
Characterization of a Virally Encoded Flavodoxin That Can Drive Bacterial Cytochrome P450 Monooxygenase Activity

 , ,
, ,
Abstract
:1. Importance
2. Introduction
3. Materials and Methods
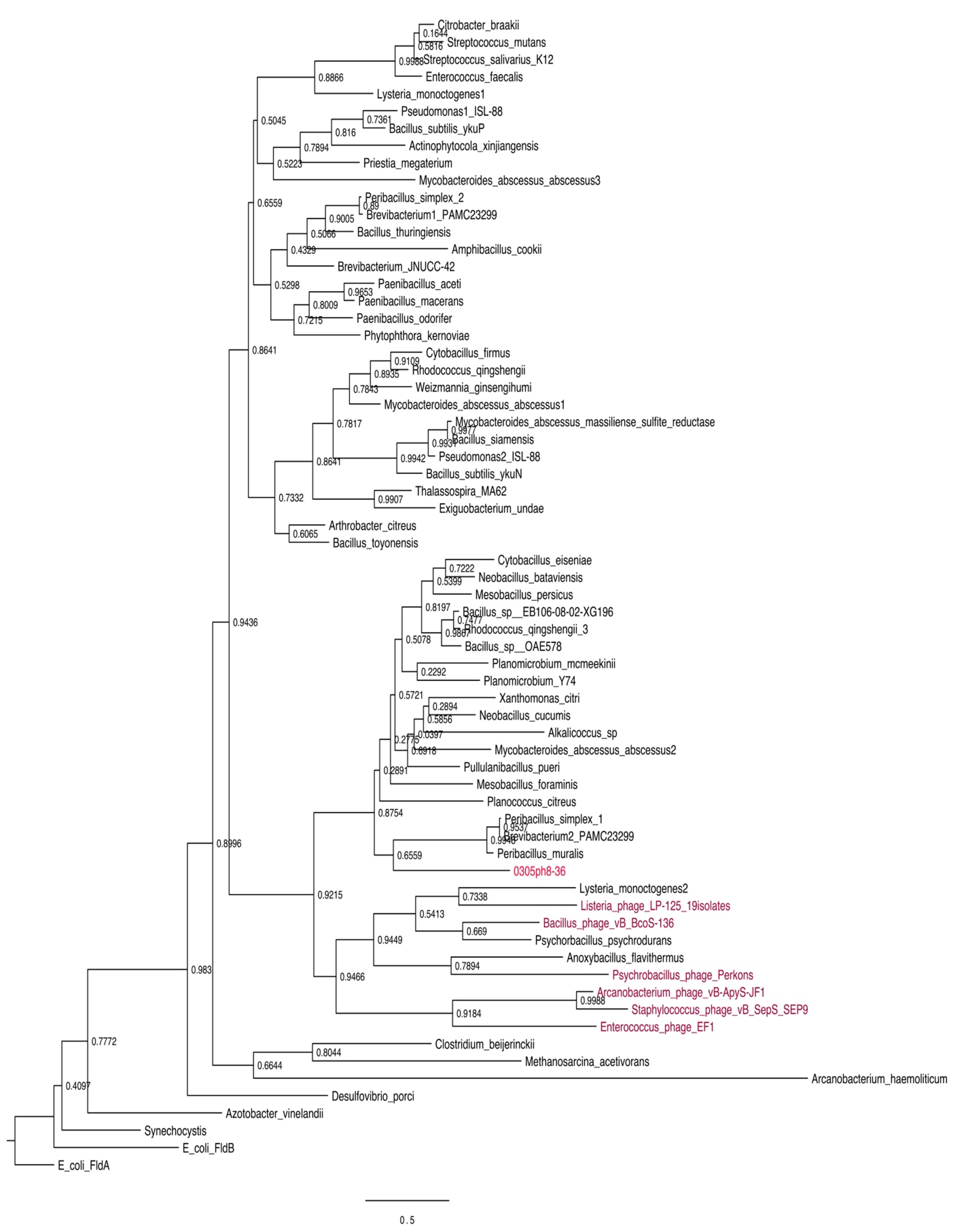
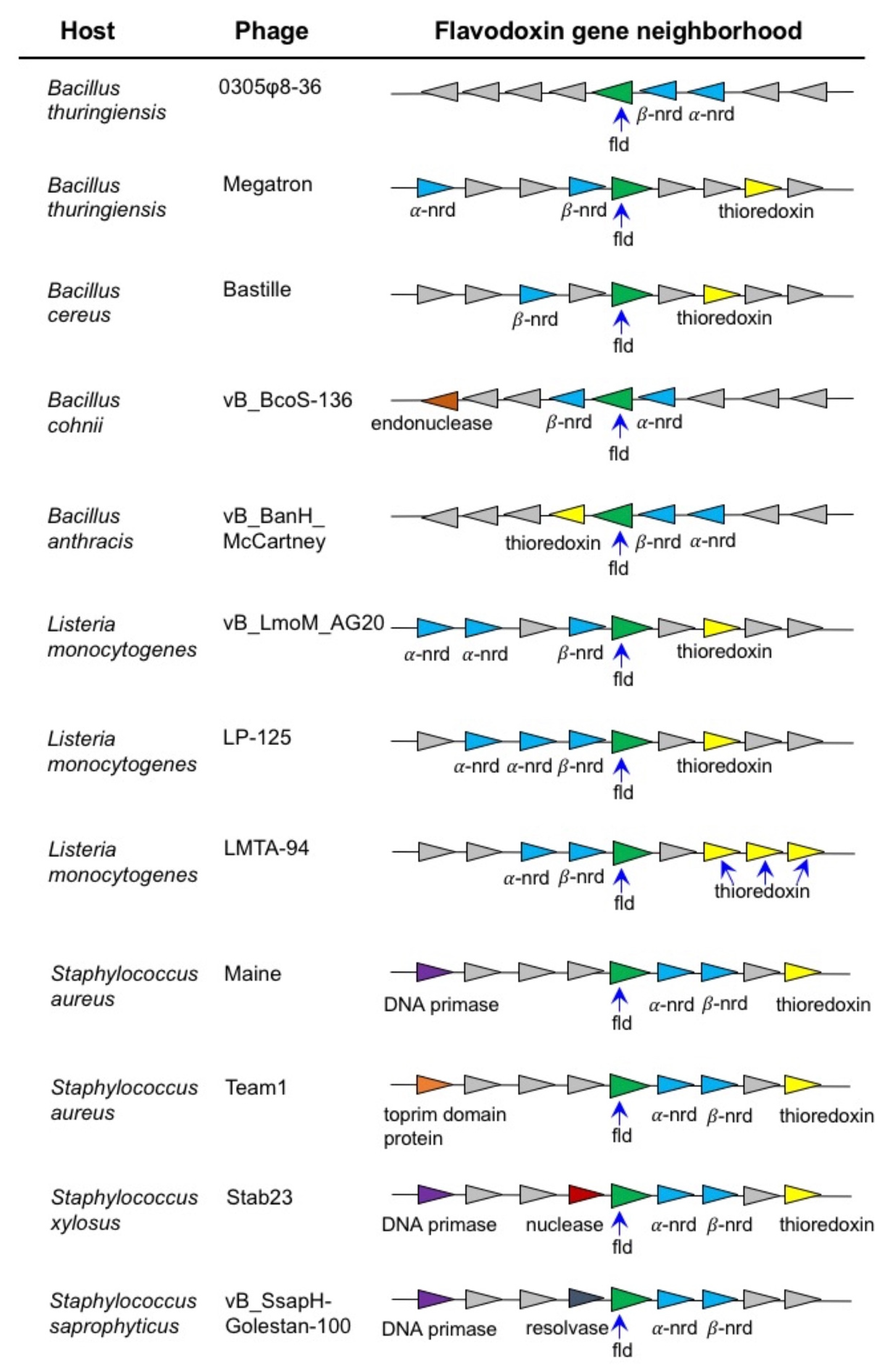
3.1. Bioinformatic and Phylogenetic Analysis
3.2. Cloning, Heterologous Expression and Purification of 0305φ8-36 Flavodoxin
3.3. Reconstitution of Streptomyces coelicolor CYP170A1 Activity with 0305φ8-36 Flavodoxin
3.4. General Methods
4. Results
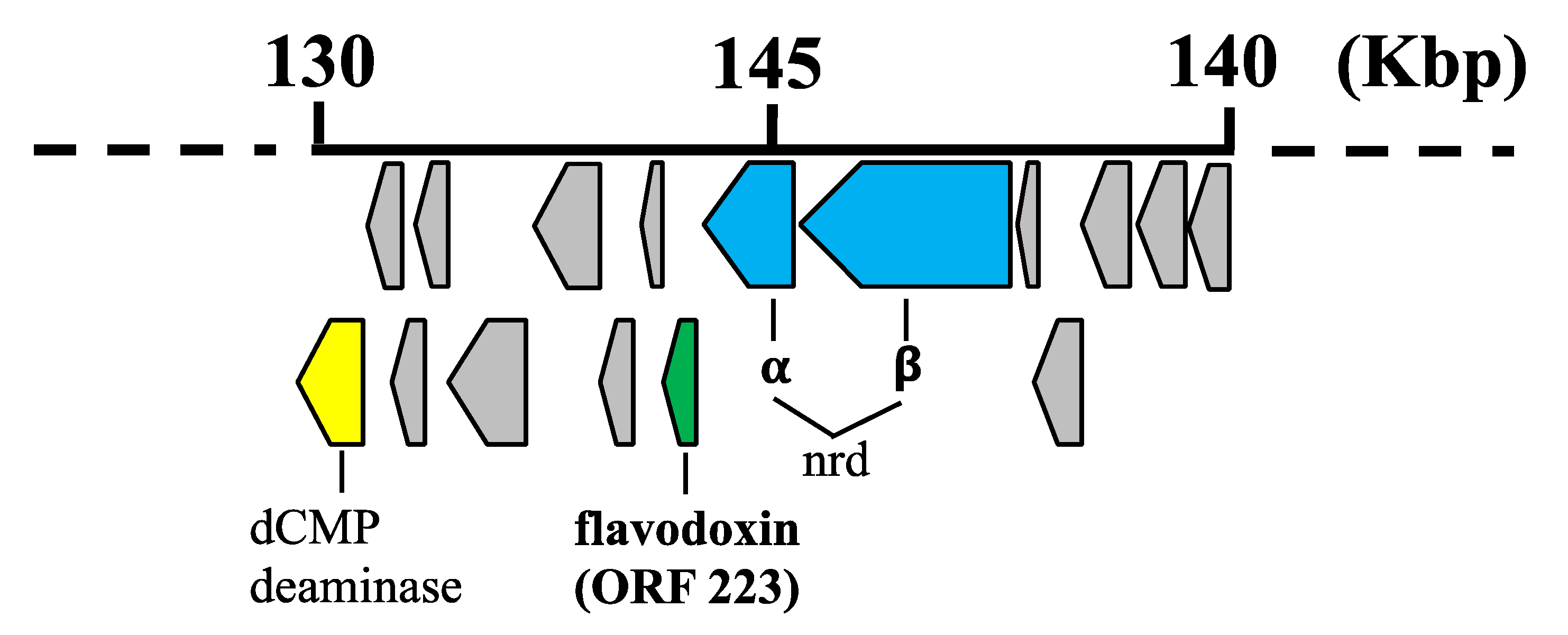
4.1. Bioinformatic and Phylogenetic Analysis of 0305φ8-36 ORF223
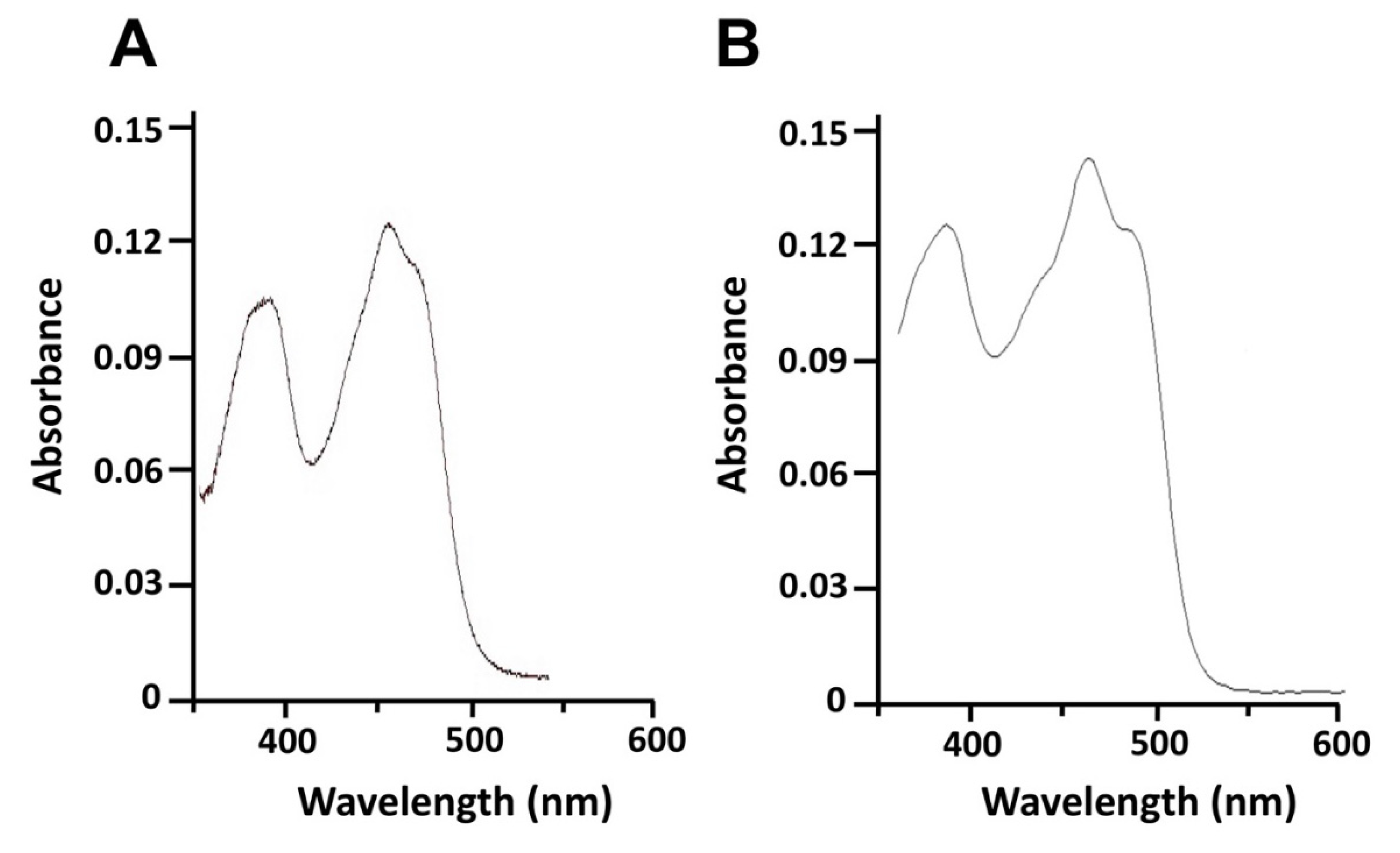
4.2. Expression, Purification and Spectral Analysis of Recombinant 0305φ8-36 ORF223
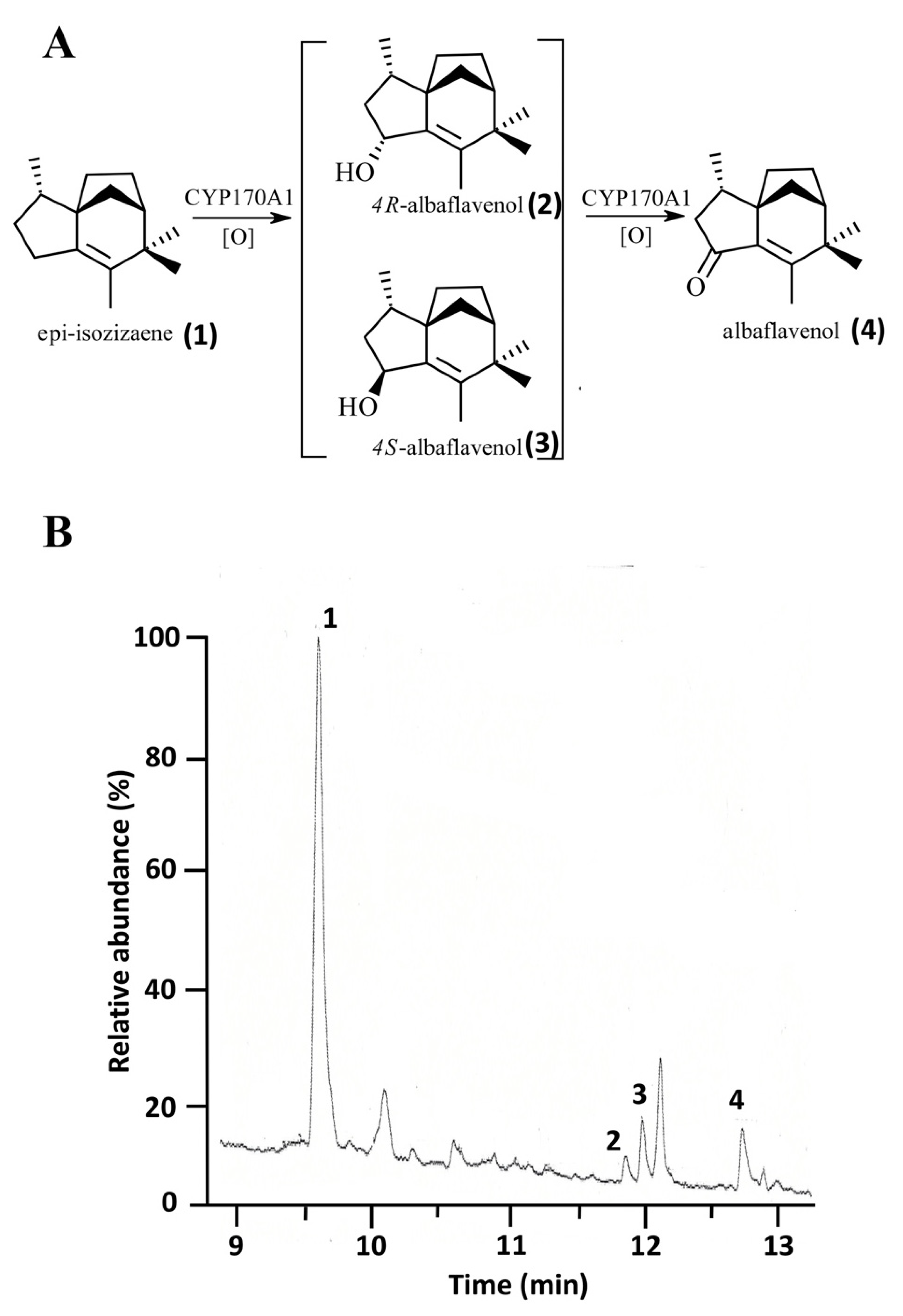
4.3. Characterization of the Catalytic Activity of 0305φ8-36 Flavodoxin in P450-Mediated Monooxygenation
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sancho, J. Flavodoxins: Sequence, folding, binding, function and beyond. Cell Mol. Life Sci. 2006, 63, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Smillie, R.M. Isolation of two proteins with chloroplast ferredoxin activity from a blue-green alga. Biochem. Biophys. Res. Commun. 1965, 20, 621–629. [Google Scholar] [CrossRef]
- Knight, E.; D’Eustachio, A.J.; Hardy, R.W. Flavodoxin: A flavoprotein with ferredoxin activity from Clostrium pasteurianum. Biochim. Biophys. Acta 1966, 113, 626–628. [Google Scholar] [CrossRef]
- Anderson, R.F. Energetics of the one-electron reduction steps of riboflavin, FMN and FAD to their fully reduced forms. Biochim. Biophys. Acta 1983, 722, 158–162. [Google Scholar] [CrossRef]
- Mayhew, S.G. The effects of pH and semiquinone formation on the oxidation-reduction potentials of flavin mononucleotide. A reappraisal. Eur. J. Biochem. 1999, 265, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Gaudu, P.; Weiss, B. Flavodoxin mutants of Escherichia coli K-12. J. Bacteriol. 2000, 182, 1788–1793. [Google Scholar] [CrossRef] [PubMed]
- Freigang, J.; Diederichs, K.; Schafer, K.P.; Welte, W.; Paul, R. Crystal structure of oxidized flavodoxin, an essential protein in Helicobacter pylori. Protein Sci. 2002, 11, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Modi, S.; Smith, G.; Paine, M.; McDonagh, P.D.; Wolf, C.R.; Tew, D.; Lian, L.Y.; Roberts, G.C.; Driessen, H.P. Crystal structure of the FMN-binding domain of human cytochrome P450 reductase at 1.93 A resolution. Protein Sci. 1999, 8, 298–306. [Google Scholar] [CrossRef]
- Lopez-Llano, J.; Maldonado, S.; Jain, S.; Lostao, A.; Godoy-Ruiz, R.; Sanchez-Ruiz, J.M.; Cortijo, M.; Fernandez-Recio, J.; Sancho, J. The long and short flavodoxins: II. The role of the differentiating loop in apoflavodoxin stability and folding mechanism. J. Biol. Chem. 2004, 279, 47184–47191. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, K.; Matsubara, H.; Rogers, L.J. Crystal structure of oxidized flavodoxin from a red alga Chondrus crispus refined at 1.8 A resolution. Description of the flavin mononucleotide binding site. J. Mol. Biol. 1992, 225, 775–789. [Google Scholar] [CrossRef]
- Peleato, M.L.; Ayora, S.; Inda, L.A.; Gomez-Moreno, C. Isolation and characterization of two different flavodoxins from the eukaryote Chlorella fusca. Biochem. J. 1994, 302, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, B.; Xue, B.; Lundin, D.; Edwards, R.A.; Breitbart, M. A bioinformatic analysis of ribonucleotide reductase genes in phage genomes and metagenomes. BMC Evol. Biol. 2013, 13, 33. [Google Scholar] [CrossRef] [PubMed]
- Harrison, A.O.; Moore, R.M.; Polson, S.W.; Wommack, K.E. Reannotation of the Ribonucleotide Reductase in a Cyanophage Reveals Life History Strategies Within the Virioplankton. Front. Microbiol. 2019, 10, 134. [Google Scholar] [CrossRef]
- Lamb, D.C.; Follmer, A.H.; Goldstone, J.V.; Nelson, D.R.; Warrilow, A.G.; Price, C.L.; True, M.Y.; Kelly, S.L.; Poulos, T.L.; Stegeman, J.J. On the occurrence of cytochrome P450 in viruses. Proc. Natl. Acad. Sci. USA 2019, 116, 12343–12352. [Google Scholar] [CrossRef] [PubMed]
- Hardies, S.C.; Thomas, J.A.; Serwer, P. Comparative genomics of Bacillus thuringiensis phage 0305phi8-36: Defining patterns of descent in a novel ancient phage lineage. Virol. J. 2007, 4, 97. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A.; Hardies, S.C.; Rolando, M.; Hayes, S.J.; Lieman, K.; Carroll, C.A.; Weintraub, S.T.; Serwer, P. Complete genomic sequence and mass spectrometric analysis of highly diverse, atypical Bacillus thuringiensis phage 0305phi8-36. Virology 2007, 368, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Chang, J.M.; Di Tommaso, P.; Notredame, C. TCS: A new multiple sequence alignment reliability measure to estimate alignment accuracy and improve phylogenetic tree reconstruction. Mol. Biol. Evol. 2014, 31, 1625–1637. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Series 1999, 41, 95–98. [Google Scholar]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, 320–324. [Google Scholar] [CrossRef]
- Kozlov, A.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Felipe, M.D.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Lei, L.; Warrilow, A.G.; Lepesheva, G.I.; Mullins, J.G.; Waterman, M.R.; Kelly, S.L. The first virally encoded cytochrome P450. J. Virol. 2009, 83, 8266–8269. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Lei, L.; Zhao, B.; Yuan, H.; Jackson, C.J.; Warrilow, A.G.; Skaug, T.; Dyson, P.J.; Dawson, E.S.; Kelly, S.L.; et al. Streptomyces coelicolor A3(2) CYP102 protein, a novel fatty acid hydroxylase encoded as a heme domain without an N-terminal redox partner. Appl. Environ. Microbiol. 2010, 76, 1975–1980. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.M.; Waterman, M.R. Flavodoxin and NADPH-flavodoxin reductase from Escherichia coli support bovine cytochrome P450c17 hydroxylase activities. J. Biol. Chem. 1994, 269, 27401–27408. [Google Scholar] [CrossRef]
- Zhao, B.; Lin, X.; Lei, L.; Lamb, D.C.; Kelly, S.L.; Waterman, M.R.; Cane, D.E. Biosynthesis of the sesquiterpene antibiotic albaflavenone in Streptomyces coelicolor A3(2). J. Biol. Chem. 2008, 283, 8183–8189. [Google Scholar] [CrossRef]
- Omura, T.; Sato, R. The Carbon Monoxide-Binding Pigment of Liver Microsomes. I. Evidence for Its Hemoprotein Nature. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar] [CrossRef]
- Jenkins, C.M.; Waterman, M.R. NADPH-flavodoxin reductase and flavodoxin from Escherichia coli: Characteristics as a soluble microsomal P450 reductase. Biochemistry 1998, 37, 6106–6113. [Google Scholar] [CrossRef]
- Zhao, B.; Guengerich, F.P.; Bellamine, A.; Lamb, D.C.; Izumikawa, M.; Lei, L.; Podust, L.M.; Sundaramoorthy, M.; Kalaitzis, J.A.; Reddy, L.M.; et al. Binding of two flaviolin substrate molecules, oxidative coupling, and crystal structure of Streptomyces coelicolor A3(2) cytochrome P450 158A2. J. Biol. Chem. 2005, 280, 11599–11607. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberon, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. 2011, 41, 423–431. [Google Scholar] [CrossRef]
- Sauder, A.B.; Quinn, M.R.; Brouillette, A.; Caruso, S.; Cresawn, S.; Erill, I.; Lewis, L.; Loesser-Casey, K.; Pate, M.; Scott, C.; et al. Genomic characterization and comparison of seven Myoviridae bacteriophage infecting Bacillus thuringiensis. Virology 2016, 489, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Erill, I.; Caruso, S.M.; 2015 UMBC Phage Hunters. Genome sequence of Bacillus cereus group phage SalinJah. Genome Announc. 2016, 4, e00953-16. [Google Scholar] [CrossRef] [PubMed]
- Erill, I.; Caruso, S.M. Genome sequences of two Bacillus cereus group Bacteriophages, Eyuki and AvesoBmore. Genome Announc. 2015, 3, e01199-15. [Google Scholar] [CrossRef] [PubMed]
- Ting, J.H.; Smyth, T.B.; Chamakura, K.R.; Kuty Everett, G.F. Complete genome of Bacillus thuringiensis myophage BigBertha. Genome Announc. 2013, 1, e00853-13. [Google Scholar] [CrossRef]
- Maroun, J.W.; Whitcher, K.J.; Chamakura, K.R.; Kuty Everett, G.F. Complete genome of Bacillus thuringiensis myophage Spock. Genome Announc. 2013, 1, e00863-13. [Google Scholar] [CrossRef]
- Breslin, E.F.; Cornell, J.; Schuhmacher, Z.; Himelright, M.; Andos, A.; Childs, A.; Clem, A.; Gerber, M.; Gordillo, A.; Harb, L.; et al. Complete genome sequence of Bacillus phage Belinda from Grand Cayman Island. Genome Announc. 2016, 4, e00571-16. [Google Scholar] [CrossRef]
- Denes, T.; Vongkamjan, K.; Ackermann, H.W.; Moreno Switt, A.I.; Wiedmann, M.; den Bakker, H.C. Comparative genomic and morphological analyses of Listeria phages isolated from farm environments. Appl. Environ. Microbiol. 2014, 80, 4616–4625. [Google Scholar] [CrossRef]
- Peters, T.L.; Song, Y.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Mutant and recombinant phages selected from in vitro coevolution conditions overcome phage-resistant Listeria monocytogenes. Appl. Environ. Microbiol. 2020, 86, e02138-20. [Google Scholar] [CrossRef]
- Duperier, J.; Bulpitt, M.; Bispo, F.; Greguske, E. Genome annotations of two Bacillus phages, Tomato and Baseball Field. Microbiol. Resour. Announc. 2021, 10, e01196-20. [Google Scholar] [CrossRef]
- Peters, T.L.; Hudson, L.K.; Song, Y.; Denes, T.G. Complete genome sequences of two Listeria phages of the genus Pecentumvirus. Microbiol. Resour. Announc. 2019, 8, e01229-19. [Google Scholar] [CrossRef]
- Kostyk, N.; Chigbu, O.; Cochran, E.; Davis, J.; Essig, J.; Do, L.; Farooque, N.; Gbadamosi, Z.; Gnanodayan, A.; Hale, A.; et al. Complete genome sequences of Bacillus cereus group phages AaronPhadgers, ALPS, Beyonphe, Bubs, KamFam, OmnioDeoPrimus, Phireball, PPIsBest, YungSlug, and Zainny. Microbiol. Resour. Announc. 2021, 10, e0030021. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, B.; Schilling, T.; Hoppert, M.; Hertel, R. Complete genome sequence of the virus isolate vB_BthM-Goe5 infecting Bacillus thuringiensis. Arch Virol. 2019, 164, 1485–1488. [Google Scholar] [CrossRef] [PubMed]
- Foltz, S.; Johnson, A.A. Complete genome sequences of nine Bacillus cereus group phages. Genome Announc. 2016, 4, e00473-16. [Google Scholar] [CrossRef] [PubMed]
- Flounlacker, K.; Miller, R.; Marquez, D.; Johnson, A.; the 2015–2016 VCU Phage Hunters. complete genome sequences of Bacillus phages DirtyBetty and Kida. Genome Announc. 2017, 5, e01385-16. [Google Scholar] [CrossRef]
- Erill, I.; Caruso, S.M. Complete genome sequence of Bacillus cereus group phage TsarBomba. Genome Announc. 2015, 3, e01178-15. [Google Scholar] [CrossRef]
- Lee, M.; Puglisi, K.M.; UMBC STEM-BUILD Cohort 1; Erill, I.; Caruso, S.M. Complete genome sequences of HonestAbe, Anthony, and Taffo16, three cluster C Bacillus cereus group bacteriophages. Genome Announc. 2018, 6, e00493-18. [Google Scholar] [CrossRef]
- Greguske, E.; Nadeau, A.; Fitzmeyer, E.; Fucikova, K. Genome sequence of Bacillus phage Saddex. Microbiol. Resour. Announc. 2018, 7, e01044-18. [Google Scholar] [CrossRef]
- Kent, B.; Raymond, T.; Mosier, P.D.; Johnson, A.A.; the 2016–2017 VCU Phage Hunters. Complete genome sequences of Bacillus phages Janet and OTooleKemple52. Genome Announc. 2018, 6, e00083-18. [Google Scholar] [CrossRef]
- Moreland, R.; Korn, A.; Newkirk, H.; Liu, M.; Gill, J.J.; Cahill, J.; Ramsey, J. Complete genome sequence of Staphylococcus aureus myophage Maine. Microbiol. Resour. Announc. 2019, 8, e01050-19. [Google Scholar] [CrossRef]
- Korn, A.M.; Hillhouse, A.E.; Sun, L.; Gill, J.J. Comparative genomics of three novel jumbo bacteriophages infecting Staphylococcus aureus. J. Virol. 2021, 95, e0239120. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Vandenheuvel, D.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Two phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono- and dual-species Staphylococcal biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [PubMed]
- El Haddad, L.; Ben Abdallah, N.; Plante, P.L.; Dumaresq, J.; Katsarava, R.; Labrie, S.; Corbeil, J.; St-Gelais, D.; Moineau, S. Improving the safety of Staphylococcus aureus polyvalent phages by their production on a Staphylococcus xylosus strain. PLoS ONE 2014, 9, e102600. [Google Scholar]
- Alves, D.R.; Gaudion, A.; Bean, J.E.; Perez Esteban, P.; Arnot, T.C.; Harper, D.R.; Kot, W.; Hansen, L.H.; Enright, M.C.; Jenkins, A.T. Combined use of bacteriophage K and a novel bacteriophage to reduce Staphylococcus aureus biofilm formation. Appl. Environ. Microbiol. 2014, 80, 6694–6703. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Liu, X.; Lu, R.; Li, Y.; Song, J.; Lei, L.; Sun, C.; Feng, X.; Du, C.; Yu, H.; et al. Complete genome sequence of Staphylococcus aureus bacteriophage GH15. J. Virol. 2012, 86, 8914–8915. [Google Scholar] [CrossRef]
- Crane, A.; Abaidoo, J.; Beltran, G.; Fry, D.; Furey, C.; Green, N.; Johal, R.; La Rosa, B.; Jimenez, C.L.; Luong, L.; et al. The complete genome sequence of the Staphylococcus bacteriophage Metroid. G3 2020, 10, 2975–2979. [Google Scholar] [CrossRef]
- Jun, J.W.; Giri, S.S.; Kim, H.J.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Park, S.C. Complete genome sequence of the novel bacteriophage pSco-10 Infecting Staphylococcus cohnii. Genome Announc. 2017, 5, e01032-17. [Google Scholar] [CrossRef]
- Sáez Moreno, D.; Visram, Z.; Mutti, M.; Restrepo-Córdoba, M.; Hartmann, S.; Kremers, A.I.; Tišáková, L.; Schertler, S.; Wittmann, J.; Kalali, B.; et al. ε2-phages are naturally bred and have a vastly improved host range in Staphylococcus aureus over wild type phages. Pharmaceuticals 2021, 14, 325. [Google Scholar] [CrossRef]
- Oduor, J.M.O.; Kadija, E.; Nyachieo, A.; Mureithi, M.W.; Skurnik, M. Bioprospecting Staphylococcus phages with therapeutic and bio-control potential. Viruses 2020, 12, 133. [Google Scholar] [CrossRef]
- Abatángelo, V.; Peressutti Bacci, N.; Boncompain, C.A.; Amadio, A.F.; Carrasco, S.; Suárez, C.A.; Morbidoni, H.R. Broad-range lytic bacteriophages that kill Staphylococcus aureus local field strains. PLoS ONE 2017, 12, e0181671. [Google Scholar]
- Hoover, D.M.; Jarrett, J.T.; Sands, R.H.; Dunham, W.R.; Ludwig, M.L.; Matthews, R.G. Interaction of Escherichia coli cobalamin-dependent methionine synthase and its physiological partner flavodoxin: Binding of flavodoxin leads to axial ligand dissociation from the cobalamin cofactor. Biochemistry 1997, 36, 127–138. [Google Scholar] [CrossRef]
- Green, A.J.; Rivers, S.L.; Cheeseman, M.; Reid, G.A.; Quaroni, L.G.; Macdonald, I.D.; Chapman, S.K.; Munro, A.W. Expression, purification and characterization of cytochrome P450 Biol: A novel P450 involved in biotin synthesis in Bacillus subtilis. J. Biol. Inorg. Chem. 2001, 6, 523–533. [Google Scholar] [CrossRef]
- Gangeswaran, R.; Eady, R.R. Flavodoxin 1 of Azotobacter vinelandii: Characterization and role in electron donation to purified assimilatory nitrate reductase. Biochem. J. 1996, 317, 103–108. [Google Scholar] [CrossRef]
- Prakash, D.; Iyer, P.R.; Suharti, S.; Walters, K.A.; Santiago-Martinez, M.G.; Golbeck, J.H.; Murakami, K.S.; Ferry, J.G. Structure and function of an unusual flavodoxin from the domain Archaea. Proc. Natl. Acad. Sci. USA 2019, 116, 25917–25922. [Google Scholar] [CrossRef]
- Jackson, R.G.; Rylott, E.L.; Fournier, D.; Hawari, J.; Bruce, N.C. Exploring the biochemical properties and remediation applications of the unusual explosive-degrading P450 system XplA/B. Proc. Natl. Acad. Sci. USA 2007, 104, 16822–16827. [Google Scholar] [CrossRef]
- Liu, L.; Schmid, R.D.; Urlacher, V.B. Cloning, expression, and characterization of a self-sufficient cytochrome P450 monooxygenase from Rhodococcus ruber DSM 44319. Appl. Microbiol. Biotechnol. 2006, 72, 876–882. [Google Scholar] [CrossRef]
- Hawkes, D.B.; Adams, G.W.; Burlingame, A.L.; Ortiz de Montellano, P.R.; De Voss, J.J. Cytochrome P450(cin) (CYP176A), isolation, expression, and characterization. J. Biol. Chem. 2002, 277, 27725–27732. [Google Scholar] [CrossRef]
- Jordan, A.; Reichard, P. Ribonucleotide reductases. Annu. Rev. Biochem. 1998, 67, 71–98. [Google Scholar] [CrossRef]
- Sheppard, A.E.; Poehlein, A.; Rosenstiel, P.; Liesegang, H.; Schulenburg, H. Complete Genome Sequence of Bacillus thuringiensis Strain 407 Cry. Genome Announc. 2013, 1, e00158-12. [Google Scholar] [CrossRef]
- Mthethwa, B.C.; Chen, W.; Ngwenya, M.L.; Kappo, A.P.; Syed, P.R.; Karpoormath, R.; Yu, J.H.; Nelson, D.R.; Syed, K. Comparative Analyses of Cytochrome P450s and Those Associated with Secondary Metabolism in Bacillus Species. Int. J. Mol. Sci. 2018, 19, 3623. [Google Scholar] [CrossRef]
- Szaleniec, M.; Wojtkiewicz, A.M.; Bernhardt, R.; Borowski, T.; Donova, M. Bacterial steroid hydroxylases: Enzyme classes, their functions and comparison of their catalytic mechanisms. Appl. Microbiol. Biotechnol. 2018, 102, 8153–8171. [Google Scholar] [CrossRef]
- Grose, J.H.; Jensen, G.L.; Burnett, S.H.; Breakwell, D.P. Genomic comparison of 93 Bacillus phages reveals 12 clusters, 14 singletons and remarkable diversity. BMC Genom. 2014, 15, 855. [Google Scholar]
- Furuya, T.; Shibata, D.; Kino, K. Phylogenetic analysis of Bacillus P450 monooxygenases and evaluation of their activity towards steroids. Steroids 2009, 74, 906–912. [Google Scholar] [CrossRef]
- Padayachee, T.; Nzuza, N.; Chen, W.; Nelson, D.R.; Syed, K. Impact of lifestyle on cytochrome P450 monooxygenase repertoire is clearly evident in the bacterial phylum Firmicutes. Sci. Rep. 2020, 10, 13982. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Z.; Wang, C.; Zhou, Y.J.; Xu, H.; Li, S. Biochemical characterization of three new α-olefin-producing P450 fatty acid decarboxylases with a halophilic property. Biotechno. Biofuels 2019, 12, 79. [Google Scholar] [CrossRef]
- Vardi, A.; Haramaty, L.; Van Mooy, B.A.; Fredricks, H.F.; Kimmance, S.A.; Larsen ABidle, K.D. Host-virus dynamics and subcellular controls of cell fate in a natural coccolithophore population. Proc. Natl. Acad. Sci. USA 2012, 109, 19327–19332. [Google Scholar] [CrossRef]
- Monier, A.; Chambouvet, A.; Milner, D.S.; Attah, V.; Terrado, R.; Lovejoy, C.; Moreau, H.; Santoro, A.E.; Derelle, E.; Richards, T.A. Host-derived viral transporter protein for nitrogen uptake in infected marine phytoplankton. Proc. Natl. Acad. Sci. USA 2017, 114, 489–498. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Martinez-Gutierrez, C.A.; Weinheimer, A.R.; Aylward, F.O. Dynamic genome evolution and complex virocell metabolism of globally-distributed giant viruses. Nat. Commun. 2020, 11, 1710. [Google Scholar] [CrossRef]
- Blanc-Mathieu, R.; Dahle, H.; Hofgaard, A.; Brandt, D.; Ban, H.; Kalinowski, J.; Ogata, H.; Sandaa, R.A. A persistent giant algal virus, with a unique morphology, encodes an unprecedented number of genes involved in energy metabolism. J. Virol. 2021, 95, e02446-20. [Google Scholar] [CrossRef]
- Brahim Belhaouari, D.; Pires De Souza, G.A.; Lamb, D.C.; Kelly, S.L.; Goldstone, J.V.; Stegeman, J.J.; Colson, P.; La Scola, B.; Aherfi, S. Metabolic arsenal of giant viruses: Host hijack or self-use? eLife 2022, 11, e78674. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Phage | Phage Host | Sample Location | Predicted Phage ORFs | Flavodoxin aa | Genbank Identifier | Ref. |
|---|---|---|---|---|---|---|---|
| Bacillus | 0305φ8-36 | Bacillus thuringiensis | soil | 246 | 148 | ABS83781.1 | [16] |
| Hakuna | Bacillus thuringiensis | soil | 294 | 159 | YP_009036585.1 | [31] | |
| Megatron | Bacillus thuringiensis | soil | 290 | 159 | YP_009036208.1 | [31] | |
| Riley | Bacillus thuringiensis | soil | 290 | 149 | YP_009055891.1 | [31] | |
| CAM003 | Bacillus thuringiensis | soil | 287 | 150 | YP_009037037.1 | [31] | |
| Evoli | Bacillus thuringiensis | soil | 294 | 150 | YP_009035659.1 | [31] | |
| Hoody T | Bacillus thuringiensis | soil | 270 | 153 | YP_009035330.1 | [31] | |
| Troll | Bacillus thuringiensis | soil | 289 | 149 | YP_008430917.1 | [31] | |
| SalinJah | Bacillus cereus | soil | 292 | 149 | ANH50605.1 | [32] | |
| Eyuki | Bacillus cereus | soil | 300 | 159 | YP_009212082.1 | [33] | |
| AvesoBmore | Bacillus thuringiensis | soil | 301 | 149 | YP_009206486.1 | [33] | |
| BigBertha | Bacillus thuringiensis | soil | 291 | 153 | YP_008771155.1 | [34] | |
| Spock | Bacillus thuringiensis | soil | 283 | 149 | YP_008770352.1 | [35] | |
| Belinda | Bacillus thuringiensis | soil | 295 | 159 | ANM46069.1 | [36] | |
| Tomato | Bacillus thuringiensis | soil | 200 | 159 | QLF85935.1 | [37] | |
| AaronPhadgers | Bacillus thuringiensis | soil | 301 | 159 | ASR78796.1 | [38] | |
| Beyonphe | Bacillus cereus | water | 300 | 149 | QDH49824.1 | [38] | |
| Bubs | Bacillus thuringiensis | soil | 302 | 159 | ASR78601.1 | [38] | |
| Kamfam | Bacillus thuringiensis | soil | 293 | 150 | AXQ67322.1 | [38] | |
| PIPsBest | Bacillus thuringiensis | soil | 301 | 159 | ASR78379.1 | [38] | |
| ALPS | Bacillus thuringiensis | soil | 295 | 159 | QDH50122.1 | [38] | |
| Phireball | Bacillus thuringiensis | soil | 299 | 159 | QDH49415.1 | [38] | |
| OmnioDeoPrimus | Bacillus thuringiensis | soil | 299 | 159 | AXQ67477.1 | [38] | |
| Zainny | Bacillus thuringiensis | soil | 303 | 159 | ASR79375.1 | [38] | |
| vB_BthM-Goe5 | Bacillus thuringiensis | soil | 272 | 150 | AZF89241.1 | [39] | |
| SageFayge | Bacillus thuringiensis | soil | 300 | 159 | YP_009280944.1 | [40] | |
| Nemo | Bacillus thuringiensis | soil | 301 | 159 | YP_009287019.1 | [40] | |
| Nigalana | Bacillus thuringiensis | soil | 302 | 159 | YP_009282535.1 | [40] | |
| DIGNKC | Bacillus thuringiensis | soil | 291 | 159 | AMW62869.1 | [40] | |
| Zuko | Bacillus thuringiensis | soil | 294 | 159 | AMW62552.1 | [40] | |
| Phrodo | Bacillus thuringiensis | soil | 288 | 149 | YP_009290006.1 | [40] | |
| NotTheCreek | Bacillus thuringiensis | soil | 296 | 159 | YP_009284467.1 | [40] | |
| Juglone | Bacillus thuringiensis | soil | 293 | 149 | AMW61744.1 | [40] | |
| Vinny | Bacillus thuringiensis | soil | 297 | 150 | AMW61891.1 | [40] | |
| DirtyBetty | Bacillus thuringiensis | soil | 302 | 159 | YP_009285086.1 | [41] | |
| Kida | Bacillus thuringiensis | soil | 304 | 159 | YP_009279309.1 | [41] | |
| TsarBomba | Bacillus thuringiensis | soil | 247 | 151 | YP_009206942.1 | [42] | |
| HonestAbe | Bacillus thuringiensis | soil | 286 | 159 | AUV57777.1 | [43] | |
| Taffo16 | Bacillus thuringiensis | soil | 284 | 149 | ASZ75860.1 | [43] | |
| Anthony | Bacillus thuringiensis | soil | 279 | 150 | ASU00988.1 | [43] | |
| Saddex | Bacillus thuringiensis | soil | 208 | 159 | AXF41931.1 | [44] | |
| Janet | Bacillus thuringiensis | soil | 285 | 150 | ASR79938.1 | [45] | |
| OTooleKemple52 | Bacillus thuringiensis | soil | 291 | 153 | ASR79614.1 | [45] | |
| SBP8a | Bacillus cereus | soil | 298 | 146 | AOZ62389.1 | DS * | |
| QCM8 | Bacillus cereus | soil | 288 | 151 | AOZ62044.1 | DS * | |
| BJ4 | Bacillus cereus | soil | 298 | 146 | AOZ61763.1 | DS * | |
| Bastille | Bacillus cereus | soil | 273 | 150 | YP_006907364.1 | DS * | |
| Chotacabras | Bacillus thuringiensis | soil | 285 | 149 | QEM43180.1 | DS * | |
| vB_BcoS-136 | Bacillus cohnii | lake sediment | 238 | 146 | AYP68319.1 | DS * | |
| Flapjack | Bacillus thuringiensis | soil | 288 | 149 | ARQ95040.1 | DS * | |
| vB_BanH_ RonSwanson | Bacillus anthracis | soil | - | 202 | UGO50429.1 | DS * | |
| vB_BanH_ Emiliahah | Bacillus anthracis | soil | - | 174 | UGO49205.1 | DS * | |
| vB_BanH_JarJar | Bacillus anthracis | soil | - | 202 | UGO48939.1 | DS * | |
| vB_BanH_ McCartney | Bacillus anthracis | soil | - | 174 | UGO47691.1 | DS * | |
| vB_BanH_ Abinadi | Bacillus anthracis | soil | - | 159 | UGO46389.1 | DS * | |
| Anthos | Bacillus thuringiensis | soil | 290 | 149 | QPY77365.1 | DS * | |
| Smudge | Bacillus thuringiensis | soil | 292 | 159 | ANI24755.1 | DS * | |
| BC-T25 | Bacillus sp. | soil | 236 | 151 | QEG04194.1 | DS * | |
| Listeria | vB_LmoM_AG20 | Listeria monocytogenes | abbatoir | 178 | 141 | YP_007676786.1 | DS * |
| LP-124 | Listeria monocytogenes | silage | 188 | 150 | YP_009784573.1 | [46] | |
| LP-064 | Listeria monocytogenes | silage | 188 | 150 | YP_009592673.1 | [46] | |
| LP-083-2 | Listeria monocytogenes | silage | 189 | 150 | YP_009044596.1 | [46] | |
| LP-048 | Listeria monocytogenes | silage | 177 | 150 | YP_009042933.1 | [46] | |
| LP-125 | Listeria monocytogenes | silage | 189 | 150 | YP_008240106.1 | [46] | |
| LP-Mix_6.2 | Listeria monocytogenes | laboratory | 197 | 150 | QNL32088.1 | [47] | |
| LP-Mix_6.1 | Listeria monocytogenes | laboratory | 195 | 150 | QNL31890.1 | [47] | |
| LP-039 | Listeria monocytogenes | silage | 201 | 150 | QEP53123.2 | [48] | |
| LP-066 | Listeria monocytogenes | silage | 189 | 150 | QDK04972.2 | [48] | |
| LMTA-94 | Listeria monocytogenes | laboratory | 189 | 150 | AID17150.1 | DS | |
| Staphylococcus | Maine | Staphylococcus aureus | pig barn swab | 219 | 130 | QEM41386.1 | [49] |
| MarsHill | Staphylococcus aureus | pig barn swab | 262 | 134 | QQM14610.1 | [50] | |
| Madawaska | Staphylococcus aureus | pig barn swab | 264 | 134 | QQO92731.1 | [50] | |
| Machias | Staphylococcus aureus | pig barn swab | 263 | 140 | QQO92468.1 | [50] | |
| phiIPLA-RODI | Staphylococcus aureus | sewage | 213 | 139 | AJA42083.1 | [51] | |
| phiIPLA-C1C | Staphylococcus aureus | sewage | 203 | 132 | YP_009214514.1 | [51] | |
| Team1 | Staphylococcus aureus | hospital | 217 | 143 | YP_009098273.1 | [52] | |
| MCE-2014 | Staphylococcus aureus | sewage | 204 | 130 | P_009098058.1 | [53] | |
| GH15 | Staphylococcus aureus | sewage | 214 | 130 | YP_007002257.1 | [54] | |
| Metroid | Staphylococcus sp. | soil | 254 | 143 | KE56191.1 | [55] | |
| pSco-10 | Staphylococcus cohnii | duck feces | 131 | 130 | ANH50479.1 | [56] | |
| BT3 | Staphylococcus aureus | animal | 232 | 139 | QVD58098.1 | [57] | |
| Stab23 | Staphylococcus xylosus | sewage | 247 | 130 | VEV88569.1 | [58] | |
| vB_Sau_S24 | Staphylococcus aureus | soil | 209 | 130 | ARM69410.1 | [59] | |
| vB_Sau_Clo6 | Staphylococcus aureus | sewage | 213 | 130 | ARM69197.1 | [59] | |
| vB_SsapH-Golestan-100 | Staphylococcus saprophyticus | - | 192 | 133 | BDA81541.1 | DS * | |
| phiSA_BS1 | Staphylococcus sp. | dairy farm | 200 | 129 | YP_009799552.1 | DS * | |
| vB_SsapH-Golestan-105-M | Staphylococcus saprophyticus | - | 203 | 149 | BDA82285.1 | DS * | |
| SA3 | Staphylococcus aureus | sewage | 223 | 130 | ASZ78055.1 | DS * | |
| PALS_1 | Staphylococcus aureus | animal | 191 | 130 | QDJ97591.1 | DS * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamb, D.C.; Goldstone, J.V.; Zhao, B.; Lei, L.; Mullins, J.G.L.; Allen, M.J.; Kelly, S.L.; Stegeman, J.J. Characterization of a Virally Encoded Flavodoxin That Can Drive Bacterial Cytochrome P450 Monooxygenase Activity. Biomolecules 2022, 12, 1107. https://doi.org/10.3390/biom12081107
Lamb DC, Goldstone JV, Zhao B, Lei L, Mullins JGL, Allen MJ, Kelly SL, Stegeman JJ. Characterization of a Virally Encoded Flavodoxin That Can Drive Bacterial Cytochrome P450 Monooxygenase Activity. Biomolecules. 2022; 12(8):1107. https://doi.org/10.3390/biom12081107
Chicago/Turabian StyleLamb, David C., Jared V. Goldstone, Bin Zhao, Li Lei, Jonathan G. L. Mullins, Michael J. Allen, Steven L. Kelly, and John J. Stegeman. 2022. "Characterization of a Virally Encoded Flavodoxin That Can Drive Bacterial Cytochrome P450 Monooxygenase Activity" Biomolecules 12, no. 8: 1107. https://doi.org/10.3390/biom12081107