
The Expression of Trace Amine-Associated Receptors (TAARs) in Breast Cancer Is Coincident with the Expression of Neuroactive Ligand–Receptor Systems and Depends on Tumor Intrinsic Subtype

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Public Resources and Databases
2.2. Data Normalization and Statistical Analysis
2.3. Survival Analysis
2.4. Gene Co-Expression Measurement
2.5. KEGG Pathway Enrichment Analysis
3. Results
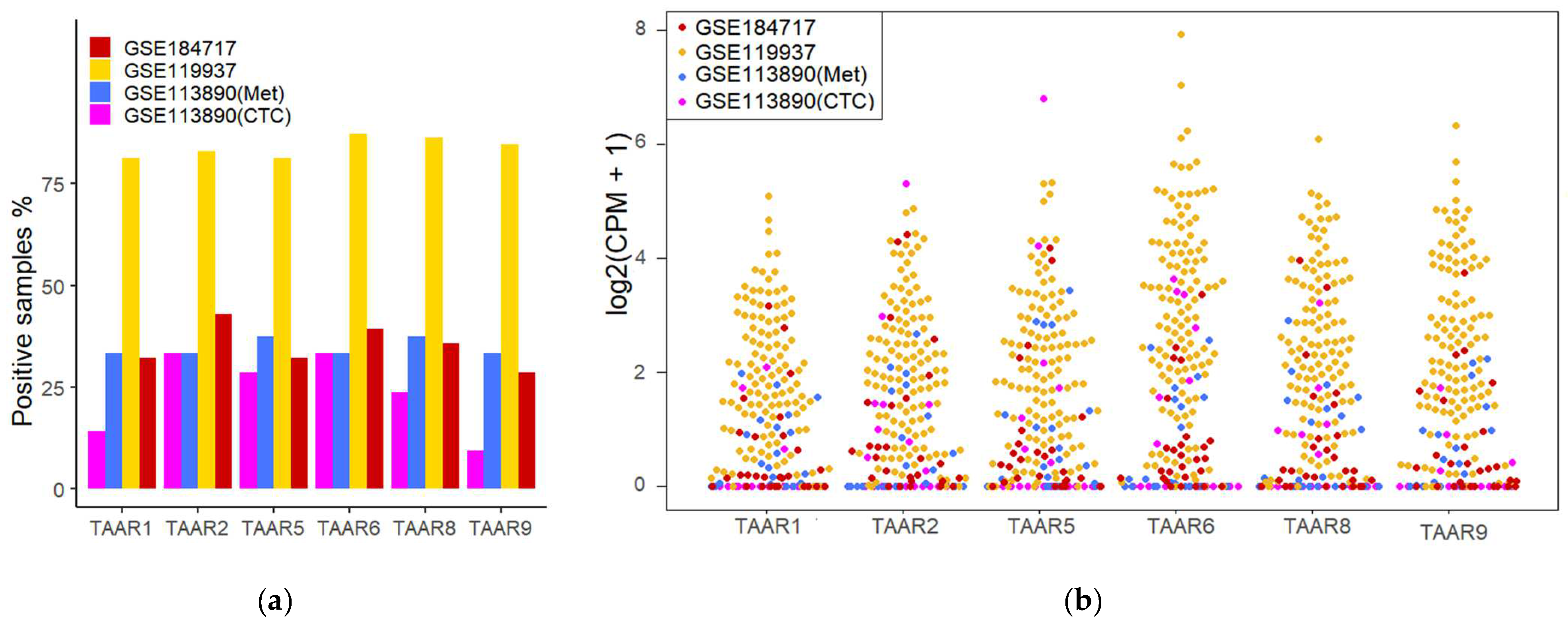
3.1. TAARs’ mRNA Expression in Primary Breast Tumors, BC Metastases, and CTCs Is Confirmed via RNA-Seq-Generated Dataset Analysis
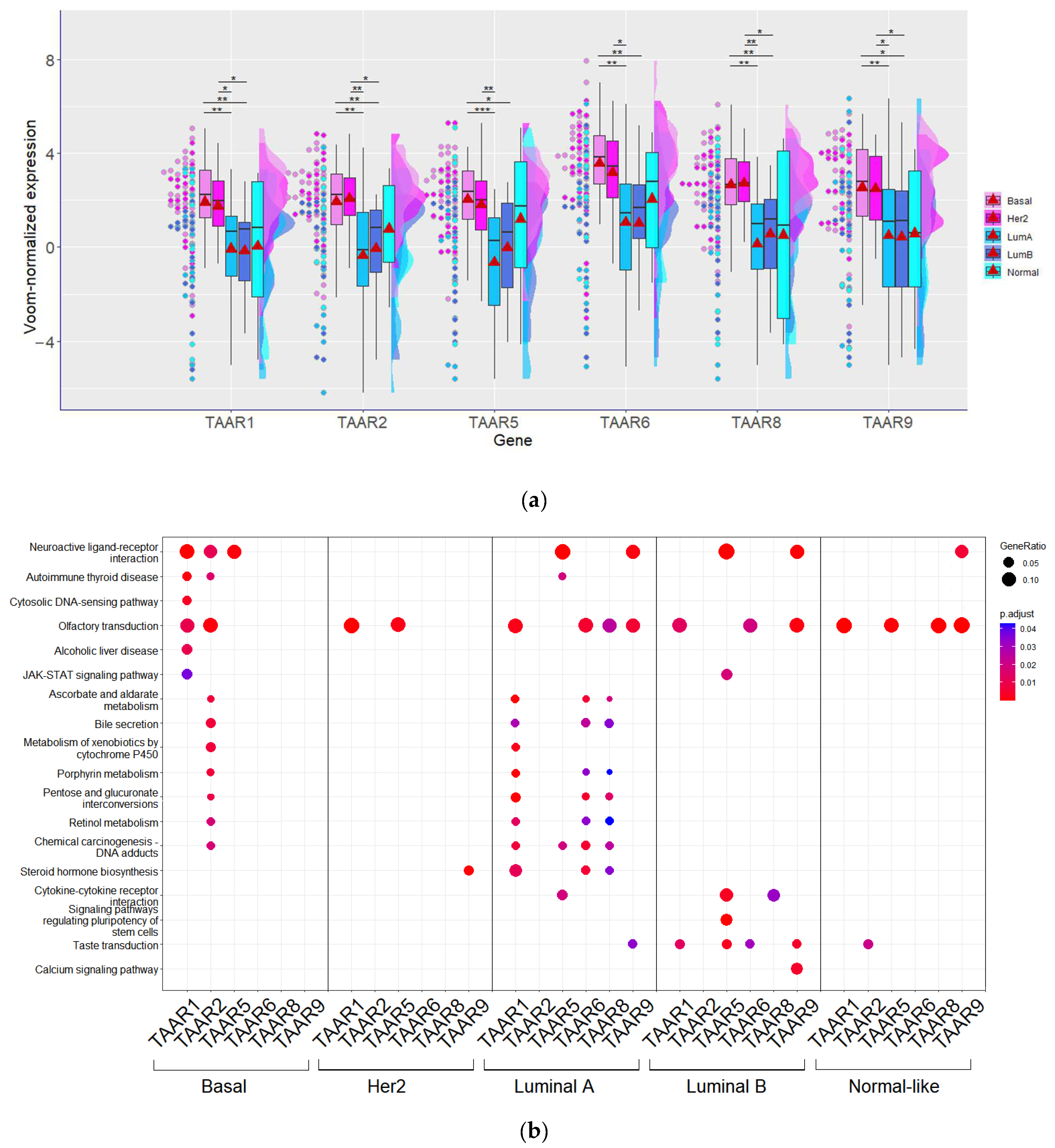
3.2. TAAR Expression Pattern in BC Depends on Tumor Intrinsic Subtype but Is Not Associated with Tumor Grade or Stage
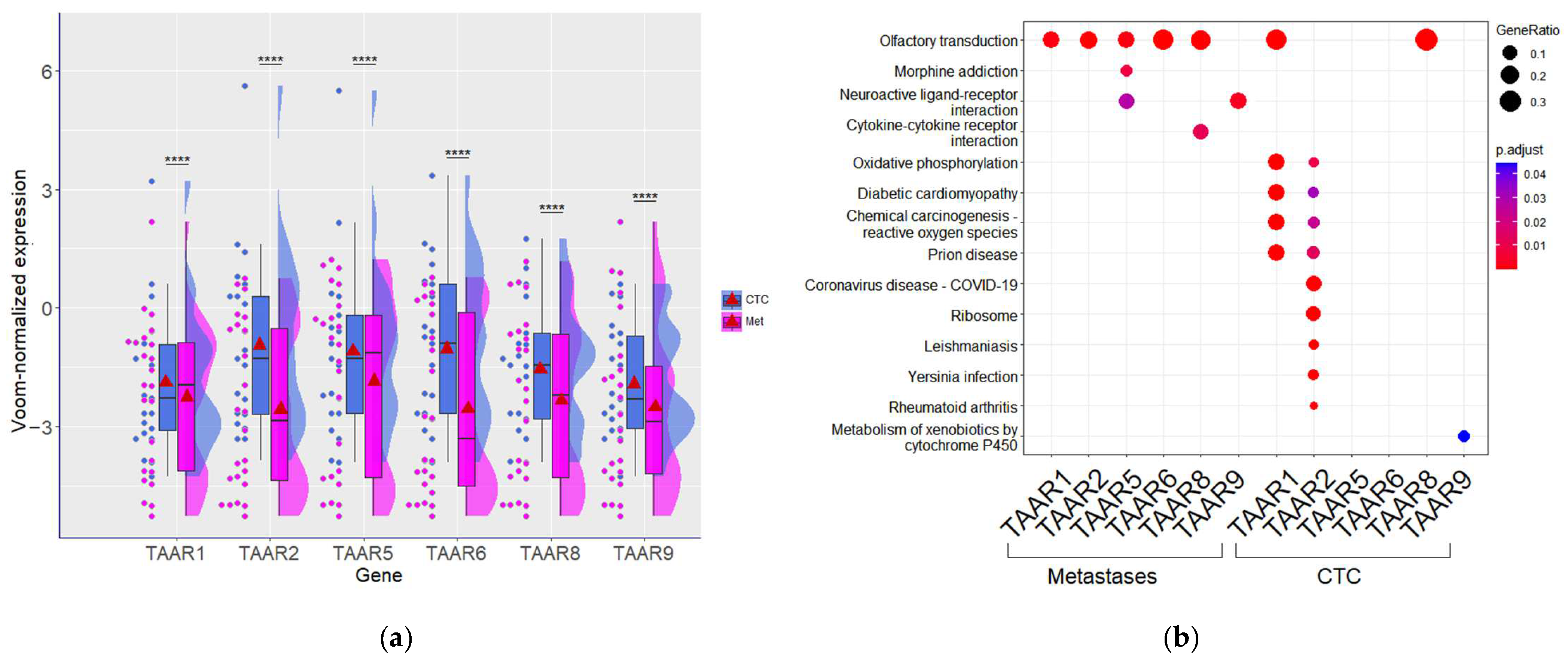
3.3. TAAR Expression Levels and Functional Associations Differ Significantly between CTCs and Metastatic Lesions in BC Patients
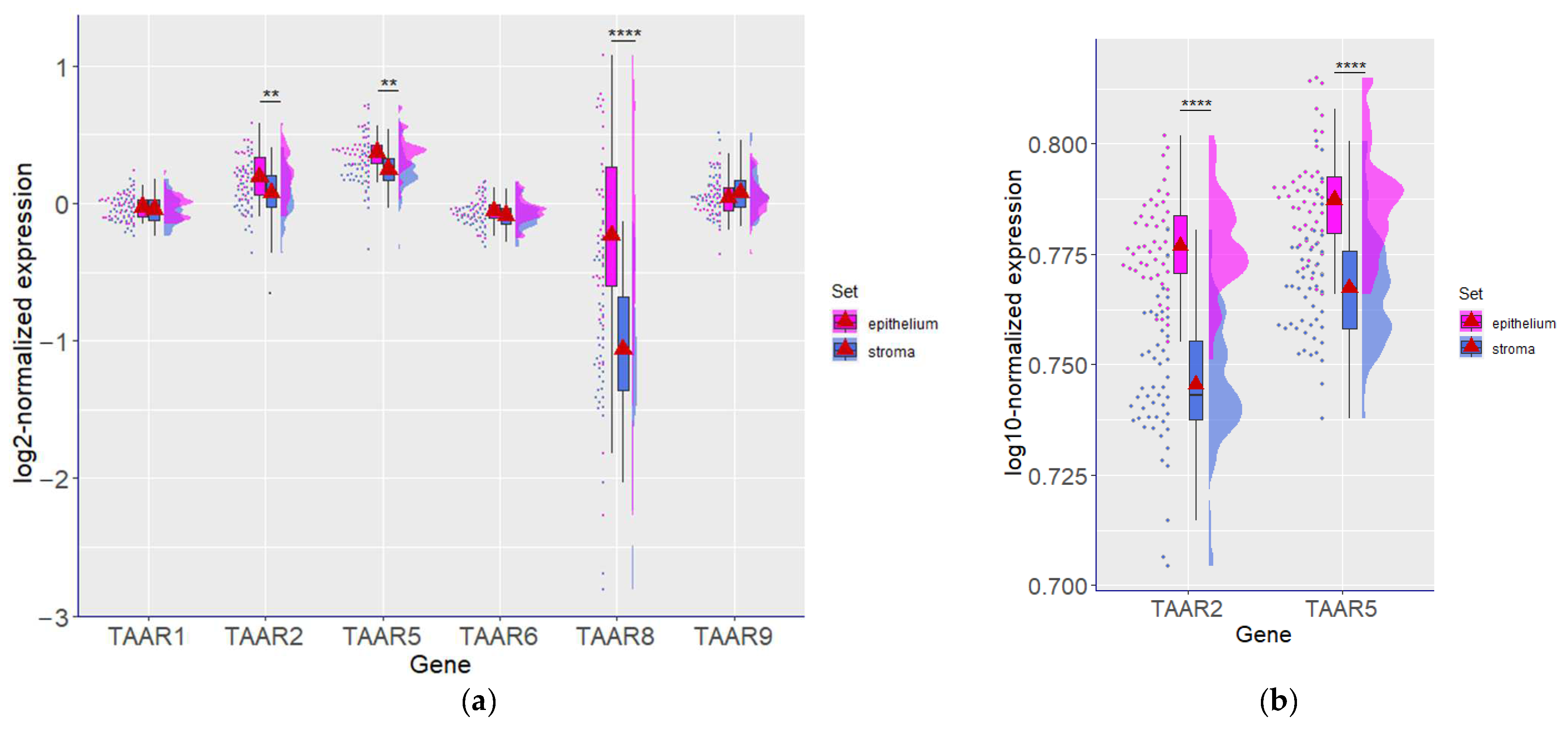
3.4. TAAR Expression Is Associated with Cancer Epithelial Cells Rather Than Stromal Cells
3.5. TAARs mRNA Expression Is Not Associated with Outcome in Studied Microarray-Generated Datasets
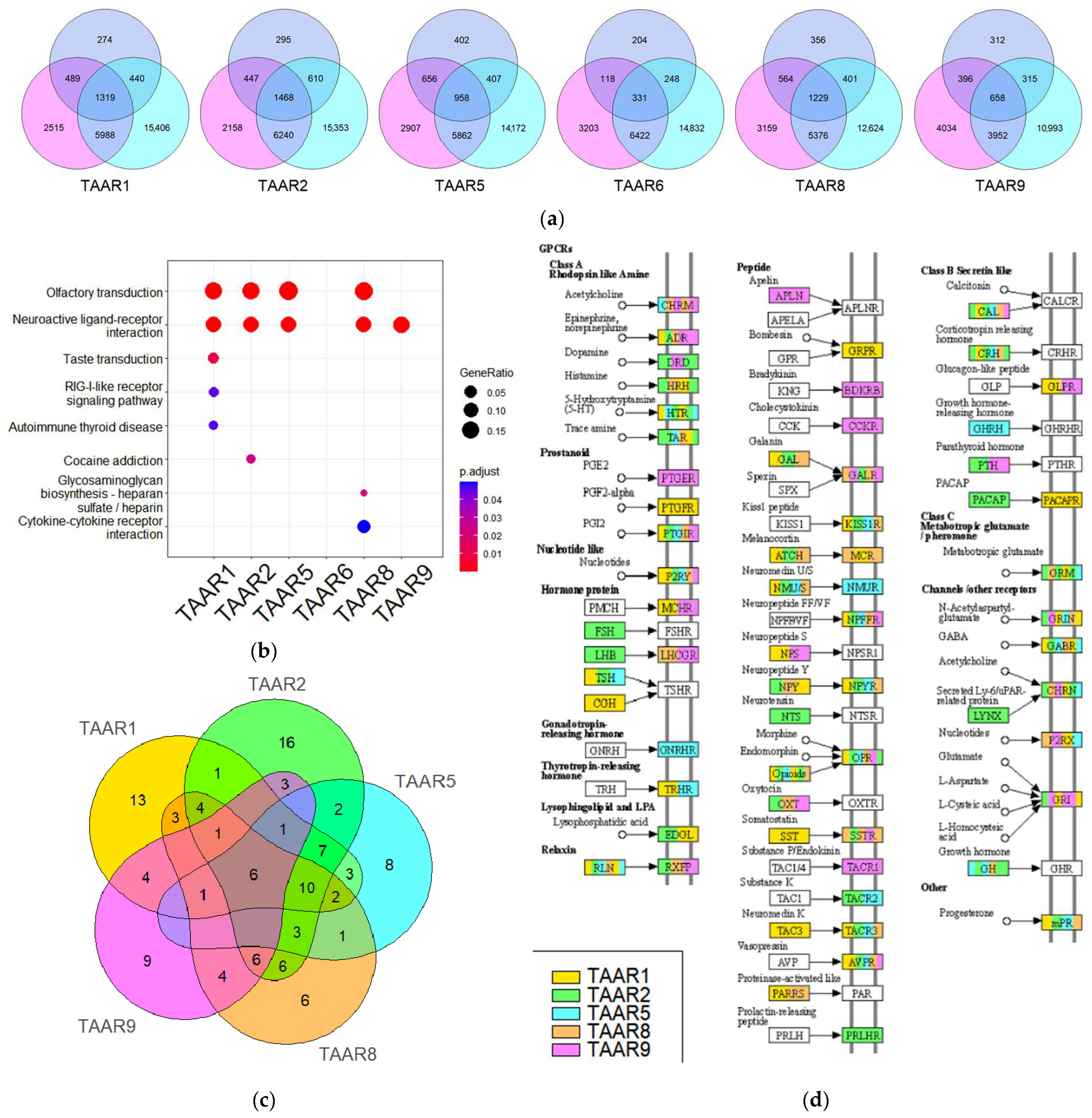
3.6. KEGG Pathway Enrichment Analysis Demonstrates TAARs’ Co-Expression with Neuroactive Ligand-Binding GPCRs, Including Monoamine Receptors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of Human Trace Amine-Associated Receptors: Therapeutic Opportunities and Challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Sukhanov, I.; Efimova, E.V.; Kozlova, A.; Antonova, K.A.; Illiano, P.; Leo, D.; Merkulyeva, N.; Kalinina, D.; Musienko, P.; et al. Trace Amine-Associated Receptor 5 Provides Olfactory Input into Limbic Brain Areas and Modulates Emotional Behaviors and Serotonin Transmission. Front. Mol. Neurosci. 2020, 13, 18. [Google Scholar] [CrossRef]
- Katolikova, N.V.; Vaganova, A.N.; Efimova, E.V.; Gainetdinov, R.R. Expression of Trace Amine-Associated Receptors in the Murine and Human Hippocampus Based on Public Transcriptomic Data. Cells 2022, 11, 1813. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, L.M.; Somaiya, R.D.; Miller, G.M. Review and Meta-Analyses of TAAR1 Expression in the Immune System and Cancers. Front. Pharmacol. 2018, 9, 683. [Google Scholar] [CrossRef] [PubMed]
- Tremmel, E.; Hofmann, S.; Kuhn, C.; Heidegger, H.; Heublein, S.; Hermelink, K.; Wuerstlein, R.; Harbeck, N.; Mayr, D.; Mahner, S.; et al. Thyronamine Regulation of TAAR1 Expression in Breast Cancer Cells and Investigation of Its Influence on Viability and Migration. Breast Cancer 2019, 11, 87–97. [Google Scholar] [CrossRef]
- Vattai, A.; Akyol, E.; Kuhn, C.; Hofmann, S.; Heidegger, H.; von Koch, F.; Hermelink, K.; Wuerstlein, R.; Harbeck, N.; Mayr, D.; et al. Increased Trace Amine-Associated Receptor 1 (TAAR1) Expression Is Associated with a Positive Survival Rate in Patients with Breast Cancer. J. Cancer Res. Clin. Oncol. 2017, 143, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Vogelsang, T.L.R.; Vattai, A.; Schmoeckel, E.; Kaltofen, T.; Chelariu-Raicu, A.; Zheng, M.; Mahner, S.; Mayr, D.; Jeschke, U.; Trillsch, F. Trace Amine-Associated Receptor 1 (TAAR1) Is a Positive Prognosticator for Epithelial Ovarian Cancer. Int. J. Mol. Sci. 2021, 22, 8479. [Google Scholar] [CrossRef]
- Kovács, T.; Mikó, E.; Vida, A.; Sebő, É.; Toth, J.; Csonka, T.; Boratkó, A.; Ujlaki, G.; Lente, G.; Kovács, P.; et al. Cadaverine, a Metabolite of the Microbiome, Reduces Breast Cancer Aggressiveness through Trace Amino Acid Receptors. Sci. Rep. 2019, 9, 1300. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Zheng, R.; Zhang, S.; Wang, S.; Chen, R.; Sun, K.; Zeng, H.; Zhou, J.; Wei, W. Global Patterns of Breast Cancer Incidence and Mortality: A Population-based Cancer Registry Data Analysis from 2000 to 2020. Cancer Commun. 2021, 41, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Pottegård, A.; Lash, T.L.; Cronin-Fenton, D.; Ahern, T.P.; Damkier, P. Use of Antipsychotics and Risk of Breast Cancer: A Danish Nationwide Case–Control Study. Br. J. Clin. Pharmacol. 2018, 84, 2152–2161. [Google Scholar] [CrossRef] [PubMed]
- Rogowski, M.; Gollahon, L.; Chellini, G.; Assadi-Porter, F.M. Uptake of 3-Iodothyronamine Hormone Analogs Inhibits the Growth and Viability of Cancer Cells. FEBS Open Bio. 2017, 7, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Dinter, J.; Mühlhaus, J.; Jacobi, S.F.; Wienchol, C.L.; Cöster, M.; Meister, J.; Hoefig, C.S.; Müller, A.; Köhrle, J.; Grüters, A.; et al. 3-Iodothyronamine Differentially Modulates α-2A-Adrenergic Receptor-Mediated Signaling. J. Mol. Endocrinol. 2015, 54, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Salahpour, A.; Masri, B.; Sotnikova, T.D.; Messa, M.; Barak, L.S.; Caron, M.G.; Gainetdinov, R.R. Functional Interaction between Trace Amine-Associated Receptor 1 and Dopamine D2 Receptor. Mol. Pharmacol. 2011, 80, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Revel, F.G.; Moreau, J.-L.; Gainetdinov, R.R.; Bradaia, A.; Sotnikova, T.D.; Mory, R.; Durkin, S.; Zbinden, K.G.; Norcross, R.; Meyer, C.A.; et al. TAAR1 Activation Modulates Monoaminergic Neurotransmission, Preventing Hyperdopaminergic and Hypoglutamatergic Activity. Proc. Natl. Acad. Sci. USA 2011, 108, 8485–8490. [Google Scholar] [CrossRef] [PubMed]
- Duarte, D.; Vale, N. Antipsychotic Drug Fluphenazine against Human Cancer Cells. Biomolecules 2022, 12, 1360. [Google Scholar] [CrossRef] [PubMed]
- Goyette, M.-A.; Cusseddu, R.; Elkholi, I.; Abu-Thuraia, A.; El-Hachem, N.; Haibe-Kains, B.; Gratton, J.-P.; Côté, J.-F. AXL Knockdown Gene Signature Reveals a Drug Repurposing Opportunity for a Class of Antipsychotics to Reduce Growth and Metastasis of Triple-Negative Breast Cancer. Oncotarget 2019, 10, 2055–2067. [Google Scholar] [CrossRef] [PubMed]
- Gwynne, W.D.; Shakeel, M.S.; Girgis-Gabardo, A.; Hassell, J.A. The Role of Serotonin in Breast Cancer Stem Cells. Molecules 2021, 26, 3171. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xia, L.; Lin, J.; Gong, L.; Xia, Y.; Xu, Y.; Liu, L.; Bao, J.; Zhang, C.; Chai, Y.; et al. Thioridazine Combined with Carboplatin Results in Synergistic Inhibition of Triple Negative Breast Cancer by Targeting Cancer Stem Cells. Transl. Oncol. 2022, 26, 101549. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.-B.; Luo, C.; Mao, X.-Y.; Li, X.; Yin, J.-Y.; Zhang, W.; Zhou, H.-H.; Liu, Z.-Q. The Prospective Value of Dopamine Receptors on Bio-Behavior of Tumor. J. Cancer 2019, 10, 1622–1632. [Google Scholar] [CrossRef]
- Yang, L.; Yao, Y.; Yong, L.; Feng, Y.; Su, H.; Yao, Q.; Xue, J.; Lu, W.; Zhou, T. Dopamine D1 Receptor Agonists Inhibit Lung Metastasis of Breast Cancer Reducing Cancer Stemness. Eur. J. Pharmacol. 2019, 859, 172499. [Google Scholar] [CrossRef] [PubMed]
- Weissenrieder, J.S.; Neighbors, J.D.; Mailman, R.B.; Hohl, R.J. Cancer and the Dopamine D2 Receptor: A Pharmacological Perspective. J. Pharmacol. Exp. Ther. 2019, 370, 111–126. [Google Scholar] [CrossRef]
- Liu, H.; Ma, L.; Li, C.; Cao, B.; Jiang, Y.; Han, L.; Xu, R.; Lin, J.; Zhang, D. The Molecular Mechanism of Chronic Stress Affecting the Occurrence and Development of Breast Cancer and Potential Drug Therapy. Transl. Oncol. 2022, 15, 101281. [Google Scholar] [CrossRef]
- Sousa, D.M.; Fernandes, V.; Lourenço, C.; Carvalho-Maia, C.; Estevão-Pereira, H.; Lobo, J.; Cantante, M.; Couto, M.; Conceição, F.; Jerónimo, C.; et al. Profiling the Adrenergic System in Breast Cancer and the Development of Metastasis. Cancers 2022, 14, 5518. [Google Scholar] [CrossRef]
- Rivero, E.M.; Martinez, L.M.; Bruque, C.D.; Gargiulo, L.; Bruzzone, A.; Lüthy, I.A. Prognostic Significance of α- and Β2-Adrenoceptor Gene Expression in Breast Cancer Patients. Br. J. Clin. Pharmacol. 2019, 85, 2143–2154. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Sturgeon, S.R.; Hankinson, S.E.; Shadyab, A.H.; Wallace, R.B.; Reeves, K.W. Psychotropic Medication Use and Postmenopausal Breast Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2020, 29, 254–256. [Google Scholar] [CrossRef]
- Rosas-Cruz, A.; Salinas-Jazmín, N.; Velázquez, M.A.V. Dopamine Receptors in Cancer: Are They Valid Therapeutic Targets? Technol. Cancer Res. Treat. 2021, 20, 15330338211027912. [Google Scholar] [CrossRef] [PubMed]
- Dakir, E.-H.; Pickard, A.; Srivastava, K.; McCrudden, C.M.; Gross, S.R.; Lloyd, S.; Zhang, S.-D.; Margariti, A.; Morgan, R.; Rudland, P.S.; et al. The Anti-Psychotic Drug Pimozide Is a Novel Chemotherapeutic for Breast Cancer. Oncotarget 2018, 9, 34889–34910. [Google Scholar] [CrossRef]
- Feng, Z.; Xia, Y.; Gao, T.; Xu, F.; Lei, Q.; Peng, C.; Yang, Y.; Xue, Q.; Hu, X.; Wang, Q.; et al. The Antipsychotic Agent Trifluoperazine Hydrochloride Suppresses Triple-Negative Breast Cancer Tumor Growth and Brain Metastasis by Inducing G0/G1 Arrest and Apoptosis. Cell Death Dis. 2018, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-E.; Lee, W.-Y.; Cheng, H.-W.; Chung, C.-H.; Mi, F.-L.; Lin, C.-W. The Antipsychotic Chlorpromazine Suppresses YAP Signaling, Stemness Properties, and Drug Resistance in Breast Cancer Cells. Chem. Biol. Interact. 2019, 302, 28–35. [Google Scholar] [CrossRef]
- Tegowski, M.; Fan, C.; Baldwin, A.S. Thioridazine Inhibits Self-Renewal in Breast Cancer Cells via DRD2-Dependent STAT3 Inhibition, but Induces a G1 Arrest Independent of DRD2. J. Biol. Chem. 2018, 293, 15977–15990. [Google Scholar] [CrossRef]
- Warchal, S.J.; Dawson, J.C.; Shepherd, E.; Munro, A.F.; Hughes, R.E.; Makda, A.; Carragher, N.O. High Content Phenotypic Screening Identifies Serotonin Receptor Modulators with Selective Activity upon Breast Cancer Cell Cycle and Cytokine Signaling Pathways. Bioorganic Med. Chem. 2020, 28, 115209. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Fan, Y.; Chen, A.; Jalali, A.; Minami, K.; Ogawa, K.; Nakshatri, H.; Li, B.-Y.; Yokota, H. Osteocyte-Driven Downregulation of Snail Restrains Effects of Drd2 Inhibitors on Mammary Tumor Cells. Cancer Res. 2018, 78, 3865–3876. [Google Scholar] [CrossRef] [PubMed]
- Pornour, M.; Ahangari, G.H.; Hejazi, S.; Deezagi, A. New Perspective Therapy of Breast Cancer Based on Selective Dopamine Receptor D2 Agonist and Antagonist Effects on MCF-7 Cell Line. Recent Pat. Anti-Cancer Drug Discov. 2015, 10, 214–223. [Google Scholar] [CrossRef]
- Heffernan, M.L.R.; Herman, L.W.; Brown, S.; Jones, P.G.; Shao, L.; Hewitt, M.C.; Campbell, J.E.; Dedic, N.; Hopkins, S.C.; Koblan, K.S.; et al. Ulotaront: A TAAR1 Agonist for the Treatment of Schizophrenia. ACS Med. Chem. Lett. 2021, 13, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, S.C.; Ogirala, A.; Worden, M.; Koblan, K.S. Depicting Safety Profile of TAAR1 Agonist Ulotaront Relative to Reactions Anticipated for a Dopamine D2-Based Pharmacological Class in FAERS. Clin. Drug Investig. 2021, 41, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Haibe-Kains, B.; Desmedt, C.; Loi, S.; Culhane, A.C.; Bontempi, G.; Quackenbush, J.; Sotiriou, C. A Three-Gene Model to Robustly Identify Breast Cancer Molecular Subtypes. JNCI J. Natl. Cancer Inst. 2012, 104, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Moreno, P.; Manning, J.; Fuentes, A.M.-P.; George, N.; Fexova, S.; Fonseca, N.A.; Füllgrabe, A.; Green, M.; Huang, N.; et al. Expression Atlas Update: From Tissues to Single Cells. Nucleic Acids Res. 2020, 48, D77–D83. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Da Silveira, W.A.; Hazard, E.S.; Chung, D.; Hardiman, G. Molecular Profiling of RNA Tumors Using High-Throughput RNA Sequencing: From Raw Data to Systems Level Analyses. In Tumor Profiling; Humana Press: New York, NY, USA, 2019; pp. 185–204. [Google Scholar]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- About GEO2R-GEO-NCBI. Available online: https://www.ncbi.nlm.nih.gov/geo/info/geo2r.html#microarray (accessed on 25 August 2023).
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Statistics for Biology and Health; Springer: New York, NY, USA, 2000; ISBN 978-1-4419-3161-0. [Google Scholar]
- Mulla, W.; Hajaj, B.; Elyagon, S.; Mor, M.; Gillis, R.; Murninkas, M.; Klapper-Goldstein, H.; Plaschkes, I.; Chalifa-Caspi, V.; Etzion, S.; et al. Rapid Atrial Pacing Promotes Atrial Fibrillation Substrate in Unanesthetized Instrumented Rats. Front. Physiol. 2019, 10, 1218. [Google Scholar] [CrossRef]
- Kernagis, D.N.; Hall, A.H.S.; Datto, M.B. Genes with Bimodal Expression Are Robust Diagnostic Targets That Define Distinct Subtypes of Epithelial Ovarian Cancer with Different Overall Survival. J. Mol. Diagn. 2012, 14, 214–222. [Google Scholar] [CrossRef]
- Huang, R.S.; Duan, S.; Shukla, S.J.; Kistner, E.O.; Clark, T.A.; Chen, T.X.; Schweitzer, A.C.; Blume, J.E.; Dolan, M.E. Identification of Genetic Variants Contributing to Cisplatin-Induced Cytotoxicity by Use of a Genomewide Approach. Am. J. Hum. Genet. 2007, 81, 427–437. [Google Scholar] [CrossRef]
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG Mapping Tools for Uncovering Hidden Features in Biological Data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Network. Comprehensive Molecular Portraits of Human Breast Tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene Expression Patterns of Breast Carcinomas Distinguish Tumor Subclasses with Clinical Implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Pitts, M.S.; McShane, J.N.; Hoener, M.C.; Christian, S.L.; Berry, M.D. TAAR1 Levels and Sub-Cellular Distribution Are Cell Line but Not Breast Cancer Subtype Specific. Histochem. Cell Biol. 2019, 152, 155–166. [Google Scholar] [CrossRef]
- Mathias, C.; Zambalde, E.P.; Rask, P.; Gradia, D.F.; de Oliveira, J.C. Long Non-Coding RNAs Differential Expression in Breast Cancer Subtypes: What Do We Know? Clin. Genet. 2019, 95, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Kothari, C.; Osseni, M.A.; Agbo, L.; Ouellette, G.; Déraspe, M.; Laviolette, F.; Corbeil, J.; Lambert, J.-P.; Diorio, C.; Durocher, F. Machine Learning Analysis Identifies Genes Differentiating Triple Negative Breast Cancers. Sci. Rep. 2020, 10, 10464. [Google Scholar] [CrossRef]
- Garcia-Recio, S.; Hinoue, T.; Wheeler, G.L.; Kelly, B.J.; Garrido-Castro, A.C.; Pascual, T.; De Cubas, A.A.; Xia, Y.; Felsheim, B.M.; McClure, M.B.; et al. Multiomics in Primary and Metastatic Breast Tumors from the AURORA US Network Finds Microenvironment and Epigenetic Drivers of Metastasis. Nat. Cancer 2022, 4, 128–147. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Niikura, N.; Ogiya, R.; Yasojima, H.; Watanabe, K.; Kanbayashi, C.; Tsuneizumi, M.; Matsui, A.; Fujisawa, T.; Iwasa, T.; et al. Distinct Gene Expression Profiles between Primary Breast Cancers and Brain Metastases from Pair-Matched Samples. Sci. Rep. 2019, 9, 13343. [Google Scholar] [CrossRef]
- Schochter, F.; Friedl, T.W.P.; de Gregorio, A.; Krause, S.; Huober, J.; Rack, B.; Janni, W. Are Circulating Tumor Cells (CTCs) Ready for Clinical Use in Breast Cancer? An Overview of Completed and Ongoing Trials Using CTCs for Clinical Treatment Decisions. Cells 2019, 8, 1412. [Google Scholar] [CrossRef]
- Cristofanilli, M.; Pierga, J.-Y.; Reuben, J.; Rademaker, A.; Davis, A.A.; Peeters, D.J.; Fehm, T.; Nolé, F.; Gisbert-Criado, R.; Mavroudis, D.; et al. The Clinical Use of Circulating Tumor Cells (CTCs) Enumeration for Staging of Metastatic Breast Cancer (MBC): International Expert Consensus Paper. Crit. Rev. Oncol. Hematol. 2019, 134, 39–45. [Google Scholar] [CrossRef]
- Lang, J.E.; Ring, A.; Porras, T.; Kaur, P.; Forte, V.A.; Mineyev, N.; Tripathy, D.; Press, M.F.; Campo, D. RNA-Seq of Circulating Tumor Cells in Stage II–III Breast Cancer. Ann. Surg. Oncol. 2018, 25, 2261–2270. [Google Scholar] [CrossRef]
- Fabisiewicz, A.; Szostakowska-Rodzos, M.; Zaczek, A.J.; Grzybowska, E.A. Circulating Tumor Cells in Early and Advanced Breast Cancer; Biology and Prognostic Value. Int. J. Mol. Sci. 2020, 21, 1671. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, H.; Song, X.; Yang, Q. Metastatic Heterogeneity of Breast Cancer: Molecular Mechanism and Potential Therapeutic Targets. Semin. Cancer Biol. 2020, 60, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Eiro, N.; Gonzalez, L.O.; Fraile, M.; Cid, S.; Schneider, J.; Vizoso, F.J. Breast Cancer Tumor Stroma: Cellular Components, Phenotypic Heterogeneity, Intercellular Communication, Prognostic Implications and Therapeutic Opportunities. Cancers 2019, 11, 664. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Keller, E.T.; Garfield, D.H.; Shen, K.; Wang, J. Stromal Cells in Tumor Microenvironment and Breast Cancer. Cancer Metastasis Rev. 2013, 32, 303–315. [Google Scholar] [CrossRef]
- Sebastian, A.; Hum, N.R.; Martin, K.A.; Gilmore, S.F.; Peran, I.; Byers, S.W.; Wheeler, E.K.; Coleman, M.A.; Loots, G.G. Single-Cell Transcriptomic Analysis of Tumor-Derived Fibroblasts and Normal Tissue-Resident Fibroblasts Reveals Fibroblast Heterogeneity in Breast Cancer. Cancers 2020, 12, 1307. [Google Scholar] [CrossRef] [PubMed]
- Itoh, G.; Takagane, K.; Fukushi, Y.; Kuriyama, S.; Umakoshi, M.; Goto, A.; Yanagihara, K.; Yashiro, M.; Tanaka, M. Cancer-associated Fibroblasts Educate Normal Fibroblasts to Facilitate Cancer Cell Spreading and T-cell Suppression. Mol. Oncol. 2022, 16, 166–187. [Google Scholar] [CrossRef] [PubMed]
- Dhage, S.; Ernlund, A.; Ruggles, K.; Axelrod, D.; Berman, R.; Roses, D.; Schneider, R.J. A Genomic Ruler to Assess Oncogenic Transition between Breast Tumor and Stroma. PLoS ONE 2018, 13, e0205602. [Google Scholar] [CrossRef]
- Babusyte, A.; Kotthoff, M.; Fiedler, J.; Krautwurst, D. Biogenic Amines Activate Blood Leukocytes via Trace Amine-Associated Receptors TAAR1 and TAAR2. J. Leukoc. Biol. 2013, 93, 387–394. [Google Scholar] [CrossRef]
- Christian, S.L.; Berry, M.D. Trace Amine-Associated Receptors as Novel Therapeutic Targets for Immunomodulatory Disorders. Front. Pharmacol. 2018, 9, 680. [Google Scholar] [CrossRef]
- Andersen, G.; Krautwurst, D. Trace Amine-Associated Receptors in the Cellular Immune System. In Trace Amines and Neurological Disorders; Elsevier: Amsterdam, The Netherlands, 2016; pp. 97–105. ISBN 978-0-12-803603-7. [Google Scholar]
- Vaganova, A.N.; Kuvarzin, S.R.; Sycheva, A.M.; Gainetdinov, R.R. Deregulation of Trace Amine-Associated Receptors (TAAR) Expression and Signaling Mode in Melanoma. Biomolecules 2022, 12, 114. [Google Scholar] [CrossRef]
- Maßberg, D.; Hatt, H. Human Olfactory Receptors: Novel Cellular Functions Outside of the Nose. Physiol. Rev. 2018, 98, 1739–1763. [Google Scholar] [CrossRef] [PubMed]
- Asadi, M.; Ahmadi, N.; Ahmadvand, S.; Jafari, A.A.; Safaei, A.; Erfani, N.; Ramezani, A. Investigation of Olfactory Receptor Family 51 Subfamily j Member 1 (OR51J1) Gene Susceptibility as a Potential Breast Cancer-Associated Biomarker. PLoS ONE 2021, 16, e0246752. [Google Scholar] [CrossRef] [PubMed]
- Kalra, S.; Mittal, A.; Gupta, K.; Singhal, V.; Gupta, A.; Mishra, T.; Naidu, S.; Sengupta, D.; Ahuja, G. Analysis of Single-Cell Transcriptomes Links Enrichment of Olfactory Receptors with Cancer Cell Differentiation Status and Prognosis. Commun. Biol. 2020, 3, 506. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Cho, H.J.; Lee, C.; Koo, J. Odorant Receptors in Cancer. BMB Rep. 2022, 55, 72–80. [Google Scholar] [CrossRef]
- Li, M.; Schweiger, M.W.; Ryan, D.J.; Nakano, I.; Carvalho, L.A.; Tannous, B.A. Olfactory Receptor 5B21 Drives Breast Cancer Metastasis. iScience 2021, 24, 103519. [Google Scholar] [CrossRef]
- Weber, L.; Maßberg, D.; Becker, C.; Altmüller, J.; Ubrig, B.; Bonatz, G.; Wölk, G.; Philippou, S.; Tannapfel, A.; Hatt, H.; et al. Olfactory Receptors as Biomarkers in Human Breast Carcinoma Tissues. Front. Oncol. 2018, 8, 33. [Google Scholar] [CrossRef]
- Masjedi, S.; Zwiebel, L.J.; Giorgio, T.D. Olfactory Receptor Gene Abundance in Invasive Breast Carcinoma. Sci. Rep. 2019, 9, 13736. [Google Scholar] [CrossRef]
- Balakrishna, P.; George, S.; Hatoum, H.; Mukherjee, S. Serotonin Pathway in Cancer. Int. J. Mol. Sci. 2021, 22, 1268. [Google Scholar] [CrossRef]
- Pouya, F.D.; Rasmi, Y.; Asl, E.R. Role of Neurotransmitters and Neuropeptides in Breast Cancer Metastasis. Biochem. Mosc. Suppl. Ser. A 2020, 14, 107–116. [Google Scholar] [CrossRef]
- Olfati, Z.; Rigi, G.; Vaseghi, H.; Zamanzadeh, Z.; Sohrabi, M.; Hejazi, S.H. Evaluation of Serotonin Receptors (5HTR2A and 5HTR3A) MRNA Expression Changes in Tumor of Breast Cancer Patients. Med. J. Islam Repub. Iran 2020, 34, 99. [Google Scholar] [CrossRef]
- Shkurnikov, M.Y.; Galatenko, V.V.; Lebedev, A.E.; Podol’skii, V.E.; Tonevitskii, E.A.; Mal’tseva, D.V. On Statistical Relationship between ADRA2A Expression and the Risk of Breast Cancer Relapse. Bull. Exp. Biol. Med. 2014, 157, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, Z.; Zhang, L.; Hu, X.; Wang, Z.; Ni, H.; Wang, Y.; Qin, J. Activation of Β2-Adrenergic Receptor Promotes Growth and Angiogenesis in Breast Cancer by Down-Regulating PPARγ. Cancer Res. Treat. 2020, 52, 830–847. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Nogueira, P.; Bragado, P.; Almendro, V.; Ametller, E.; Rios, J.; Choudhury, S.; Mancino, M.; Gascón, P. Differential Expression of Neurogenes among Breast Cancer Subtypes Identifies High Risk Patients. Oncotarget 2015, 7, 5313–5326. [Google Scholar] [CrossRef] [PubMed]
- Massari, N.A.; Nicoud, M.B.; Medina, V.A. Histamine Receptors and Cancer Pharmacology: An Update. Br. J. Pharmacol. 2020, 177, 516–538. [Google Scholar] [CrossRef]
- Español, A.; Salem, A.; Sanchez, Y.; Sales, M.E. Breast Cancer: Muscarinic Receptors as New Targets for Tumor Therapy. World J. Clin. Oncol. 2021, 12, 404–428. [Google Scholar] [CrossRef]
- Reader, J.; Holt, D.; Fulton, A. Prostaglandin E2 EP Receptors as Therapeutic Targets in Breast Cancer. Cancer Metastasis Rev. 2011, 30, 449–463. [Google Scholar] [CrossRef]
- Sánchez, M.L.; Coveñas, R. The Galaninergic System: A Target for Cancer Treatment. Cancers 2022, 14, 3755. [Google Scholar] [CrossRef]
- Watt, H.L.; Kharmate, G.; Kumar, U. Biology of Somatostatin in Breast Cancer. Mol. Cell. Endocrinol. 2008, 286, 251–261. [Google Scholar] [CrossRef]
- Song, Y.; Li, L.; Chen, J.; Chen, H.; Cui, B.; Feng, Y.; Zhang, P.; Zhang, Q.; Xia, Y.; Luo, M. Thioridazine Hydrochloride: An Antipsychotic Agent That Inhibits Tumor Growth and Lung Metastasis in Triple-Negative Breast Cancer via Inducing G0/G1 Arrest and Apoptosis. Cell Cycle 2020, 19, 3521–3533. [Google Scholar] [CrossRef]
- Xie, Q.-E.; Du, X.; Wang, M.; Xie, F.; Zhang, Z.; Cao, Y.; Liu, Y. Identification of Serotonin as a Predictive Marker for Breast Cancer Patients. Int. J. Gen. Med. 2021, 14, 1939–1948. [Google Scholar] [CrossRef]
- Peters, M.A.M.; Meijer, C.; Fehrmann, R.S.N.; Walenkamp, A.M.E.; Kema, I.P.; de Vries, E.G.E.; Hollema, H.; Oosting, S.F. Serotonin and Dopamine Receptor Expression in Solid Tumours Including Rare Cancers. Pathol. Oncol. Res. 2020, 26, 1539–1547. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset ID | Title | n | Samples’ Characteristics |
|---|---|---|---|
| GSE113890 | The whole transcriptional landscape of circulating tumor cells compared to metastases in stage IV breast cancer | 45 | Stage IV patients, 21 circulating tumor cells samples and 24 metastatic lesions were analyzed |
| GSE119937 | Molecular Determinants of Post-Mastectomy Breast Cancer Recurrence | 118 1 | Primary breast cancer samples, grade I–III (refer to Table S1 for details) |
| GSE184717 | Novel temporal and spatial patterns of metastatic colonization from breast cancer rapid-autopsy tumor biopsies | 28 | Breast cancer metastatic lesions |
| Dataset ID | Title | n | Samples Characteristics | Platform |
|---|---|---|---|---|
| GSE5847 | Tumor and stroma from breast by LCM | 95 | 47 stromal microdissected samples, 48 epithelial microdissected samples from 15 invasive and 35 non-invasive breast cancers | Affymetrix Human Genome U133A Array |
| GSE20685 | Microarray-based molecular subtyping of breast cancer | 327 | Primary breast cancer samples, Stage I–III (refer to Table S1 for details) | Affymetrix Human Genome U133 Plus 2.0 Array |
| GSE25066 | Genomic predictor of response and survival following neoadjuvant taxane-anthracycline chemotherapy in breast cancer | 508 | Primary breast cancer samples, Stage I–III (refer to Table S1 for details) | Affymetrix Human Genome U133A Array |
| GSE58215 | Integrated analysis reveals microRNA networks coordinately expressed with key proteins in breast cancer | 566 | Primary breast cancer samples, grade I–III (refer to Table S1 for details) | Agilent-028004 SurePrint G3 Human GE 8x60K Microarray |
| GSE80999 | Integrative clustering reveals a novel split in the luminal A subtype of breast cancer with impact on outcome | 356 | Primary breast cancer samples, grade I–III (refer to Table S1 for details) | Agilent-028004 SurePrint G3 Human GE 8x60K Microarray |
| GSE88715 | Gene expression profiles of microdissected tumor epithelium and stroma from TN breast tumors | 76 | 38 triple negative breast cancer samples, tissue compartments isolated by laser capture microdissection | Agilent-028004 SurePrint G3 Human GE 8x60K Microarray |
| GSE102484 | Expression data from invasive breast cancer patient | 683 | Primary breast cancer samples, Stage I–III (refer to Table S1 for details) | Affymetrix Human Genome U133 Plus 2.0 Array |
| GSE131769 | Genomic signature of the standardized uptake value in 18F-fluorodeoxyglucose positron emission tomography in breast cancer | 301 | Surgically resected breast cancer, Stage I–III (refer to Table S1 for details) | Illumina HumanHT-12 V3.0 expression beadchip |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaganova, A.N.; Maslennikova, D.D.; Konstantinova, V.V.; Kanov, E.V.; Gainetdinov, R.R. The Expression of Trace Amine-Associated Receptors (TAARs) in Breast Cancer Is Coincident with the Expression of Neuroactive Ligand–Receptor Systems and Depends on Tumor Intrinsic Subtype. Biomolecules 2023, 13, 1361. https://doi.org/10.3390/biom13091361
Vaganova AN, Maslennikova DD, Konstantinova VV, Kanov EV, Gainetdinov RR. The Expression of Trace Amine-Associated Receptors (TAARs) in Breast Cancer Is Coincident with the Expression of Neuroactive Ligand–Receptor Systems and Depends on Tumor Intrinsic Subtype. Biomolecules. 2023; 13(9):1361. https://doi.org/10.3390/biom13091361
Chicago/Turabian StyleVaganova, Anastasia N., Daria D. Maslennikova, Valeria V. Konstantinova, Evgeny V. Kanov, and Raul R. Gainetdinov. 2023. "The Expression of Trace Amine-Associated Receptors (TAARs) in Breast Cancer Is Coincident with the Expression of Neuroactive Ligand–Receptor Systems and Depends on Tumor Intrinsic Subtype" Biomolecules 13, no. 9: 1361. https://doi.org/10.3390/biom13091361