
Public Transcriptomic Data Meta-Analysis Demonstrates TAAR6 Expression in the Mental Disorder-Related Brain Areas in Human and Mouse Brain

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection and Inclusion Criteria for Datasets
2.2. Raw Data Normalization and Analysis
2.3. Gene Ontology Terms Enrichment Analysis of TAAR6 Co-Genes
2.4. Genotype-Tissue Expression (GTEx) Data
2.5. Public In Situ Hybridization (ISH) Data
2.6. Tissue Sampling and RNA Extraction
2.7. RT-PCR Analysis
3. Results
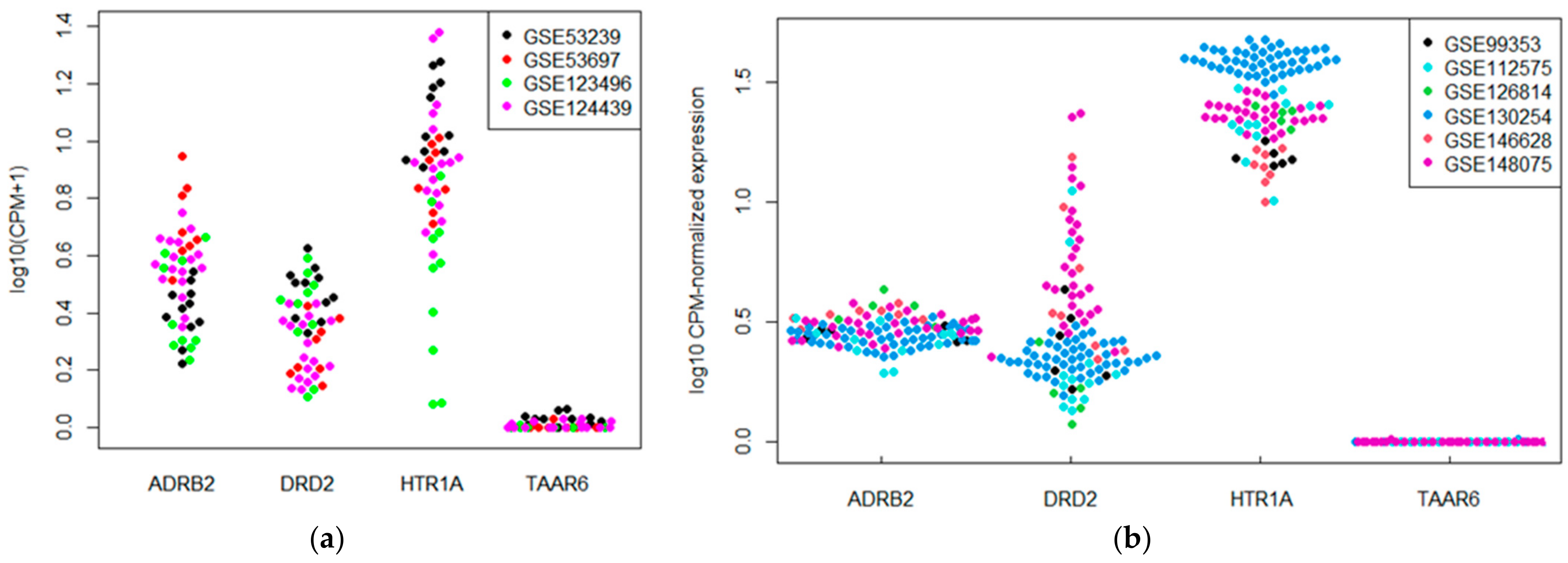
3.1. TAAR6 Expression in the Brain Neocortical Areas and Hippocampus in Datasets from GEO
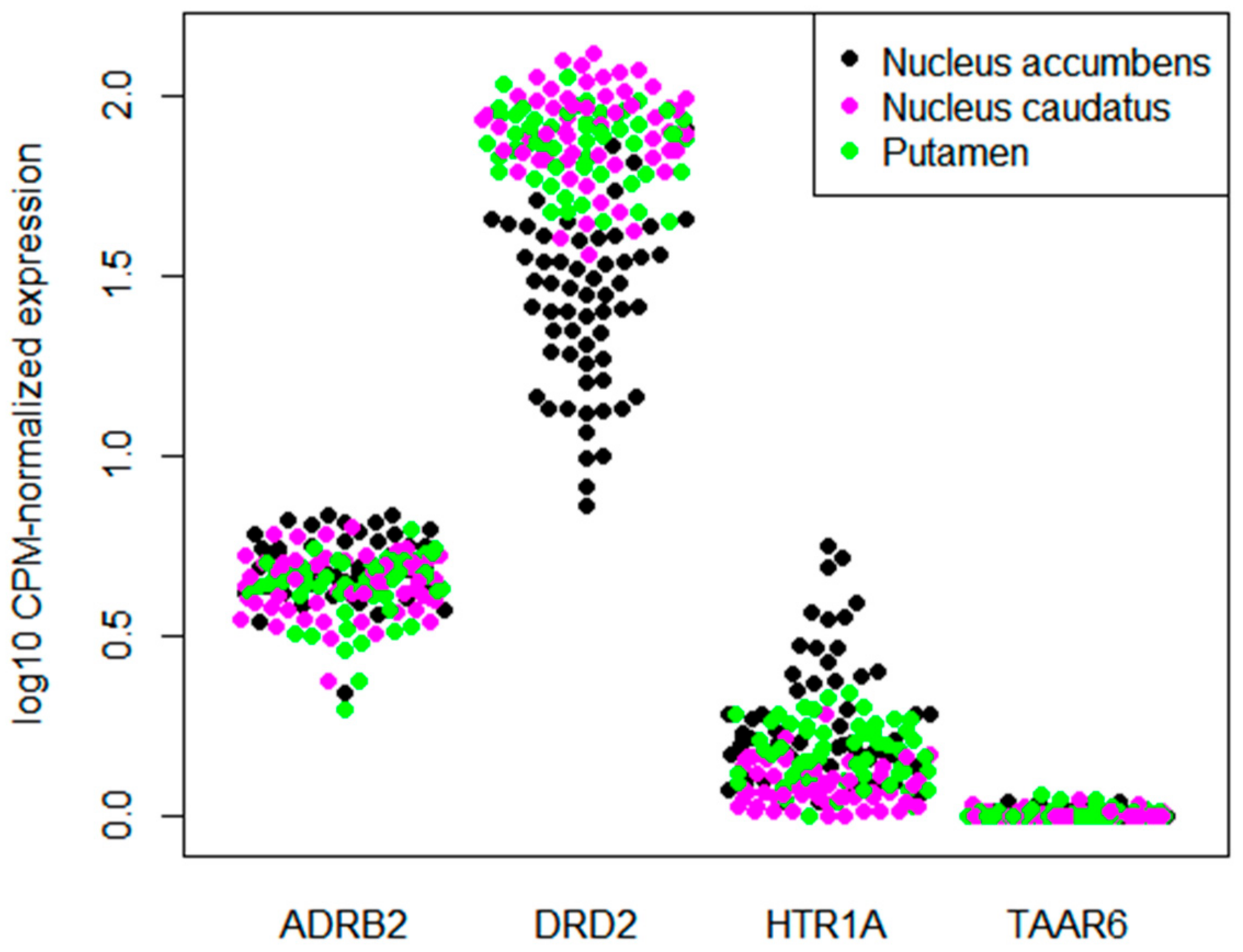
3.2. TAAR6 Expression in the Basal Ganglia in Datasets from GEO
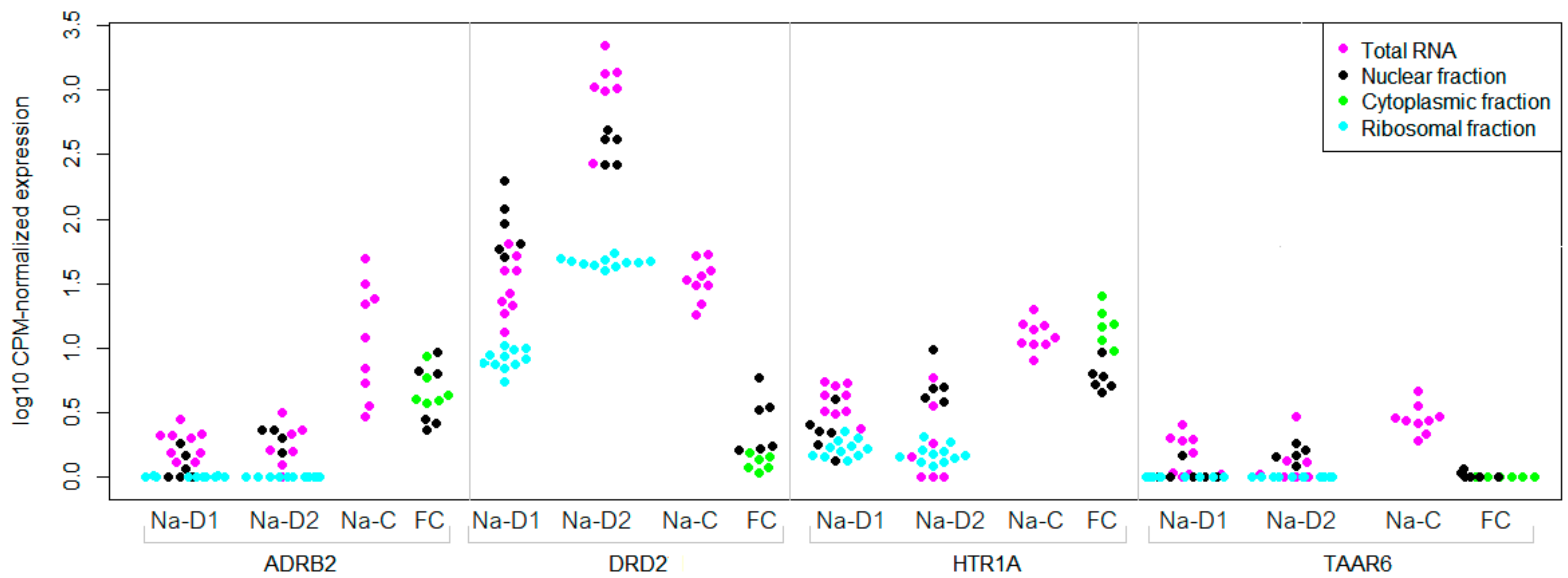
3.3. TAAR6 mRNA Distribution in the Brain Neuronal or Subcellular Fractions
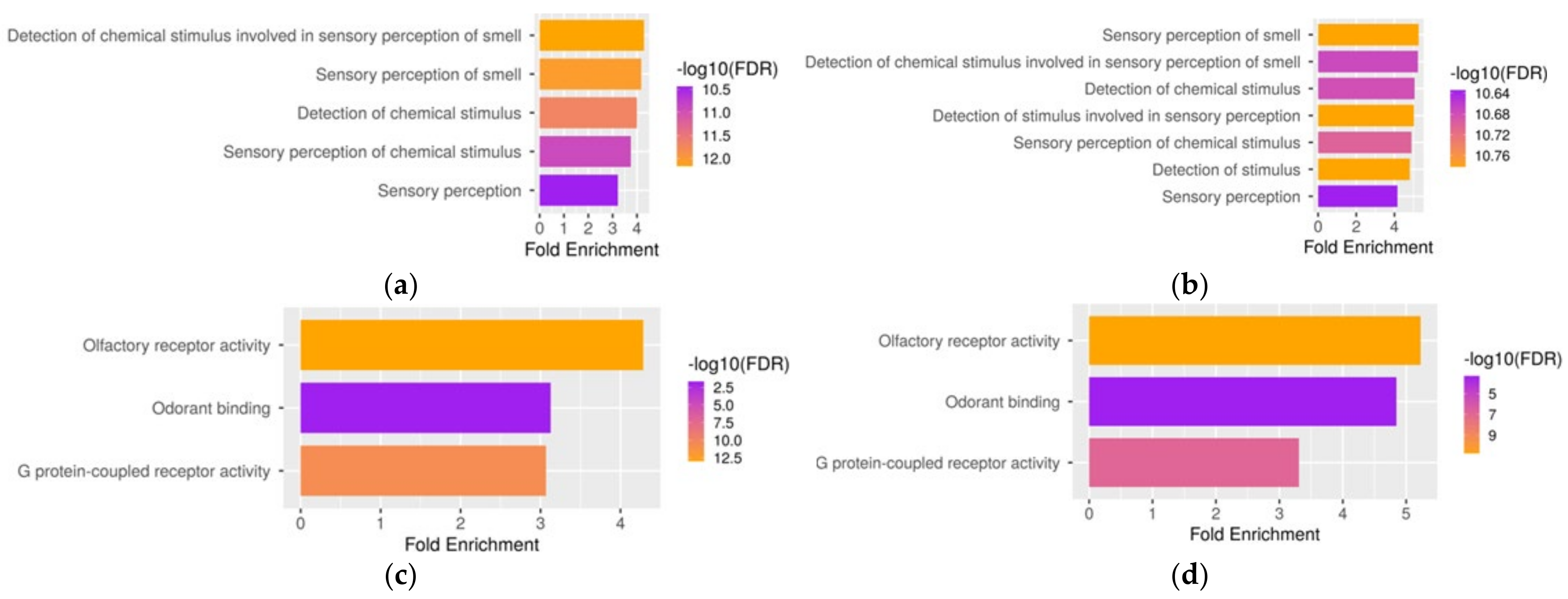
3.4. Functional Analysis of Genes Co-Expressed with TAAR6 in Different Neuronal Populations in the Mouse Nucleus Accumbens Samples
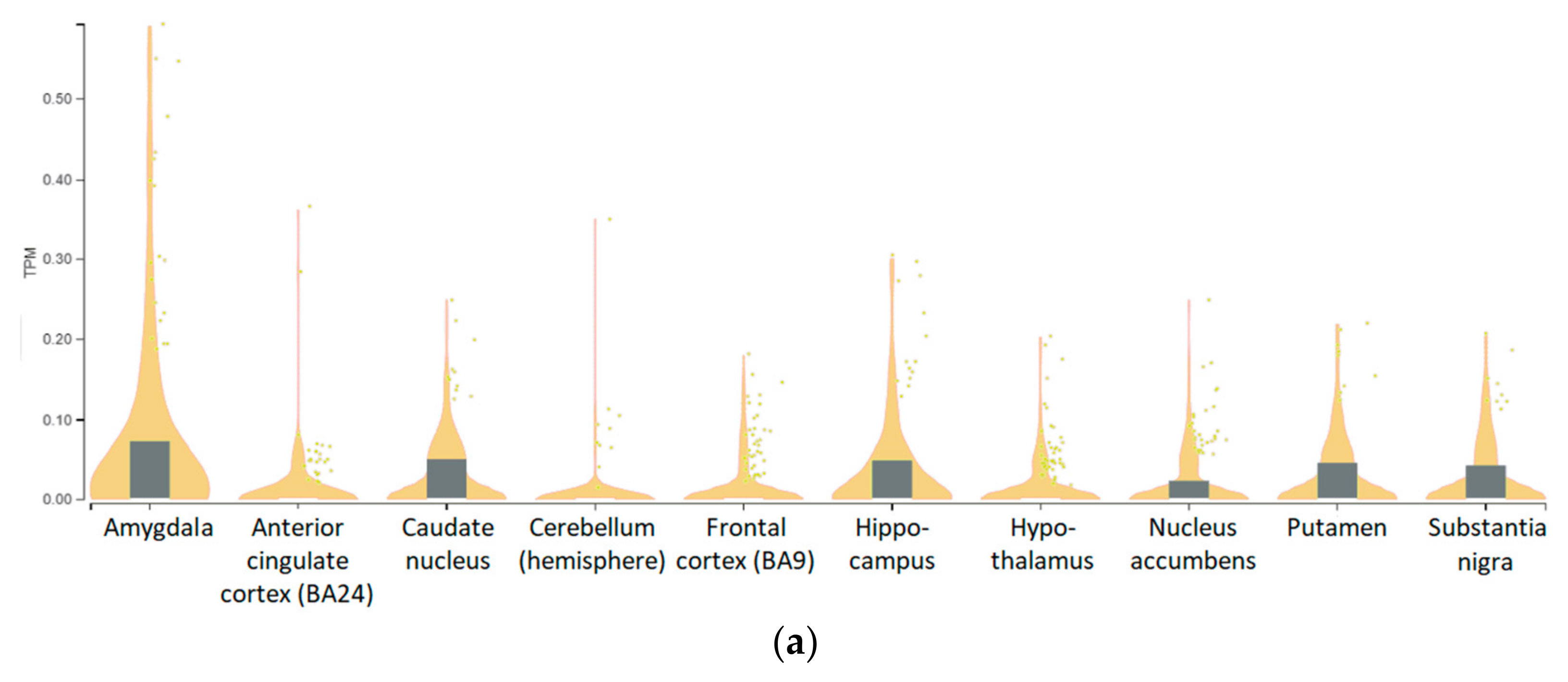
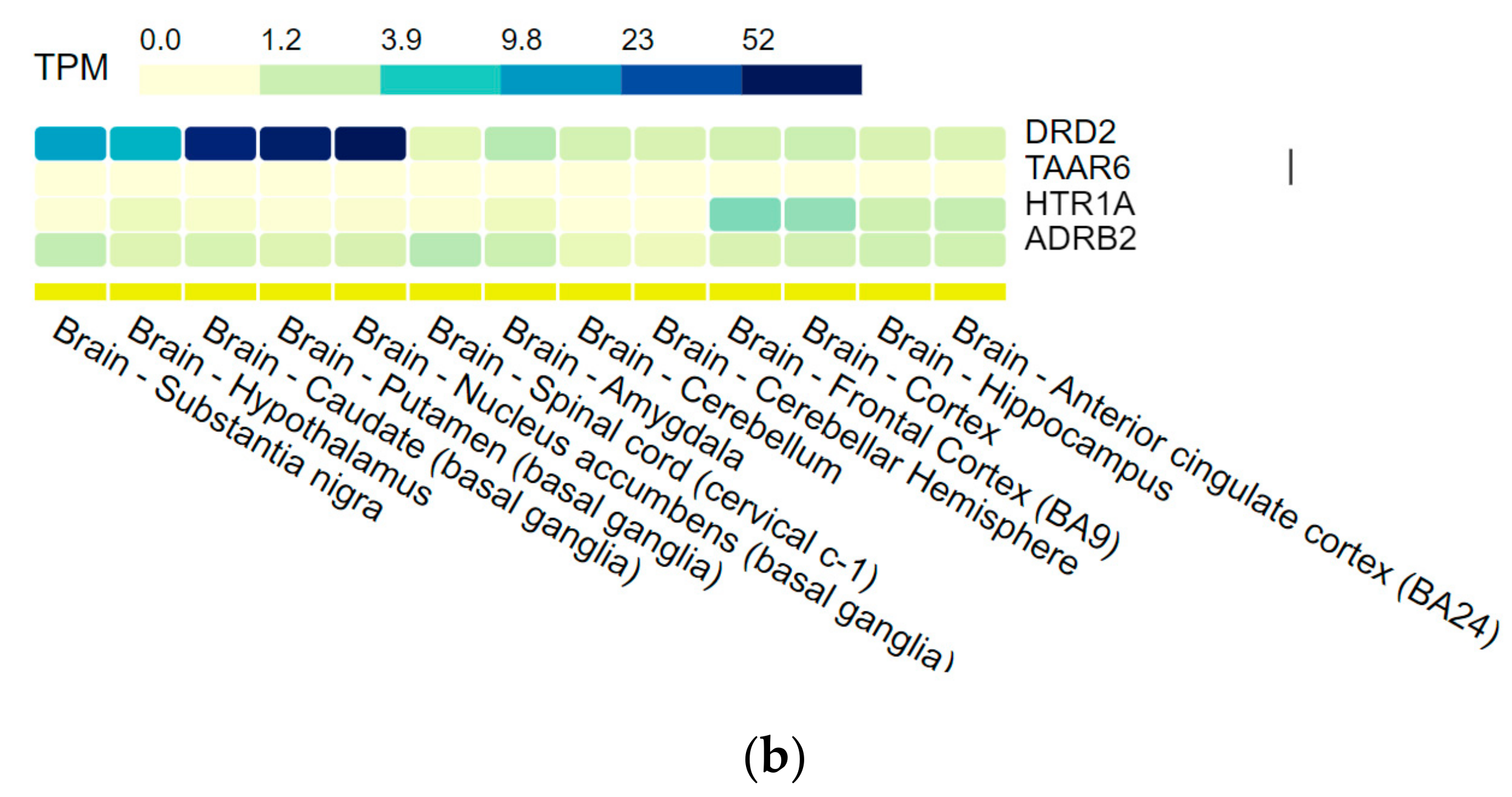
3.5. Genotype-Tissue Expression (GTEx) Data
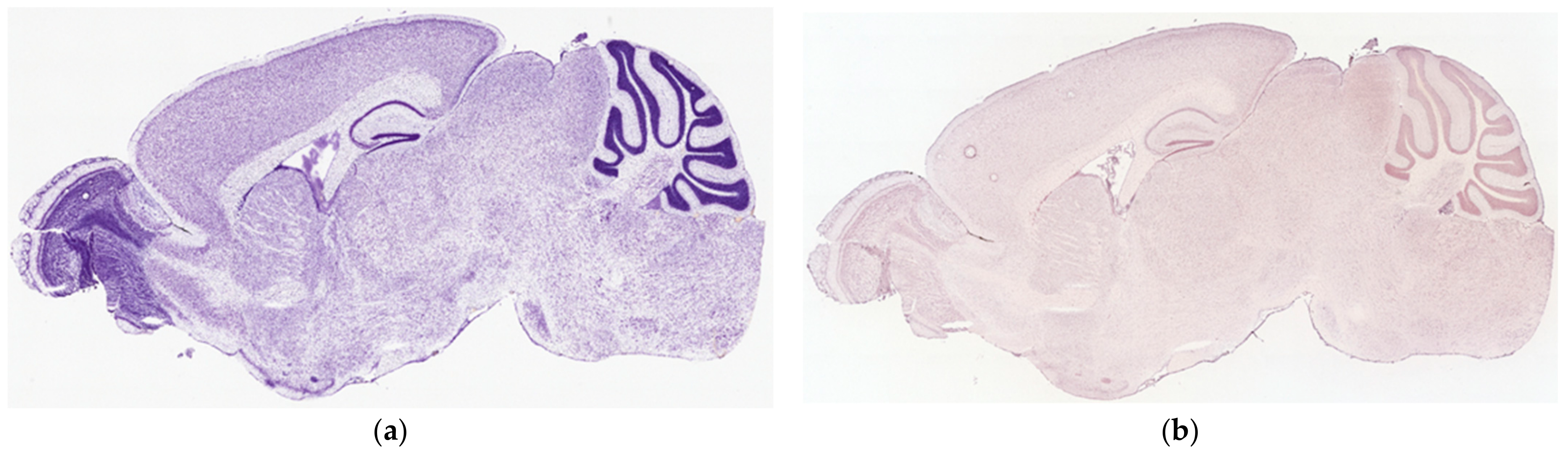
3.6. ISH Visualization of TAAR6 Expression in the Mouse Brain
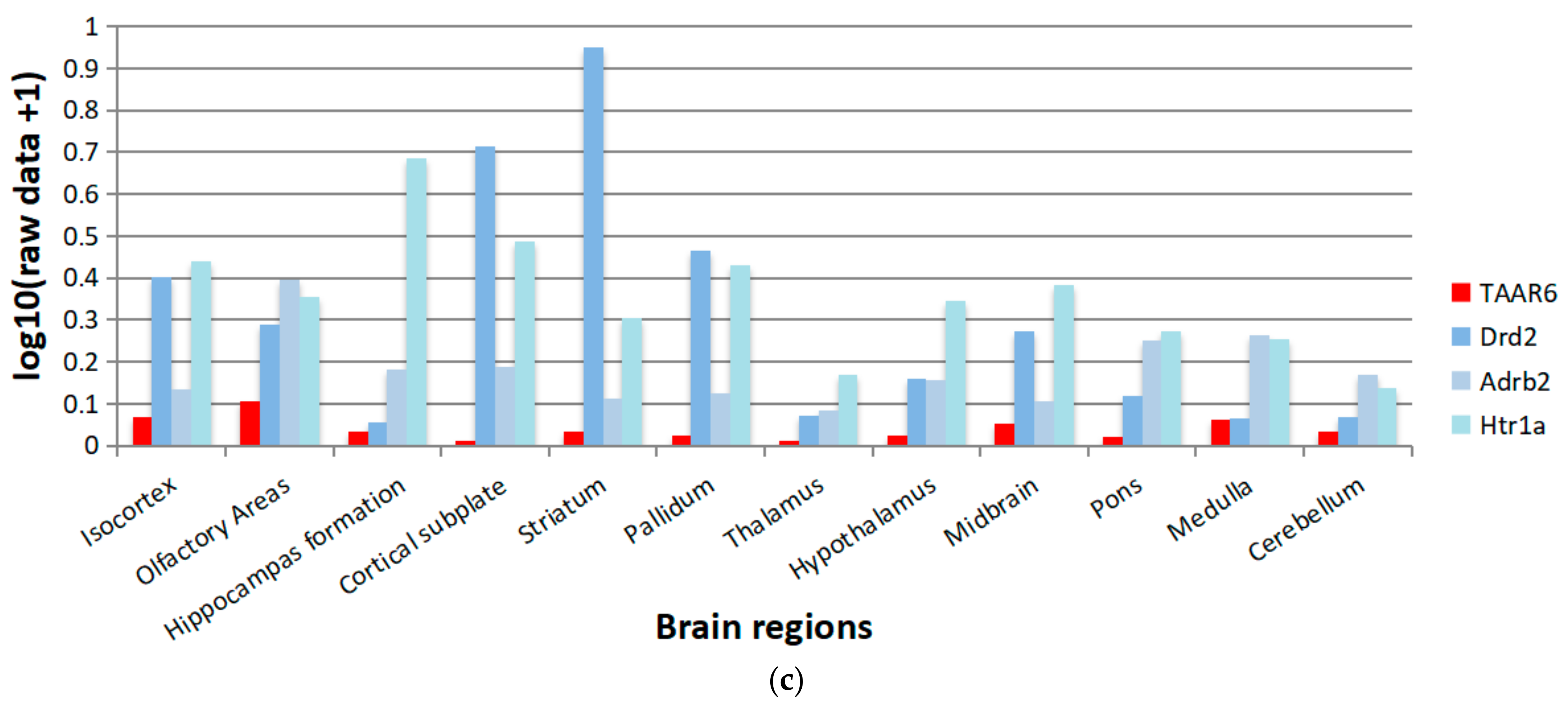
3.7. TAAR6 mRNA Expression in the Mouse Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boulton, A.A. Letter: Amines and Theories in Psychiatry. Lancet 1974, 2, 52–53. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef]
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of Human Trace Amine-Associated Receptors: Therapeutic Opportunities and Challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Sukhanov, I.; Efimova, E.V.; Kozlova, A.; Antonova, K.A.; Illiano, P.; Leo, D.; Merkulyeva, N.; Kalinina, D.; Musienko, P.; et al. Trace amine-associated receptor 5 provides olfactory input into limbic brain areas and modulates emotional behaviors and serotonin transmission. Front. Mol. Neurosci. 2020, 13, 18. [Google Scholar] [CrossRef]
- Efimova, E.V.; Katolikova, N.V.; Kanov, E.V.; Gainetdinov, R.R. Trace amine-associated receptors at the cross-road between innate olfaction of amines, emotions, and adult neurogenesis. Neural Regen. Res. 2021, 17, 1257–1258. [Google Scholar] [CrossRef]
- Kalinina, D.S.; Ptukha, M.A.; Goriainova, A.V.; Merkulyeva, N.S.; Kozlova, A.A.; Murtazina, R.Z.; Shemiakova, T.S.; Kuvarzin, S.R.; Vaganova, A.N.; Volnova, A.B.; et al. Role of the trace amine associated receptor 5 (TAAR5) in the sensorimotor functions. Sci. Rep. 2021, 11, 23092. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Kuvarzin, S.R.; Mor, M.S.; Katolikova, N.V.; Shemiakova, T.; Razenkova, V.; Ptukha, M.; Kozlova, A.A.; Murtazina, R.Z.; Smirnova, D.; et al. Trace amine-associated receptor 2 (TAAR2) is expressed in the limbic brain areas and is involved in dopamine regulation and adult neurogenesis. Front. Behav. Neurosci. 2022, 16, 847410. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Saraiva, L.R.; Korsching, S.I. Positive Darwinian Selection and the Birth of an Olfactory Receptor Clade in Teleosts. Proc. Natl. Acad. Sci. USA 2009, 106, 4313–4318. [Google Scholar] [CrossRef] [PubMed]
- Vallender, E.J.; Xie, Z.; Westmoreland, S.V.; Miller, G.M. Functional Evolution of the Trace Amine Associated Receptors in Mammals and the Loss of TAAR1 in Dogs. BMC Evol. Biol. 2010, 10, 51. [Google Scholar] [CrossRef]
- Glyakina, A.V.; Pavlov, C.D.; Sopova, J.V.; Gainetdinov, R.R.; Leonova, E.I.; Galzitskaya, O.V. Search for Structural Basis of Interactions of Biogenic Amines with Human TAAR1 and TAAR6 Receptors. Int. J. Mol. Sci. 2021, 23, 209. [Google Scholar] [CrossRef]
- Xie, Z.; Vallender, E.J.; Yu, N.; Kirstein, S.L.; Yang, H.; Bahn, M.E.; Westmoreland, S.V.; Miller, G.M. Cloning, Expression, and Functional Analysis of Rhesus Monkey Trace Amine-Associated Receptor 6: Evidence for Lack of Monoaminergic Association. J. Neurosci. Res. 2008, 86, 3435–3446. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, C.; Gómez-Tamayo, J.C.; Nebel, J.-C.; Pardo, L.; Gonzalez, A. Identifying Human Diamine Sensors for Death Related Putrescine and Cadaverine Molecules. PLoS Comput. Biol. 2018, 14, e1005945. [Google Scholar] [CrossRef]
- Ferrero, D.M.; Wacker, D.; Roque, M.A.; Baldwin, M.W.; Stevens, R.C.; Liberles, S.D. Agonists for 13 Trace Amine-Associated Receptors Provide Insight into the Molecular Basis of Odor Selectivity. ACS Chem. Biol. 2012, 7, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Dai, W.; Xu, Z.; Liang, Q.; Miller, E.; Li, S.; Gao, X.; Baldwin, M.; Chai, R.; Li, Q. Evolution of Brain-Expressed Biogenic Amine Receptors into Olfactory Trace Amine-Associated Receptors. Mol. Biol. Evol. 2022, 39, msac006. [Google Scholar] [CrossRef]
- Wallach, J. Endogenous Hallucinogens as Ligands of the Trace Amine Receptors: A Possible Role in Sensory Perception. Med. Hypotheses 2009, 72, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Serretti, A.; Pae, C.-U.; Chiesa, A.; Mandelli, L.; De Ronchi, D. Influence of TAAR6 Polymorphisms on Response to Aripiprazole. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Tahir, R.A.; Wu, H.; Javed, N.; Khalique, A.; Khan, S.A.F.; Mir, A.; Ahmed, M.S.; Barreto, G.E.; Qing, H.; Ashraf, G.M.; et al. Pharmacoinformatics and Molecular Docking Reveal Potential Drug Candidates against Schizophrenia to Target TAAR6. J. Cell. Physiol. 2019, 234, 13263–13276. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.; Ragoczy, T.; Lu, Z.; Kondoh, K.; Kuang, D.; Groudine, M.; Buck, L.B. Olfactory Receptor Genes Expressed in Distinct Lineages Are Sequestered in Different Nuclear Compartments. Proc. Natl. Acad. Sci. USA 2015, 112, E2403–E2409. [Google Scholar] [CrossRef]
- Fleischer, J.; Schwarzenbacher, K.; Breer, H. Expression of Trace Amine–Associated Receptors in the Grueneberg Ganglion. Chem. Senses 2007, 32, 623–631. [Google Scholar] [CrossRef]
- Johnson, M.A.; Tsai, L.; Roy, D.S.; Valenzuela, D.H.; Mosley, C.; Magklara, A.; Lomvardas, S.; Liberles, S.D.; Barnea, G. Neurons Expressing Trace Amine-Associated Receptors Project to Discrete Glomeruli and Constitute an Olfactory Subsystem. Proc. Natl. Acad. Sci. USA 2012, 109, 13410–13415. [Google Scholar] [CrossRef] [Green Version]
- Liberles, S.D.; Buck, L.B. A second class of chemosensory receptors in the olfactory epithelium. Nature 2006, 442, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace Amines: Identification of a Family of Mammalian G Protein-Coupled Receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Martinez, M.; Sanders, A.R.; Hou, C.; Saitou, N.; Kitano, T.; Mowry, B.J.; Crowe, R.R.; Silverman, J.M.; Levinson, D.F.; et al. Polymorphisms in the Trace Amine Receptor 4 (TRAR4) Gene on Chromosome 6q23.2 Are Associated with Susceptibility to Schizophrenia. Am. J. Hum. Genet. 2004, 75, 624–638. [Google Scholar] [CrossRef]
- Pae, C.-U.; Yu, H.-S.; Amann, D.; Kim, J.-J.; Lee, C.-U.; Lee, S.-J.; Jun, T.-Y.; Lee, C.; Paik, I.-H.; Patkar, A.A.; et al. Association of the Trace Amine Associated Receptor 6 (TAAR6) Gene with Schizophrenia and Bipolar Disorder in a Korean Case Control Sample. J. Psychiatr. Res. 2008, 42, 35–40. [Google Scholar] [CrossRef]
- Pae, C.-U.; Drago, A.; Patkar, A.A.; Jun, T.-Y.; Serretti, A. Epistasis between a Set of Variations Located in the TAAR6 and HSP-70 Genes toward Schizophrenia and Response to Antipsychotic Treatment. Eur. Neuropsychopharmacol. 2009, 19, 806–811. [Google Scholar] [CrossRef]
- Pae, C.-U.; Drago, A.; Forlani, M.; Patkar, A.A.; Serretti, A. Investigation of an Epistastic Effect between a Set of TAAR6 and HSP-70 Genes Variations and Major Mood Disorders. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153, 680–683. [Google Scholar] [CrossRef]
- Pae, C.-U.; Drago, A.; Mandelli, L.; De Ronchi, D.; Serretti, A. TAAR 6 and HSP-70 Variations Associated with Bipolar Disorder. Neurosci. Lett. 2009, 465, 257–261. [Google Scholar] [CrossRef]
- Pae, C.-U.; Drago, A.; Kim, J.-J.; Patkar, A.A.; Jun, T.-Y.; Lee, C.; Mandelli, L.; De Ronchi, D.; Paik, I.-H.; Serretti, A. TAAR6 Variation Effect on Clinic Presentation and Outcome in a Sample of Schizophrenic In-Patients: An Open Label Study. Eur. Psychiatry 2008, 23, 390–395. [Google Scholar] [CrossRef]
- Pae, C.-U.; Drago, A.; Kim, J.-J.; Patkar, A.A.; Jun, T.-Y.; De Ronchi, D.; Serretti, A. TAAR6 Variations Possibly Associated with Antidepressant Response and Suicidal Behavior. Psychiatry Res. 2010, 180, 20–24. [Google Scholar] [CrossRef]
- Vladimirov, V.; Thiselton, D.L.; Kuo, P.-H.; McClay, J.; Fanous, A.; Wormley, B.; Vittum, J.; Ribble, R.; Moher, B.; van den Oord, E.; et al. A Region of 35 Kb Containing the Trace Amine Associate Receptor 6 (TAAR6) Gene Is Associated with Schizophrenia in the Irish Study of High-Density Schizophrenia Families. Mol. Psychiatry 2007, 12, 842–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets--Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- GEO Browser-GEO-NCBI. Available online: https://www.ncbi.nlm.nih.gov/geo/browse/ (accessed on 23 March 2022).
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- GTEx Portal. Available online: https://www.gtexportal.org/home/ (accessed on 22 March 2022).
- Brain Map-Brain-Map.Org. Available online: https://portal.brain-map.org/ (accessed on 20 June 2022).
- ISH Data :: Allen Brain Atlas: Mouse Brain. Available online: https://mouse.brain-map.org/ (accessed on 16 March 2022).
- Daigle, T.L.; Madisen, L.; Hage, T.A.; Valley, M.T.; Knoblich, U.; Larsen, R.S.; Takeno, M.M.; Huang, L.; Gu, H.; Larsen, R.; et al. A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality. Cell 2018, 174, 465–480. [Google Scholar] [CrossRef]
- Harris, J.A.; Mihalas, S.; Hirokawa, K.E.; Whitesell, J.D.; Choi, H.; Bernard, A.; Bohn, P.; Caldejon, S.; Casal, L.; Cho, A.; et al. Hierarchical Organization of Cortical and Thalamic Connectivity. Nature 2019, 575, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Lein, E.S.; Hawrylycz, M.J.; Ao, N.; Ayres, M.; Bensinger, A.; Bernard, A.; Boe, A.F.; Boguski, M.S.; Brockway, K.S.; Byrnes, E.J.; et al. Genome-Wide Atlas of Gene Expression in the Adult Mouse Brain. Nature 2007, 445, 168–176. [Google Scholar] [CrossRef]
- Oh, S.W.; Harris, J.A.; Ng, L.; Winslow, B.; Cain, N.; Mihalas, S.; Wang, Q.; Lau, C.; Kuan, L.; Henry, A.M.; et al. A Mesoscale Connectome of the Mouse Brain. Nature 2014, 508, 207–214. [Google Scholar] [CrossRef]
- ISH Data :: Allen Brain Atlas: Human Brain. Available online: https://human.brain-map.org/ish/search (accessed on 23 March 2022).
- Experiment Detail :: Allen Brain Atlas: Mouse Brain. Available online: https://mouse.brain-map.org/experiment/show/70724989 (accessed on 16 March 2022).
- Experiment Detail :: Allen Brain Atlas: Mouse Brain. Available online: https://mouse.brain-map.org/experiment/show/68744522 (accessed on 16 March 2022).
- Experiment Detail :: Allen Brain Atlas: Mouse Brain. Available online: https://mouse.brain-map.org/experiment/show/358 (accessed on 23 March 2022).
- Experiment Detail :: Allen Brain Atlas: Mouse Brain. Available online: https://mouse.brain-map.org/experiment/show/79394355 (accessed on 23 March 2022).
- Liberles, S.D. Trace amine-associated receptors: Ligands, neural circuits, and behaviors. Curr. Opin. Neurobiol. 2015, 34, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zaghlool, A.; Niazi, A.; Björklund, Å.K.; Westholm, J.O.; Ameur, A.; Feuk, L. Characterization of the Nuclear and Cytosolic Transcriptomes in Human Brain Tissue Reveals New Insights into the Subcellular Distribution of RNA Transcripts. Sci. Rep. 2021, 11, 4076. [Google Scholar] [CrossRef] [PubMed]
- Vaganova, A.N.; Murtazina, R.Z.; Shemyakova, T.S.; Prjibelski, A.D.; Katolikova, N.V.; Gainetdinov, R.R. Pattern of TAAR5 Expression in the Human Brain Based on Transcriptome Datasets Analysis. Int. J. Mol. Sci. 2021, 22, 8802. [Google Scholar] [CrossRef]
- Khan, M.Z.; Nawaz, W. The Emerging Roles of Human Trace Amines and Human Trace Amine-Associated Receptors (HTAARs) in Central Nervous System. Biomed. Pharmacother. Biomed. Pharmacother. 2016, 83, 439–449. [Google Scholar] [CrossRef]
- Abou Jamra, R.; Sircar, I.; Becker, T.; Freudenberg-Hua, Y.; Ohlraun, S.; Freudenberg, J.; Brockschmidt, F.; Schulze, T.G.; Gross, M.; Spira, F.; et al. A Family-Based and Case–Control Association Study of Trace Amine Receptor Genes on Chromosome 6q23 in Bipolar Affective Disorder. Mol. Psychiatry 2005, 10, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Baumann, B.; Bogerts, B. Neuroanatomical Studies on Bipolar Disorder. Br. J. Psychiatry 2001, 178, s142–s147. [Google Scholar] [CrossRef]
- Rutigliano, G.; Zucchi, R. Molecular Variants in Human Trace Amine-Associated Receptors and Their Implications in Mental and Metabolic Disorders. Cell. Mol. Neurobiol. 2020, 40, 239–255. [Google Scholar] [CrossRef]
- Ikeda, M.; Iwata, N.; Suzuki, T.; Kitajima, T.; Yamanouchi, Y.; Kinoshita, Y.; Inada, T.; Ozaki, N. No Association of Haplotype-Tagging SNPs in TRAR4 with Schizophrenia in Japanese Patients. Schizophr. Res. 2005, 78, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Du, J.; Xu, Y.; Xing, Q.; Wang, H.; Wu, S.; Chen, Q.; Li, X.; Li, X.; Shen, J.; et al. Failure to Find Association between TRAR4 and Schizophrenia in the Chinese Han Population. J. Neural. Transm. 2006, 113, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Venken, T.; Alaerts, M.; Adolfsson, R.; Van Broeckhoven, C.; Del-Favero, J. No Association of Trace Amine-Associated Receptor 6 with Bipolar Disorder in a Northern Swedish Population. Psychiatr. Genet. 2006, 16, 1–2. [Google Scholar] [CrossRef]
- Vladimirov, V.I.; Maher, B.S.; Wormley, B.; O’Neill, F.A.; Walsh, D.; Kendler, K.S.; Riley, B.P. The Trace Amine Associated Receptor (TAAR6) Gene Is Not Associated with Schizophrenia in the Irish Case-Control Study of Schizophrenia (ICCSS) Sample. Schizophr. Res. 2009, 107, 249–254. [Google Scholar] [CrossRef]
- Xu, L.; Nan, J.; Lan, Y.; 2020. The Nucleus Accumbens: A Common Target in the Comorbidity of Depression and Addiction. Front. Neural Circuits 14. McCollum, L.A.; Roberts, R.C. Uncovering the Role of the Nucleus Accumbens in Schizophrenia: A Postmortem Analysis of Tyrosine Hydroxylase and Vesicular Glutamate Transporters. Schizophr. Res. 2015, 169, 369–373. [Google Scholar] [CrossRef]
- Manduca, A.; Servadio, M.; Damsteegt, R.; Campolongo, P.; Vanderschuren, L.J.; Trezza, V. Dopaminergic Neurotransmission in the Nucleus Accumbens Modulates Social Play Behavior in Rats. Neuropsychopharmacology 2016, 41, 2215–2223. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Aida, T.; Ito, H.; Kobayashi, K.; Wada, Y.; Kato, S.; Nakano, T.; Zhu, M.; Isa, K.; Isa, T.; et al. Dopaminergic Signaling in the Nucleus Accumbens Modulates Stress-Coping Strategies during Inescapable Stress. J. Neurosci. Off. J. Soc. Neurosci. 2020, 40, 7241–7254. [Google Scholar] [CrossRef] [PubMed]
- Soares-Cunha, C.; de Vasconcelos, N.A.P.; Coimbra, B.; Domingues, A.V.; Silva, J.M.; Loureiro-Campos, E.; Gaspar, R.; Sotiropoulos, I.; Sousa, N.; Rodrigues, A.J. Nucleus accumbens medium spiny neurons subtypes signal both reward and aversion. Mol. Psychiatry 2020, 25, 3241–3255. [Google Scholar] [CrossRef] [PubMed]
- Kronman, H.; Richter, F.; Labonté, B.; Chandra, R.; Zhao, S.; Hoffman, G.; Lobo, M.K.; Schadt, E.E.; Nestler, E.J. Biology and Bias in Cell Type-Specific RNAseq of Nucleus Accumbens Medium Spiny Neurons. Sci. Rep. 2019, 9, 8350. [Google Scholar] [CrossRef]
- Pavan, B.; Dalpiaz, A. Odorants could elicit repair processes in melanized neuronal and skin cells. Neural Regen. Res. 2007, 12, 1401–1404. [Google Scholar] [CrossRef] [PubMed]
- Grison, A.; Zucchelli, S.; Urzì, A.; Zamparo, I.; Lazarevic, D.; Pascarella, G.; Roncaglia, P.; Giorgetti, A.; Garcia-Esparcia, P.; Vlachouli, C.; et al. Mesencephalic dopaminergic neurons express a repertoire of olfactory receptors and respond to odorant-like molecules. BMC Genom. 2014, 15, 729. [Google Scholar] [CrossRef]
- Whittaker, J.R.; Foley, S.F.; Ackling, E.; Murphy, K.; Caseras, X. The Functional Connectivity Between the Nucleus Accumbens and the Ventromedial Prefrontal Cortex as an Endophenotype for Bipolar Disorder. Biol. Psychiatry 2018, 84, 803–809. [Google Scholar] [CrossRef]
- Gaudel, F.; Guiraudie-Capraz, G.; Feron, F. Limbic Expression of mRNA Coding for Chemoreceptors in Human Brain-Lessons from Brain Atlases. Int. J. Mol. Sci. 2021, 22, 6858. [Google Scholar] [CrossRef]
- Correll, C.U.; Koblan, K.S.; Hopkins, S.C.; Li, Y.; Dworak, H.; Goldman, R.; Loebel, A. Safety and Effectiveness of Ulotaront (SEP-363856) in Schizophrenia: Results of a 6-Month, Open-Label Extension Study. Npj Schizophr. 2021, 7, 63. [Google Scholar] [CrossRef]
- Galluppi, G.R.; Polhamus, D.G.; Fisher, J.M.; Hopkins, S.C.; Koblan, K.S. Population Pharmacokinetic Analysis of Ulotaront in Subjects with Schizophrenia. CPT Pharmacomet. Syst. Pharmacol. 2021, 10, 1245–1254. [Google Scholar] [CrossRef]
- Heffernan, M.L.R.; Herman, L.W.; Brown, S.; Jones, P.G.; Shao, L.; Hewitt, M.C.; Campbell, J.E.; Dedic, N.; Hopkins, S.C.; Koblan, K.S.; et al. Ulotaront: A TAAR1 Agonist for the Treatment of Schizophrenia. ACS Med. Chem. Lett. 2022, 13, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, S.C.; Ogirala, A.; Worden, M.; Koblan, K.S. Depicting Safety Profile of TAAR1 Agonist Ulotaront Relative to Reactions Anticipated for a Dopamine D2-Based Pharmacological Class in FAERS. Clin. Drug Investig. 2021, 41, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Steward, O.; Schuman, E.M. Compartmentalized Synthesis and Degradation of Proteins in Neurons. Neuron 2003, 40, 347–359. [Google Scholar] [CrossRef]
- Bahar Halpern, K.; Caspi, I.; Lemze, D.; Levy, M.; Landen, S.; Elinav, E.; Ulitsky, I.; Itzkovitz, S. Nuclear Retention of mRNA in Mammalian Tissues. Cell Rep. 2015, 13, 2653–2662. [Google Scholar] [CrossRef] [PubMed]
- De Abreu, R.S.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
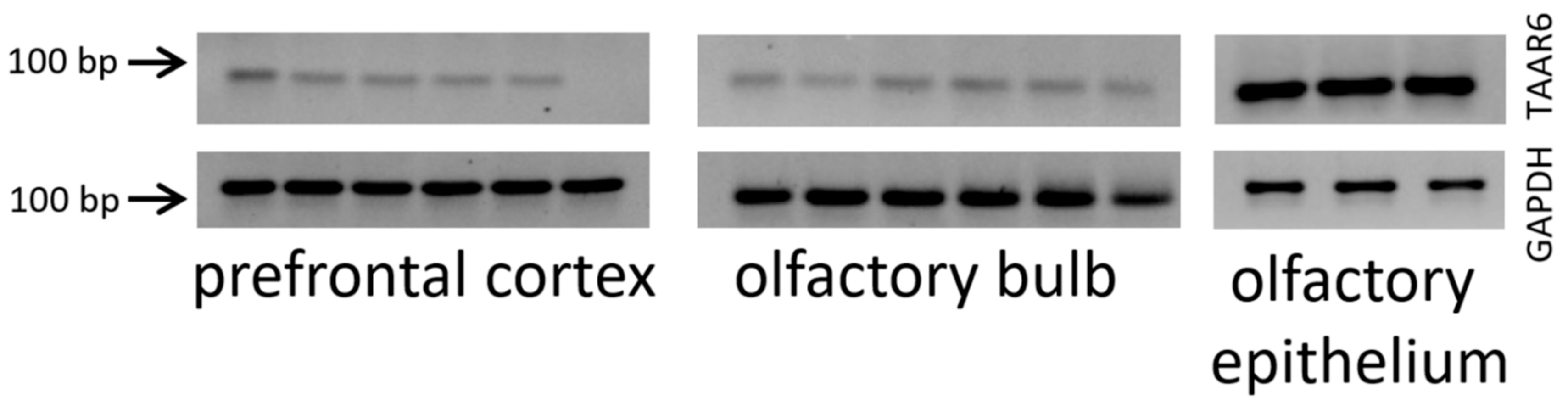{kind=link}
| Accession Number | Title | Samples | n 1 |
|---|---|---|---|
| GSE53239 | RNA-sequencing of the brain transcriptome implicates dysregulation of neuroplasticity, circadian rhythms, and GTPase binding in bipolar disorder | Dorsolateral prefrontal cortex | 11 |
| GSE53697 | RNAseq in Alzheimer’s disease patients | Dorsolateral prefrontal cortex | 8 |
| GSE123496 | Human brain tissues from healthy controls and multiple sclerosis patients | Frontal cortex | 5 |
| Parietal cortex | 5 | ||
| Hippocampus | 5 | ||
| Corpus callosum | 5 | ||
| Internal capsule | 5 | ||
| GSE124439 | Postmortem Cortex Samples Identify Distinct Molecular Subtypes of ALS: Retrotransposon Activation, Oxidative Stress, and Activated Glia | Frontal cortex | 9 |
| Motor cortex | 8 | ||
| GSE160521 | Diurnal rhythms across the human dorsal and ventral striatum | Nucleus accumbens | 59 |
| Nucleus caudatus | 59 | ||
| Putamen | 59 |
| Accession Number | Title | Samples | n 1 |
|---|---|---|---|
| GSE107183 | RNA changes in hippocampus of transgenic murine model of tauopathy (rTg4510 mice) compared to controls at asymptomatic stage (2 months) of neurodegeneration as determined by mRNA deep sequencing. | Hippocampus | 6 |
| GSE112575 | Frontal cortex transcriptomic analysis of a TDP-43 Q331K knock-in mouse (20 month) | Frontal cortex | 8 |
| GSE116752 | Striatal transcriptome of a mouse model of ADHD reveals a pattern of synaptic remodeling | Hippocampus | 5 |
| GSE126814 | Quantitative Analysis of Wild Type and Neat1 -/- Cerebral Frontal Cortex Transcriptomes | Frontal cortex | 5 |
| GSE130254 | Regional Analysis of The Brain Transcriptome in Mice Bred For High And Low Methamphetamine Consumption | Prefrontal cortex | 48 |
| Nucleus accumbens | 47 | ||
| Ventral midbrain | 48 | ||
| GSE136869 | Transcriptome analysis using RNA sequencing of the hippocampus of aged LPAR2-/- versus wildtype control mice | Hippocampus | 7 |
| GSE146628 | Identification of Natural Antisense Transcripts in Mouse Brain and Their Association with Autism Spectrum Disorder Risk Genes | Prefrontal cortex | 12 |
| Striatum | 12 | ||
| GSE147842 | Adult mouse hippocampal transcriptome changes associated with long-term behavioral and metabolic effects of gestational air pollution toxicity | Hippocampus | 10 |
| GSE148075 | Wild mice with different social network sizes vary in brain gene expression | Prefrontal cortex | 29 |
| Hippocampus | 29 | ||
| Hypothalamus | 28 | ||
| GSE166831 | Altered hippocampal transcriptome dynamics following sleep deprivation | Hippocampus | 9 |
| GSE170997 | Transcriptomics data of blood and brain from the YAC128 Huntington’s disease mouse model (brain) | Striatum | 8 |
| Cerebellum | 8 | ||
| GSE99353 | Frontal cortex transcriptomic analysis of a TDP-43 Q331K knock-in mouse (5 month) | Prefrontal cortex | 6 |
| Accession Number | Title | Samples | n 1 |
|---|---|---|---|
| GSE110727 | Characterization of the nuclear and cytosolic transcriptomes in human brain tissue | Frontal cortex (human) | 12 |
| GSE121199 | Biology and Bias in Cell Type-Specific RNAseq of Nucleus Accumbens Medium Spiny Neurons | Nucleus accumbens (mouse) | 9 |
| GSE130376 | Next-generation sequencing of cholinergic interneurons in the nucleus accumbens of cocaine-addicted and non-addicted mice | Nucleus accumbens (mouse) | 49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaganova, A.N.; Katolikova, N.V.; Murtazina, R.Z.; Kuvarzin, S.R.; Gainetdinov, R.R. Public Transcriptomic Data Meta-Analysis Demonstrates TAAR6 Expression in the Mental Disorder-Related Brain Areas in Human and Mouse Brain. Biomolecules 2022, 12, 1259. https://doi.org/10.3390/biom12091259
Vaganova AN, Katolikova NV, Murtazina RZ, Kuvarzin SR, Gainetdinov RR. Public Transcriptomic Data Meta-Analysis Demonstrates TAAR6 Expression in the Mental Disorder-Related Brain Areas in Human and Mouse Brain. Biomolecules. 2022; 12(9):1259. https://doi.org/10.3390/biom12091259
Chicago/Turabian StyleVaganova, Anastasia N., Nataliia V. Katolikova, Ramilya Z. Murtazina, Savelii R. Kuvarzin, and Raul R. Gainetdinov. 2022. "Public Transcriptomic Data Meta-Analysis Demonstrates TAAR6 Expression in the Mental Disorder-Related Brain Areas in Human and Mouse Brain" Biomolecules 12, no. 9: 1259. https://doi.org/10.3390/biom12091259