
Aquaporin-1 and Aquaporin-4 Expression in Ependyma, Choroid Plexus and Surrounding Transition Zones in the Human Brain
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Post-Mortem Specimens
2.2. Immunohistochemistry
2.3. Light Microscopy
2.4. Electron Microscopy
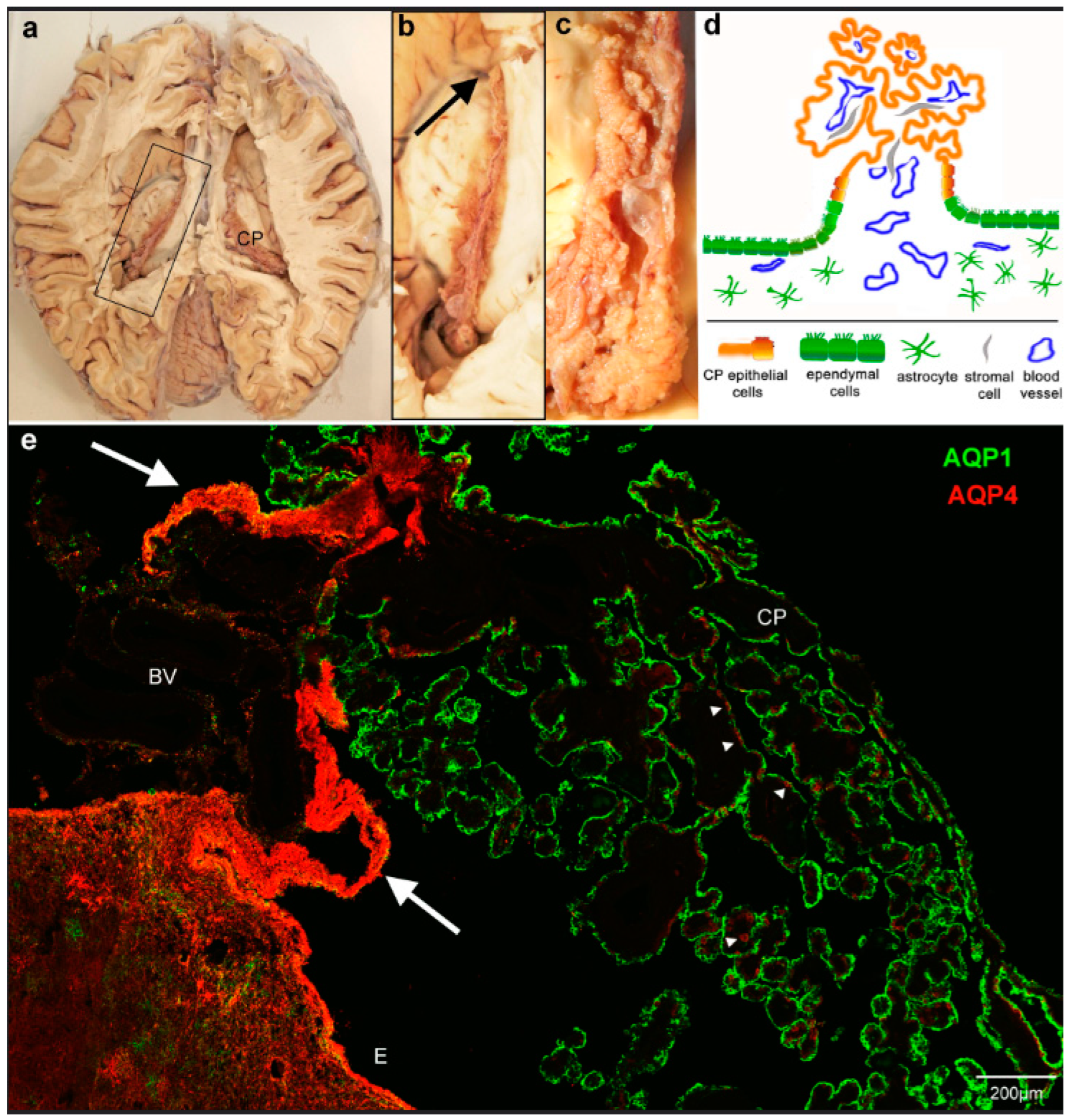
3. Results
4. Discussion
4.1. Topography of the Transitional Glial Plate

4.2. Astrocytic Polarity in the Transitional Glial Plate
4.3. Functional Implications of High AQP4 Expression in the Transitional Glial Plate
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Denker, B.M.; Smith, B.L.; Kuhajda, F.P.; Agre, P. Identification, Purification, and Partial Characterization of a Novel Mr 28,000 Integral Membrane-Protein from Erythrocytes and Renal Tubules. J. Biol. Chem. 1988, 263, 15634–15642. [Google Scholar] [CrossRef] [PubMed]
- Frigeri, A.; Gropper, M.A.; Turck, C.W.; Verkman, A.S. Immunolocalization of the Mercurial-Insensitive Water Channel and Glycerol Intrinsic Protein in Epithelial-Cell Plasma-Membranes. Proc. Natl. Acad. Sci. USA 1995, 92, 4328–4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.S.; Bhat, R.V.; Preston, G.M.; Guggino, W.B.; Baraban, J.M.; Agre, P. Molecular characterization of an aquaporin cDNA from brain: Candidate osmoreceptor and regulator of water balance. Proc. Natl. Acad. Sci. USA 1994, 91, 13052–13056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, S.; Smith, B.L.; Christensen, E.I.; Agre, P. Distribution of the Aquaporin Chip in Secretory and Resorptive Epithelia and Capillary Endothelia. Proc. Natl. Acad. Sci. USA 1993, 90, 7275–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagelhus, E.A.; Ottersen, O.P. Physiological roles of aquaporin-4 in brain. Physiol. Rev. 2013, 93, 1543–1562. [Google Scholar] [CrossRef] [Green Version]
- Rash, J.E.; Yasumura, T.; Hudson, C.S.; Agre, P.; Nielsen, S. Direct immunogold labeling of aquaporin-4 in square arrays of astrocyte and ependymocyte plasma membranes in rat brain and spinal cord. Proc. Natl. Acad. Sci. USA 1998, 95, 11981–11986. [Google Scholar] [CrossRef] [Green Version]
- Dermietzel, R. Junctions in the Central Nervous System of the Cat. III. Gap Junctions and Membrane-associated Orthogonal Particle Complexes (MOPC) in Astrocytic Membranes. Cell Tissue Res. 1974, 149, 121–135. [Google Scholar]
- Landis, D.M.D.; Reese, T.S. Arrays of particles in freeze-fractured astrocytic membranes. J. Cell Biol. 1974, 60, 316–320. [Google Scholar] [CrossRef] [Green Version]
- Wolburg, H.; Wolburg-Buchholz, K.; Fallier-Becker, P.; Noell, S.; Mack, A.F. Structure and Functions of Aquaporin-4-Based Orthogonal Arrays of Particles. Int. Rev. Cell Mol. Biol. 2011, 287, 1–41. [Google Scholar]
- Praetorius, J.; Nielsen, S. Distribution of sodium transporters and aquaporin-1 in the human choroid plexus. Am. J. Physiol.—Cell Physiol. 2006, 291, C59–C67. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.; Arnulf Nagelhus, E.; Amiry-Moghaddam, M.; Bourque, C.; Agre, P.; Petter Ottersen, O. Specialized Membrane Domains for Water Transport in Glial Cells: High-Resolution Immunogold Cytochemistry of Aquaporin-4 in Rat Brain. J. Neurosci. 1997, 17, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agre, P.; Nielsen, S.; Ottersen, O.P. Towards a molecular understanding of water homeostasis in the brain. Neuroscience 2004, 129, 849–850. [Google Scholar] [CrossRef] [PubMed]
- Hablitz, L.M.; Nedergaard, M. The Glymphatic System: A Novel Component of Fundamental Neurobiology. J. Neurosci. 2021, 41, 7698–7711. [Google Scholar] [CrossRef] [PubMed]
- Mack, A.F.; Wolburg, H. A Novel Look at Astrocytes: Aquaporins, Ionic Homeostasis, and the Role of the Microenvironment for Regeneration in the CNS. Neuroscientist 2013, 19, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Deffner, F.; Gleiser, C.; Mattheus, U.; Wagner, A.; Neckel, P.H.; Fallier-Becker, P.; Hirt, B.; Mack, A.F. Aquaporin-4 expression in the human choroid plexus. Cell. Mol. Life Sci. 2022, 79, 90. [Google Scholar] [CrossRef]
- Guadagno, E.; Moukhles, H. Laminin-induced aggregation of the inwardly rectifying potassium channel, Kir4.1, and the water-permeable channel, AQP4, via a dystroglycan-containing complex in astrocytes. Glia 2004, 47, 138–149. [Google Scholar] [CrossRef]
- Hoddevik, E.H.; Rao, S.B.; Zahl, S.; Boldt, H.B.; Ottersen, O.P.; Amiry-Moghaddam, M. Organisation of extracellular matrix proteins laminin and agrin in pericapillary basal laminae in mouse brain. Brain Struct. Funct. 2020, 225, 805–816. [Google Scholar] [CrossRef] [Green Version]
- Mack, A.; Neuhaus, J.; Wolburg, H. Relationship between orthogonal arrays of particles and tight junctions as demonstrated in cells of the ventricular wall of the rat brain. Cell Tissue Res. 1987, 248, 619–625. [Google Scholar] [CrossRef]
- Wolburg, H.; Paulus, W. Choroid plexus: Biology and pathology. Acta Neuropathol. 2010, 119, 75–88. [Google Scholar] [CrossRef]
- Amiry-Moghaddam, M.; Frydenlund, D.S.; Ottersen, O.P. Anchoring of aquaporin-4 in brain: Molecular mechanisms and implications for the physiology and pathophysiology of water transport. Neuroscience 2004, 129, 997–1008. [Google Scholar] [CrossRef]
- Nagelhus, E.A.; Veruki, M.L.; Torp, R.; Haug, F.M.; Laake, J.H.; Nielsen, S.; Agre, P.; Ottersen, O.P. Aquaporin-4 water channel protein in the rat retina and optic nerve: Polarized expression in Muller cells and fibrous astrocytes. J. Neurosci. 1998, 18, 2506–2519. [Google Scholar] [CrossRef] [Green Version]
- Belmaati Cherkaoui, M.; Vacca, O.; Izabelle, C.; Boulay, A.-C.; Boulogne, C.; Gillet, C.; Barnier, J.-V.; Rendon, A.; Cohen-Salmon, M.; Vaillend, C. Dp71 contribution to the molecular scaffold anchoring aquaporine-4 channels in brain macroglial cells. Glia 2021, 69, 954–970. [Google Scholar] [CrossRef] [PubMed]
- Wolburg, H.; Noell, S.; Wolburg-Buchholz, K.; Mack, A.; Fallier-Becker, P. Agrin, Aquaporin-4, and Astrocyte Polarity as an Important Feature of the Blood-Brain Barrier. Neuroscientist 2009, 15, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Noell, S.; Fallier-Becker, P.; Deutsch, U.; Mack, A.; Wolburg, H. Agrin defines polarized distribution of orthogonal arrays of particles in astrocytes. Cell Tissue Res. 2009, 337, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Neely, J.D.; Amiry-Moghaddam, M.; Ottersen, O.P.; Froehner, S.C.; Agre, P.; Adams, M.E. Syntrophin-dependent expression and localization of Aquaporin-4 water channel protein. Proc. Natl. Acad. Sci. USA 2001, 98, 14108–14113. [Google Scholar] [CrossRef] [Green Version]
- Noël, G.; Tham, D.K.L.; Guadagno, E.; MacVicar, B.; Moukhles, H. The Laminin-Induced Phosphorylation of PKCδ Regulates AQP4 Distribution and Water Permeability in Rat Astrocytes. Cell. Mol. Neurobiol. 2021, 41, 1743–1757. [Google Scholar] [CrossRef]
- Rao, S.B.; Skauli, N.; Jovanovic, N.; Katoozi, S.; Frigeri, A.; Froehner, S.C.; Adams, M.E.; Ottersen, O.P.; Amiry-Moghaddam, M. Orchestrating aquaporin-4 and connexin-43 expression in brain: Differential roles of α1- and β1-syntrophin. Biochim. Biophys. Acta 2021, 1863, 183616. [Google Scholar] [CrossRef]
- Nicchia, G.P.; Rossi, A.; Nudel, U.; Svelto, M.; Frigeri, A. Dystrophin-dependent and -independent AQP4 pools are expressed in the mouse brain. Glia 2008, 56, 869–876. [Google Scholar] [CrossRef]
- Haenggi, T.; Soontornmalai, A.; Schaub, M.C.; Fritschy, J.M. The role of utrophin and Dp71 for assembly of different dystrophin-associated protein complexes (DPCS) in the choroid plexus and microvasculature of the brain. Neuroscience 2004, 129, 403–413. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Ghersi-Egea, J.F.; Strazielle, N.; Catala, M.; Silva-Vargas, V.; Doetsch, F.; Engelhardt, B. Molecular anatomy and functions of the choroidal blood-cerebrospinal fluid barrier in health and disease. Acta Neuropathol. 2018, 135, 337–361. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, F.; Mack, A.F.; Wolburg, H. Topological Aspects of the Blood-Brain and Blood-Cerebrospinal Fluid Barriers and Their Relevance in Inflammation. In Blood Brain Barrier and Inflammation; Lyck, R., Enzmann, G., Eds.; Springer: Cham, Switzerland, 2017; pp. 23–48. [Google Scholar] [CrossRef]
- Matyszak, M.K.; Lawson, L.J.; Perry, V.H.; Gordon, S. Stromal macrophages of the choroid plexus situated at an interface between the brain and peripheral immune system constitutively express major histocompatibility class II antigens. J. Neuroimmunol. 1992, 40, 173–181. [Google Scholar] [CrossRef]
- Mogk, S.; Meiwes, A.; Boßelmann, C.M.; Wolburg, H.; Duszenko, M. The lane to the brain: How African trypanosomes invade the CNS. Trends Parasitol. 2014, 30, 470–477. [Google Scholar] [CrossRef]
- Schwerk, C.; Tenenbaum, T.; Kim, K.S.; Schroten, H. The choroid plexus-a multi-role player during infectious diseases of the CNS. Front. Cell. Neurosci. 2015, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Deffner, F.; Scharr, M.; Klingenstein, S.; Klingenstein, M.; Milazzo, A.; Scherer, S.; Wagner, A.; Hirt, B.; Mack, A.F.; Neckel, P.H. Histological Evidence for the Enteric Nervous System and the Choroid Plexus as Alternative Routes of Neuroinvasion by SARS-CoV2. Front. Neuroanat. 2020, 14, 596439. [Google Scholar] [CrossRef]
- Brightman, M.W.; Reese, T.S. Junctions between intimately apposed cell membranes in the vertebrate brain. J. Cell Biol. 1969, 40, 648–677. [Google Scholar] [CrossRef]
- Alisch, J.S.R.; Kiely, M.; Triebswetter, C.; Alsameen, M.H.; Gong, Z.; Khattar, N.; Egan, J.M.; Bouhrara, M. Characterization of Age-Related Differences in the Human Choroid Plexus Volume, Microstructural Integrity, and Blood Perfusion Using Multiparameter Magnetic Resonance Imaging. Front. Aging Neurosci. 2021, 13, 734992. [Google Scholar] [CrossRef]
- Masseguin, C.; LePanse, S.; Corman, B.; Verbavatz, J.M.; Gabrion, J. Aging affects choroidal proteins involved in CSF production in Sprague-Dawley rats. Neurobiol. Aging 2005, 26, 917–927. [Google Scholar] [CrossRef]
- Chen, R.L.; Kassem, N.A.; Redzic, Z.B.; Chen, C.P.C.; Segal, M.B.; Preston, J.E. Age-related changes in choroid plexus and blood–cerebrospinal fluid barrier function in the sheep. Exp. Gerontol. 2009, 44, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Quintela, T.; Marcelino, H.; Deery, M.J.; Feret, R.; Howard, J.; Lilley, K.S.; Albuquerque, T.; Gonçalves, I.; Duarte, A.C.; Santos, C.R.A. Sex-Related Differences in Rat Choroid Plexus and Cerebrospinal Fluid: A cDNA Microarray and Proteomic Analysis. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef]
- Santos, C.R.A.; Duarte, A.C.; Quintela, T.; Tomas, J.; Albuquerque, T.; Marques, F.; Palha, J.A.; Goncalves, I. The choroid plexus as a sex hormone target: Functional implications. Front. Neuroendocrinol. 2017, 44, 103–121. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [Green Version]
- Mestre, H.; Du, T.; Sweeney, A.M.; Liu, G.; Samson, A.J.; Peng, W.; Mortensen, K.N.; Stæger, F.F.; Bork, P.A.R.; Bashford, L.; et al. Cerebrospinal fluid influx drives acute ischemic tissue swelling. Science 2020, 367, eaax7171. [Google Scholar] [CrossRef]
- Ishida, K.; Yamada, K.; Nishiyama, R.; Hashimoto, T.; Nishida, I.; Abe, Y.; Yasui, M.; Iwatsubo, T. Glymphatic system clears extracellular tau and protects from tau aggregation and neurodegeneration. J. Exp. Med. 2022, 219, e20211275. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary AB | |||
| AQP-4 | Santa Cruz sc-20812 | rabbit | 1:100 |
| AQP-4 | Santa Cruz sc-9888 | goat | 1:100 |
| AQP-1 | Thermo Fischer PA5-78805 | rabbit | 1:100 |
| AQP-1 | Santa Cruz, sc-32737 | mouse | 1:100 |
| Dystrophin | abcam ab15277 | rabbit | 1:100 |
| ß-Dystroglycan | abcam ab49515 | mouse | 1:100 |
| GFAP | Santa Cruz sc-58766 | mouse | 1:100 |
| Laminin | abcam ab11575 | rabbit | 1:100 |
| Secondary AB | |||
| Anti-mouse Alexa 488 | Thermofisher | goat | 1:400 |
| Anti-mouse Alexa 546 | Thermofisher | goat | 1:400 |
| Anti-rabbit Alexa 488 | Thermofisher | goat | 1:400 |
| Anti-rabbit Alexa 546 | Thermofisher | goat | 1:400 |
| Anti-rabbit Alexa 488 | Thermofisher | donkey | 1:400 |
| Anti-mouse Alexa 546 | Thermofisher | donkey | 1:400 |
| Anti-goat Alexa 660 | Thermofisher | donkey | 1:400 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bihlmaier, R.; Deffner, F.; Mattheus, U.; Neckel, P.H.; Hirt, B.; Mack, A.F. Aquaporin-1 and Aquaporin-4 Expression in Ependyma, Choroid Plexus and Surrounding Transition Zones in the Human Brain. Biomolecules 2023, 13, 212. https://doi.org/10.3390/biom13020212
Bihlmaier R, Deffner F, Mattheus U, Neckel PH, Hirt B, Mack AF. Aquaporin-1 and Aquaporin-4 Expression in Ependyma, Choroid Plexus and Surrounding Transition Zones in the Human Brain. Biomolecules. 2023; 13(2):212. https://doi.org/10.3390/biom13020212
Chicago/Turabian StyleBihlmaier, Ronja, Felix Deffner, Ulrich Mattheus, Peter H. Neckel, Bernhard Hirt, and Andreas F. Mack. 2023. "Aquaporin-1 and Aquaporin-4 Expression in Ependyma, Choroid Plexus and Surrounding Transition Zones in the Human Brain" Biomolecules 13, no. 2: 212. https://doi.org/10.3390/biom13020212