
Do Media Extracellular Vesicles and Extracellular Vesicles Bound to the Extracellular Matrix Represent Distinct Types of Vesicles?

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Discovery of Matrix Vesicles
3. Discovery of Media Vesicles
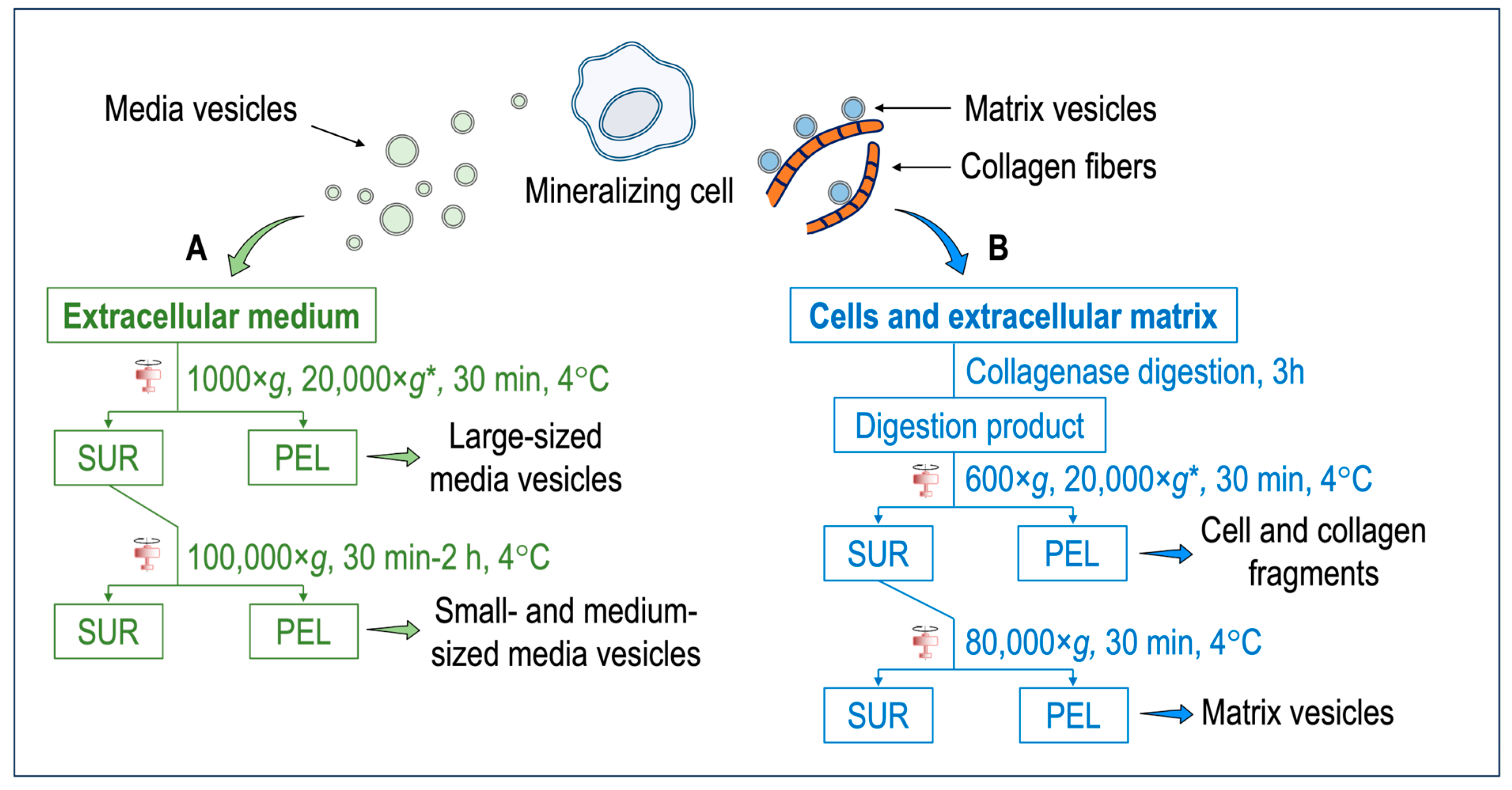
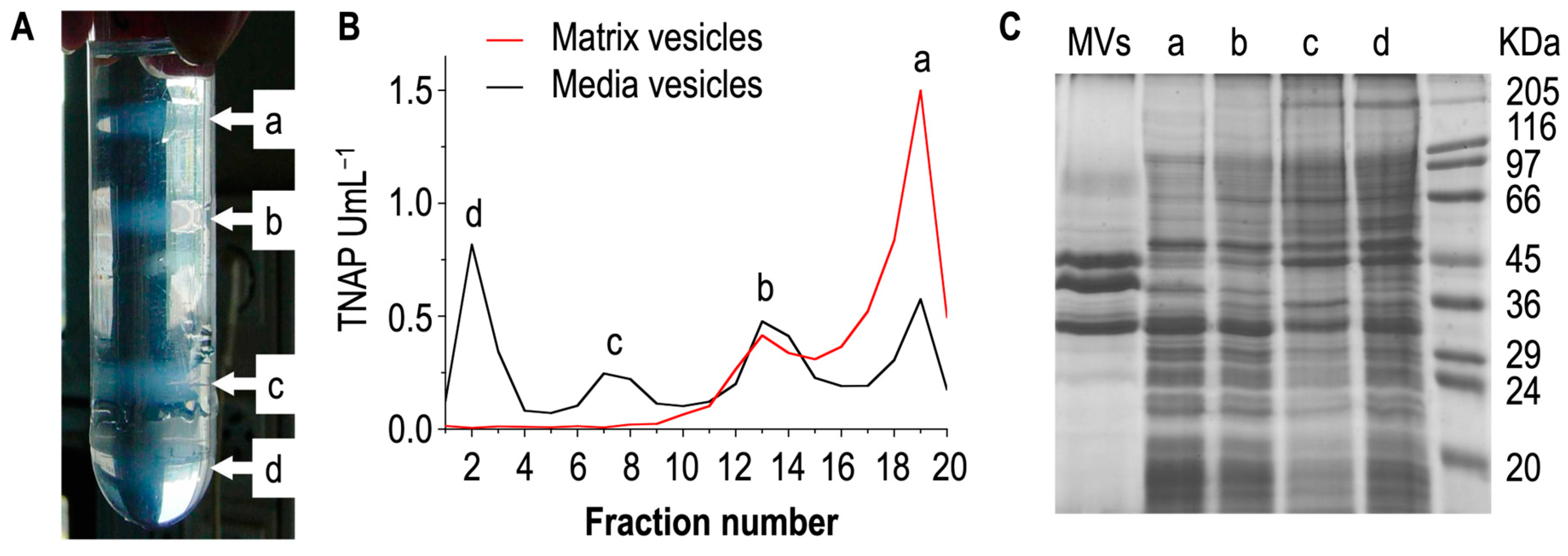
4. How to Differentiate Matrix Vesicles and Media Vesicles

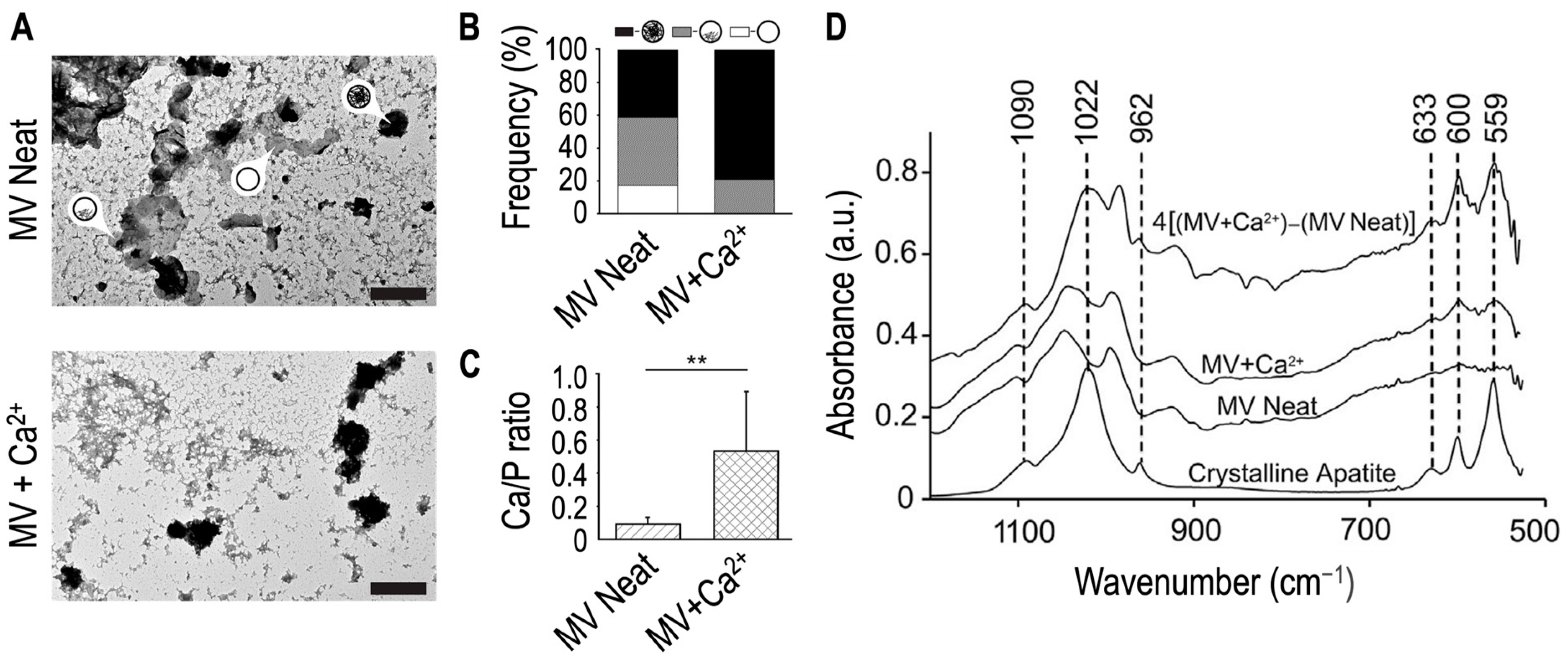
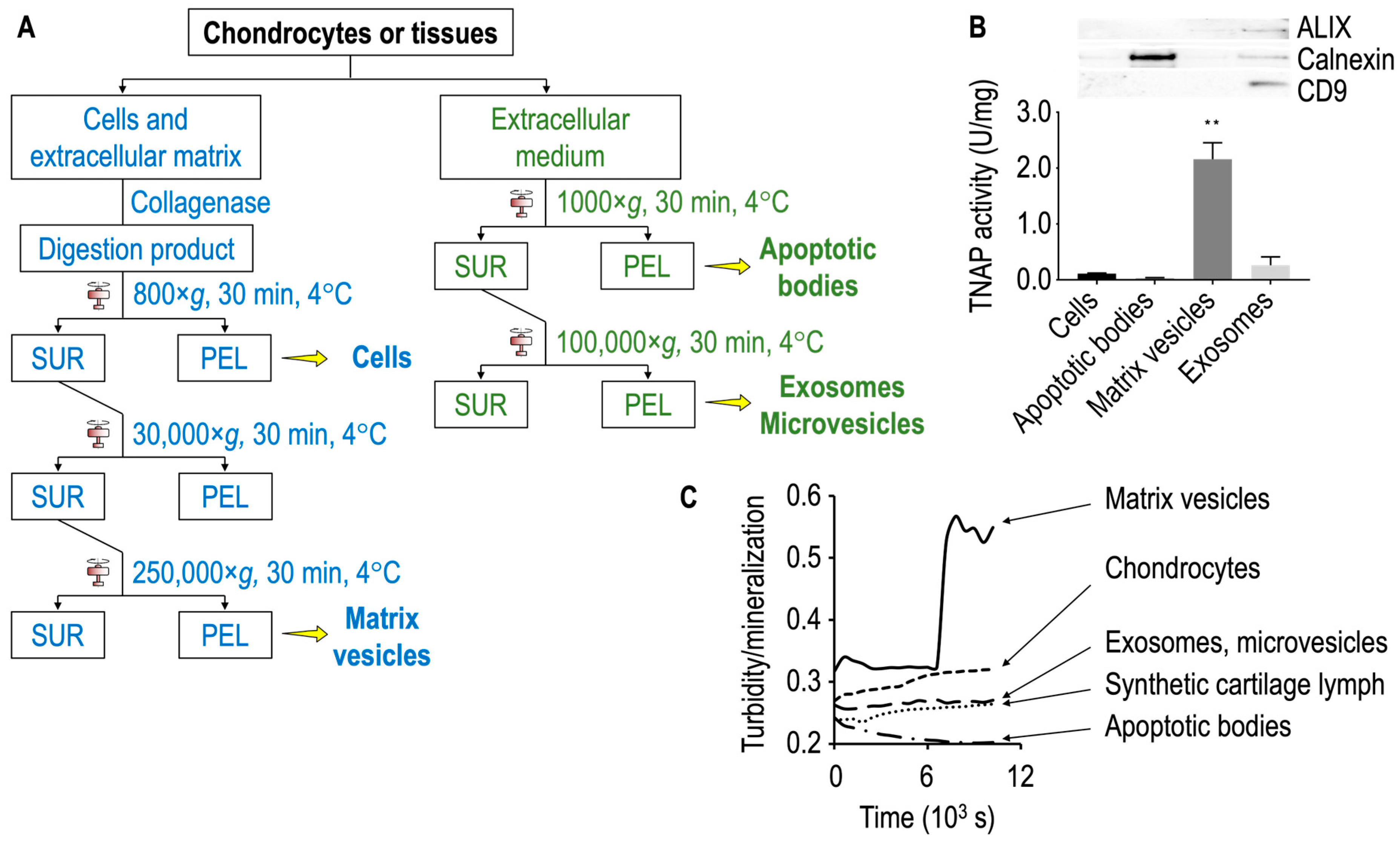
5. Matrix Vesicles and Media Vesicles from Growth Plate Cartilage and Hypertrophic Chondrocytes
6. Matrix Vesicles and Media Vesicles from Differentiated Osteoblasts
7. Matrix Vesicles and Media Vesicles from Smooth Muscle Cells
8. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, C.M.; Loyer, X.; Rautou, P.-E.; Amabile, N. Extracellular Vesicles in Coronary Artery Disease. Nat. Rev. Cardiol. 2017, 14, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Atukorala, I.; Mathivanan, S. Biogenesis of Extracellular Vesicles. Subcell. Biochem. 2021, 97, 19–43. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of Secretion and Uptake of Exosomes and Other Extracellular Vesicles for Cell-to-Cell Communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Stahl, P.D.; Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles, Integrators of Homeostasis. Physiology 2019, 34, 169–177. [Google Scholar] [CrossRef]
- Szabo, G.; Momen-Heravi, F. Extracellular Vesicles in Liver Disease and Potential as Biomarkers and Therapeutic Targets. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 455–466. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445.e18. [Google Scholar] [CrossRef]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of Distinct Nanoparticles and Subsets of Extracellular Vesicles by Asymmetric Flow Field-Flow Fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef]
- Zhang, Q.; Jeppesen, D.K.; Higginbotham, J.N.; Graves-Deal, R.; Trinh, V.Q.; Ramirez, M.A.; Sohn, Y.; Neininger, A.C.; Taneja, N.; McKinley, E.T.; et al. Supermeres Are Functional Extracellular Nanoparticles Replete with Disease Biomarkers and Therapeutic Targets. Nat. Cell Biol. 2021, 23, 1240–1254. [Google Scholar] [CrossRef]
- Buzas, E.I. The Roles of Extracellular Vesicles in the Immune System. Nat. Rev. Immunol. 2023, 23, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Dixson, A.C.; Dawson, T.R.; Di Vizio, D.; Weaver, A.M. Context-Specific Regulation of Extracellular Vesicle Biogenesis and Cargo Selection. Nat. Rev. Mol. Cell Biol. 2023, 24, 454–476. [Google Scholar] [CrossRef] [PubMed]
- Grange, C.; Bussolati, B. Extracellular Vesicles in Kidney Disease. Nat. Rev. Nephrol. 2022, 18, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Karpman, D.; Ståhl, A.-L.; Arvidsson, I. Extracellular Vesicles in Renal Disease. Nat. Rev. Nephrol. 2017, 13, 545–562. [Google Scholar] [CrossRef]
- Malda, J.; Boere, J.; van de Lest, C.H.A.; van Weeren, P.R.; Wauben, M.H.M. Extracellular Vesicles—New Tool for Joint Repair and Regeneration. Nat. Rev. Rheumatol. 2016, 12, 243–249. [Google Scholar] [CrossRef]
- Möller, A.; Lobb, R.J. The Evolving Translational Potential of Small Extracellular Vesicles in Cancer. Nat. Rev. Cancer 2020, 20, 697–709. [Google Scholar] [CrossRef]
- Thompson, A.G.; Gray, E.; Heman-Ackah, S.M.; Mäger, I.; Talbot, K.; Andaloussi, S.E.; Wood, M.J.; Turner, M.R. Extracellular Vesicles in Neurodegenerative Disease—Pathogenesis to Biomarkers. Nat. Rev. Neurol. 2016, 12, 346–357. [Google Scholar] [CrossRef]
- Xu, R.; Rai, A.; Chen, M.; Suwakulsiri, W.; Greening, D.W.; Simpson, R.J. Extracellular Vesicles in Cancer—Implications for Future Improvements in Cancer Care. Nat. Rev. Clin. Oncol. 2018, 15, 617–638. [Google Scholar] [CrossRef]
- Witwer, K.W.; Théry, C. Extracellular Vesicles or Exosomes? On Primacy, Precision, and Popularity Influencing a Choice of Nomenclature. J. Extracell. Vesicles 2019, 8, 1648167. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.J.; Ashraf, A.; Chung, E.J. Extracellular Vesicles as Regulators of the Extracellular Matrix. Bioengineering 2023, 10, 136. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yu, R.; Xue, J.; Tang, Y.; Su, S.; Liao, C.; Guo, Q.; Guo, W.; Zheng, J. How Do Extracellular Vesicles Play a Key Role in the Maintenance of Bone Homeostasis and Regeneration? A Comprehensive Review of Literature. Int. J. Nanomed. 2022, 17, 5375–5389. [Google Scholar] [CrossRef] [PubMed]
- Irving, J.T. A Histological Staining Method for Sites of Calcification in Teeth and Bone. Arch. Oral. Biol. 1959, 1, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Irving, J.T. The Sudanophil Material at Sites of Calcification. Arch. Oral. Biol. 1963, 8, 735–745. [Google Scholar] [CrossRef]
- Irving, J.T.; Wuthier, R.E. Further Observations on the Sudan Black Stain for Calcification. Arch. Oral. Biol. 1961, 5, 323–324. [Google Scholar] [CrossRef]
- Anderson, H.C. Electron Microscopic Studies of Induced Cartilage Development and Calcification. J. Cell Biol. 1967, 35, 81–101. [Google Scholar] [CrossRef]
- Anderson, H.C. Vesicles Associated with Calcification in the Matrix of Epiphyseal Cartilage. J. Cell Biol. 1969, 41, 59–72. [Google Scholar] [CrossRef]
- Bonucci, E. Fine Structure of Early Cartilage Calcification. J. Ultrastruct. Res. 1967, 20, 33–50. [Google Scholar] [CrossRef]
- Ali, S.Y.; Anderson, H.C.; Sajdera, S.W. Enzymic and Electron-Microscopic Analysis of Extracellular Matrix Vesicles Associated with Calcification in Cartilage. Biochem. J. 1971, 122, 56P. [Google Scholar] [CrossRef]
- Ali, S.Y.; Sajdera, S.W.; Anderson, H.C. Isolation and Characterization of Calcifying Matrix Vesicles from Epiphyseal Cartilage. Proc. Natl. Acad. Sci. USA 1970, 67, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Majeska, R.J.; Wuthier, R.E. Studies on Matrix Vesicles Isolated from Chick Epiphyseal Cartilage. Association of Pyrophosphatase and ATPase Activities with Alkaline Phosphatase. Biochim. Biophys. Acta 1975, 391, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Peress, N.S.; Anderson, H.C.; Sajdera, S.W. The Lipids of Matrix Vesicles from Bovine Fetal Epiphyseal Cartilage. Calcif. Tissue Res. 1974, 14, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Wuthier, R.E. Lipid Composition of Isolated Epiphyseal Cartilage Cells, Membranes and Matrix Vesicles. Biochim. Biophys. Acta 1975, 409, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Wuthier, R.E. Lipids of Matrix Vesicles. Fed. Proc. 1976, 35, 117–121. [Google Scholar] [PubMed]
- Ketenjian, A.Y.; Arsenis, C. Morphological and Biochemical Studies during Differentiation and Calcification of Fracture Callus Cartilage. Clin. Orthop. Relat. Res. 1975, 107, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Jafri, A.M.; Lewis, N.J.; Arsenis, C. Purification of Alkaline Phosphatase from Extracellular Vesicles of Fracture Callus Cartilage. Calcif. Tissue Res. 1978, 25, 85–92. [Google Scholar] [CrossRef]
- Wuthier, R.E.; Lipscomb, G.F. Matrix Vesicles: Structure, Composition, Formation and Function in Calcification. Front. Biosci. 2011, 16, 2812–2902. [Google Scholar] [CrossRef]
- Bernard, G.W.; Pease, D.C. An Electron Microscopic Study of Initial Intramembranous Osteogenesis. Am. J. Anat. 1969, 125, 271–290. [Google Scholar] [CrossRef]
- Ketenjian, A.Y.; Jafri, A.M.; Arsenis, C. Studies on the Mechanism of Callus Cartilage Differentiation and Calcification during Fracture Healing. Orthop. Clin. N. Am. 1978, 9, 43–65. [Google Scholar] [CrossRef]
- Sisca, R.F.; Provenza, D.V. Initial Dentin Formation in Human Deciduous Teeth. An Electron Microscope Study. Calcif. Tissue Res. 1972, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M. Calcification of Matrix Vesicles in Human Aortic Valve and Aortic Media. Fed. Proc. 1976, 35, 156–162. [Google Scholar] [PubMed]
- Sela, J.; Bab, I.A. The Relationship between Extracellular Matrix Vesicles and Calcospherities in Primary Mineralization of Neoplastic Bone Tissue. TEM and SEM Studies on Osteosarcoma. Virchows Arch. A Pathol. Anat. Histol. 1979, 382, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Couch, Y.; Buzàs, E.I.; Di Vizio, D.; Gho, Y.S.; Harrison, P.; Hill, A.F.; Lötvall, J.; Raposo, G.; Stahl, P.D.; Théry, C.; et al. A Brief History of Nearly EV-Erything—The Rise and Rise of Extracellular Vesicles. J. Extracell. Vesicles 2021, 10, e12144. [Google Scholar] [CrossRef] [PubMed]
- Veziroglu, E.M.; Mias, G.I. Characterizing Extracellular Vesicles and Their Diverse RNA Contents. Front. Genet. 2020, 11, 700. [Google Scholar] [CrossRef]
- Davies, O.G. Extracellular Vesicles: From Bone Development to Regenerative Orthopedics. Mol. Ther. 2023, 31, 1251–1274. [Google Scholar] [CrossRef] [PubMed]
- Chargaff, E.; West, R. The Biological Significance of the Thromboplastic Protein of Blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar] [CrossRef]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Sun, C.N. Lattice Structures and Osmiophilic Bodies in the Developing Respiratory Tissue of Rats. J. Ultrastruct. Res. 1966, 15, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Mercer, E.H.; Shaffer, B.M. Electron Microscopy of Solitary and Aggregated Slime Mould Cells. J. Biophys. Biochem. Cytol. 1960, 7, 353–356. [Google Scholar] [CrossRef]
- Vickerman, K. Patterns of Cellular Organisation in Limax Amoebae. An Electron Microscope Study. Exp. Cell Res. 1962, 26, 497–519. [Google Scholar] [CrossRef] [PubMed]
- Grillo, M.A. Extracellular Synaptic Vesicles in the Mouse Heart. J. Cell Biol. 1970, 47, 547–553. [Google Scholar] [CrossRef]
- Dermietzel, R.; Venjakob, K.; Brettschneider, H. Occurrence of Extracellular Synaptic Vesicles in the Autonomic Nervous System. Naturwissenschaften 1972, 59, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Basso, M.; Bonetto, V. Extracellular Vesicles and a Novel Form of Communication in the Brain. Front. Neurosci. 2016, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- Budnik, V.; Ruiz-Cañada, C.; Wendler, F. Extracellular Vesicles Round off Communication in the Nervous System. Nat. Rev. Neurosci. 2016, 17, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Allan, D.; Billah, M.M.; Finean, J.B.; Michell, R.H. Release of Diacylglycerol-Enriched Vesicles from Erythrocytes with Increased Intracellular (Ca2+). Nature 1976, 261, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Aaronson, S.; Behrens, U.; Orner, R.; Haines, T.H. Ultrastructure of Intracellular and Extracellular Vesicles, Membranes, and Myelin Figures Produced by Ochromonas Danica. J. Ultrastruct. Res. 1971, 35, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Chigaleĭchik, A.G.; Belova, L.A.; Grishchenko, V.M.; Rylkin, S.S. Several properties of the extracellular vesicles of Candida tropicalis yeasts grown on n-alkanes. Mikrobiologiia 1977, 46, 467–471. [Google Scholar]
- Vysotskiĭ, V.V.; Mazurova, I.K.; Shmeleva, E.A. Extracellular material of some representatives of the genus Corynebacterium (the electron microscopic aspect). Zh. Mikrobiol. Epidemiol. Immunobiol. 1977, 8, 90–95. [Google Scholar]
- Käppeli, O.; Finnerty, W.R. Partition of Alkane by an Extracellular Vesicle Derived from Hexadecane-Grown Acinetobacter. J. Bacteriol. 1979, 140, 707–712. [Google Scholar] [CrossRef]
- da Silveira, J.F.; Abrahamsohn, P.A.; Colli, W. Plasma Membrane Vesicles Isolated from Epimastigote Forms of Trypanosoma Cruzi. Biochim. Biophys. Acta 1979, 550, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B Lymphocytes Secrete Antigen-Presenting Vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, J.; Miekus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M.Z. Embryonic Stem Cell-Derived Microvesicles Reprogram Hematopoietic Progenitors: Evidence for Horizontal Transfer of mRNA and Protein Delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of mRNAs and microRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Trams, E.G.; Lauter, C.J.; Salem, N.; Heine, U. Exfoliation of Membrane Ecto-Enzymes in the Form of Micro-Vesicles. Biochim. Biophys. Acta 1981, 645, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Shekari, F.; Alibhai, F.J.; Baharvand, H.; Börger, V.; Bruno, S.; Davies, O.; Giebel, B.; Gimona, M.; Salekdeh, G.H.; Martin-Jaular, L.; et al. Cell Culture-Derived Extracellular Vesicles: Considerations for Reporting Cell Culturing Parameters. J. Extracell. Biol. 2023, 2, e115. [Google Scholar] [CrossRef]
- Bottini, M.; Mebarek, S.; Anderson, K.L.; Strzelecka-Kiliszek, A.; Bozycki, L.; Simão, A.M.S.; Bolean, M.; Ciancaglini, P.; Pikula, J.B.; Pikula, S.; et al. Matrix Vesicles from Chondrocytes and Osteoblasts: Their Biogenesis, Properties, Functions and Biomimetic Models. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 532–546. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef]
- Williams, S.; Fernandez-Rhodes, M.; Law, A.; Peacock, B.; Lewis, M.P.; Davies, O.G. Comparison of Extracellular Vesicle Isolation Processes for Therapeutic Applications. J. Tissue Eng. 2023, 14, 20417314231174609. [Google Scholar] [CrossRef]
- Fernández-Rhodes, M.; Adlou, B.; Williams, S.; Lees, R.; Peacock, B.; Aubert, D.; Jalal, A.R.; Lewis, M.P.; Davies, O.G. Defining the Influence of Size-Exclusion Chromatography Fraction Window and Ultrafiltration Column Choice on Extracellular Vesicle Recovery in a Skeletal Muscle Model. J. Extracell. Biol. 2023, 2, e85. [Google Scholar] [CrossRef]
- Wu, L.N.; Genge, B.R.; Dunkelberger, D.G.; LeGeros, R.Z.; Concannon, B.; Wuthier, R.E. Physicochemical Characterization of the Nucleational Core of Matrix Vesicles. J. Biol. Chem. 1997, 272, 4404–4411. [Google Scholar] [CrossRef]
- Wuthier, R.E. Electrolytes of Isolated Epiphyseal Chondrocytes, Matrix Vesicles, and Extracellular Fluid. Calcif. Tissue Res. 1977, 23, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Garimella, R.; Bi, X.; Camacho, N.; Sipe, J.B.; Anderson, H.C. Primary Culture of Rat Growth Plate Chondrocytes: An in Vitro Model of Growth Plate Histotype, Matrix Vesicle Biogenesis and Mineralization. Bone 2004, 34, 961–970. [Google Scholar] [CrossRef]
- Kirsch, T.; von der Mark, K. Remodelling of Collagen Types I, II and X and Calcification of Human Fetal Cartilage. Bone Miner. 1992, 18, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, T.; Nah, H.D.; Shapiro, I.M.; Pacifici, M. Regulated Production of Mineralization-Competent Matrix Vesicles in Hypertrophic Chondrocytes. J. Cell Biol. 1997, 137, 1149–1160. [Google Scholar] [CrossRef]
- Descalzi Cancedda, F.; Gentili, C.; Manduca, P.; Cancedda, R. Hypertrophic Chondrocytes Undergo Further Differentiation in Culture. J. Cell Biol. 1992, 117, 427–435. [Google Scholar] [CrossRef]
- Gentili, C.; Bianco, P.; Neri, M.; Malpeli, M.; Campanile, G.; Castagnola, P.; Cancedda, R.; Cancedda, F.D. Cell Proliferation, Extracellular Matrix Mineralization, and Ovotransferrin Transient Expression during in Vitro Differentiation of Chick Hypertrophic Chondrocytes into Osteoblast-like Cells. J. Cell Biol. 1993, 122, 703–712. [Google Scholar] [CrossRef]
- Roach, H.I.; Erenpreisa, J.; Aigner, T. Osteogenic Differentiation of Hypertrophic Chondrocytes Involves Asymmetric Cell Divisions and Apoptosis. J. Cell Biol. 1995, 131, 483–494. [Google Scholar] [CrossRef]
- Rosenthal, A.K.; Gohr, C.M.; Ninomiya, J.; Wakim, B.T. Proteomic Analysis of Articular Cartilage Vesicles from Normal and Osteoarthritic Cartilage. Arthritis Rheum. 2011, 63, 401–411. [Google Scholar] [CrossRef]
- Ak, R. Articular Cartilage Vesicles and Calcium Crystal Deposition Diseases. Curr. Opin. Rheumatol. 2016, 28, 127. [Google Scholar] [CrossRef]
- Dean, D.D.; Schwartz, Z.; Bonewald, L.; Muniz, O.E.; Morales, S.; Gomez, R.; Brooks, B.P.; Qiao, M.; Howell, D.S.; Boyan, B.D. Matrix Vesicles Produced by Osteoblast-like Cells in Culture Become Significantly Enriched in Proteoglycan-Degrading Metalloproteinases after Addition of Beta-Glycerophosphate and Ascorbic Acid. Calcif. Tissue Int. 1994, 54, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Thouverey, C.; Strzelecka-Kiliszek, A.; Balcerzak, M.; Buchet, R.; Pikula, S. Matrix Vesicles Originate from Apical Membrane Microvilli of Mineralizing Osteoblast-like Saos-2 Cells. J. Cell Biochem. 2009, 106, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Buchet, R.; Pikula, S.; Magne, D.; Mebarek, S. Isolation and Characteristics of Matrix Vesicles. Methods Mol. Biol. 2013, 1053, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Boyan, B.D.; Asmussen, N.C.; Lin, Z.; Schwartz, Z. The Role of Matrix-Bound Extracellular Vesicles in the Regulation of Endochondral Bone Formation. Cells 2022, 11, 1619. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, Z.; Knight, G.; Swain, L.D.; Boyan, B.D. Localization of Vitamin D3-Responsive Alkaline Phosphatase in Cultured Chondrocytes. J. Biol. Chem. 1988, 263, 6023–6026. [Google Scholar] [CrossRef]
- Bessueille, L.; Briolay, A.; Como, J.; Mebarek, S.; Mansouri, C.; Gleizes, M.; El Jamal, A.; Buchet, R.; Dumontet, C.; Matera, E.L.; et al. Tissue-Nonspecific Alkaline Phosphatase Is an Anti-Inflammatory Nucleotidase. Bone 2020, 133, 115262. [Google Scholar] [CrossRef]
- Balcerzak, M.; Radisson, J.; Azzar, G.; Farlay, D.; Boivin, G.; Pikula, S.; Buchet, R. A Comparative Analysis of Strategies for Isolation of Matrix Vesicles. Anal. Biochem. 2007, 361, 176–182. [Google Scholar] [CrossRef]
- Walker, S.A.; Davidovich, I.; Yang, Y.; Lai, A.; Goncalves, J.P.; Deliwala, V.; Busatto, S.; Shapiro, S.; Koifman, N.; Salomon, C.; et al. Sucrose-Based Cryoprotective Storage of Extracellular Vesicles. Extracell. Vesicle 2022, 1, 100016. [Google Scholar] [CrossRef]
- Boyan, B.D.; Schwartz, Z.; Swain, L.D.; Carnes, D.L.; Zislis, T. Differential Expression of Phenotype by Resting Zone and Growth Region Costochondral Chondrocytes In Vitro. Bone 1988, 9, 185–194. [Google Scholar] [CrossRef]
- Kirsch, T.; Ishikawa, Y.; Mwale, F.; Wuthier, R.E. Roles of the Nucleational Core Complex and Collagens (Types II and X) in Calcification of Growth Plate Cartilage Matrix Vesicles. J. Biol. Chem. 1994, 269, 20103–20109. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Camalier, C.E.; Nagashima, K.; Chan, K.C.; Lucas, D.A.; de la Cruz, M.J.; Gignac, M.; Lockett, S.; Issaq, H.J.; Veenstra, T.D.; et al. Analysis of the Extracellular Matrix Vesicle Proteome in Mineralizing Osteoblasts. J. Cell Physiol. 2007, 210, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Skelton, A.M.; Cohen, D.J.; Boyan, B.D.; Schwartz, Z. Osteoblast-Derived Matrix Vesicles Exhibit Exosomal Traits and a Unique Subset of microRNA: Their Caveolae-Dependent Endocytosis Results in Reduced Osteogenic Differentiation. Int. J. Mol. Sci. 2023, 24, 12770. [Google Scholar] [CrossRef] [PubMed]
- Balcerzak, M.; Malinowska, A.; Thouverey, C.; Sekrecka, A.; Dadlez, M.; Buchet, R.; Pikula, S. Proteome Analysis of Matrix Vesicles Isolated from Femurs of Chicken Embryo. Proteomics 2008, 8, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Golub, E.E. Role of Matrix Vesicles in Biomineralization. Biochim. Biophys. Acta 2009, 1790, 1592–1598. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.C. Molecular Biology of Matrix Vesicles. Clin. Orthop. Relat. Res. 1995, 314, 266–280. [Google Scholar] [CrossRef]
- Kirsch, T.; Pfäffle, M. Selective Binding of Anchorin CII (Annexin V) to Type II and X Collagen and to Chondrocalcin (C-Propeptide of Type II Collagen). Implications for Anchoring Function between Matrix Vesicles and Matrix Proteins. FEBS Lett. 1992, 310, 143–147. [Google Scholar] [CrossRef]
- Kirsch, T.; Harrison, G.; Golub, E.E.; Nah, H.D. The Roles of Annexins and Types II and X Collagen in Matrix Vesicle-Mediated Mineralization of Growth Plate Cartilage. J. Biol. Chem. 2000, 275, 35577–35583. [Google Scholar] [CrossRef]
- von der Mark, K.; Mollenhauer, J. Annexin V Interactions with Collagen. Cell Mol. Life Sci. 1997, 53, 539–545. [Google Scholar] [CrossRef]
- Wu, L.N.; Genge, B.R.; Lloyd, G.C.; Wuthier, R.E. Collagen-Binding Proteins in Collagenase-Released Matrix Vesicles from Cartilage. Interaction between Matrix Vesicle Proteins and Different Types of Collagen. J. Biol. Chem. 1991, 266, 1195–1203. [Google Scholar] [CrossRef]
- D’Angelo, M.; Billings, P.C.; Pacifici, M.; Leboy, P.S.; Kirsch, T. Authentic Matrix Vesicles Contain Active Metalloproteases (MMP). a Role for Matrix Vesicle-Associated MMP-13 in Activation of Transforming Growth Factor-Beta. J. Biol. Chem. 2001, 276, 11347–11353. [Google Scholar] [CrossRef] [PubMed]
- Katsura, N.; Yamada, K. Isolation and Characterization of a Metalloprotease Associated with Chicken Epiphyseal Cartilage Matrix Vesicles. Bone 1986, 7, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.C. Matrix Vesicles and Calcification. Curr. Rheumatol. Rep. 2003, 5, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Watkins, E.L.; Stillo, J.V.; Wuthier, R.E. Subcellular Fractionation of Epiphyseal Cartilage: Isolation of Matrix Vesicles and Profiles of Enzymes, Phospholipids, Calcium and Phosphate. Biochim. Biophys. Acta 1980, 631, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Wuthier, R.E.; Chin, J.E.; Hale, J.E.; Register, T.C.; Hale, L.V.; Ishikawa, Y. Isolation and Characterization of Calcium-Accumulating Matrix Vesicles from Chondrocytes of Chicken Epiphyseal Growth Plate Cartilage in Primary Culture. J. Biol. Chem. 1985, 260, 15972–15979. [Google Scholar] [CrossRef] [PubMed]
- Warner, G.P.; Hubbard, H.L.; Lloyd, G.C.; Wuthier, R.E. 32Pi- and 45Ca-Metabolism by Matrix Vesicle-Enriched Microsomes Prepared from Chicken Epiphyseal Cartilage by Isosmotic Percoll Density-Gradient Fractionation. Calcif. Tissue Int. 1983, 35, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Plaut, J.S.; Strzelecka-Kiliszek, A.; Bozycki, L.; Pikula, S.; Buchet, R.; Mebarek, S.; Chadli, M.; Bolean, M.; Simao, A.M.S.; Ciancaglini, P.; et al. Quantitative Atomic Force Microscopy Provides New Insight into Matrix Vesicle Mineralization. Arch. Biochem. Biophys. 2019, 667, 14–21. [Google Scholar] [CrossRef]
- Bommanavar, S.; Hosmani, J.; Togoo, R.A.; Baeshen, H.A.; Raj, A.T.; Patil, S.; Bhandi, S.; Birkhed, D. Role of Matrix Vesicles and Crystal Ghosts in Bio-Mineralization. J. Bone Miner. Metab. 2020, 38, 759–764. [Google Scholar] [CrossRef]
- Anderson, H.C.; Stechschulte, D.J.; Collins, D.E.; Jacobs, D.H.; Morris, D.C.; Hsu, H.H.; Redford, P.A.; Zeiger, S. Matrix Vesicle Biogenesis in Vitro by Rachitic and Normal Rat Chondrocytes. Am. J. Pathol. 1990, 136, 391–398. [Google Scholar]
- Boyan, B.D.; Schwartz, Z.; Bonewald, L.F.; Swain, L.D. Localization of 1,25-(OH)2D3-Responsive Alkaline Phosphatase in Osteoblast-like Cells (ROS 17/2.8, MG 63, and MC 3T3) and Growth Cartilage Cells in Culture. J. Biol. Chem. 1989, 264, 11879–11886. [Google Scholar] [CrossRef]
- Chin, J.E.; Schalk, E.M.; Kemick, M.L.; Wuthier, R.E. Effect of Synthetic Human Parathyroid Hormone on the Levels of Alkaline Phosphatase Activity and Formation of Alkaline Phosphatase-Rich Matrix Vesicles by Primary Cultures of Chicken Epiphyseal Growth Plate Chondrocytes. Bone Miner. 1986, 1, 421–436. [Google Scholar] [PubMed]
- Genge, B.R.; Cao, X.; Wu, L.N.; Buzzi, W.R.; Showman, R.W.; Arsenault, A.L.; Ishikawa, Y.; Wuthier, R.E. Establishment of the Primary Structure of the Major Lipid-Dependent Ca2+ Binding Proteins of Chicken Growth Plate Cartilage Matrix Vesicles: Identity with Anchorin CII (Annexin V) and Annexin II. J. Bone Miner. Res. 1992, 7, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.E.; Chin, J.E.; Ishikawa, Y.; Paradiso, P.R.; Wuthier, R.E. Correlation between Distribution of Cytoskeletal Proteins and Release of Alkaline Phosphatase-Rich Vesicles by Epiphyseal Chondrocytes in Primary Culture. Cell Motil. 1983, 3, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Hale, L.V.; Kemick, M.L.; Wuthier, R.E. Effect of Vitamin D Metabolites on the Expression of Alkaline Phosphatase Activity by Epiphyseal Hypertrophic Chondrocytes in Primary Cell Culture. J. Bone Miner. Res. 1986, 1, 489–495. [Google Scholar] [CrossRef]
- Hale, J.E.; Wuthier, R.E. The Mechanism of Matrix Vesicle Formation. Studies on the Composition of Chondrocyte Microvilli and on the Effects of Microfilament-Perturbing Agents on Cellular Vesiculation. J. Biol. Chem. 1987, 262, 1916–1925. [Google Scholar] [CrossRef]
- Macrae, V.E.; Davey, M.G.; McTeir, L.; Narisawa, S.; Yadav, M.C.; Millan, J.L.; Farquharson, C. Inhibition of PHOSPHO1 Activity Results in Impaired Skeletal Mineralization during Limb Development of the Chick. Bone 2010, 46, 1146–1155. [Google Scholar] [CrossRef]
- Schalk, E.M.; Wuthier, R.E. Effect of Trifluoperazine and Other Drugs on Matrix Vesicle Formation by Chicken Growth Plate Chondrocytes in Primary Cell Culture. Biochem. Pharmacol. 1986, 35, 2373–2379. [Google Scholar] [CrossRef]
- Schwartz, Z.; Langston, G.G.; Swain, L.D.; Boyan, B.D. Inhibition of 1,25-(OH)2D3- and 24,25-(OH)2D3-Dependent Stimulation of Alkaline Phosphatase Activity by A23187 Suggests a Role for Calcium in the Mechanism of Vitamin D Regulation of Chondrocyte Cultures. J. Bone Miner. Res. 1991, 6, 709–718. [Google Scholar] [CrossRef]
- Swain, L.D.; Schwartz, Z.; Caulfield, K.; Brooks, B.P.; Boyan, B.D. Nongenomic Regulation of Chondrocyte Membrane Fluidity by 1,25-(OH)2D3 and 24,25-(OH)2D3 Is Dependent on Cell Maturation. Bone 1993, 14, 609–617. [Google Scholar] [CrossRef]
- Väänänen, H.K.; Morris, D.C.; Anderson, H.C. Calcification of Cartilage Matrix in Chondrocyte Cultures Derived from Rachitic Rat Growth Plate Cartilage. Metab. Bone Dis. Relat. Res. 1983, 5, 87–92. [Google Scholar] [CrossRef]
- Wu, L.N.; Sauer, G.R.; Genge, B.R.; Wuthier, R.E. Induction of Mineral Deposition by Primary Cultures of Chicken Growth Plate Chondrocytes in Ascorbate-Containing Media. Evidence of an Association between Matrix Vesicles and Collagen. J. Biol. Chem. 1989, 264, 21346–21355. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.C.; Simão, A.M.S.; Narisawa, S.; Huesa, C.; McKee, M.D.; Farquharson, C.; Millán, J.L. Loss of Skeletal Mineralization by the Simultaneous Ablation of PHOSPHO1 and Alkaline Phosphatase Function: A Unified Model of the Mechanisms of Initiation of Skeletal Calcification. J. Bone Miner. Res. 2011, 26, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.C.; Bottini, M.; Cory, E.; Bhattacharya, K.; Kuss, P.; Narisawa, S.; Sah, R.L.; Beck, L.; Fadeel, B.; Farquharson, C.; et al. Skeletal Mineralization Deficits and Impaired Biogenesis and Function of Chondrocyte-Derived Matrix Vesicles in Phospho1(-/-) and Phospho1/Pi T1 Double-Knockout Mice. J. Bone Miner. Res. 2016, 31, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Bougault, C.; El Jamal, A.; Briolay, A.; Mebarek, S.; Boutet, M.-A.; Garraud, T.; Le Goff, B.; Blanchard, F.; Magne, D.; Brizuela, L. Involvement of Sphingosine Kinase/Sphingosine 1-Phosphate Metabolic Pathway in Spondyloarthritis. Bone 2017, 103, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Magne, D.; Bluteau, G.; Faucheux, C.; Palmer, G.; Vignes-Colombeix, C.; Pilet, P.; Rouillon, T.; Caverzasio, J.; Weiss, P.; Daculsi, G.; et al. Phosphate Is a Specific Signal for ATDC5 Chondrocyte Maturation and Apoptosis-Associated Mineralization: Possible Implication of Apoptosis in the Regulation of Endochondral Ossification. J. Bone Miner. Res. 2003, 18, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic Comparison Defines Novel Markers to Characterize Heterogeneous Populations of Extracellular Vesicle Subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, T.; Wang, W.; Pfander, D. Functional Differences between Growth Plate Apoptotic Bodies and Matrix Vesicles. J. Bone Miner. Res. 2003, 18, 1872–1881. [Google Scholar] [CrossRef]
- Shapiro, I.M.; Landis, W.J.; Risbud, M.V. Matrix Vesicles: Are They Anchored Exosomes? Bone 2015, 79, 29–36. [Google Scholar] [CrossRef]
- Thouverey, C.; Malinowska, A.; Balcerzak, M.; Strzelecka-Kiliszek, A.; Buchet, R.; Dadlez, M.; Pikula, S. Proteomic Characterization of Biogenesis and Functions of Matrix Vesicles Released from Mineralizing Human Osteoblast-like Cells. J. Proteom. 2011, 74, 1123–1134. [Google Scholar] [CrossRef]
- Cmoch, A.; Strzelecka-Kiliszek, A.; Palczewska, M.; Groves, P.; Pikula, S. Matrix Vesicles Isolated from Mineralization-Competent Saos-2 Cells Are Selectively Enriched with Annexins and S100 Proteins. Biochem. Biophys. Res. Commun. 2011, 412, 683–687. [Google Scholar] [CrossRef]
- Strzelecka-Kiliszek, A.; Bozycki, L.; Mebarek, S.; Buchet, R.; Pikula, S. Characteristics of Minerals in Vesicles Produced by Human Osteoblasts hFOB 1.19 and Osteosarcoma Saos-2 Cells Stimulated for Mineralization. J. Inorg. Biochem. 2017, 171, 100–107. [Google Scholar] [CrossRef]
- Jiang, L.; Cui, Y.; Luan, J.; Zhou, X.; Zhou, X.; Han, J. A Comparative Proteomics Study on Matrix Vesicles of Osteoblast-like Saos-2 and U2-OS Cells. Intractable Rare Dis. Res. 2013, 2, 59–62. [Google Scholar] [CrossRef]
- Li, C.; Cui, Y.; Luan, J.; Zhou, X.; Li, H.; Wang, H.; Shi, L.; Han, J. Tenascin C Affects Mineralization of SaOS2 Osteoblast-like Cells through Matrix Vesicles. Drug Discov. Ther. 2016, 10, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Fedde, K.N. Human Osteosarcoma Cells Spontaneously Release Matrix-Vesicle-like Structures with the Capacity to Mineralize. Bone Miner. 1992, 17, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F.; Schwartz, Z.; Swain, L.D.; Ramirez, V.; Poser, J.; Boyan, B.D. Stimulation of Plasma Membrane and Matrix Vesicle Enzyme Activity by Transforming Growth Factor-Beta in Osteosarcoma Cell Cultures. J. Cell Physiol. 1990, 145, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F.; Schwartz, Z.; Swain, L.D.; Boyan, B.D. Stimulation of Matrix Vesicle Enzyme Activity in Osteoblast-like Cells by 1,25(OH)2D3 and Transforming Growth Factor Beta (TGF Beta). Bone Miner. 1992, 17, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, Z.; Dennis, R.; Bonewald, L.; Swain, L.; Gomez, R.; Boyan, B.D. Differential Regulation of Prostaglandin E2 Synthesis and Phospholipase A2 Activity by 1,25-(OH)2D3 in Three Osteoblast-like Cell Lines (MC-3T3-E1, ROS 17/2.8, and MG-63). Bone 1992, 13, 51–58. [Google Scholar] [CrossRef]
- Davies, O.G.; Cox, S.C.; Williams, R.L.; Tsaroucha, D.; Dorrepaal, R.M.; Lewis, M.P.; Grover, L.M. Annexin-Enriched Osteoblast-Derived Vesicles Act as an Extracellular Site of Mineral Nucleation within Developing Stem Cell Cultures. Sci. Rep. 2017, 7, 12639. [Google Scholar] [CrossRef]
- Ferreira, E.; Porter, R.M.; Wehling, N.; O’Sullivan, R.P.; Liu, F.; Boskey, A.; Estok, D.M.; Harris, M.B.; Vrahas, M.S.; Evans, C.H.; et al. Inflammatory Cytokines Induce a Unique Mineralizing Phenotype in Mesenchymal Stem Cells Derived from Human Bone Marrow. J. Biol. Chem. 2013, 288, 29494–29505. [Google Scholar] [CrossRef]
- Yamada, M. Ultrastructural and Cytochemical Studies on the Calcification of the Tendon-Bone Joint. Arch. Histol. Jpn. 1976, 39, 347–378. [Google Scholar] [CrossRef]
- Amizuka, N.; Hasegawa, T.; Oda, K.; Luiz de Freitas, P.H.; Hoshi, K.; Li, M.; Ozawa, H. Histology of Epiphyseal Cartilage Calcification and Endochondral Ossification. Front. Biosci.-Elite 2012, 4, 2085–2100. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T. Ultrastructure and Biological Function of Matrix Vesicles in Bone Mineralization. Histochem. Cell Biol. 2018, 149, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Bozycki, L.; Komiazyk, M.; Mebarek, S.; Buchet, R.; Pikula, S.; Strzelecka-Kiliszek, A. Analysis of Minerals Produced by hFOB 1.19 and Saos-2 Cells Using Transmission Electron Microscopy with Energy Dispersive X-Ray Microanalysis. J. Vis. Exp. 2018, 136, 57423. [Google Scholar] [CrossRef]
- Bilen, M.A.; Pan, T.; Lee, Y.-C.; Lin, S.-C.; Yu, G.; Pan, J.; Hawke, D.; Pan, B.-F.; Vykoukal, J.; Gray, K.; et al. Proteomics Profiling of Exosomes from Primary Mouse Osteoblasts under Proliferation versus Mineralization Conditions and Characterization of Their Uptake into Prostate Cancer Cells. J. Proteome Res. 2017, 16, 2709–2728. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Névo, N.; Jouve, M.; Valenzuela, J.I.; Maurin, M.; Verweij, F.J.; Palmulli, R.; Lankar, D.; Dingli, F.; Loew, D.; et al. Specificities of Exosome versus Small Ectosome Secretion Revealed by Live Intracellular Tracking of CD63 and CD9. Nat. Commun. 2021, 12, 4389. [Google Scholar] [CrossRef]
- Chen, N.X.; O’Neill, K.D.; Moe, S.M. Matrix Vesicles Induce Calcification of Recipient Vascular Smooth Muscle Cells through Multiple Signaling Pathways. Kidney Int. 2018, 93, 343–354. [Google Scholar] [CrossRef]
- Mizukami, Y.; Kawao, N.; Takafuji, Y.; Ohira, T.; Okada, K.; Jo, J.-I.; Tabata, Y.; Kaji, H. Matrix Vesicles Promote Bone Repair after a Femoral Bone Defect in Mice. PLoS ONE 2023, 18, e0284258. [Google Scholar] [CrossRef]
- Uenaka, M.; Yamashita, E.; Kikuta, J.; Morimoto, A.; Ao, T.; Mizuno, H.; Furuya, M.; Hasegawa, T.; Tsukazaki, H.; Sudo, T.; et al. Osteoblast-Derived Vesicles Induce a Switch from Bone-Formation to Bone-Resorption in Vivo. Nat. Commun. 2022, 13, 1066. [Google Scholar] [CrossRef]
- Chen, N.X.; Moe, S.M. Pathophysiology of Vascular Calcification. Curr. Osteoporos. Rep. 2015, 13, 372–380. [Google Scholar] [CrossRef]
- Tanimura, A.; McGregor, D.H.; Anderson, H.C. Matrix Vesicles in Atherosclerotic Calcification. Proc. Soc. Exp. Biol. Med. 1983, 172, 173–177. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Akbulut, A.C.; Kaczor, D.M.; Halder, M.; Koenen, R.R.; Kramann, R. Initiation and Propagation of Vascular Calcification Is Regulated by a Concert of Platelet- and Smooth Muscle Cell-Derived Extracellular Vesicles. Front. Cardiovasc. Med. 2018, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Bakhshian Nik, A.; Hutcheson, J.D.; Aikawa, E. Extracellular Vesicles As Mediators of Cardiovascular Calcification. Front. Cardiovasc. Med. 2017, 4, 78. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, A.N.; Davies, J.D.; Reynolds, J.L.; McNair, R.; Jones, G.T.; Sidibe, A.; Schurgers, L.J.; Skepper, J.N.; Proudfoot, D.; Mayr, M.; et al. Calcium Regulates Key Components of Vascular Smooth Muscle Cell-Derived Matrix Vesicles to Enhance Mineralization. Circ. Res. 2011, 109, e1–e12. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, A.N.; Shanahan, C.M. Emerging Roles for Vascular Smooth Muscle Cell Exosomes in Calcification and Coagulation. J. Physiol. 2016, 594, 2905–2914. [Google Scholar] [CrossRef] [PubMed]
- Zazzeroni, L.; Faggioli, G.; Pasquinelli, G. Mechanisms of Arterial Calcification: The Role of Matrix Vesicles. Eur. J. Vasc. Endovasc. Surg. 2018, 55, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, E.; Blaser, M.C. 2020 Jeffrey M. Hoeg Award Lecture: Calcifying Extracellular Vesicles as Building Blocks of Microcalcifications in Cardiovascular Disorders. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yu, H.; Zhang, D.; Feng, T.; Miao, M.; Li, J.; Liu, X. Matrix Vesicles as a Therapeutic Target for Vascular Calcification. Front. Cell Dev. Biol. 2022, 10, 825622. [Google Scholar] [CrossRef]
- Krohn, J.B.; Hutcheson, J.D.; Martínez-Martínez, E.; Aikawa, E. Extracellular Vesicles in Cardiovascular Calcification: Expanding Current Paradigms. J. Physiol. 2016, 594, 2895–2903. [Google Scholar] [CrossRef]
- Hutcheson, J.D.; Goettsch, C.; Bertazzo, S.; Maldonado, N.; Ruiz, J.L.; Goh, W.; Yabusaki, K.; Faits, T.; Bouten, C.; Franck, G.; et al. Genesis and Growth of Extracellular-Vesicle-Derived Microcalcification in Atherosclerotic Plaques. Nat. Mater. 2016, 15, 335–343. [Google Scholar] [CrossRef]
- Yang, S.; Zeng, Z.; Yuan, Q.; Chen, Q.; Wang, Z.; Xie, H.; Liu, J. Vascular Calcification: From the Perspective of Crosstalk. Mol. Biomed. 2023, 4, 35. [Google Scholar] [CrossRef]
- Chaudhary, S.C.; Kuzynski, M.; Bottini, M.; Beniash, E.; Dokland, T.; Mobley, C.G.; Yadav, M.C.; Poliard, A.; Kellermann, O.; Millán, J.L.; et al. Phosphate Induces Formation of Matrix Vesicles during Odontoblast-Initiated Mineralization in Vitro. Matrix Biol. 2016, 52–54, 284–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.X.; Chen, X.; O’Neill, K.D.; Atkinson, S.J.; Moe, S.M. RhoA/Rho Kinase (ROCK) Alters Fetuin-A Uptake and Regulates Calcification in Bovine Vascular Smooth Muscle Cells (BVSMC). Am. J. Physiol. Renal Physiol. 2010, 299, F674–F680. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, P.; Chen, N.X.; O’Neill, K.; McClintick, J.N.; Moe, S.M.; Janga, S.C. Differential miRNA Expression in Cells and Matrix Vesicles in Vascular Smooth Muscle Cells from Rats with Kidney Disease. PLoS ONE 2015, 10, e0131589. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.X.; O’Neill, K.D.; Dominguez, J.M.; Moe, S.M. Regulation of Reactive Oxygen Species in the Pathogenesis of Matrix Vesicles Induced Calcification of Recipient Vascular Smooth Muscle Cells. Vasc. Med. 2021, 26, 585–594. [Google Scholar] [CrossRef]
- Reynolds, J.L.; Joannides, A.J.; Skepper, J.N.; McNair, R.; Schurgers, L.J.; Proudfoot, D.; Jahnen-Dechent, W.; Weissberg, P.L.; Shanahan, C.M. Human Vascular Smooth Muscle Cells Undergo Vesicle-Mediated Calcification in Response to Changes in Extracellular Calcium and Phosphate Concentrations: A Potential Mechanism for Accelerated Vascular Calcification in ESRD. J. Am. Soc. Nephrol. 2004, 15, 2857–2867. [Google Scholar] [CrossRef]
- Shroff, R.C.; McNair, R.; Skepper, J.N.; Figg, N.; Schurgers, L.J.; Deanfield, J.; Rees, L.; Shanahan, C.M. Chronic Mineral Dysregulation Promotes Vascular Smooth Muscle Cell Adaptation and Extracellular Matrix Calcification. J. Am. Soc. Nephrol. 2010, 21, 103–112. [Google Scholar] [CrossRef]
- Kapustin, A.N.; Chatrou, M.L.L.; Drozdov, I.; Zheng, Y.; Davidson, S.M.; Soong, D.; Furmanik, M.; Sanchis, P.; De Rosales, R.T.M.; Alvarez-Hernandez, D.; et al. Vascular Smooth Muscle Cell Calcification Is Mediated by Regulated Exosome Secretion. Circ. Res. 2015, 116, 1312–1323. [Google Scholar] [CrossRef]
- Kapustin, A.N.; Schoppet, M.; Schurgers, L.J.; Reynolds, J.L.; McNair, R.; Heiss, A.; Jahnen-Dechent, W.; Hackeng, T.M.; Schlieper, G.; Harrison, P.; et al. Prothrombin Loading of Vascular Smooth Muscle Cell-Derived Exosomes Regulates Coagulation and Calcification. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e22–e32. [Google Scholar] [CrossRef]
- New, S.E.P.; Goettsch, C.; Aikawa, M.; Marchini, J.F.; Shibasaki, M.; Yabusaki, K.; Libby, P.; Shanahan, C.M.; Croce, K.; Aikawa, E. Macrophage-Derived Matrix Vesicles: An Alternative Novel Mechanism for Microcalcification in Atherosclerotic Plaques. Circ. Res. 2013, 113, 72–77. [Google Scholar] [CrossRef]
- Cui, L.; Rashdan, N.A.; Zhu, D.; Milne, E.M.; Ajuh, P.; Milne, G.; Helfrich, M.H.; Lim, K.; Prasad, S.; Lerman, D.A.; et al. End Stage Renal Disease-Induced Hypercalcemia May Promote Aortic Valve Calcification via Annexin VI Enrichment of Valve Interstitial Cell Derived-Matrix Vesicles. J. Cell Physiol. 2017, 232, 2985–2995. [Google Scholar] [CrossRef]
- Chaudhary, S.C.; Khalid, S.; Smethurst, V.; Monier, D.; Mobley, J.; Huet, A.; Conway, J.F.; Napierala, D. Proteomic Profiling of Extracellular Vesicles Released from Vascular Smooth Muscle Cells during Initiation of Phosphate-Induced Mineralization. Connect. Tissue Res. 2018, 59, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Roszkowska, M.; Strzelecka-Kiliszek, A.; Bessueille, L.; Buchet, R.; Magne, D.; Pikula, S. Collagen Promotes Matrix Vesicle-Mediated Mineralization by Vascular Smooth Muscle Cells. J. Inorg. Biochem. 2018, 186, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-X.; Luo, Z.-W.; Li, F.-X.-Z.; Cao, J.; Rao, S.-S.; Liu, Y.-W.; Wang, Y.-Y.; Zhu, G.-Q.; Gong, J.-S.; Zou, J.-T.; et al. Aged Bone Matrix-Derived Extracellular Vesicles as a Messenger for Calcification Paradox. Nat Commun 2022, 13, 1453. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Hu, Y.; Yang, P.; Xie, X.; Fang, B. Extracellular Vesicle-Loaded Hydrogels for Tissue Repair and Regeneration. Mater. Today Bio 2023, 18, 100522. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
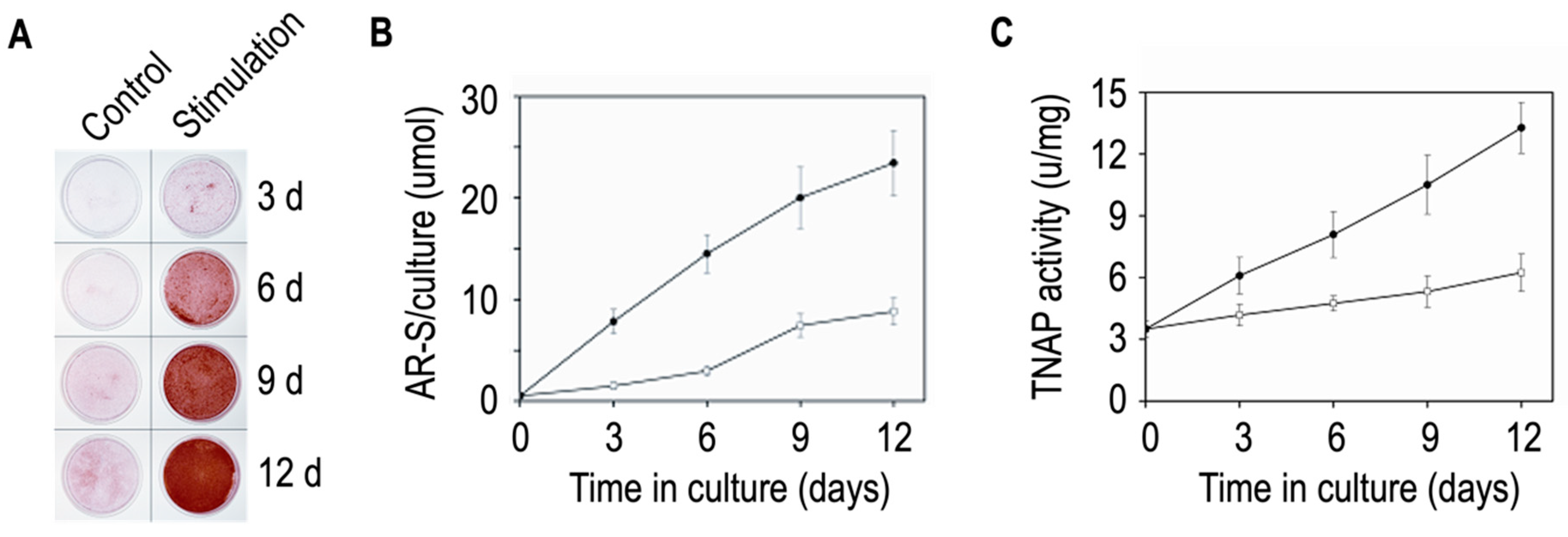
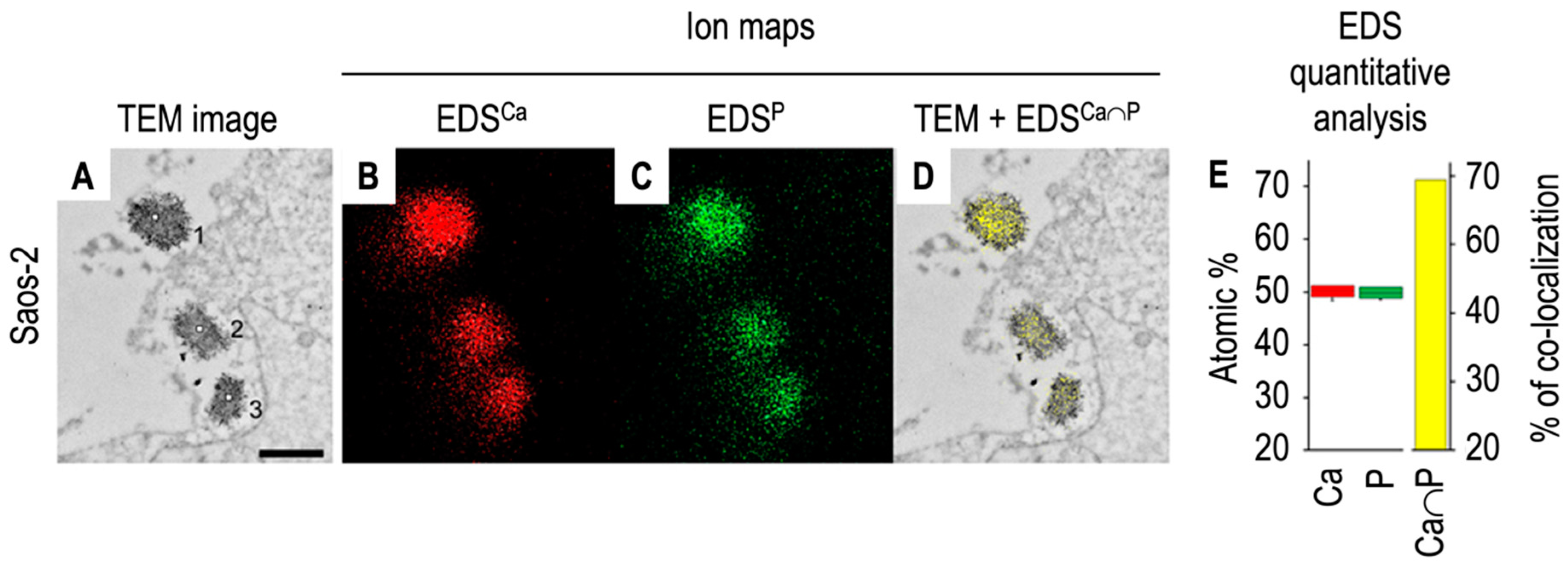
| Digestion | Properties | Samples | Ratio TNAP | Mineralization | Apatite in | References |
|---|---|---|---|---|---|---|
| Process | Activity | Assay | Lumen | |||
| MVs: Media EVs | ||||||
| Growth plate | From 4 to 6 | YES | YES (IR) | [88] | ||
| cartilage chicken | ||||||
| Collagenase | Widely used | Primary | From 8 to 12 | YES | ND | [87] |
| Chondrocytes | ||||||
| Saos2 cells | Around 16 * | YES | YES (IR) | [83] | ||
| Collagenase and | MVs without | Growth plate | ND | Calcium uptake | Calcium uptake | [91] |
| hyaluronidase | surface collagens | cartilage chicken | was impaired | was impaired | ||
| Collagenase and | MVs with surface | Growth plate | ND | Calcium uptake | Calcium uptake | [91] |
| trypsin | attached collagens | cartilage chicken | was optimum | was optimum | ||
| Hyaluronidase | Used for | Non mineralizing | ND | ND | ND | [80] |
| articular | articular | |||||
| chondrocytes | chondrocytes | |||||
| Trypsin | MG-63 cells | Around 6 ** | ND | ND | [93] | |
| Liberase and | Gentle digestion | MC3T3-E1 | From 0.7 to 0.8 | ND | ND | [92] |
| blendzyme-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mebarek, S.; Buchet, R.; Pikula, S.; Strzelecka-Kiliszek, A.; Brizuela, L.; Corti, G.; Collacchi, F.; Anghieri, G.; Magrini, A.; Ciancaglini, P.; et al. Do Media Extracellular Vesicles and Extracellular Vesicles Bound to the Extracellular Matrix Represent Distinct Types of Vesicles? Biomolecules 2024, 14, 42. https://doi.org/10.3390/biom14010042
Mebarek S, Buchet R, Pikula S, Strzelecka-Kiliszek A, Brizuela L, Corti G, Collacchi F, Anghieri G, Magrini A, Ciancaglini P, et al. Do Media Extracellular Vesicles and Extracellular Vesicles Bound to the Extracellular Matrix Represent Distinct Types of Vesicles? Biomolecules. 2024; 14(1):42. https://doi.org/10.3390/biom14010042
Chicago/Turabian StyleMebarek, Saida, Rene Buchet, Slawomir Pikula, Agnieszka Strzelecka-Kiliszek, Leyre Brizuela, Giada Corti, Federica Collacchi, Genevieve Anghieri, Andrea Magrini, Pietro Ciancaglini, and et al. 2024. "Do Media Extracellular Vesicles and Extracellular Vesicles Bound to the Extracellular Matrix Represent Distinct Types of Vesicles?" Biomolecules 14, no. 1: 42. https://doi.org/10.3390/biom14010042