
Multifaceted Roles of the N6-Methyladenosine RNA Methyltransferase METTL3 in Cancer and Immune Microenvironment
by
 ,
,
Chenxi Hu
1,2, 
Jiacheng Liu
1,3,
Yue Li
1,2,
Wei Jiang
1,2,
Ding Ji
1,2,
Wei Liu
1,* and
Teng Ma
4,* 1
Department of Immunology, School of Basic Medicine, Hebei Medical University, 361 Zhongshan East Rd., Shijiazhuang 050017, China
2
The First Hospital of Hebei Medical University, Shijiazhuang 050030, China
3
The Forth Hospital of Hebei Medical University, Shijiazhuang 050011, China
4
Cancer Research Center, Beijing Chest Hospital, Capital Medical University/Beijing Tuberculosis and Thoracic Tumor Research Institute, Beijing 101149, China
*
Authors to whom correspondence should be addressed.
Biomolecules 2022, 12(8), 1042; https://doi.org/10.3390/biom12081042
Submission received: 29 June 2022
/
Revised: 18 July 2022
/
Accepted: 18 July 2022
/
Published: 28 July 2022
(This article belongs to the Special Issue Metabolic Modifications in Genomic Stability Maintenance)
Abstract
:As the most abundant internal mRNA modification in eukaryotic cells, N6-methyladenosine (m6A) has emerged as an important regulator of gene expression and has a profound impact on cancer initiation and progression. mRNA m6A modification is regulated by m6A methyltransferases, demethylases and reader proteins to fine tune gene expression at the post-transcriptional level. The most well-studied m6A methyltransferase, METTL3, plays critical roles in regulating gene expression and affecting the outcome of various cancers. In this review, we discuss the multifaceted roles of METTL3 in regulating specific molecular signaling pathways in different types of cancers and the recent progress on how METTL3 impacts the tumor immune microenvironment. Finally, we discuss future directions and the potential for therapeutic targeting of METTL3 in cancer treatment.
1. Introduction
To date, more than 100 chemical modifications on RNA have been identified in all kingdoms of life [1]. Among these modifications, N6-methyladenosine (m6A) is one of the most abundant internal modifications in eukaryotic messenger and noncoding RNAs and plays important roles in regulating RNA metabolism, such as pre-mRNA splicing [2], mRNA nuclear export [3], mRNA stability [4], mRNA translation [5], miRNA maturation [6], and long noncoding RNA (lncRNA) processing [7]. By selectively ‘marking’ mRNA with m6A, the cell can control gene expression, which broadly affects many processes, including cancer progression [8,9], stress response [10], stem cell differentiation [11,12], and gametogenesis [13].
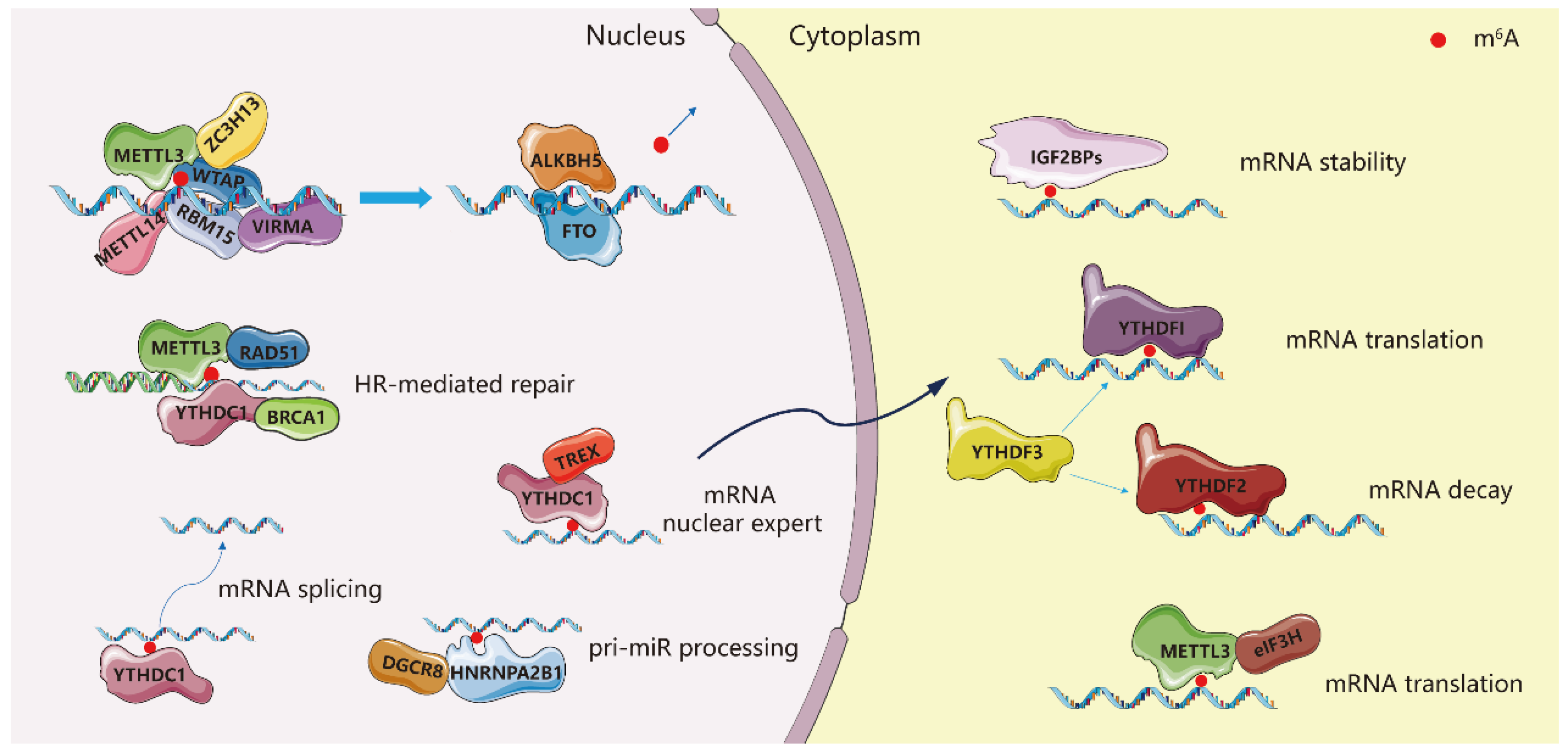
N6-methyladenosine is catalyzed by a large methyltransferase complex which is localized in the nucleus. The majority of research has focused on two main subunits of this complex—methyltransferase-like 3 (METTL3) and methyltransferase-like 14 (METTL14) [14,15]. METTL3 is the sole subunit capable of independently catalyzing the deposition of m6A on mRNA, but direct protein–protein interactions with METTL14 significantly improve catalytic efficiency compared to METTL3 alone.
Other subunits in the m6A methyltransferase complex, such as Wilm’s Tumor 1-Associating Protein (WTAP) [16], RNA Binding Motif Protein 15 (RBM15) [7], Zinc finger CCCH domain-containing protein 13 (ZC3H13) [17] and VIRMA (KIAA1429) [18], play regulatory roles that confer specificity for different target mRNAs or lncRNAs. Since m6A is involved in pre-mRNA splicing and miRNA maturation, the deposition of m6A on nascent RNA transcripts must occur rapidly. Indeed, m6A methylation of pre-mRNA and the binding of m6A-binding proteins such as hnRNPG on newly methylated transcripts occurs co-transcriptionally and before transcription termination [19,20]. There are reports that METTL3 can be bound to chromatin by interacting with histone modifications such as the transcription factor CEBPZ, setting the stage for rapid m6A methylation of certain actively transcribed genes [8].
Demethylases remove m6A modification from RNA, such as FTO and ALKBH5 [21,22]. The fate of methylated mRNA is decided by the binding of m6A-binding proteins such as YTH domain-containing proteins (YTHDF1, YTHDF2, YTHDF3, YTHDC1, and YTHDC2) [23,24,25], insulin-like growth factor 2 mRNA binding proteins (IGF2BP1 and IGF2BP2) and others [26]. Some readers affect the stability of methylated transcripts in the cytoplasm whereas others may affect splicing, nuclear export and miRNA processing [2,3,4]. Splicing of pre-mRNA can be promoted by FTO, YTHDC1 and HNRNPA2B1 [6,27,28]. To facilitate nuclear export of mRNA, the nuclear export complex TREX is recruited by the m6A methyltransferase complex and TREX also stimulates the recruitment of YTHDC1 to mRNA, which in turn can recruit SR proteins to promote export of m6A-modified mRNA [3,29]. METTL3 itself can facilitate mRNA translation by recruiting eIF3H to initiate translation in a manner independent of its catalytic activity [30] (Figure 1). In summary, throughout the various stages of pre-mRNA processing and pri-miRNA maturation, m6A modification is dynamically added, removed, and read by various enzymes responding to the present needs of the cell.
Furthermore, METTL3 is intimately involved in DNA damage repair [31]. Ultraviolet-induced cyclobutane pyrimidine and abasic sites with complete sugar-phosphate backbones are two important frequent DNA damages. Ultraviolet causes strand breaks, while abasic sites can lead to mutations and strand breaks [32]. The METTL3-METTL14 complex is recruited at the DNA-damaged sites to modulate nucleotide-excision repair and homologous recombination (HR)-mediated repair by recruiting RAD51 and BRCA1 (Figure 1). Mechanistically, the METTL3-METTL14 complex is active on unpaired DNA [33]; METTL3 is phosphorylated at S43 (serine phosphorylation sites) by ATM (ATM is a critical master regulator of DNA damage response signaling) and localized to double-strand breaks (DSBs) sites [34].
This review compiles and discusses the current literature regarding the diverse functions of METTL3 in cancer. It will also illuminate how METTL3 regulates the expression of genes and pathways in various cancers (Figure 2), which may be useful in identifying novel therapies for cancer.
2. Multiple Roles of the METTL3 in Cancers
2.1. Acute Myeloid Leukemia (AML)
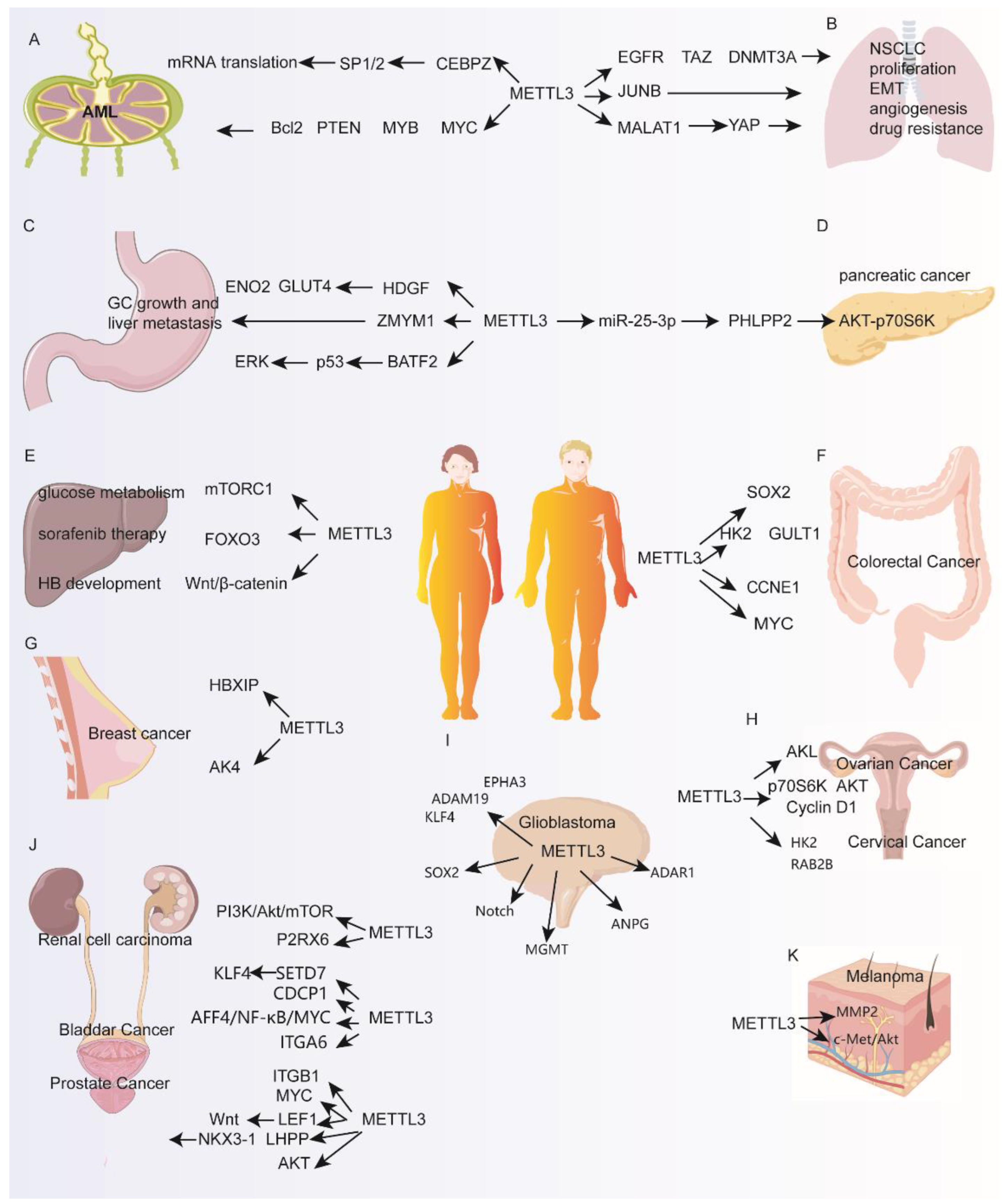
AML is a malignant cancer of myeloid hematopoietic stem or progenitor cells [35]. In normal human hematopoietic stem/progenitor cells (HSPCs), METTL3 inhibits differentiation and promotes proliferation, thus playing a crucial role in normal myeloid differentiation. AML cells have higher expression of METTL3 compared to HSPCs and a CRISPR dropout screen identified METTL3, METTL14, WTAP and KIAA1429 as essential for the survival of AML cells, indicating a potential oncogenic role for m6A modification in AML [36]. Indeed, genetic depletion of METTL3 in AML cell lines and primary leukemic blasts lead to cell growth inhibition, cell cycle arrest, induction of apoptosis and differentiation [8,36]. Mechanistically, METTL3’s oncogenic role in AML was linked to enhancing translation of anti-apoptotic and pro-growth transcripts such as c-MYC, MYB, PTEN and BCL2 in an m6A-dependent manner [36]. METTL3, but not METTL14, was also found to associate with chromatin at transcription initiating sites in a CEBPZ-dependent manner [8]. Promoter-bound METTL3 methylates the coding region of target mRNAs such as SP1 and SP2, leading to an increase in translation efficiency of these mRNAs, thus promoting AML cell survival [8] (Figure 2A). Taken together, AML cells expressing a higher abundance of METTL3 compared to HSPCs can utilize m6A modification to promote the expression of oncogenic pathways, thus promoting leukemia.
2.2. Lung Cancer
Lung cancer is one of the most prevalent malignant tumors, contributing to a quarter of all cancer deaths in the USA in 2020 [37]. In lung cancer, METTL3 but not METTL14 is significantly elevated in tumors compared to normal tissue [30]. Genetic ablation of METTL3 expression in A549 cells significantly reduces cell proliferation and survival. METTL3 exerts its oncogenic role in lung cancer by enhancing the translation of epidermal growth factor receptor (EGFR), transcriptional co-activator with PDZ-binding motif (TAZ) and DNA methyltransferase 3A (DNMT3A) in a methyltransferase- or m6A binding protein-independent manner by recruiting the translation initiation factor eIF3h to METTL3 bound transcripts [30]. In non-small-cell lung carcinoma (NSCLC), the expression of METTL3 was negatively correlated with miR-33a expression in 32 patient-derived tissues and showed that METTL3 is a potential target of miR-33a [38]. Overexpression of miR-33a in A549 or NCI-H460 cells led to decreased METTL3 expression at both the RNA and protein levels and decreased cell proliferation [38]. Furthermore, METTL3 promotes NSCLC apoptosis, migration and invasion by mediating mature miR-1246, which targets paternally expressed gene 3 (PEG3) [39]. METTL3 promotes metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) translation by recruiting YTHDF1/3. MALAT1 combines miR-1914-3p to increase yes association protein (YAP), inducing NSCLC metastasis and drug resistance [40]. Treatment of A549 and LC2/ad lung cancer cells with TGF-β caused upregulation of METTL3 and promoted epithelial to mesenchymal transition (EMT) [41]. Knockdown of METTL3 decreased m6A modification and mRNA stability of JUNB, one of the important transcription regulators of EMT, and also led to the expression changes in EMT-related genes such as E-cadherin, fibronectin and vimentin [41]. miR-143-3p/VASH1 was related to poor survival outcomes in lung cancer patients [42]. METTL3 elevates miR-143-3p expression by enhancing splicing of miR-143-3p to generate mature miRNA [42]. In turn, miR-143-3p can promote invasion in an in vitro blood–brain barrier model and inhibit vasohibin-1 (VASH1) expression to increase angiogenesis in lung cancer tissue [42]. Other studies found that METTL3 elevated the m6A level of Zinc finger and BTB domain-containing 4 (ZBTB4) mRNA in cigarette smoke extract-induced transformed HBE (T-HBE) cells. Thus, ZBTB4, a transcriptional repressor, was dependent on YTHDF2. Lower expression of ZBTB4 upregulated EZH2, which enhanced H3K27me3 combining with E-cadherin promoter, causing lower E-cadherin levels and EMT and T-HBE cell malignancy [43] (Figure 2B).
2.3. Gastric Cancer (GC)
METTL3 is upregulated in GC tumor cells and is an independent prognostic factor of poor survival and an effective predictor for severity of GC [44,45]. The suppression of METTL3 expression inhibits cell proliferation, migration and invasion of SGC7901 and AGS cells [44]. P300 was found to mediate H3K27 acetylation of the METTL3 promoter, thus upregulating METTL3 transcription. One of METTL3’s targets in GC cells, HDGF, is methylated by METTL3 and stabilized by the m6A reader IGF2BP3 [46]. HDGF can accelerate tumor angiogenesis and enhance glycolysis by increasing the expression of GLUT4 and ENO2 [46]. METTL3 also promoted liver metastasis in a mouse model of gastric cancer [46]. Another study demonstrated METTL3’s ability to facilitate EMT in vitro and promote metastasis in vivo [47]. Zinc finger MYM type containing 1 (ZMYM1) was identified as the target of METTL3 for m6A modification in GC cells [47]. Mechanistically, METTL3 methylates ZMYM1 mRNA and is stabilized by Human antigen R (HuR), which acts as a m6A “reader” protein located in the nucleus [47]. HuR is also well acknowledged as a general RNA binding protein [48]. In turn, ZMYM1 promotes EMT and metastasis by recruiting the CtBP/LSD1/CoREST complex to the E-cadherin promoter to repress its transcription. Importantly, this study reveals that the METTL3/ZMYM1/E-cadherin signaling pathway is a potential therapeutic target for GC metastasis [47]. Another study showed that BATF2 increases p53 protein stability, inhibiting the phosphorylation of extracellular signal-regulated kinase (ERK). METTL3 represses BATF2 expression, thus promoting GC development [49]. Silencing hepatitis B X-interacting protein (HBXIP) causes low METTL3 expression. METTL3 increases MYC translation by m6A methylation [44]. Embryonic ectoderm development protein (EED) increases METTL3 by methylating miR-338-5p. METTL3 increases oncogene CDCP1 [50], thus promoting GC cells proliferation, migration and invasion. (Figure 2C).
2.4. Pancreatic Cancer
Pancreatic cancer is a tumor of the digestive tract with lethal malignancy and poor prognosis with 5-year survival rates at 9% [37]. METTL3 expression and RNA m6A modification is higher in pancreatic cancer tissues compared to normal tissues and high METTL3 expression is related to high pathological stage and high lymph node metastasis. Conversely, METTL3 knockdown promoted pancreatic cancer cell proliferation, invasion, and migration in vitro [51]. Since cigarette smoking is linked to pancreatic cancer, a study found that cigarette smoke condensate can increase expression of METTL3 by causing DNA hypomethylation of the METTL3 promoter, thus allowing the transcription factor NFIC to activate METTL3 transcription [52]. Then, METTL3 promotes the maturation of miR-25-3p mediated by NF-κB associated protein (NKAP), which leads to increased carcinogenicity in pancreatic ductal epithelial cells [52]. High levels of miR-25-3p in smokers and in pancreatic cancer tissues are associated with poor prognosis in pancreatic cancer patients [52] (Figure 2D). miR-25-3p inhibits PHLPP2, thereby activating the oncogenic AKT-p70S6K signaling pathway, which induces malignant phenotypes of pancreatic cancer [52]. METTL3-depleted pancreatic cancer cells show higher sensitivity to anticancer reagents and irradiation. This means that METTL3 is probably a potential target for increasing therapeutic effect in patients with pancreatic cancer [53].
2.5. Hepatocellular Carcinoma (HCC) and Hepatoblastoma
Hepatocellular carcinoma (HCC) ranks as the sixth common malignancy worldwide and has higher mortality among patients [54]. METTL3 is expressed at higher levels in tumor samples compared to normal liver cells and higher expression of METTL3 is correlated with worse survival outcomes in HCC patients [55,56,57]. METTL3 knockdown of HEPG2 and Huh7 cells slows cell proliferation, reduces colony formation, suppresses cell migration and impairs in vivo tumor formation [57]. Mechanistically, METTL3 methylates the tumor suppressor SOCS2 and its mRNA is degraded in a YTHDF2-dependent manner, thus contributing to tumorigenesis [57]. In another study, METTL3 was found to methylate and stabilize LINC00958 lncRNA leading to its upregulation [56]. LINC00958 exerts oncogenic properties in HCC by downregulating miR-3619-5p to upregulate hepatoma-derived growth factor (HDGF) expression, leading to reprogramming of lipid metabolism to promote HCC [56]. METTL3 is also involved in elevating mTORC1 activity and glycolysis, thus contributing to tumor progression of HCC cell lines [55]. METTL3 also plays a role in sorafenib sensitivity in HCC as METTL3 knockdown cells exhibited a two-fold increase in the IC50 of sorafenib compared to METTL3 expressing cells [58]. Interestingly, METTL3 was also shown to suppress autophagy in HCC cells by methylating FOXO3 mRNA, leading to its upregulation in a YTHDF1-dependent manner [58]. This study identifies METTL3 and FOXO3 as critical regulators of autophagy-induced sorafenib resistance [58].
The m6A regulators METTL3, WTAP, FTO and YTHDF2 are elevated in tumor vs control tissues from hepatoblastoma patients and all four regulators contributed to the proliferation and colony formation of HepG2 cells [59]. This study chose to focus on the role of METTL3 due to elevated m6A levels in tumor vs normal patient total RNA samples although this was measured in only five patient samples. METTL3 knockdown decreased CTNNB1 mRNA methylation level and stability to inhibit Wnt/β-catenin signaling pathway in Huh6 and HepG2 cells [59]. In another study, METTL3 was identified as a direct target of microRNA-186, which is weakly expressed in hepatoblastoma tissue [60]. In addition, overexpression of miR-186 significantly inhibited the invasive phenotype in vitro and in vivo, which could be reversed by METTL3 overexpression. This study also corroborates with the previously mentioned study by demonstrating that the miR-186/METTL3 axis regulates the Wnt/β-catenin signaling pathway, contributing to tumorigenesis [60] (Figure 2E).
2.6. Colorectal Carcinoma (CRC)
METTL3 shows oncogenic effects in promoting the proliferation and metastasis of CRC using in vitro and in vivo models and could also act as a potential biomarker for CRC prognosis [61]. CRC metastasis tissues have high expression of METTL3 and elevated METTL3 correlates with poor prognosis [62]. Down regulation of METTL3 significantly inhibits the self-renewal and migration of CRC stem cells in vitro [62]. MeRIP-seq results showed that SOX2 is the downstream target gene of METTL3. Since SOX2 is an important transcription factor for maintaining self-renewal and proliferation of pluripotent stem cells, it plays an important part in the development of malignant tumors. The coding region of methylated SOX2 mRNA can be recognized by IGF2BP2 to inhibit the degradation of SOX2 mRNA. Supporting this, the expression of SOX2 target genes is positively correlated with METTL3 and IGF2BP2 in CRC [62]. In a separate study, METTL3 promotes activation of the glycolytic pathway in CRC patients by interacting directly with the 5′/3′UTR region of HK2 and the 3′UTR region of GLUT1. HK2 and GLUT1 transcripts are then stabilized by the m6A reader IGF2BP2 [61]. GLUT1 promotes glucose uptake and lactate production, which causes mTORC1 signaling activation and CRC development [63]. METTL3 and its pathway are potential therapeutic targets for CRC patients with high glucose metabolism [63]. Xiang et al. found that METTL3 promoted CRC progression through enhancing MYC expression in an IGF2BP1-dependent manner [64]. Another study demonstrated a link between the gut microbiome and cellular m6A levels by showing that butyrate, a bacterial short-chain fatty acid, decreases METTL3 levels in vitro, thus suppressing tumor cell growth [65]. Further elucidation of this mechanism of microbiome-mediated tumor suppression through m6A regulation would be an interesting avenue of research [65] (Figure 2F).
2.7. Breast Cancer
Breast cancer is the most common cancer in females in the USA [37]. Upregulation of METTL3 is observed in breast cancer tissues and cells and its elevated expression is correlated with worse patient survival [66]. Since breast cancer is a heterogenous disease divided into various molecular subtypes, METTL3 expression is found to be higher in normal-like and Luminal A/B subtypes compared to triple negative and HER2 positive subtypes [67]. Knockdown of METTL3 in Luminal A and triple negative cell lines can reduce methylation levels, reduce tumor cell proliferation, accelerate apoptosis and inhibit tumor growth in a BALB/c xenograft model [66]. Bcl-2 transcripts are upregulated following METTL3 methylation, indicating that METTL3 may promote breast cancer proliferation by regulating apoptosis [66]. Another study found that METTL3 expression in breast cancer tissues is positively correlated with HBXIP expression and HBXIP is indeed a target of METTL3 methylation [68]. HBXIP upregulates METTL3 by inhibiting miRNA let-7g, which downregulates METTL3 expression [68]. The positive feedback loop of HBXIP/let-7g/METTL3/HBXIP promotes the proliferation of breast cancer cells [68]. In one study, by utilizing a genetically defined immortalized and oncogenic transformed human mammary epithelial cell (HMEC) model, METTL3 expression was downregulated and m6A levels on mRNA were lower in immortalized and transformed HMECs compared to primary, untransformed HMECs [69]. Given the nature of this phenotype, the authors surprisingly found that overexpression of METTL3 and METTL14 in transformed cells but not immortalized cells can promote cell proliferation and migration [69]. These results show that the function of METTL3 in this model is nuanced, possibly playing different roles in primary HMECs vs transformed or immortalized HMECs [69]. Tamoxifen contributes to treating estrogen receptor (ER)-positive breast cancer. In tamoxifen-resistant MCF-7 cells, increased METTL3 confers a greater resistance to tamoxifen by promoting AK4 expression. AK4 inhibits mitochondrial apoptosis and promotes ROS production [70]. Another study found that METTL3 decreases Collagen type III alpha 1 chain (COL3A1), thus inhibiting triple-negative breast cancer (TNBC) cell metastasis [71]. The numerous genetic mutations present in breast cancers and the various molecular subtypes warrants a more in-depth analysis of the role of METTL3 and m6A in breast cancer (Figure 2G).
2.8. Ovarian Cancer
Using immunohistochemistry on 52 patient samples, in one study METTL3 was highly expressed in ovarian cancer tissues compared to adjacent normal tissues, and was significantly correlated to tumor grade [72]. METTL3 knockdown in SKOV3 and OVCAR3 cells caused decreased cell proliferation, increased apoptosis, and perturbed migration and invasion in Transwell assays due to lower levels of phosphorylated Akt [72]. In another study using a larger cohort of 162 ovarian carcinoma samples, METTL3 expression was positively correlated with tumor histological grades pT, pN, pM and FIGO stage, suggesting that METTL3 is important for metastasis of ovarian cancer [73]. This study further shows that METTL3 can promote AXL translation and the EMT process [73]. METTL3 accelerated the maturation of miR-126-5p via the m6A modification of pri-miR-126-5p. miR-126-5p can activate the PI3K/Akt/mTOR pathway by targeting PTEN, thus facilitating ovarian cancer cell proliferation, migration, and invasion [74]. These studies demonstrate the feasibility of METTL3 as a new prognostic indicator and therapeutic target in ovarian cancer (Figure 2H).
2.9. Cervical Cancer
METTL3 is independent indicator for poor prognosis in early stage cervical cancer patients [75]. METTL3 targets the 3′-UTR of hexokinase 2 (HK2) mRNA to enhance HK2 stability in a YTHDF1-dependent manner, which promotes the proliferation and aerobic glycolysis of cervical cancer cells [76]. Hu et al. found that METTL3 increased the mRNA stability of RAB2B in an IGF2BP3-dependent manner, which can facilitate cervical cancer development [77]. These studies suggest that METTL3 might be a potential target for cervical cancer therapy (Figure 2H).
2.10. Glioblastoma (GBM)
Glioma stem-like cells (GSCs) contribute to poor prognosis as they are responsible for the high rate of recurrence and chemotherapy resistance in GBM [78]. Since METTL3 has been shown to play a role in regulating stem cell pluripotency and differentiation [79], its role in GSCs has been studied. METTL3 plays a crucial role in the maintenance and dedifferentiation of GSCs [78,80]. Knockdown of METTL3 in GSCs promotes growth and self-renewal in vitro and sphere formation [80]. The expression of several oncogenes including ADAM metallopeptidase domain 19 (ADAM19), EPH receptor A3 (EPHA3) and Kruppel like factor 4 (KLF4) are elevated in METTL3 and METTL14 knockdown GSCs [80,81]. Conversely, a separate study suggested that METTL3 was upregulated in GBM tumors and its knockdown inhibited tumor growth in U87/shMETTL3-injected mice. Further research suggested that SOX2 stabilized by METTL3 with recruitment of HuR was related to the maintenance and radiation resistance of glioma stem-like cells [78]. Furthermore, METTL3 promotes the temozolomide (TMZ) resistance of glioma cells by increasing DNA repair genes O6-methylguanine-DNA methyltransferase (MGMT) and alkylpurine–DNA–N-glycosylase (ANPG) [82]. GSEA analysis of METTL3 knockdown vs control GSCs identified reduction of Notch signaling, a pathway promoting keratinocytes differentiation and inhibiting tumorigenesis including NOTCH1, NOTCH3, NOTCH4, and HES1. METTL3 knockdown downregulated NOTCH1 and HES1 protein expression [83]. METTL3 promotes cell proliferation dependent on YTHDF1 by targeting RNA-binding protein adenosine deaminases acting on RNA-1 (ADAR1) which catalyzes A-to-I RNA editing. Furthermore, ADAR1 stabilizes cyclin-dependent kinase 2 (CDK2) mRNA, which is an important cell cycle kinase that promotes cell proliferation. Interestingly, A-to-I RNA editing is decreased in GBM, because METTL3 boosts the amount of transcripts depositing m6A, inhibiting editing at multiple sites [84] (Figure 2I).
2.11. Renal Cell Carcinoma (RCC)
Renal cell carcinoma (RCC) is a frequent malignant tumor of the adult kidney [54]. Samples from a cohort of 145 patients found that high expression of METTL3 correlated with longer patient survival, indicating a tumor suppressive role for METTL3 in RCC [85]. Consistent with the clinical findings, METTL3 was expressed at lower levels in CAKI-1, CAKI-2, and ACHN RCC cell lines compared to a normal human renal tubular epithelial cell line, HK-2 [85]. In CAKI-1 and CAKI-2 cells, METTL3 was found to slow the proliferation, colony formation, migration, and invasion of RCC cells [85]. METTL3 mediates RCC cell proliferation possibly by negative regulation of the PI3K-Akt-mTOR pathway. In a recent TCGA analysis of kidney renal papillary cell carcinoma (KIRP) (n = 289, normal samples = 32), there was no significant different expression of METTL3, WTAP, RBM15, FTO YTHDC2 and YTHDC1 in KIRP compared to normal kidney tissue samples [86]. In another study, METTL14 decreased P2RX6 (ligand-gated ion channel receptors) expression activation-facilitated RCC cell migration and invasion by inhibiting ATP-induced Ca2+ influx to increase the ERK1/2 and MMP9 signaling pathway in SN12-PM6 and 786-O RCC cell lines compared to HK2 [87] (Figure 2J). In RCC, it is not known whether the methylation-dependent or methylation-independent mechanisms of METTL3 contribute to tumorigenesis. Given the controversial findings of these studies, more in-depth studies that experimentally determine the direct mRNA targets of METTL3 should be done to clarify the role of METTL3 in RCC.
2.12. Bladder Cancer
Compared with the paracancerous bladder urothelial tissue, the expression of METTL3 in the bladder cancer tissue is increased. Knockdown of METTL3 can significantly inhibit the proliferation, colony formation, migration and invasion of bladder cancer cells in vitro [88]. Using nude mice models, METTL3-depleted UM-UC-3 cells produced smaller tumors compared to control cells and had reduced metastasis [88]. METTL3 methylates the mRNA of the tumor suppressors SETD7 and KLF4, which are then degraded by YTHDF2 to promote the development of bladder cancer [88]. In a chemically induced bladder cancer model of normal human uroepithelial SV-HUC-1 cells and prostate epithelial RWPE-1 cells, METTL3 can upregulate CUB domain-containing protein 1 (CDCP1) in bladder cancer tissues using both methylation-dependent and methylation-independent mechanisms of METTL3 [89]. Inhibition of the METTL3-m6A-CDCP1 axis can slow down the growth and progression of bladder cancer cells [89]. This axis can also promote the malignant transformation of urothelial cells and the occurrence of bladder cancer in vivo and in vitro [89]. METTL3 can also promote the progression of bladder cancer through both the NF-κB and MYC signaling networks since AFF4, IKBKB, and p65 as well as MYC transcripts are methylated and stabilized by METTL3 [90]. METTL3 can enhance cell adhesion by upregulating ITGA6, which is linked to poor prognosis in bladder cancer [91]. Since ITGA6 enhances the growth and metastasis of bladder cancer cells, METTL3 knockout leads to reduced adhesion, proliferation, migration and invasion of bladder cancer cells [91]. METTL3 can downregulate PTEN expression by interacting with the microprocessor protein DGCR8 and positively regulating pri-miR221/222 processing in an m6A-dependent manner, thus exerting oncogenic effects in bladder cancer [92]. These results collectively suggest that METTL3 plays an important role in the progression of bladder cancer (Figure 2J).
2.13. Prostate Cancer
METTL3 is upregulated in prostate cancer cells compared to normal cells and is a poor prognostic factor for patient survival [93,94]. Genetic depletion and overexpression of METTL3 in various prostate cancer cell lines demonstrate that METTL3 can significantly promote the proliferation, migration and invasion of prostate cancer cells [93,94]. In terms of mechanisms, one study revealed that METTL3 can elevate the expression of MYC mRNA and protein levels by increasing the m6A level of the MYC transcript, which leads to carcinogenesis of prostate cancer [94]. Another study showed that METTL3 can methylate LEF1 mRNA leading to increased LEF1 protein expression. Since LEF1 is a positive regulator of the Wnt signaling pathway, METTL3 can promote prostate cancer progression through the METTL3-LEF1-Wnt axis [93]. METTL3 can also positively regulate the sonic hedgehog (SHH) pathway component GLI1, which is a transcription factor that activates c-Myc and cyclin D expression [95]. Li et al. showed that YTHDF2 degraded the tumor suppressors LHPP and NKX3-1 mRNA in a METTL3-m6A-dependent manner to regulate AKT phosphorylation-induced prostate tumor progression [96]. Another study found that METTL3 advanced the expression of Integrin β1 (ITGB1) in a m6A-HuR-dependent manner, promoting the bone metastasis of prostate cancer cells [97] (Figure 2J).
2.14. Melanoma
Studies have shown that melanoma cell lines express higher levels of METTL3 than normal melanocytes, resulting in increased m6A methylation on total RNA [98]. shRNA-mediated knockdown of METTL3 attenuates melanoma cell colony formation and invasion in vitro [98]. In A375 and WM793 cells, METTL3 regulated matrix metalloprotein-2 (MMP2) and N-cadherin protein levels, respectively [98]. Uveal melanoma (UM) is the most common intraocular tumor with high morbidity because of frequent metastasis [99]. METTL3 and METTL14 are significantly upregulated in UM cell lines compared to normal uveal cells [99]. METTL3 knockdown in UM cell lines decreases proliferation, colony formation, migration, and invasion by decreasing c-Met expression in a methylation dependent manner [99]. Since c-Met is upstream of the Akt pathway, METTL3 knockdown also decreases total Akt levels, indicating that the METTL3/c-Met/Akt axis is important for melanoma cell survival and migration. METTL3 inhibitor, cycloleucine, also shows similar effects to METTL3 knockdown cells, providing new insight for the treatment of UM [99] (Figure 2K).
3. METTL3 Functions in Tumor Immune Microenvironment
Though immunotherapy with programmed cell death-1 (PD-1) checkpoint blockade has achieved great progress in many types of cancers, a subset of cancer types still shows less or no response. Increasing evidence is depicting the roles of METTL3 in the tumor immune microenvironment and METTL3 as a potential therapeutic target for tumor immunotherapy.
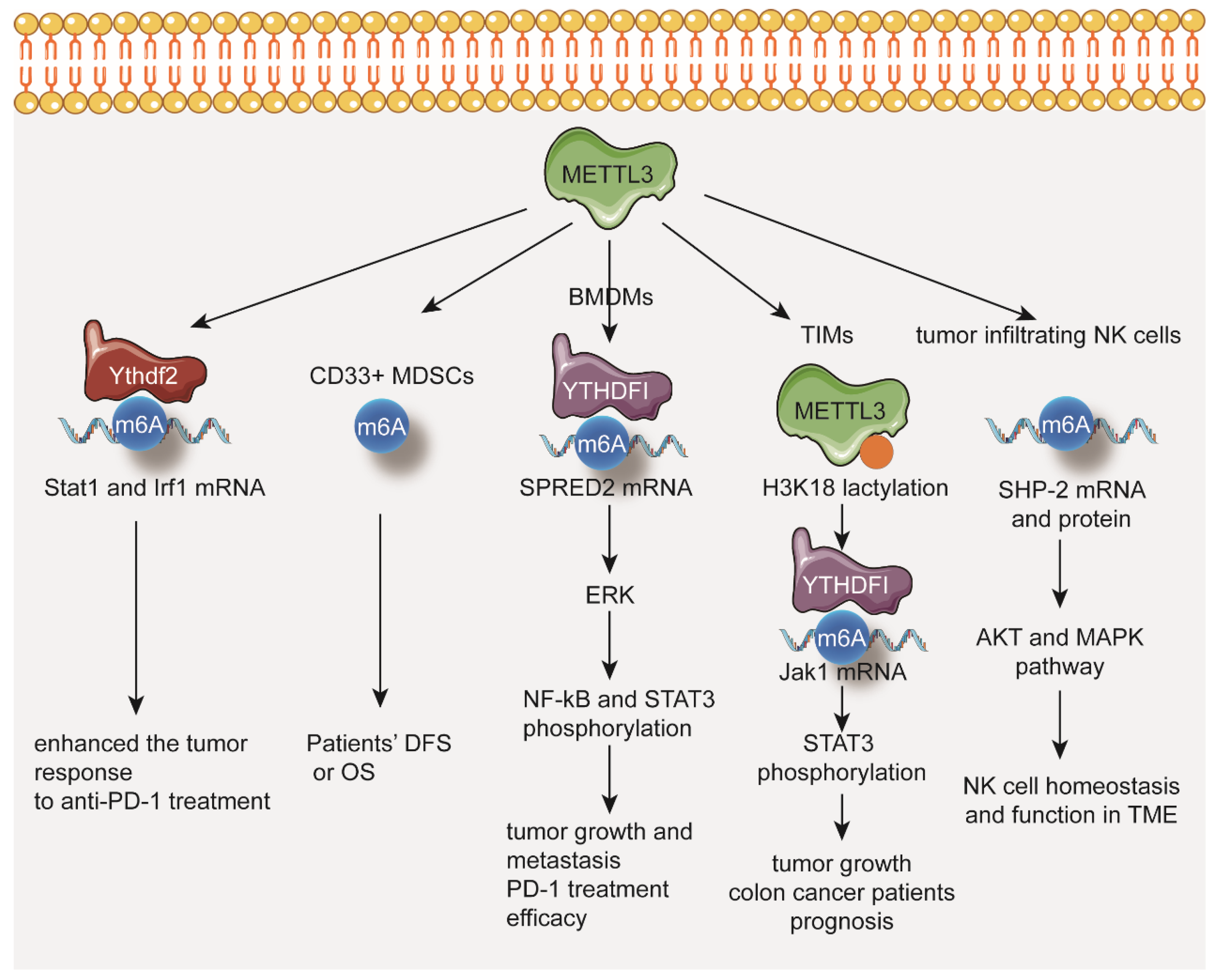
In mismatch-repair-proficient or microsatellite instability-low (pMMR-MSI-L) CRC and melanoma mouse models, depletion of METTL3 enhances the tumor response to anti-PD-1 treatment through stabilizing the Stat1 and Irf1 mRNA via YTHDF2 [100] (Figure 3).
In cervical cancer patients, the levels of METTL3 and CD33+ MDSCs in tumor tissues are significantly associated with reduced DFS or OS [101] (Figure 3).
In METTL3-deficient bone marrow-derived macrophages (BMDMs), the expression of M1-associated genes (Tnf-α and Il-6) and M2-associated genes (Arg1) is significantly increased. Loss of METTL3 impairs the YTHDF1-mediated translation of SPRED2, which increases the phosphorylation of NF-kB and STAT3 through the ERK pathway. Depletion of METTL3 in myeloid cells promotes tumor growth and metastasis. Furthermore, the efficacy of PD-1 checkpoint blockade is impaired in METTL3-deficient mice [102] (Figure 3).
Increased expression of METTL3 in tumor infiltrating myeloid cells (TIMs) correlates with the poor prognosis of colon cancer patients. Increasing lactate in tumor microenvironment induces METTL3 upregulation in TIMs via H3K18 lactylation. Depletion of METTL3 in myeloid cells decreases tumor growth in mice. METTL3 mediates m6A modification on Jak1 mRNA in TIMs and enhances JAK1 protein translation through the m6A-YTHDF1 axis, subsequently increasing phosphorylation of STAT3 (Figure 3).
A positive correlation between protein expression levels of METTL3 and effector molecules in tumor infiltrating NK cells has also been found. NK-cell-specific deletion of METTL3 in mice disrupts NK cell infiltration and function in the tumor microenvironment, leading to tumor progression. Mechanistically, METTL3 regulates SHP-2 m6A modification which subsequently stabilizes SHP-2 protein expression in NK cells. In METTL3-deficient NK cells, reduced SHP-2 activity suppresses the activation of the AKT and MAPK from IL-15 signaling (Figure 3).
These findings indicate that METTL3-mediated m6A methylation plays important functions in the tumor immune microenvironment.
4. Conclusions and Perspectives
Cancer cells utilize METTL3 to exploit specific molecular pathways to favor proliferation, metastasis, invasion, drug resistance, and the maintenance of cancer progenitor cells (see Table 1). Targeting m6A modifying enzymes such as METTL3 or METTL14 by selective inhibitors represents a promising state-of-the-art therapeutic strategy to treat cancers as it acts upstream of notable oncogenic pathways such as MYC, Akt and SHH. MYC is one of the most sought-after drug targets and developing specific MYC inhibitors has proved challenging. Since METTL3 is an upstream regulator of MYC in a number of cancers (AML, bladder cancer, and prostate cancer), targeting METTL3 can modulate this historically difficult drug target. Presently, multiple METTL3 specific inhibitors are already in various stages of development [103]. STM2457, a highly selective catalyst inhibitor of METTL3, results in decreased m6A levels of leukemogenic mRNAs, causing decreased growth, increased differentiation and apoptosis in a mouse AML model [104].
In addition, mRNA m6A methylation is interrelated with cellular immunity. Han, D et al. found that loss of YTHDF1 strengthened the function of dendritic cells tumor antigen cross-presentation and CD8+ T response to tumor [105]. Recent research has showed that FTO enhanced melanoma tumorigenesis and PD-1 antibody treatment controlled melanoma development in FTO knockdown tumors on the complete immunity system [106]. The significant investment and research into therapeutics targeting METTL3 underscores the novelty and promise of targeting mRNA modifications in cancer.
Author Contributions
Concept and design: T.M. and W.L. Administrative support: W.J. and D.J. Collection and assembly of data: C.H. Data analysis and interpretation: C.H., J.L., Y.L., W.L. and T.M. Manuscript writing: all authors. Final approval of manuscript: all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Lung Cancer Project from Beijing Municipal Health Commission 2020–2022.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| METTL3 | Methyltransferase-like 3 |
| YTHDF1 | YTH N6-Methyladenosine RNA Binding Protein 1 |
| YTHDF2 | YTH N6-Methyladenosine RNA Binding Protein 2 |
| YTHDC2 | YTH Domain Containing 2 |
| AML | Acute myeloid leukemia |
| HCC | Hepatocellular carcinoma |
| CRC | Colorectal carcinoma |
| RCC | Renal cell carcinoma |
References
- Lewis, C.; Pan, T.; Kalsotra, A. RNA modifications and structures cooperate to guide RNA–protein interactions. Nat. Rev. Mol. Cell Biol. 2017, 18, 202–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Lesbirel, S.; Viphakone, N.; Parker, M.; Parker, J.; Heath, C.; Sudbery, I.; Wilson, S.A. The m6A-methylase complex recruits TREX and regulates mRNA export. Sci. Rep. 2018, 8, 13827. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m6A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, D.P.; Chen, C.-K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m6A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, I.; Tzelepis, K.; Pandolfini, L.; Namshik, H.; Millan-Zambrano, G.; Robson, S.C.; Aspris, D.; Migliori, V.; Bannister, A.J.; Hannes, P.; et al. Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature 2017, 552, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Choe, J.; Lin, S.; Zhang, W.; Liu, Q.; Wang, L.; Ramirez-Moya, J.; Du, P.; Kim, W.; Tang, S.; Sliz, P.; et al. mRNA circularization by METTL3–eIF3h enhances translation and promotes oncogenesis. Nature 2018, 561, 556–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.; Qian, S.-B. Dynamic m6A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Chen, Y.; Sun, B.; Wang, L.; Yang, Y.; Ma, D.; Lv, J.; Yusheng, C.; Ding, Y.; Xue, Y.; et al. m6A modulates haematopoietic stem and progenitor cell specification. Nature 2017, 549, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Batista, P.J.; Molinie, B.; Wang, J.; Qu, K.; Zhang, J.; Li, L.; Bouley, D.M.; Lujan, E.; Haddad, B.; Daneshvar, K.; et al. m6A RNA Modification Controls Cell Fate Transition in Mammalian Embryonic Stem Cells. Cell Stem Cell 2014, 15, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.-M.; Li, C.J.; Vagbo, C.B.; Shi, Y.; Wang, W.-L.; Song, S.-H.; et al. ALKBH5 Is a Mammalian RNA Demethylase that Impacts RNA Metabolism and Mouse Fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3–METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Feng, J.; Xue, Y.; Guan, Z.; Zhang, D.; Liu, Z.; Gong, Z.; Wang, Q.; Huang, J.; Tang, C.; et al. Structural basis of N6-adenosine methylation by the METTL3–METTL14 complex. Nature 2016, 534, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.-L.; Sun, B.-F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Lv, R.; Ma, H.; Shen, H.; He, C.; Wang, J.; Jiao, F.; Liu, H.; Yang, P.; Tan, L.; et al. Zc3h13 Regulates Nuclear RNA m6A Methylation and Mouse Embryonic Stem Cell Self-Renewal. Mol. Cell 2018, 69, 1028–1038.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.; Mumbach, M.R.; Jovanovic, M.; Wang, T.; Maciag, K.; Bushkin, G.G.; Mertins, P.; Ter-Ovanesyan, D.; Habib, N.; Cacchiarelli, D.; et al. Perturbation of m6A Writers Reveals Two Distinct Classes of mRNA Methylation at Internal and 5′ Sites. Cell Rep. 2014, 8, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slobodin, B.; Han, R.; Calderone, V.; Vrielink, J.A.O.; Loayza-Puch, F.; Elkon, R.; Agami, R. Transcription Impacts the Efficiency of mRNA Translation via Co-transcriptional N6-adenosine Methylation. Cell 2017, 169, 326–337.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.I.; Shi, H.; Lyu, R.; Wylder, A.C.; Matuszek, Z.; Pan, J.N.; He, C.; Parisien, M.; Pan, T. Regulation of Co-transcriptional Pre-mRNA Splicing by m6A through the Low-Complexity Protein hnRNPG. Mol. Cell 2019, 76, 70–81.e9. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yan, J.; Li, Q.; Li, J.; Gong, S.; Zhou, H.; Gan, J.; Jiang, H.; Jia, G.-F.; Luo, C.; et al. Meclofenamic acid selectively inhibits FTO demethylation of m6A over ALKBH5. Nucleic Acids Res. 2015, 43, 373–384. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887, Erratum in Nat. Chem. Biol. 2012, 8, 1008. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Zhang, Z.; Theler, D.; Kaminska, K.H.; Hiller, M.; de la Grange, P.; Pudimat, R.; Rafalska, I.; Heinrich, B.; Bujnicki, J.M.; Allain, F.H.-T.; et al. The YTH Domain Is a Novel RNA Binding Domain. J. Biol. Chem. 2010, 285, 14701–14710. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295, Correction in Nat. Cell Biol. 2018, 20, 1098; Correction in Nat. Cell Biol. 2020, 22, 1288. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.-S.; Hao, Y.-J.; Sun, B.-F.; Sun, H.-Y.; Li, A.; Ping, X.-L.; Lai, W.-Y.; et al. Nuclear m 6 A Reader YTHDC1 Regulates mRNA Splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Yang, Y.; Sun, B.-F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.-J.; Ping, X.-L.; Chen, Y.-S.; Wang, W.-J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Luo, G.-Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. YTHDC1 mediates nuclear export of N6-methyladenosine methylated mRNAs. eLife 2017, 6, e31311. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Choe, J.; Du, P.; Triboulet, R.; Gregory, R.I. The m 6 A Methyltransferase METTL3 Promotes Translation in Human Cancer Cells. Mol. Cell 2016, 62, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Laurent, B.; Hsu, C.-H.; Nachtergaele, S.; Lu, Z.; Sheng, W.; Xu, C.; Chen, H.; Ouyang, J.; Wang, S.; et al. RNA m6A methylation regulates the ultraviolet-induced DNA damage response. Nature 2017, 543, 573–576. [Google Scholar] [CrossRef]
- Thompson, P.S.; Cortez, D. New insights into abasic site repair and tolerance. DNA Repair 2020, 90, 102866. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, C.B.; Yu, D.; Hajian, T.; Li, J.; Huang, Y.; Dai, N.; Corrêa, I.R., Jr.; Wu, T.; Vedadi, M.; Zhang, X.; et al. Human MettL3–MettL14 complex is a sequence-specific DNA adenine methyltransferase active on single-strand and unpaired DNA in vitro. Cell Discov. 2019, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, L.; Peng, D.; Jiang, A.; He, Y.; Zeng, Y.; Xie, C.; Zhou, H.; Luo, X.; Liu, H.; et al. METTL3 and N6-Methyladenosine Promote Homologous Recombination-Mediated Repair of DSBs by Modulating DNA-RNA Hybrid Accumulation. Mol. Cell 2020, 79, 425–442.e7. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Su, R.; Weng, H.; Huang, H.; Li, Z.; Chen, J. RNA N6-methyladenosine modification in cancers: Current status and perspectives. Cell Res. 2018, 28, 507–517. [Google Scholar] [CrossRef]
- Vu, L.P.; Pickering, B.F.; Cheng, Y.; Zaccara, S.; Nguyen, D.; Minuesa, G.; Chou, T.; Chow, A.; Saletore, Y.; Mackay, M.; et al. The N6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat. Med. 2017, 23, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Du, M.; Zhang, Y.; Mao, Y.; Mou, J.; Zhao, J.; Xue, Q.; Wang, D.; Huang, J.; Gao, S.; Gao, Y. MiR-33a suppresses proliferation of NSCLC cells via targeting METTL3 mRNA. Biochem. Biophys. Res. Commun. 2017, 482, 582–589. [Google Scholar] [CrossRef]
- Huang, S.; Luo, S.; Gong, C.; Liang, L.; Xiao, Y.; Li, M.; He, J. MTTL3 upregulates microRNA-1246 to promote occurrence and progression of NSCLC via targeting paternally expressed gene 3. Mol. Ther. Nucleic Acids 2021, 24, 542–553. [Google Scholar] [CrossRef]
- Jin, D.; Guo, J.; Wu, Y.; Du, J.; Yang, L.; Wang, X.; Di, W.; Hu, B.; An, J.; Kong, L.; et al. m6A mRNA methylation initiated by METTL3 directly promotes YAP translation and increases YAP activity by regulating the MALAT1-miR-1914-3p-YAP axis to induce NSCLC drug resistance and metastasis. J. Hematol. Oncol. 2019, 12, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanna-Udom, S.; Terashima, M.; Lyu, H.; Ishimura, A.; Takino, T.; Sakari, M.; Tsukahara, T.; Suzuki, T. The m6A methyltransferase METTL3 contributes to Transforming Growth Factor-beta-induced epithelial-mesenchymal transition of lung cancer cells through the regulation of JUNB. Biochem. Biophys. Res. Commun. 2020, 524, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Deng, Q.; Lv, Z.; Ling, Y.; Hou, X.; Chen, Z.; Dinglin, X.; Ma, S.; Li, D.; Wu, Y.; et al. N6-methyladenosine induced miR-143-3p promotes the brain metastasis of lung cancer via regulation of VASH1. Mol. Cancer 2019, 18, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.; Wu, Y.; Xiao, T.; Xue, J.; Sun, J.; Xia, H.; Ma, H.; Lu, L.; Li, J.; Shi, A.; et al. METTL3-mediated m6A modification of ZBTB4 mRNA is involved in the smoking-induced EMT in cancer of the lung. Mol. Ther. Nucleic Acids 2021, 23, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-D.; Chen, Z.-H.; Yu, K.; Lu, J.-H.; Wu, Q.-N.; Wang, Y.; Ju, H.-Q.; Xu, R.-H.; Liu, Z.-X.; Zeng, Z.-L. METTL3 Promotes the Progression of Gastric Cancer via Targeting the MYC Pathway. Front. Oncol. 2020, 10, 115. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Yang, S.; Sui, J.; Xu, S.; Cheng, Y.; Shen, B.; Zhang, Y.; Zhang, X.; Yin, L.; Pu, Y.; et al. Dysregulated N6-methyladenosine methylation writer METTL3 contributes to the proliferation and migration of gastric cancer. J. Cell. Physiol. 2020, 235, 548–562. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, C.; Ding, Q.; Zhao, Y.; Wang, Z.; Chen, J.; Jiang, Z.; Zhang, Y.; Xu, G.; Zhang, J.; et al. METTL3-mediated m6A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut 2019, 69, 1193–1205. [Google Scholar] [CrossRef]
- Yue, B.; Song, C.; Yang, L.; Cui, R.; Cheng, X.; Zhang, Z.; Zhao, G. METTL3-mediated N6-methyladenosine modification is critical for epithelial-mesenchymal transition and metastasis of gastric cancer. Mol. Cancer 2019, 18, 142. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Xu, L. The RNA-binding protein HuR in human cancer: A friend or foe? Adv. Drug Deliv. Rev. 2022, 184, 114179. [Google Scholar] [CrossRef]
- Xie, J.-W.; Huang, X.-B.; Chen, Q.-Y.; Ma, Y.-B.; Zhao, Y.-J.; Liu, L.-C.; Wang, J.-B.; Lin, J.-X.; Lu, J.; Cao, L.-L.; et al. m6A modification-mediated BATF2 acts as a tumor suppressor in gastric cancer through inhibition of ERK signaling. Mol. Cancer 2020, 19, 114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yan, Y.; Cao, X.; Zhang, J.; Li, Y.; Guo, C. Methylation of microRNA-338-5p by EED promotes METTL3-mediated translation of oncogene CDCP1 in gastric cancer. Aging 2021, 13, 12224–12238. [Google Scholar] [CrossRef]
- Xia, T.; Wu, X.; Cao, M.; Zhang, P.; Shi, G.; Zhang, J.; Lu, Z.; Wu, P.; Cai, B.; Miao, Y.; et al. The RNA m6A methyltransferase METTL3 promotes pancreatic cancer cell proliferation and invasion. Pathol. Res. Pract. 2019, 215, 152666. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bai, R.; Li, M.; Ye, H.; Wu, C.; Wang, C.; Li, S.; Tan, L.; Mai, D.; Li, G.; et al. Excessive miR-25-3p maturation via N6-methyladenosine stimulated by cigarette smoke promotes pancreatic cancer progression. Nat. Commun. 2019, 10, 1858. [Google Scholar] [CrossRef] [PubMed]
- Taketo, K.; Konno, M.; Asai, A.; Koseki, J.; Toratani, M.; Satoh, T.; Doki, Y.; Mori, M.; Ishii, H.; Ogawa, K. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int. J. Oncol. 2018, 52, 621–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Wei, X.; Jian, Z.; Zhang, X. METTL3 expression is associated with glycolysis metabolism and sensitivity to glycolytic stress in hepatocellular carcinoma. Cancer Med. 2020, 9, 2859–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, X.; Chen, Z.; Gao, W.; Zhang, Y.; Wang, J.; Wang, J.; Cao, M.; Cai, J.; Wu, J.; Wang, X. M6A-mediated upregulation of LINC00958 increases lipogenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. J. Hematol. Oncol. 2020, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wei, L.; Law, C.-T.; Tsang, F.H.-C.; Shen, J.; Cheng, C.L.-H.; Tsang, L.-H.; Ho, D.W.H.; Chiu, D.K.-C.; Lee, J.M.-F.; et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Niu, Y.; Wan, A.; Chen, D.; Liang, H.; Chen, X.; Sun, L.; Zhan, S.; Chen, L.; Cheng, C.; et al. RNA m6A methylation regulates sorafenib resistance in liver cancer through FOXO 3-mediated autophagy. EMBO J. 2020, 39, e103181. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, J.; Sun, G.; Wu, Q.; Ma, J.; Zhang, X.; Huang, N.; Bian, Z.; Gu, S.; Xu, M.; et al. m6A mRNA methylation regulates CTNNB1 to promote the proliferation of hepatoblastoma. Mol. Cancer 2019, 18, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Wang, Z.; Li, J.; Zhu, J.; Ren, Z.; Zhang, D.; Zhao, W.; Fan, Y.; Zhang, D.; Sun, R. Cross talk between RNA N6-methyladenosine methyltransferase-like 3 and miR-186 regulates hepatoblastoma progression through Wnt/β-catenin signalling pathway. Cell Prolif. 2020, 53, e12768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Xuan, B.; Yan, T.; Ma, Y.; Xu, P.; Tian, X.; Zhang, X.; Cao, Y.; Ma, D.; Zhu, X.; et al. m6A-dependent glycolysis enhances colorectal cancer progression. Mol. Cancer 2020, 19, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Hu, P.-S.; Zuo, Z.; Lin, J.-F.; Li, X.; Wu, Q.-N.; Chen, Z.-H.; Zeng, Z.-L.; Wang, F.; Zheng, J.; et al. METTL3 facilitates tumor progression via an m6A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol. Cancer 2019, 18, 112. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gao, S.; Liu, W.; Wong, C.-C.; Wu, J.; Liu, D.; Gou, H.; Kang, W.; Zhai, J.; Li, C.; et al. RNA N6-Methyladenosine Methyltransferase METTL3 Facilitates Colorectal Cancer by Activating the m6A-GLUT1-mTORC1 Axis and Is a Therapeutic Target. Gastroenterology 2021, 160, 1284–1300.e16. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Liang, X.; Yin, S.; Liu, J.; Xiang, Z. N6-methyladenosine methyltransferase METTL3 promotes colorectal cancer cell proliferation through enhancing MYC expression. Am. J. Transl. Res. 2020, 12, 1789–1806. [Google Scholar] [PubMed]
- Zhu, W.; Si, Y.; Xu, J.; Lin, Y.; Wang, J.; Cao, M.; Sun, S.; Ding, Q.; Zhu, L.; Wei, J. Methyltransferase like 3 promotes colorectal cancer proliferation by stabilizing CCNE1 mRNA in an m6A-dependent manner. J. Cell. Mol. Med. 2020, 24, 3521–3533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Xu, B.; Shi, J. N6-methyladenosine METTL3 promotes the breast cancer progression via targeting Bcl-2. Gene 2020, 722, 144076. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wu, D.; Ning, J.; Liu, W.; Zhang, D.; Wu, L.; Wu, D.; Ning, J.; Liu, W.; Zhang, D. Changes of N6-methyladenosine modulators promote breast cancer progression. BMC Cancer 2019, 19, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Wang, X.; Cao, C.; Gao, Y.; Zhang, S.; Yang, Z.; Liu, Y.; Zhang, X.; Zhang, W.; Ye, L. HBXIP-elevated methyltransferase METTL3 promotes the progression of breast cancer via inhibiting tumor suppressor let-7g. Cancer Lett. 2018, 415, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Fry, N.J.; Law, B.A.; Ilkayeva, O.R.; Carraway, K.R.; Holley, C.; Mansfield, K.D. N6-methyladenosine contributes to cellular phenotype in a genetically-defined model of breast cancer progression. Oncotarget 2018, 9, 31231–31243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Gonzalez, G.; Dai, X.; Miao, W.; Yuan, J.; Huang, M.; Bade, D.; Li, L.; Sun, Y.; Wang, Y. Adenylate Kinase 4 Modulates the Resistance of Breast Cancer Cells to Tamoxifen through an m6A-Based Epitranscriptomic Mechanism. Mol. Ther. 2020, 28, 2593–2604. [Google Scholar] [CrossRef]
- Shi, Y.; Zheng, C.; Jin, Y.; Bao, B.; Wang, D.; Hou, K.; Feng, J.; Tang, S.; Qu, X.; Liu, Y.; et al. Reduced Expression of METTL3 Promotes Metastasis of Triple-Negative Breast Cancer by m6A Methylation-Mediated COL3A1 Up-Regulation. Front. Oncol. 2020, 10, 1126. [Google Scholar] [CrossRef]
- Liang, S.; Guan, H.; Lin, X.; Li, N.; Geng, F.; Li, J. METTL3 serves an oncogenic role in human ovarian cancer cells partially via the AKT signaling pathway. Oncol. Lett. 2020, 19, 3197–3204. [Google Scholar] [CrossRef]
- Hua, W.; Zhao, Y.; Jin, X.; Yu, D.; He, J.; Xie, D.; Duan, P. METTL3 promotes ovarian carcinoma growth and invasion through the regulation of AXL translation and epithelial to mesenchymal transition. Gynecol. Oncol. 2018, 151, 356–365. [Google Scholar] [CrossRef]
- Bi, X.; Lv, X.; Liu, D.; Guo, H.; Yao, G.; Wang, L.; Liang, X.; Yang, Y. METTL3-mediated maturation of miR-126-5p promotes ovarian cancer progression via PTEN-mediated PI3K/Akt/mTOR pathway. Cancer Gene Ther. 2020, 28, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, Y.; Fang, Y.; Ma, S.; Zheng, H.; Liu, K.; Wang, R. Elevated Expression of Inhibitor of Apoptosis-stimulating Protein of p53 (iASPP) and Methyltransferase-like 3 (METTL3) Correlate with Poor Prognosis in FIGO Ib1-IIa Squamous Cell Cervical Cancer. J. Cancer 2020, 11, 2382–2389. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, X.; Li, L.; Gao, Z.; Su, X.; Ji, M.; Liu, J. N6-methyladenosine METTL3 promotes cervical cancer tumorigenesis and Warburg effect through YTHDF1/HK2 modification. Cell Death Dis. 2020, 11, 911. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, Y.; Huang, Y.; Jin, Z.; Wang, C.; Wang, H.; Xu, J. METTL3 regulates the malignancy of cervical cancer via post-transcriptional regulation of RAB2B. Eur. J. Pharmacol. 2020, 879, 173134. [Google Scholar] [CrossRef]
- Visvanathan, A.; Patil, V.; Arora, A.; Hegde, A.S.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Essential role of METTL3-mediated m6A modification in glioma stem-like cells maintenance and radioresistance. Oncogene 2018, 37, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.S.; He, C. Fate by RNA methylation: m6A steers stem cell pluripotency. Genome Biol. 2015, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- Cui, Q.; Shi, H.; Ye, P.; Li, L.; Qu, Q.; Sun, G.; Lu, Z.; Huang, Y.; Yang, C.-G.; Riggs, A.D.; et al. m6A RNA Methylation Regulates the Self-Renewal and Tumorigenesis of Glioblastoma Stem Cells. Cell Rep. 2017, 18, 2622–2634. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.-C.; Tsai, C.-Y.; Tsai, M.-M.; Lin, K.-H. Impact of DNA and RNA Methylation on Radiobiology and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Chen, G.; Dong, X.; Li, H.; Li, S.; Cheng, S.; Li, Y.; Wang, L.; Yuan, J.; Qian, Z.; et al. METTL3 Promotes the Resistance of Glioma to Temozolomide via Increasing MGMT and ANPG in a m6A Dependent Manner. Front. Oncol. 2021, 11, 702983. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, A.; Patil, V.; Abdulla, S.; Hoheisel, J.D.; Somasundaram, K. N6-Methyladenosine Landscape of Glioma Stem-Like Cells: METTL3 Is Essential for the Expression of Actively Transcribed Genes and Sustenance of the Oncogenic Signaling. Genes 2019, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Tassinari, V.; Cesarini, V.; Tomaselli, S.; Ianniello, Z.; Silvestris, D.A.; Ginistrelli, L.C.; Martini, M.; De Angelis, B.; De Luca, G.; Vitiani, L.R.; et al. ADAR1 is a new target of METTL3 and plays a pro-oncogenic role in glioblastoma by an editing-independent mechanism. Genome Biol. 2021, 22, 51. [Google Scholar] [CrossRef]
- Li, X.; Tang, J.; Huang, W.; Wang, F.; Li, P.; Qin, C.; Qin, Z.; Zou, Q.; Wei, J.; Hua, L.; et al. The M6A methyltransferase METTL3: Acting as a tumor suppressor in renal cell carcinoma. Oncotarget 2017, 8, 96103–96116. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Jing, C.; Xiao, C.; Li, T.; Wang, Y. Prognostic risk signature based on the expression of three m6A RNA methylation regulatory genes in kidney renal papillary cell carcinoma. Aging 2020, 12, 22078–22094. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Zhang, J.; Chen, Y.; Xu, Y.; Ma, J.; Hu, G.; Huang, Y.; Zheng, J.; Zhai, W.; Xue, W. The m6A-suppressed P2RX6 activation promotes renal cancer cells migration and invasion through ATP-induced Ca2+ influx modulating ERK1/2 phosphorylation and MMP9 signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Li, J.; Ying, Y.; Yan, H.; Jin, K.; Ma, X.; He, L.; Xu, X.; Liu, B.; Wang, X.; et al. METTL3/YTHDF2 m6A axis promotes tumorigenesis by degrading SETD7 and KLF4 mRNAs in bladder cancer. J. Cell. Mol. Med. 2020, 24, 4092–4104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Jin, H.; Que, B.; Chao, Y.; Zhang, H.; Ying, X.; Zhou, Z.; Yuan, Z.; Su, J.; Wu, B.; et al. Dynamic m6A mRNA methylation reveals the role of METTL3-m6A-CDCP1 signaling axis in chemical carcinogenesis. Oncogene 2019, 38, 4755–4772. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Sheng, L.; Gao, Q.; Xiong, Q.; Zhang, H.; Wu, M.; Liang, Y.; Zhu, F.; Zhang, Y.; Zhang, X.; et al. The m6A methyltransferase METTL3 promotes bladder cancer progression via AFF4/NF-κB/MYC signaling network. Oncogene 2019, 38, 3667–3680. [Google Scholar] [CrossRef]
- Jin, H.; Ying, X.; Que, B.; Wang, X.; Chao, Y.; Zhang, H.; Yuan, Z.; Qi, D.; Lin, S.; Min, W.; et al. N6-methyladenosine modification of ITGA6 mRNA promotes the development and progression of bladder cancer. eBioMedicine 2019, 47, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Wang, J.-Z.; Yang, X.; Yu, H.; Zhou, R.; Lu, H.-C.; Yuan, W.-B.; Lu, J.-C.; Zhou, Z.-J.; Lu, Q.; et al. METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner. Mol. Cancer 2019, 18, 110. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.-X.; Cao, Z.-G.; Zhao, S.-L. m6A methyltransferase METTL3 promotes the progression of prostate cancer via m6A-modified LEF1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3565–3571. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Du, Y.; Wang, L.; Liu, X. The M6A methyltransferase METTL3 promotes the development and progression of prostate carcinoma via mediating MYC methylation. J. Cancer 2020, 11, 3588–3595. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Yang, F.; Zhan, H.; Situ, J.; Li, W.; Mao, Y.; Luo, Y. RNA m6A Methyltransferase METTL3 Promotes The Growth Of Prostate Cancer By Regulating Hedgehog Pathway. OncoTargets Ther. 2019, 12, 9143–9152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xie, H.; Ying, Y.; Chen, H.; Yan, H.; He, L.; Xu, M.; Xu, X.; Liang, Z.; Liu, B.; et al. YTHDF2 mediates the mRNA degradation of the tumor suppressors to induce AKT phosphorylation in N6-methyladenosine-dependent way in prostate cancer. Mol. Cancer 2020, 19, 152. [Google Scholar] [CrossRef]
- Li, E.; Wei, B.; Wang, X.; Kang, R. METTL3 enhances cell adhesion through stabilizing integrin β1 mRNA via an m6A-HuR-dependent mechanism in prostatic carcinoma. Am. J. Cancer Res. 2020, 10, 1012–1025. [Google Scholar]
- Dahal, U.; Le, K.; Gupta, M. RNA m6A methyltransferase METTL3 regulates invasiveness of melanoma cells by matrix metallopeptidase 2. Melanoma Res. 2019, 29, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Xu, W.; Zhao, Y.; Jin, S.; Wang, S.; Liu, Q.; Chen, X.; Wang, J.; Dong, F.; Hu, D.; et al. RNA m6A methylation regulates uveal melanoma cell proliferation, migration, and invasion by targeting c-Met. J. Cell. Physiol. 2020, 235, 7107–7119. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hui, H.; Agrawal, K.; Kang, Y.; Li, N.; Tang, R.; Yuan, J.; Rana, T.M. m6A RNA methyltransferases METTL3/14 regulate immune responses to anti-PD-1 therapy. EMBO J. 2020, 39, e104514. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.-H.; Zhang, L.; Huang, H.; Dai, S.-Q.; Li, J. Connecting METTL3 and intratumoural CD33+ MDSCs in predicting clinical outcome in cervical cancer. J. Transl. Med. 2020, 18, 393. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhang, X.; Yang, P.; Zhang, X.; Peng, Y.; Li, D.; Yu, Y.; Wu, Y.; Wang, Y.; Zhang, J.; et al. RNA m6A methylation orchestrates cancer growth and metastasis via macrophage reprogramming. Nat. Commun. 2021, 12, 1394. [Google Scholar] [CrossRef] [PubMed]
- Cully, M. Chemical inhibitors make their RNA epigenetic mark. Nat. Rev. Drug Discov. 2019, 18, 892–894. [Google Scholar] [CrossRef]
- Yankova, E.; Blackaby, W.; Albertella, M.; Rak, J.; De Braekeleer, E.; Tsagkogeorga, G.; Pilka, E.S.; Aspris, D.; Leggate, D.; Hendrick, A.G.; et al. Small-molecule inhibition of METTL3 as a strategy against myeloid leukaemia. Nature 2021, 593, 597–601. [Google Scholar] [CrossRef]
- Han, D.; Liu, J.; Chen, C.; Dong, L.; Liu, Y.; Chang, R.; Huang, X.; Wang, J.; Dougherty, U.; Bissonnette, M.B.; et al. Anti-tumour immunity controlled through mRNA m6A methylation and YTHDF1 in dendritic cells. Nature 2019, 566, 270–274. [Google Scholar] [CrossRef]
- Yang, S.; Wei, J.; Cui, Y.-H.; Park, G.; Shah, P.; Deng, Y.; Aplin, A.E.; Lu, Z.; Hwang, S.; He, C.; et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat. Commun. 2019, 10, 2782. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
METTL3-dependent biological functions in cells.

Figure 2.
Multifaceted functions of METTL3 in different types of cancer. (A–K) propose how METTL3 regulates various genes and molecular pathways. AML, Acute myeloid leukemia; NSCLC, non-small-cell lung carcinoma; GC, Gastric cancer.
Figure 2.
Multifaceted functions of METTL3 in different types of cancer. (A–K) propose how METTL3 regulates various genes and molecular pathways. AML, Acute myeloid leukemia; NSCLC, non-small-cell lung carcinoma; GC, Gastric cancer.

Figure 3.
METTL3 in regulation of tumor immune microenvironment.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Expression, role, biological performance and molecular mechanisms of METTL3 in various cancers.
Table 1.
Expression, role, biological performance and molecular mechanisms of METTL3 in various cancers.
| Cancer Type | Expression | Role | Biological Performance | Molecular Mechanism | Ref. |
|---|---|---|---|---|---|
| Acute myeloid leukemia | Upregulated | Oncogene | Stem cell differentiation, proliferation, cell cycle, tumorigenesis | c-MYC, MYB, PTEN, BCL2, SP1 and SP2, ect | [8,36] |
| Lung cancer | Upregulated | Oncogene | Survival, proliferation, migration, invasion, metastasis, angiogenesis | EGFR, TAZ, DNMT3A, miR-1246/PEG3, MALAT1, JUNB, VASH1, ZBTB4/EZH2, ect | [30,39,40,41,42,43] |
| Gastric cancer | Upregulated | Oncogene | Proliferation, migration, invasion, metastasis | HDGF, ZMYM1, BATF2, CDCP1, ect | [46,47,49,50] |
| Pancreatic cancer | Upregulated | Oncogene | Proliferation, migration, invasion, chemoresistance | miR-25-3p/PHLPP2/ AKT-p70S6K, ect | [51,52] |
| Hepatocellular carcinoma and hepatoblastoma | Upregulated | Oncogene | Proliferation, colony formation, migration, tumorigenesis | SOCS2, HDGF, mTORC1, FOXO3, CTNNB1/Wnt/β-catenin, ect | [55,56,57,58,59] |
| Colorectal carcinoma | Upregulated | Oncogene | Self-renewal, proliferation, metastasis, tumorigenesis | SOX2, HK2, GLUT1, MYC, ect | [61,62,63] |
| Breast cancer | Upregulated | Oncogene | Proliferation, apoptosis, migration | Bcl2, HBXIP, AK4, ect | [66,68,70] |
| Ovarian cancer | Upregulated | Oncogene | Proliferation, migration, invasion, metastasis | AXL, PTEN, ect | [73,74] |
| Cervical cancer | Upregulated | Oncogene | Proliferation, aerobic glycolysis | HK2, RAB2B, ect | [76,77] |
| Glioblastoma | --- | Tumor suppressor | Self-renewal, stem cell differentiation | ADAM19, EPHA3, KLF4, ect | [80,81] |
| Upregulated | Oncogene | Proliferation | SOX2, ADAR1/CDK2, ect | [78,84] | |
| Renal cell carcinoma | Downregulated | Tumor suppressor | Proliferation, colony formation, migration, invasion | PI3K/Akt/mTOR, P2RX6, ect | [85,86,87] |
| Bladder cancer | Upregulated | Oncogene | Proliferation, colony formation, migration invasion | SETD7, KLF4, CDCP1, MYC, ITGA6, ect | [88,89,90,91] |
| Prostate cancer | Upregulated | Oncogene | Proliferation, migration, invasion, tumorigenesis | MYC, LEF1/Wnt, GLI1, LHPP, NKX3-1, ITGB1, ect | [93,94,95,96,97] |
| Melanoma | Upregulated | Oncogene | Colony formation, invasion | MMP2, N-cadherin, c-Met, ect | [98,99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, C.; Liu, J.; Li, Y.; Jiang, W.; Ji, D.; Liu, W.; Ma, T. Multifaceted Roles of the N6-Methyladenosine RNA Methyltransferase METTL3 in Cancer and Immune Microenvironment. Biomolecules 2022, 12, 1042. https://doi.org/10.3390/biom12081042
AMA Style
Hu C, Liu J, Li Y, Jiang W, Ji D, Liu W, Ma T. Multifaceted Roles of the N6-Methyladenosine RNA Methyltransferase METTL3 in Cancer and Immune Microenvironment. Biomolecules. 2022; 12(8):1042. https://doi.org/10.3390/biom12081042
Chicago/Turabian StyleHu, Chenxi, Jiacheng Liu, Yue Li, Wei Jiang, Ding Ji, Wei Liu, and Teng Ma. 2022. "Multifaceted Roles of the N6-Methyladenosine RNA Methyltransferase METTL3 in Cancer and Immune Microenvironment" Biomolecules 12, no. 8: 1042. https://doi.org/10.3390/biom12081042
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.