mice | Intraperitoneal (100 mg/kg) | ||
| Tacrolimus-induced pruritus in chronic contact hypersensitivity mice | Topical (30 mg/kg) | [140] | ||
| TRPV1 | Resolvin D3 (antagonist) | Imiquimod-induced spontaneous scratching and alloknesis in mice | Intraperitoneal (2.8 mg/kg) Intradermal (100 ng/100 µL) Intrathecal (10 ng/10 µL) | [141] |
| PAC-14028 (antagonist) | Df extract-induced AD in mice | Oral (10–30 mg/kg) | [142] | |
| Oxazolone-induced chronic dermatitis in mice | Topical (50 µL of 1% cream) | [143] | ||
| Mild to moderate AD in human | Topical (1% cream) | [144] | ||
| Capsaicin (agonist) | Notalgia paraesthetica in human | Topical (8% patch) | [145] | |
| Brachioradial pruritus in human | [146] | |||
| SB705498 (antagonist) | Histamine-induced pruritus in human | Topical (3% cream) | [147] | |
| TRPV3 | Citrusinine-II (antagonist) | AEW- and histamine-induced scratching in mice | Intradermal, transdermal (5–10 µM/50 µL) | [148] |
| Histamine-induced pruritus in mice | Intradermal, transdermal (10 µM/50 µL) | |||
| Osthole (antagonist) | AEW- and histamine-induced scratching in mice | Intradermal (30–300 nM/50 µL) | [149] | |
| Histamine-induced pruritus in mice | Intradermal (300 nM/50 µL) | |||
| Forsythoside B (antagonist) | AEW- and histamine-induced scratching in mice | Intradermal (3–30 µM/50 µL) | [150] | |
| Histamine-induced pruritus in mice | Intradermal (0.3–30 µM/50 µL) | |||
| Carvacrol-induced pruritus in mice | Intradermal (30–300 µM/50 µL) | |||
| Verbascoside (antagonist) | Carvacrol-induced pruritus in mice | Intradermal (300 µM/50 µL) | [151] | |
| Isochlorogenic acid A (antagonist) | Carvacrol-induced pruritus in mice | Transdermal (1 mM/50 µL) | [152] | |
| Isochlorogenic acid B (antagonist) | Transdermal (1 mM/50 µL) | |||
| Scutellarein (antagonist) | Carvacrol-induced pruritus in mice | Subcutaneous (0.2–0.5 mg/kg) | [153] | |
| 2,4-dinitrofluorobenzene-induceddermatitis and pruritus in mice | Subcutaneous (0.2–0.5 mg/kg) | |||
| Dyclonine (antagonist) | Carvacrol-induced pruritus in mice | Intradermal (10–50 µM/50 µL) | [154] | |
| Trpvicin (antagonist) | SLIGRL-induced pruritus in mice | Intradermal (10–100 µM/50 µL) | [155] | |
| Calcipotriol-induced pruritus in mice | Oral (100 mg/kg) | |||
| TRPV4 | Vitexin (antagonist) | Histamine-, C48/80-, chloroquine-, and BAM8-22-induced acute, and dry-skin-induced chronic itch in mice | Intravenous (7.5 mg/kg) | [156] |
| AEW-induced dry skin causing chronic itch in mice | Intraperitoneal (7.5 mg/kg) | |||
| Cimifugin (antagonist) | GSK101-induced acute and Imiquimod-induced chronic itch in mice | Intragastric (75 mg/kg, 100 µL) | [157] |
DNCB, 2,4-dinitrochlorbenzene; AD, atopic dermatitis; Df, dermatophagoides farina; NP, notalgia paraesthetica; BRP, brachioradial pruritus; AEW, acetone–ether–water; TRPV, transient receptor potential vanilloid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Go, E.J.; Lee, J.Y.; Kim, Y.H.; Park, C.-K. Site-Specific Transient Receptor Potential Channel Mechanisms and Their Characteristics for Targeted Chronic Itch Treatment. Biomolecules 2024, 14, 107. https://doi.org/10.3390/biom14010107
AMA Style
Go EJ, Lee JY, Kim YH, Park C-K. Site-Specific Transient Receptor Potential Channel Mechanisms and Their Characteristics for Targeted Chronic Itch Treatment. Biomolecules. 2024; 14(1):107. https://doi.org/10.3390/biom14010107
Chicago/Turabian StyleGo, Eun Jin, Ji Yeon Lee, Yong Ho Kim, and Chul-Kyu Park. 2024. "Site-Specific Transient Receptor Potential Channel Mechanisms and Their Characteristics for Targeted Chronic Itch Treatment" Biomolecules 14, no. 1: 107. https://doi.org/10.3390/biom14010107
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.